
Lower Zinc but Higher Calcium Content in Rodent Spinal Cord Compared to Brain

 , ,
, , 
Abstract

1. Introduction
2. Materials and Methods
2.1. Animals and Sample Preparation
2.2. Inductively Coupled Plasma Optic Emission Spectroscopy
2.3. Statistical Analysis of the ICP-OES Results
2.4. ZnT3-HA Imaging by Confocal Microscopy in Brain and Spinal Cord Sections
3. Results

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Organ | Ca | Mg | P | Zn | Fe | Cu |
|---|---|---|---|---|---|---|---|
| #1 male untreated (n = 11) | Brain | 0.4 ± 0.2 | 0.64 ± 0.08 | 9.4 ± 3.1 | 0.041 ± 0.015 | 0.047 ± 0.014 | 0.016 ± 0.005 |
| Spinal cord | 4.2 ± 2.3 | 0.68 ± 0.18 | 13.7 ± 3.2 | 0.030 ± 0.010 | 0.040 ± 0.013 | 0.008 ± 0.003 | |
| #2 male mock inj.* (n = 11) | Brain | 0.2 ± 0.1 | 0.67 ± 0.08 | 10.1 ± 2.9 | 0.040 ± 0.017 | 0.046 ± 0.010 | 0.013 ± 0.004 |
| Spinal cord | 2.4 ± 1.4 | 0.48 ± 0.14 | 11.5 ± 3.9 | 0.020 ± 0.007 | 0.033 ± 0.011 | 0.010 ± 0.005 | |
| #3 female untreated (n = 8) | Brain | 0.4 ± 0.3 | 0.57 ± 0.03 | 7.0 ± 0.3 | 0.047 ± 0.005 | 0.039 ± 0.023 | 0.015 ± 0.001 |
| Spinal cord | 1.8 ± 1.2 | 0.44 ± 0.04 | 7.7 ± 0.7 | 0.025 ± 0.003 | 0.024 ± 0.005 | 0.007 ± 0.001 | |
| Mean (n = 3) | Brain | 0.3 ± 0.1 | 0.63 ± 0.05 | 8.8 ± 1.7 | 0.043 ± 0.004 | 0.044 ± 0.004 | 0.015 ± 0.001 |
| Spinal cord | 2.8 ± 1.1 | 0.53 ± 0.13 | 11.0 ± 3.1 | 0.025 ± 0.005 | 0.032 ± 0.008 | 0.009 ± 0.001 | |
| Fold difference | 7.1 | −0.2 | 0.2 | −0.4 | −0.3 | −0.4 | |
| T-test (p-value) | 0.03 | 0.30 | 0.35 | 0.01 | 0.09 | 0.004 |
| Experiment | Organ | Ca | Mg | P | Zn | Fe | Cu |
|---|---|---|---|---|---|---|---|
| #1 male ND (n = 11) | Brain | 0.3 ± 0.2 | 0.75 ± 0.11 | 14.7 ± 3.4 | 0.063 ± 0.015 | 0.086 ± 0.017 | 0.011 ± 0.003 |
| Spinal cord | 4.1 ± 2.0 | 0.73 ± 0.08 | 20.2 ± 4.1 | 0.035 ± 0.011 | 0.073 ± 0.037 | 0.009 ± 0.008 | |
| #2 male RD (n = 11) | Brain | 0.3 ± 0.2 | 0.80 ± 0.07 | 16.2 ± 1.7 | 0.067 ± 0.015 | 0.100 ± 0.046 | 0.011 ± 0.002 |
| Spinal cord | 2.5 ± 2.2 | 0.63 ± 0.08 | 17.8 ± 4.1 | 0.033 ± 0.005 | 0.057 ± 0.022 | 0.006 ± 0.003 | |
| #3 female ND (n = 10) | Brain | 0.6 ± 0.6 | 0.73 ± 0.08 | 15.0 ± 1.8 | 0.065 ± 0.010 | 0.078 ± 0.014 | 0.011 ± 0.002 |
| Spinal cord | 4.3 ± 3.9 | 0.58 ± 0.08 | 16.9 ± 3.0 | 0.028 ± 0.004 | 0.049 ± 0.014 | 0.004 ± 0.002 | |
