

Advances in Food Allergy Immunotherapy: Current Strategies and Role of Antibodies Isotypes


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Humoral Response
2.1. Immunoglobulin E (IgE) in Immunity: Biological Humoral Response
2.2. Allergen-Specific IgE Antibodies
2.3. IgG and IgA Biological Humoral Response
3. Cellular Response
3.1. Isotype Class Switching, Affinity Maturation, and IgE Production
3.2. IgE Plasma Cells and Memory B Cells in Food Allergies
4. Food Allergy Treatments
4.1. Anti-IgE Targeted Immunotherapy
4.2. Allergen-Specific Immunotherapy (AIT)
5. Changes in Allergen-Specific Immunity After AIT
5.1. Immunophenotypic Changes in Circulating Allergen-Specific Memory B Cells
5.2. Serum Allergen-Specific Immunoglobulin Changes
5.3. Allergen-Specific Isotypes Shifting After AIT: Do They Possess Neutralizing Capabilities?
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Loh, W.; Tang, M.L.K. The Epidemiology of Food Allergy in the Global Context. Int. J. Environ. Res. Public Health 2018, 15, 2043. [Google Scholar] [CrossRef] [PubMed]
- Sicherer, S.H.; Sampson, H.A. Food allergy: A review and update on epidemiology, pathogenesis, diagnosis, prevention, and management. J. Allergy Clin. Immunol. 2018, 141, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Knol, E.F.; van Neerven, R.J.J. IgE versus IgG and IgA: Differential roles of allergen-specific antibodies in sensitization, tolerization, and treatment of allergies. Immunol. Rev. 2024, 328, 314–333. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C.M.; Yarrarapu, S.N.S.; Mendez, M.D. Food Allergies. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]
- Neuman-Sunshine, D.L.; Eckman, J.A.; Keet, C.A.; Matsui, E.C.; Peng, R.D.; Lenehan, P.J.; Wood, R.A. The natural history of persistent peanut allergy. Ann. Allergy Asthma Immunol. 2012, 108, 326–331.e3. [Google Scholar] [CrossRef]
- Wood, R.A.; Sicherer, S.H.; Vickery, B.P.; Jones, S.M.; Liu, A.H.; Fleischer, D.M.; Henning, A.K.; Mayer, L.; Burks, A.W.; Grishin, A.; et al. The natural history of milk allergy in an observational cohort. J. Allergy Clin. Immunol. 2013, 131, 805–812. [Google Scholar] [CrossRef]
- Savage, J.; Sicherer, S.; Wood, R. The Natural History of Food Allergy. J. Allergy Clin. Immunol. practice 2016, 4, 196–203. quiz 204. [Google Scholar] [CrossRef]
- Burks, A.W.; Sampson, H.A.; Plaut, M.; Lack, G.; Akdis, C.A. Treatment for food allergy. J. Allergy Clin. Immunol. 2018, 141, 1–9. [Google Scholar] [CrossRef]
- Berin, M.C. Targeting type 2 immunity and the future of food allergy treatment. J. Exp. Med. 2023, 220, e20221104. [Google Scholar] [CrossRef]
- Furiness, K.N.; El Ansari, Y.S.; Oettgen, H.C.; Kanagaratham, C. Allergen-specific IgA and IgG antibodies as inhibitors of mast cell function in food allergy. Front. Allergy 2024, 5, 1389669. [Google Scholar] [CrossRef]
- Ando, T.; Kitaura, J. Tuning IgE: IgE-Associating Molecules and Their Effects on IgE-Dependent Mast Cell Reactions. Cells 2021, 10, 1697. [Google Scholar] [CrossRef]