| #4 female RD (n = 9) | Brain | 0.4 ± 0.2 | 0.79 ± 0.06 | 15.6 ± 0.9 | 0.071 ± 0.006 | 0.077 ± 0.012 | 0.012 ± 0.002 |
| Spinal cord | 2.8 ± 4.3 | 0.57 ± 0.07 | 15.7 ± 2.9 | 0.035 ± 0.016 | 0.051 ± 0.016 | 0.004 ± 0.002 | |
| Mean (n = 4) | Brain | 0.4 ± 0.1 | 0.77 ± 0.04 | 15.4 ± 0.6 | 0.066 ± 0.003 | 0.085 ± 0.011 | 0.012 ± 0.001 |
| Spinal cord | 3.4 ± 0.9 | 0.63 ± 0.08 | 17.7 ± 1.9 | 0.033 ± 0.000 | 0.057 ± 0.011 | 0.006 ± 0.002 | |
| Fold difference | 7.8 | −0.2 | 0.2 | −0.5 | −0.3 | −0.5 | |
| T-test (p-value) | 0.0006 | 0.02 | 0.06 | 0.000006 | 0.01 | 0.003 |
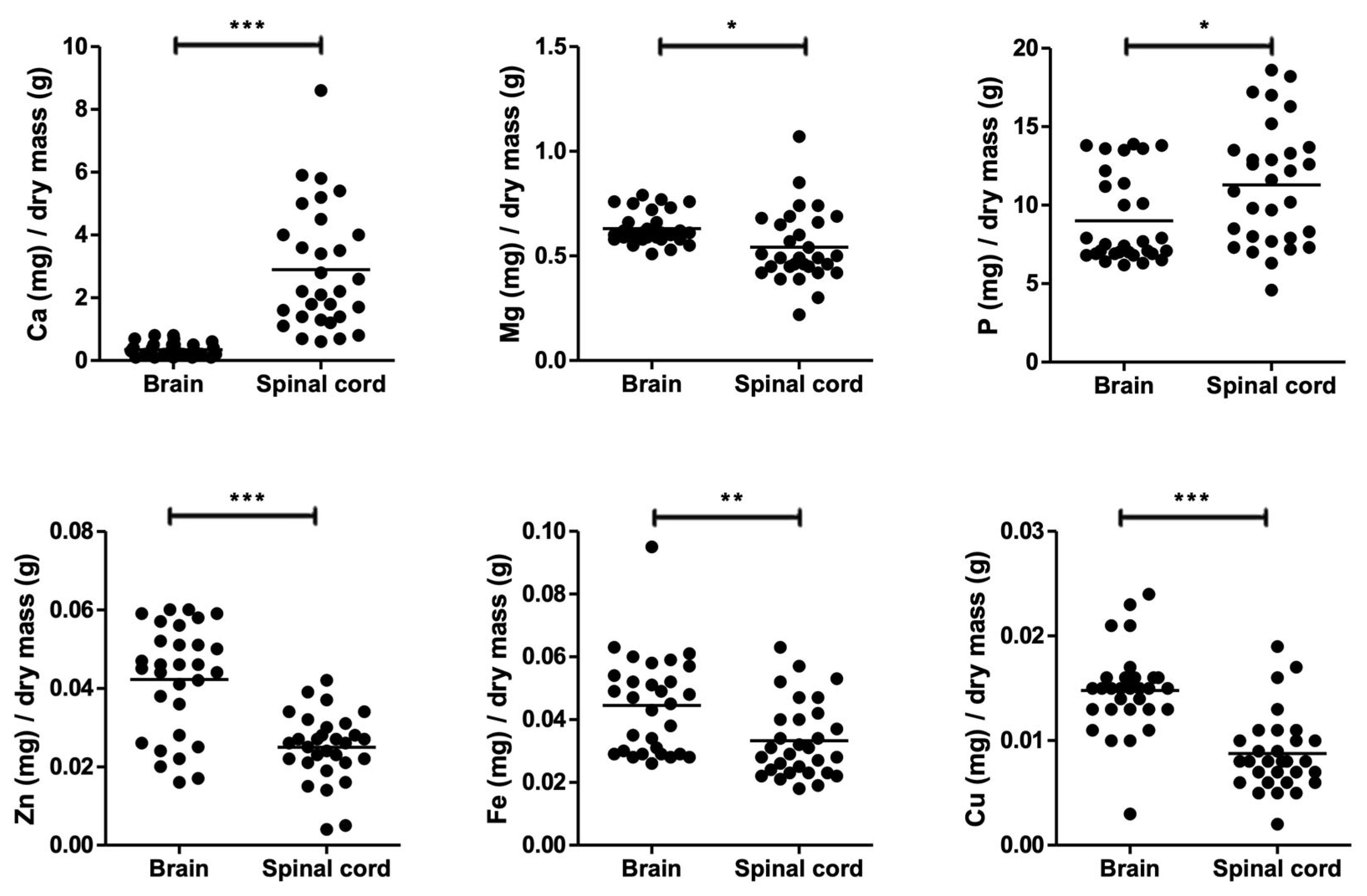
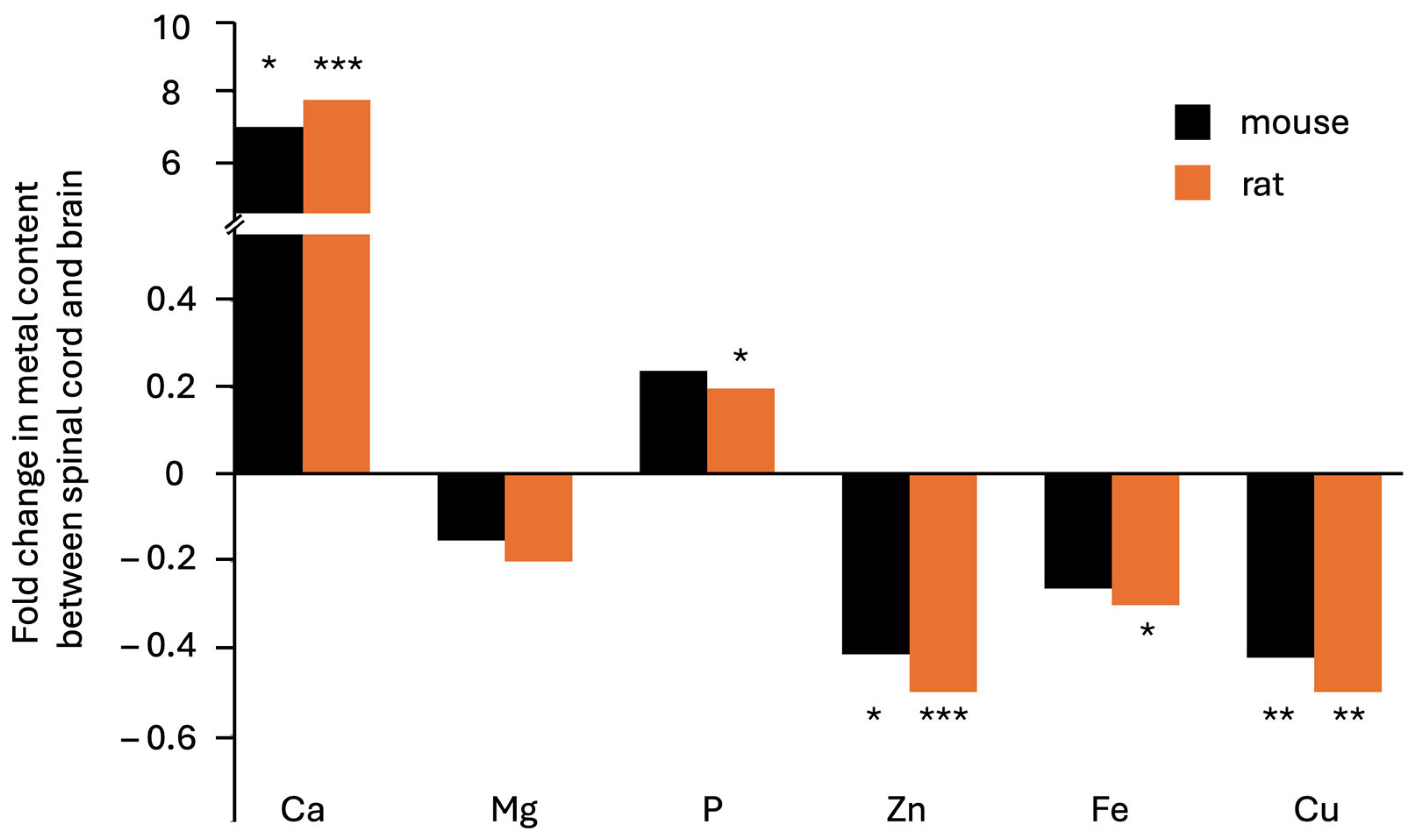
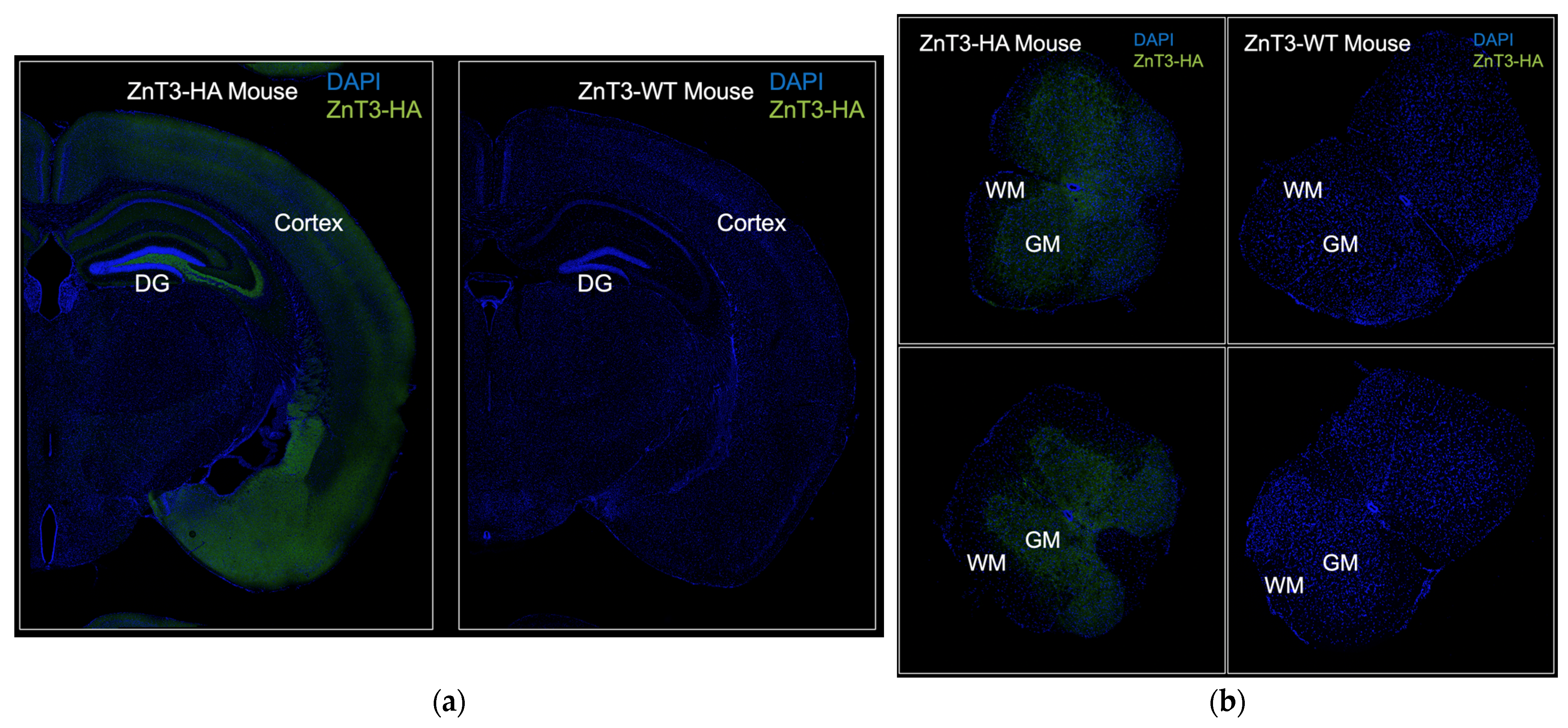
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Krall, R.F.; Tzounopoulos, T.; Aizenman, E. The Function and Regulation of Zinc in the Brain. Neuroscience 2021, 457, 235–258. [Google Scholar] [CrossRef] [PubMed]
- Hollings, A.L.; Ellison, G.C.; Willans, M.; Lam, V.; Munyard, T.; Remy, A.R.; Takechi, R.; Mamo, J.C.L.; Webb, S.; New, E.J.; et al. Subventricular Accumulation of Cu in the Aging Mouse Brain Does Not Associate with Anticipated Increases in Markers of Oxidative Stress. ACS Chem. Neurosci. 2025, 16, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Möller, H.E.; Bossoni, L.; Connor, J.R.; Crichton, R.R.; Does, M.D.; Ward, R.J.; Zecca, L.; Zucca, F.A.; Ronen, I. Iron, Myelin, and the Brain: Neuroimaging Meets Neurobiology. Trends Neurosci. 2019, 42, 384–401. [Google Scholar] [CrossRef] [PubMed]
- Posadas, Y.; Lopez-Guerrero, V.E.; Arcos-Lopez, T.; Sayler, R.I.; Sanchez-Lopez, C.; Perez-Cruz, C.; Quintanar, L. The role of d-block metal ions in neurodegenerative diseases. In Comprehensive Inorganic Chemistry III, 3rd ed.; Reedijk, J., Poeppelmeier, K.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2023; pp. 575–628. [Google Scholar]
- Yoo, J.; Han, J.; Lim, M.H. Transition metal ions and neurotransmitters: Coordination chemistry and implications for neurodegeneration. RSC Chem. Biol. 2023, 4, 548–563. [Google Scholar] [CrossRef]
- Kenkhuis, B.; Bush, A.I.; Ayton, S. How iron can drive neurodegeneration. Trends Neurosci. 2023, 46, 333–335. [Google Scholar] [CrossRef]