- Hellman, L.T.; Akula, S.; Thorpe, M.; Fu, Z. Tracing the Origins of IgE, Mast Cells, and Allergies by Studies of Wild Animals. Front. Immunol. 2017, 8, 1749. [Google Scholar] [CrossRef] [PubMed]
- Dullaers, M.; De Bruyne, R.; Ramadani, F.; Gould, H.J.; Gevaert, P.; Lambrecht, B.N. The who, where, and when of IgE in allergic airway disease. J. Allergy Clin. Immunol. 2012, 129, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.S.; Greer, A.M. The role of FcεRI expressed in dendritic cells and monocytes. Cell. Mol. Life Sci. 2015, 72, 2349–2360. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Grobe, W.; Walgenbach-Brünagel, G.; Flicker, S.; Yu, C.; Sylvester, M.; Allam, J.P.; Oldenburg, J.; Garbi, N.; Valenta, R.; et al. Distinct Expression and Function of FcεRII in Human B Cells and Monocytes. J. Immunol. 2017, 198, 3033–3044. [Google Scholar] [CrossRef]
- Poole, J.A.; Rosenwasser, L.J. The role of immunoglobulin E and immune inflammation: Implications in allergic rhinitis. Curr. Allergy Asthma Rep. 2005, 5, 252–258. [Google Scholar] [CrossRef]
- Wu, L.C.; Zarrin, A.A. The production and regulation of IgE by the immune system. Nat. Rev. Immunol. 2014, 14, 247–259. [Google Scholar] [CrossRef]
- Geha, R.S.; Jabara, H.H.; Brodeur, S.R. The regulation of immunoglobulin E class-switch recombination. Nat. Rev. Immunol. 2003, 3, 721–732. [Google Scholar] [CrossRef]
- Lawrence, M.G.; Woodfolk, J.A.; Schuyler, A.J.; Stillman, L.C.; Chapman, M.D.; Platts-Mills, T.A. Half-life of IgE in serum and skin: Consequences for anti-IgE therapy in patients with allergic disease. J. Allergy Clin. Immunol. 2017, 139, 422–428.e4. [Google Scholar] [CrossRef]
- Gowthaman, U.; Chen, J.S.; Eisenbarth, S.C. Regulation of IgE by T follicular helper cells. J. Leukoc. Biol. 2020, 107, 409–418. [Google Scholar] [CrossRef]
- Lama, J.K.; Kita, H.T. TFH cells regulate antibody affinity and determine the outcomes of anaphylaxis. J. Allergy Clin. Immunol. 2022, 150, 1042–1044. [Google Scholar] [CrossRef]
- Bax, H.J.; Keeble, A.H.; Gould, H.J. Cytokinergic IgE Action in Mast Cell Activation. Front. Immunol. 2012, 3, 229. [Google Scholar] [CrossRef] [PubMed]
- Vidarsson, G.; Dekkers, G.; Rispens, T. IgG subclasses and allotypes: From structure to effector functions. Front. Immunol. 2014, 5, 520. [Google Scholar] [CrossRef] [PubMed]
- Rispens, T.; Huijbers, M.G. The unique properties of IgG4 and its roles in health and disease. Nat. Rev. Immunol. 2023, 23, 763–778. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Ojalvo, D.; Chen, X.; Kazmi, W.; Menchén-Martínez, D.; Pérez-Rodríguez, L.; Fernandes-Braga, W.; Tyler, S.; Benkov, K.; Pittman, N.; Lai, J.; et al. Differential T follicular helper cell phenotypes distinguish IgE-mediated milk allergy from eosinophilic esophagitis in children. J. Allergy Clin. Immunol. 2025, 155, 909–922. [Google Scholar] [CrossRef]
- Leser, T.D.; Molbak, L. Better living through microbial action: The benefits of the mammalian gastrointestinal microbiota on the host. Environ. Microbiol. 2009, 11, 2194–2206. [Google Scholar] [CrossRef]