- Popescu, B.F.; Robinson, C.A.; Rajput, A.; Rajput, A.H.; Harder, S.L.; Nichol, H. Iron, copper, and zinc distribution of the cerebellum. Cerebellum 2009, 8, 74–79. [Google Scholar] [CrossRef]
- Ellison, G.; Hollings, A.L.; Hackett, M.J. A review of the “metallome” within neurons and glia, as revealed by elemental mapping of brain tissue. BBA Adv. 2021, 2, 100038. [Google Scholar] [CrossRef]
- Pushie, M.J.; Sylvain, N.J.; Hou, H.; Pendleton, N.; Wang, R.; Zimmermann, L.; Pally, M.; Cayabyab, F.S.; Peeling, L.; Kelly, M.E. X-Ray fluorescence mapping of brain tissue reveals the profound extent of trace element dysregulation in stroke pathophysiology. Metallomics 2024, 16, mfae054. [Google Scholar] [CrossRef]
- Goutman, S.A.; Hardiman, O.; Al-Chalabi, A.; Chió, A.; Savelieff, M.G.; Kiernan, M.C.; Feldman, E.L. Emerging insights into the complex genetics and pathophysiology of amyotrophic lateral sclerosis. Lancet Neurol. 2022, 21, 465–479. [Google Scholar] [CrossRef]
- Muccilli, A.; Seyman, E.; Oh, J. Spinal Cord MRI in Multiple Sclerosis. Neurol. Clin. 2018, 36, 35–57. [Google Scholar] [CrossRef]
- Calabro, F.J.; Perez, M.A. Bilateral reach-to-grasp movement asymmetries after human spinal cord injury. J. Neurophysiol. 2016, 115, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Sirabella, R.; Valsecchi, V.; Anzilotti, S.; Cuomo, O.; Vinciguerra, A.; Cepparulo, P.; Brancaccio, P.; Guida, N.; Blondeau, N.; Canzoniero, L.M.T.; et al. Ionic Homeostasis Maintenance in ALS: Focus on New Therapeutic Targets. Front. Neurosci. 2018, 12, 510. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.H.; Elliot, J.L. Metallothionein expression is altered in a transgenic murine model of familial amyotrophic lateral sclerosis. Exp. Neurol. 2000, 162, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.J.; Zhou, Q.S.; Huang, H.B.; Wan, Y.L.; Tian, S.F.; Duan, D.M. Effects of ketamine on the balance of ions Ca2+, Mg2+, Cu2+ and Zn2+ in the ischemia-reperfusion affected spinal cord tissues in rabbits. Neurochem. Res. 2009, 34, 2192–2196. [Google Scholar] [CrossRef]