- Woof, J.M.; Kerr, M.A. The function of immunoglobulin A in immunity. J. Pathol. 2006, 208, 270–282. [Google Scholar] [CrossRef]
- Mkaddem, S.B.; Christou, I.; Rossato, E.; Berthelot, L.; Lehuen, A.; Monteiro, R.C. IgA, IgA receptors, and their anti-inflammatory properties. Curr. Top. Microbiol. Immunol. 2014, 382, 221–235. [Google Scholar] [CrossRef]
- Macpherson, A.J.; McCoy, K.D.; Johansen, F.E.; Brandtzaeg, P. The immune geography of IgA induction and function. Mucosal Immunol. 2008, 1, 11–22. [Google Scholar] [CrossRef]
- Bottcher, M.F.; Haggstrom, P.; Bjorksten, B.; Jenmalm, M.C. Total and allergen-specific immunoglobulin A levels in saliva in relation to the development of allergy in infants up to 2 years of age. Clin. Exp. Allergy 2002, 32, 1293–1298. [Google Scholar] [CrossRef]
- Sandin, A.; Bjorksten, B.; Bottcher, M.F.; Englund, E.; Jenmalm, M.C.; Braback, L. High salivary secretory IgA antibody levels are associated with less late-onset wheezing in IgE-sensitized infants. Pediatr. Allergy Immunol. 2011, 22, 477–481. [Google Scholar] [CrossRef]
- Ludviksson, B.R.; Arason, G.J.; Thorarensen, O.; Ardal, B.; Valdimarsson, H. Allergic diseases and asthma in relation to serum immunoglobulins and salivary immunoglobulin A in pre-school children: A follow-up community-based study. Clin. Exp. Allergy 2005, 35, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Fageras, M.; Tomicic, S.; Voor, T.; Bjorksten, B.; Jenmalm, M.C. Slow salivary secretory IgA maturation may relate to low microbial pressure and allergic symptoms in sensitized children. Pediatr. Res. 2011, 70, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Kukkonen, K.; Kuitunen, M.; Haahtela, T.; Korpela, R.; Poussa, T.; Savilahti, E. High intestinal IgA associates with reduced risk of IgE-associated allergic diseases. Pediatr. Allergy Immunol. 2010, 21, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, M.; Kinoshita, K.; Fagarasan, S.; Yamada, S.; Shinkai, Y.; Honjo, T. Class switch recombination and hypermutation require activation-induced cytidine deaminase (AID), a potential RNA editing enzyme. Cell 2000, 102, 553–563. [Google Scholar] [CrossRef]
- Finkelman, F.D.; Holmes, J.; Katona, I.M.; Urban, J.F.; Beckmann, M.P.; Park, L.S.; Schooley, K.A.; Coffman, R.L.; Mosmann, T.R.; Paul, W.E. Lymphokine control of in vivo immunoglobulin isotype selection. Annu. Rev. Immunol. 1990, 8, 303–333. [Google Scholar] [CrossRef]
- Miranda-Waldetario, M.C.G.; Redes, J.; Fernandes-Braga, W.; Curotto de Lafaille, M.A. B cell memory of IgE responses in food allergy. In Encyclopedia of Food Allergy; Reference Module in Food Allergy; Elsevier: New York, NY, USA, 2023. [Google Scholar] [CrossRef]
- Xiong, H.; Dolpady, J.; Wabl, M.; Curotto de Lafaille, M.A.; Lafaille, J.J. Sequential class switching is required for the generation of high affinity IgE antibodies. J. Exp. Med. 2012, 209, 353–364. [Google Scholar] [CrossRef]
- Erazo, A.; Kutchukhidze, N.; Leung, M.; Christ, A.P.; Urban, J.F., Jr.; Curotto de Lafaille, M.A.; Lafaille, J.J. Unique maturation program of the IgE response in vivo. Immunity 2007, 26, 191–203. [Google Scholar] [CrossRef]