- Tomik, B.; Chwiej, J.; Szczerbowska-Boruchowska, M.; Lankosz, M.; Wójcik, S.; Adamek, D.; Falkenberg, G.; Bohic, S.; Simionovici, A.; Stegowski, Z.; et al. Implementation of X-Ray fluorescence microscopy for investigation of elemental abnormalities in amyotrophic lateral sclerosis. Neurochem. Res. 2006, 31, 321–331. [Google Scholar] [CrossRef]
- Kinebuchi, M.; Matsuura, A.; Kiyono, T.; Nomura, Y.; Kimura, S. Diagnostic copper imaging of Menkes disease by synchrotron radiation-generated X-Ray fluorescence analysis. Sci. Rep. 2016, 6, 33247. [Google Scholar] [CrossRef]
- Santos-Díaz, A.I.; Solís-López, J.; Díaz-Torres, E.; Guadarrama-Olmos, J.C.; Osorio, B.; Kroll, T.; Webb, S.M.; Hiriart, M.; Jiménez-Estrada, I.; Missirlis, F. Metal ion content of internal organs in the calorically restricted Wistar rat. J. Trace Elem. Med. Biol. 2023, 78, 127182. [Google Scholar] [CrossRef]
- Weed, L.H. The Cells of the Arachnoid. Bull. Johns Hopkins Hosp. 1920, 31, 343. [Google Scholar]
- Herren, Y.R. Occurrence and distribution of calcified plaques in the spinal arachnoid in man. Arch. Neurol. Psych. 1939, 41, 1180–1186. [Google Scholar] [CrossRef]
- Weed, L.H. Studies on Cerebro-Spinal Fluid. No. III: The pathways of escape from the Subarachnoid Spaces with particular reference to the Arachnoid Villi. J. Med. Res. 1914, 31, 51–91. [Google Scholar]
- Tanaka, K.; Nishiura, I.; Koyama, T. Arachnoiditis ossificans after repeated myelographies and spinal operations–A case report and review of the literature. Neurol. Surg. 1987, 15, 89–93. [Google Scholar]
- Young, W. Role of calcium in central nervous system injuries. J. Neurotrauma 1992, 9, S9–S25. [Google Scholar]
- Frizzell, B.; Kaplan, P.; Dussault, R.; Sevick, R. Arachnoiditis ossificans: MR imaging features in five patients. AJR Am. J. Roentgenol. 2001, 177, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Faure, A.; Khalfallah, M.; Perrouin-Verbe, B.; Caillon, F.; Deschamps, C.; Bord, E.; Mathe, J.F.; Robert, R. Arachnoiditis ossificans of the cauda equina. Case report and review of the literature. J. Neurosurg. 2002, 97, 239–243. [Google Scholar] [PubMed]
- Wright, M.H.; Denney, L.C. A comprehensive review of spinal arachnoiditis. Orthop. Nurs. 2003, 22, 215–219. [Google Scholar] [CrossRef]
- Slavin, K.V.; Nixon, R.R.; Nesbit, G.M.; Burchiel, K.J. Extensive arachnoid ossification with associated syringomyelia presenting as thoracic myelopathy. Case report and review of the literature. J. Neurosurg. 1999, 91, 223–229. [Google Scholar]
- Haug, F.M. Electron microscopical localization of the zinc in hippocampal mossy fibre synapses by a modified sulfide silver procedure. Histochemie 1967, 8, 355–368. [Google Scholar] [CrossRef]
- Crawford, J.D.; Connor, J.D. Zinc in maturing rat brain: Hippocampal concentration and localization. J. Neurochem. 1972, 19, 1451–1458. [Google Scholar] [CrossRef]
- Fjerdingstad, E.; Danscher, G.; Fjerdingstad, E.J. Zinc content in hippocampus and whole brain of normal rats. Brain Res. 1974, 79, 338–342. [Google Scholar] [CrossRef]
- Wenzel, H.J.; Cole, T.B.; Born, D.E.; Schwartzkroin, P.A.; Palmiter, R.D. Ultrastructural localization of zinc transporter-3 (ZnT-3) to synaptic vesicle membranes within mossy fiber boutons in the hippocampus of mouse and monkey. Proc. Natl. Acad. Sci. USA 1997, 94, 12676–12681. [Google Scholar] [CrossRef]
- Cole, T.B.; Wenzel, H.J.; Kafer, K.E.; Schwartzkroin, P.A.; Palmiter, R.D. Elimination of zinc from synaptic vesicles in the intact mouse brain by disruption of the ZnT3 gene. Proc. Natl. Acad. Sci. USA 1999, 96, 1716–1721. [Google Scholar] [CrossRef] [PubMed]
- Kantheti, P.; Qiao, X.; Diaz, M.E.; Peden, A.A.; Meyer, G.E.; Carskadon, S.L.; Kapfhamer, D.; Sufalko, D.; Robinson, M.S.; Noebels, J.L.; et al. Mutation in AP-3 delta in the mocha mouse links endosomal transport to storage deficiency in platelets, melanosomes, and synaptic vesicles. Neuron 1998, 21, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Tejeda-Guzmán, C.; Rosas-Arellano, A.; Kroll, T.; Webb, S.M.; Barajas-Aceves, M.; Osorio, B.; Missirlis, F. Biogenesis of zinc storage granules in Drosophila melanogaster. J. Exp. Biol. 2018, 221, jeb168419. [Google Scholar]
- Howell, G.A.; Welch, M.G.; Frederickson, C.J. Stimulation-induced uptake and release of zinc in hippocampal slices. Nature 1984, 308, 736–738. [Google Scholar] [CrossRef]
- Wang, Z.; Li, J.Y.; Dahlström, A.; Danscher, G. Zinc-enriched GABAergic terminals in mouse spinal cord. Brain Res. 2001, 921, 165–172. [Google Scholar] [CrossRef]