- Chen, Q.; Liu, H.; Luling, N.; Reinke, J.; Dent, A.L. Evidence that High-Affinity IgE Can Develop in the Germinal Center in the Absence of an IgG1-Switched Intermediate. J. Immunol. 2023, 210, 905–915. [Google Scholar] [CrossRef]
- Koenig, J.F.E.; Wade-Vallance, A.K.; Jiménez-Saiz, R.; Bruton, K.; Gadkar, S.; Grydziuszko, E.; Walker, T.D.; Gordon, M.E.; Gillgrass, A.E.; Taylor, J.J.; et al. Allergic Reactivity and Memory Occur Independently of Sequential Switching Through IgG1. Allergy 2025, Epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- He, J.S.; Meyer-Hermann, M.; Xiangying, D.; Zuan, L.Y.; Jones, L.A.; Ramakrishna, L.; de Vries, V.C.; Dolpady, J.; Aina, H.; Joseph, S.; et al. The distinctive germinal center phase of IgE+ B lymphocytes limits their contribution to the classical memory response. J. Exp. Med. 2013, 210, 2755–2771. [Google Scholar] [CrossRef]
- Asrat, S.; Kaur, N.; Liu, X.; Ben, L.H.; Kajimura, D.; Murphy, A.J.; Sleeman, M.A.; Limnander, A.; Orengo, J.M. Chronic allergen exposure drives accumulation of long-lived IgE plasma cells in the bone marrow, giving rise to serological memory. Sci. Immunol. 2020, 5, eaav8402. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.U.; Ogunniyi, A.O.; Calatroni, A.; Tadigotla, V.R.; Ruiter, B.; Ma, A.; Moon, J.; Love, J.C.; Shreffler, W.G. Peanut oral immunotherapy transiently expands circulating Ara h 2-specific B cells with a homologous repertoire in unrelated subjects. J. Allergy Clin. Immunol. 2015, 136, 125–134.e12. [Google Scholar] [CrossRef] [PubMed]
- Hoh, R.A.; Joshi, S.A.; Liu, Y.; Wang, C.; Roskin, K.M.; Lee, J.Y.; Pham, T.; Looney, T.J.; Jackson, K.J.; Dixit, V.P.; et al. Single B-cell deconvolution of peanut-specific antibody responses in allergic patients. J. Allergy Clin. Immunol. 2016, 137, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Croote, D.; Darmanis, S.; Nadeau, K.C.; Quake, S.R. High-affinity allergen-specific human antibodies cloned from single IgE B cell transcriptomes. Science 2018, 362, 1306–1309. [Google Scholar] [CrossRef]
- Saunders, S.P.; Ma, E.G.M.; Aranda, C.J.; Curotto de Lafaille, M.A. Non-classical B Cell Memory of Allergic IgE Responses. Front. Immunol. 2019, 10, 715. [Google Scholar] [CrossRef]
- Fernandes-Braga, W.; Curotto de Lafaille, M.A. B cell memory of Immunoglobulin E (IgE) antibody responses in allergy. Curr. Opin. Immunol. 2024, 91, 102488. [Google Scholar] [CrossRef]
- Mitre, E.; Nutman, T.B. IgE memory: Persistence of antigen-specific IgE responses years after treatment of human filarial infections. J. Allergy Clin. Immunol. 2006, 117, 939–945. [Google Scholar] [CrossRef]
- Achatz, G.; Nitschke, L.; Lamers, M.C. Effect of transmembrane and cytoplasmic domains of IgE on the IgE response. Science 1997, 276, 409–411. [Google Scholar] [CrossRef]
- Le Gros, G.; Schultze, N.; Walti, S.; Einsle, K.; Finkelman, F.; Kosco-Vilbois, M.H.; Heusser, C. The development of IgE+ memory B cells following primary IgE immune responses. Eur. J. Immunol. 1996, 26, 3042–3047. [Google Scholar] [CrossRef]