- Qian, J.; Noebels, J.L. Visualization of transmitter release with zinc fluorescence detection at the mouse hippocampal mossy fibre synapse. J. Physiol. 2005, 566, 747–758. [Google Scholar] [CrossRef]
- Brown, C.E.; Dyck, R.H. Modulation of synaptic zinc in barrel cortex by whisker stimulation. Neuroscience 2005, 134, 355–359. [Google Scholar] [CrossRef]
- Nakashima, A.S.; Dyck, R.H. Enhanced plasticity in zincergic, cortical circuits after exposure to enriched environments. J. Neurosci. 2008, 28, 13995–13999. [Google Scholar] [CrossRef]
- Nakashima, A.S.; Dyck, R.H. Dynamic, experience-dependent modulation of synaptic zinc within the excitatory synapses of the mouse barrel cortex. Neuroscience 2010, 170, 1015–1019. [Google Scholar] [CrossRef]
- Sindreu, C.; Palmiter, R.D.; Storm, D.R. Zinc transporter ZnT-3 regulates presynaptic Erk1/2 signaling and hippocampus-dependent memory. Proc. Natl. Acad. Sci. USA 2011, 108, 3366–3370. [Google Scholar] [CrossRef]
- Amico-Ruvio, S.A.; Murthy, S.E.; Smith, T.P.; Popescu, G.K. Zinc effects on NMDA receptor gating kinetics. Biophys. J. 2011, 100, 1910–1918. [Google Scholar] [CrossRef] [PubMed]
- Perez-Rosello, T.; Anderson, C.T.; Schopfer, F.J.; Zhao, Y.; Gilad, D.; Salvatore, S.R.; Freeman, B.A.; Hershfinkel, M.; Aizenman, E.; Tzounopoulos, T. Synaptic Zn2+ inhibits neurotransmitter release by promoting endocannabinoid synthesis. J. Neurosci. 2013, 33, 9259–9272. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.T.; Radford, R.J.; Zastrow, M.L.; Zhang, D.Y.; Apfel, U.P.; Lippard, S.J.; Tzounopoulos, T. Modulation of extrasynaptic NMDA receptors by synaptic and tonic zinc. Proc. Natl. Acad. Sci. USA 2015, 112, 2705–2714. [Google Scholar] [CrossRef] [PubMed]
- Kalappa, B.I.; Anderson, C.T.; Goldberg, J.M.; Lippard, S.J.; Tzounopoulos, T. AMPA receptor inhibition by synaptically released zinc. Proc. Natl. Acad. Sci. USA 2015, 112, 15749–15754. [Google Scholar] [CrossRef]
- Perez-Rosello, T.; Anderson, C.T.; Ling, C.; Lippard, S.J.; Tzounopoulos, T. Tonic zinc inhibits spontaneous firing in dorsal cochlear nucleus principal neurons by enhancing glycinergic neurotransmission. Neurobiol. Dis. 2015, 81, 14–19. [Google Scholar] [CrossRef]
- Patrick-Wu, H.P.; Dyck, R.H. Signaling by Synaptic Zinc is Required for Whisker-Mediated, Fine Texture Discrimination. Neuroscience 2018, 369, 242–247. [Google Scholar] [CrossRef]
- McAllister, B.B.; Wright, D.K.; Wortman, R.C.; Shultz, S.R.; Dyck, R.H. Elimination of vesicular zinc alters the behavioural and neuroanatomical effects of social defeat stress in mice. Neurobiol. Stress 2018, 9, 199–213. [Google Scholar] [CrossRef]
- Kouvaros, S.; Kumar, M.; Tzounopoulos, T. Synaptic Zinc Enhances Inhibition Mediated by Somatostatin, but not Parvalbumin, Cells in Mouse Auditory Cortex. Cereb. Cortex 2020, 30, 3895–3909. [Google Scholar] [CrossRef]
- Kouvaros, S.; Bizup, B.; Solis, O.; Kumar, M.; Ventriglia, E.; Curry, F.P.; Michaelides, M.; Tzounopoulos, T. A CRE/DRE dual recombinase transgenic mouse reveals synaptic zinc-mediated thalamocortical neuromodulation. Sci. Adv. 2023, 9, eadf3525. [Google Scholar] [CrossRef]
- Bender, P.T.R.; McCollum, M.; Boyd-Pratt, H.; Mendelson, B.Z.; Anderson, C.T. Synaptic zinc potentiates AMPA receptor function in mouse auditory cortex. Cell Rep. 2023, 42, 112932. [Google Scholar] [CrossRef]
- Chrusch, M.J.; Fu, S.; Spanswick, S.C.; Vecchiarelli, H.A.; Patel, P.P.; Hill, M.N.; Dyck, R.H. Environmental enrichment engages vesicular zinc signaling to enhance hippocampal neurogenesis. Cells 2023, 12, 883. [Google Scholar] [CrossRef] [PubMed]
- Bizup, B.; Tzounopoulos, T. On the genesis and unique functions of zinc neuromodulation. J. Neurophysiol. 2024, 132, 1241–1254. [Google Scholar] [CrossRef] [PubMed]
- Linkous, D.H.; Flinn, J.M.; Koh, J.Y.; Lanzirotti, A.; Bertsch, P.M.; Jones, B.F.; Giblin, L.J.; Frederickson, C.J. Evidence that the ZNT3 protein controls the total amount of elemental zinc in synaptic vesicles. J. Histochem. Cytochem. 2008, 56, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Upmanyu, N.; Jin, J.; Emde, H.V.; Ganzella, M.; Bösche, L.; Malviya, V.N.; Zhuleku, E.; Politi, A.Z.; Ninov, M.; Silbern, I.; et al. Colocalization of different neurotransmitter transporters on synaptic vesicles is sparse except for VGLUT1 and ZnT3. Neuron 2022, 110, 1483–1497. [Google Scholar] [CrossRef]
- Bizup, B.; Brutsaert, S.; Cunningham, C.L.; Thathiah, A.; Tzounopoulos, T. Cochlear zinc signaling dysregulation is associated with noise-induced hearing loss, and zinc chelation enhances cochlear recovery. Proc. Natl. Acad. Sci. USA 2024, 121, e2310561121. [Google Scholar] [CrossRef]