- Satitsuksanoa, P.; Daanje, M.; Akdis, M.; Boyd, S.D.; van de Veen, W. Biology and dynamics of B cells in the context of IgE-mediated food allergy. Allergy 2021, 76, 1707–1717. [Google Scholar] [CrossRef]
- He, J.S.; Subramaniam, S.; Narang, V.; Srinivasan, K.; Saunders, S.P.; Carbajo, D.; Wen-Shan, T.; Hidayah Hamadee, N.; Lum, J.; Lee, A.; et al. IgG1 memory B cells keep the memory of IgE responses. Nat. Commun. 2017, 8, 641. [Google Scholar] [CrossRef] [PubMed]
- Eisen, H.N.; Siskind, G.W. Variations in Affinities of Antibodies during the Immune Response. Biochemistry 1964, 3, 996–1008. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Sullivan, B.M.; Allen, C.D. Fluorescent in vivo detection reveals that IgE(+) B cells are restrained by an intrinsic cell fate predisposition. Immunity 2012, 36, 857–872. [Google Scholar] [CrossRef] [PubMed]
- Turqueti-Neves, A.; Otte, M.; Schwartz, C.; Schmitt, M.E.; Lindner, C.; Pabst, O.; Yu, P.; Voehringer, D. The Extracellular Domains of IgG1 and T Cell-Derived IL-4/IL-13 Are Critical for the Polyclonal Memory IgE Response In Vivo. PLoS Biol. 2015, 13, e1002290. [Google Scholar] [CrossRef]
- Ramadani, F.; Bowen, H.; Gould, H.J.; Fear, D.J. Transcriptional Analysis of the Human IgE-Expressing Plasma Cell Differentiation Pathway. Front. Immunol. 2019, 10, 402. [Google Scholar] [CrossRef]
- Zuccarino-Catania, G.V.; Sadanand, S.; Weisel, F.J.; Tomayko, M.M.; Meng, H.; Kleinstein, S.H.; Good-Jacobson, K.L.; Shlomchik, M.J. CD80 and PD-L2 define functionally distinct memory B cell subsets that are independent of antibody isotype. Nat. Immunol. 2014, 15, 631–637. [Google Scholar] [CrossRef]
- Bar-Or, A.; Oliveira, E.M.; Anderson, D.E.; Krieger, J.I.; Duddy, M.; O’Connor, K.C.; Hafler, D.A. Immunological memory: Contribution of memory B cells expressing costimulatory molecules in the resting state. J. Immunol. 2001, 167, 5669–5677. [Google Scholar] [CrossRef]
- Ota, M.; Hoehn, K.B.; Fernandes-Braga, W.; Ota, T.; Aranda, C.J.; Friedman, S.; Miranda-Waldetario, M.G.C.; Redes, J.; Suprun, M.; Grishina, G.; et al. CD23(+)IgG1(+) memory B cells are poised to switch to pathogenic IgE production in food allergy. Sci. Transl. Med. 2024, 16, eadi0673. [Google Scholar] [CrossRef]
- Aranda, C.J.; Gonzalez-Kozlova, E.; Saunders, S.P.; Fernandes-Braga, W.; Ota, M.; Narayanan, S.; He, J.S.; Del Duca, E.; Swaroop, B.; Gnjatic, S.; et al. IgG memory B cells expressing IL4R and FCER2 are associated with atopic diseases. Allergy 2023, 78, 752–766. [Google Scholar] [CrossRef]
- Koenig, J.F.E.; Knudsen, N.P.H.; Phelps, A.; Bruton, K.; Hoof, I.; Lund, G.; Libera, D.D.; Lund, A.; Christensen, L.H.; Glass, D.R.; et al. Type 2-polarized memory B cells hold allergen-specific IgE memory. Sci. Transl. Med. 2024, 16, eadi0944. [Google Scholar] [CrossRef]
- Heeringa, J.J.; Rijvers, L.; Arends, N.J.; Driessen, G.J.; Pasmans, S.G.; van Dongen, J.J.M.; de Jongste, J.C.; van Zelm, M.C. IgE-expressing memory B cells and plasmablasts are increased in blood of children with asthma, food allergy, and atopic dermatitis. Allergy 2018, 73, 1331–1336. [Google Scholar] [CrossRef] [PubMed]