- Palmiter, R.D.; Cole, T.B.; Quaife, C.J.; Findley, S.D. ZnT-3, a putative transporter of zinc into synaptic vesicles. Proc. Natl. Acad. Sci. USA 1996, 93, 14934–14939. [Google Scholar] [CrossRef]
- Brown, C.E.; Dyck, R.H. Distribution of zincergic neurons in the mouse forebrain. J. Comp. Neurol. 2004, 479, 156–167. [Google Scholar] [CrossRef]
- Chi, T.; Kim, M.S.; Lang, S.; Bose, N.; Kahn, A.; Flechner, L.; Blaschko, S.D.; Zee, T.; Muteliefu, G.; Bond, N.; et al. A Drosophila model identifies a critical role for zinc in mineralization for kidney stone disease. PLoS ONE 2015, 10, e0124150. [Google Scholar] [CrossRef]
- Ustriyana, P.; Hennefarth, M.R.; Srirangapatanam, S.; Jung, H.; Wang, Y.; Chen, L.; Lue, T.F.; Lin, G.; Kang, M.; Stoller, M.L.; et al. Mineralized Peyronie’s plaque has a phenotypic resemblance to bone. Acta Biomater. 2022, 140, 457–466. [Google Scholar] [CrossRef]
- Kwon, J.H.; Kim, D.K.; Cho, Y.E.; Kwun, I.S. Zinc Action in Vascular Calcification. Prev. Nutr. Food Sci. 2024, 29, 118–124. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos-Díaz, A.I.; Bizup, B.; Pantaleón-Gómez, A.K.; Osorio, B.; Barbier, O.C.; Tzounopoulos, T.; Missirlis, F. Lower Zinc but Higher Calcium Content in Rodent Spinal Cord Compared to Brain. Cells 2025, 14, 922. https://doi.org/10.3390/cells14120922
Santos-Díaz AI, Bizup B, Pantaleón-Gómez AK, Osorio B, Barbier OC, Tzounopoulos T, Missirlis F. Lower Zinc but Higher Calcium Content in Rodent Spinal Cord Compared to Brain. Cells. 2025; 14(12):922. https://doi.org/10.3390/cells14120922
Chicago/Turabian StyleSantos-Díaz, Alma I., Brandon Bizup, Ana Karen Pantaleón-Gómez, Beatriz Osorio, Olivier Christophe Barbier, Thanos Tzounopoulos, and Fanis Missirlis. 2025. "Lower Zinc but Higher Calcium Content in Rodent Spinal Cord Compared to Brain" Cells 14, no. 12: 922. https://doi.org/10.3390/cells14120922
APA StyleSantos-Díaz, A. I., Bizup, B., Pantaleón-Gómez, A. K., Osorio, B., Barbier, O. C., Tzounopoulos, T., & Missirlis, F. (2025). Lower Zinc but Higher Calcium Content in Rodent Spinal Cord Compared to Brain. Cells, 14(12), 922. https://doi.org/10.3390/cells14120922