- Berkowska, M.A.; Heeringa, J.J.; Hajdarbegovic, E.; van der Burg, M.; Thio, H.B.; van Hagen, P.M.; Boon, L.; Orfao, A.; van Dongen, J.J.; van Zelm, M.C. Human IgE(+) B cells are derived from T cell-dependent and T cell-independent pathways. J. Allergy Clin. Immunol. 2014, 134, 688–697.e6. [Google Scholar] [CrossRef] [PubMed]
- Leung, D.Y.; Sampson, H.A.; Yunginger, J.W.; Burks, A.W., Jr.; Schneider, L.C.; Wortel, C.H.; Davis, F.M.; Hyun, J.D.; Shanahan, W.R., Jr.; Avon Longitudinal Study of, P.; et al. Effect of anti-IgE therapy in patients with peanut allergy. N. Engl. J. Med. 2003, 348, 986–993. [Google Scholar] [CrossRef] [PubMed]
- Dantzer, J.A.; Wood, R.A. Anti-IgE and food allergy. J. Allergy Clin. Immunol. 2025, 155, 1–11. [Google Scholar] [CrossRef]
- Azzano, P.; Paquin, M.; Langlois, A.; Morin, C.; Parizeault, G.; Lacombe-Barrios, J.; Samaan, K.; Graham, F.; Paradis, L.; Des Roches, A.; et al. Determinants of omalizumab dose-related efficacy in oral immunotherapy: Evidence from a cohort of 181 patients. J. Allergy Clin. Immunol. 2021, 147, 233–243. [Google Scholar] [CrossRef]
- Sampson, H.A.; Leung, D.Y.; Burks, A.W.; Lack, G.; Bahna, S.L.; Jones, S.M.; Wong, D.A. A phase II, randomized, double-blind, parallel-group, placebo-controlled oral food challenge trial of Xolair (omalizumab) in peanut allergy. J. Allergy Clin. Immunol. 2011, 127, 1309–1310.e1. [Google Scholar] [CrossRef]
- Wood, R.A.; Togias, A.; Sicherer, S.H.; Shreffler, W.G.; Kim, E.H.; Jones, S.M.; Leung, D.Y.M.; Vickery, B.P.; Bird, J.A.; Spergel, J.M.; et al. Omalizumab for the Treatment of Multiple Food Allergies. N. Engl. J. Med. 2024, 390, 889–899. [Google Scholar] [CrossRef]
- Eggel, A.; Pennington, L.F.; Jardetzky, T.S. Therapeutic monoclonal antibodies in allergy: Targeting IgE, cytokine, and alarmin pathways. Immunol. Rev. 2024, 328, 387–411. [Google Scholar] [CrossRef]
- Feuille, E.; Nowak-Wegrzyn, A. Allergen-Specific Immunotherapies for Food Allergy. Allergy Asthma Immunol. Res. 2018, 10, 189–206. [Google Scholar] [CrossRef]
- Ozdemir, P.G.; Sato, S.; Yanagida, N.; Ebisawa, M. Oral Immunotherapy in Food Allergy: Where Are We Now? Allergy Asthma Immunol. Res. 2023, 15, 125–144. [Google Scholar] [CrossRef]
- Csonka, P.; Lee, B.; Kuitunen, I. Epicutaneous immunotherapy for food allergy: A systematic review and meta-analysis. Clin. Transl. Allergy 2025, 15, e70045. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Nagakura, K.I.; Yanagida, N.; Ebisawa, M. Current perspective on allergen immunotherapy for food allergies. Allergol. Int. 2024, 73, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Pouessel, G.; Lezmi, G. Oral immunotherapy for food allergy: Translation from studies to clinical practice? World Allergy Organ. J. 2023, 16, 100747. [Google Scholar] [CrossRef] [PubMed]
- Burks, A.W.; Jones, S.M.; Wood, R.A.; Fleischer, D.M.; Sicherer, S.H.; Lindblad, R.W.; Stablein, D.; Henning, A.K.; Vickery, B.P.; Liu, A.H.; et al. Oral immunotherapy for treatment of egg allergy in children. N. Engl. J. Med. 2012, 367, 233–243. [Google Scholar] [CrossRef]
- Smith, S.S.; Hilas, O. Peanut (Arachis hypogaea) Allergen Powder-dnfp: The First FDA-approved Oral Immunotherapy for Desensitization of Peanut Allergy in Children. J. Pediatr. Pharmacol. Ther. 2021, 26, 669–674. [Google Scholar] [CrossRef]
- The Palisade Group of Clinical Investigators; Vickery, B.P.; Vereda, A.; Casale, T.B.; Beyer, K.; du Toit, G.; Hourihane, J.O.; Jones, S.M.; Shreffler, W.G.; Marcantonio, A.; et al. AR101 Oral Immunotherapy for Peanut Allergy. N. Engl. J. Med. 2018, 379, 1991–2001. [Google Scholar] [CrossRef]
- Ciaccio, C.; Goldsobel, A.B.; Anagnostou, A.; Beyer, K.; Casale, T.B.; Deschildre, A.; Fernández-Rivas, M.; Hourihane, J.O.; Krawiec, M.; Lieberman, J.; et al. Participant characteristics and safety outcomes of peanut oral immunotherapy in the RAMSES and ARC011 trials. Ann. Allergy Asthma Immunol. 2022, 129, 758–768.e4. [Google Scholar] [CrossRef]
- Du Toit, G.; Brown, K.R.; Vereda, A.; Irani, A.M.; Tilles, S.; Ratnayake, A.; Jones, S.M.; Vickery, B.P. Oral Immunotherapy for Peanut Allergy in Children 1 to Less Than 4 Years of Age. NEJM Evid. 2023, 2, EVIDoa2300145. [Google Scholar] [CrossRef]
- Uhl, C.; Klevebro, S.; Sverremark-Ekstrom, E.; Tedner, S.G.; Brandstrom, J.; Papageorgiou, C.; Melen, E.; Konradsen, J.R.; Nilsson, C.; Asarnoj, A. High Degree of Desensitization After 1 Year of Early-Life Peanut Oral Immunotherapy: Small Children Oral Immunotherapy (SmaChO) Randomized Controlled Trial. J. Allergy Clin. Immunol. Pract. 2024, 12, 1297–1305. [Google Scholar] [CrossRef]
- Sicherer, S.H.; Bunyavanich, S.; Berin, M.C.; Lo, T.; Groetch, M.; Schaible, A.; Perry, S.A.; Wheatley, L.M.; Fulkerson, P.C.; Chang, H.L.; et al. Peanut Oral Immunotherapy in Children with High-Threshold Peanut Allergy. NEJM Evid. 2025, 4, EVIDoa2400306. [Google Scholar] [CrossRef]
- Lee, A.S.E.; Baker, M.G.; Cox, A.L.; Oriel, R.C.; Tsuang, A.; Sicherer, S.H.; Kattan, J.D. Long-term follow-up of children who achieved sustained unresponsiveness after peanut oral immunotherapy. J. Allergy Clin. Immunol. practice 2024, 12, 255–256. [Google Scholar] [CrossRef] [PubMed]
- Satitsuksanoa, P.; van de Veen, W.; Tan, G.; Lopez, J.F.; Wirz, O.; Jansen, K.; Sokolowska, M.; Mirer, D.; Globinska, A.; Boonpiyathad, T.; et al. Allergen-specific B cell responses in oral immunotherapy-induced desensitization, remission, and natural outgrowth in cow’s milk allergy. Allergy 2025, 80, 161–180. [Google Scholar] [CrossRef] [PubMed]
- Vickery, B.P.; Lin, J.; Kulis, M.; Fu, Z.; Steele, P.H.; Jones, S.M.; Scurlock, A.M.; Gimenez, G.; Bardina, L.; Sampson, H.A.; et al. Peanut oral immunotherapy modifies IgE and IgG4 responses to major peanut allergens. J. Allergy Clin. Immunol. 2013, 131, 128–134.e3. [Google Scholar] [CrossRef] [PubMed]
- Akdis, C.A.; Akdis, M. Mechanisms of immune tolerance to allergens: Role of IL-10 and Tregs. J. Clin. Investig. 2014, 124, 4678–4680. [Google Scholar] [CrossRef]
- McKenzie, C.I.; Reinwald, S.; Averso, B.; Spurrier, B.; Satz, A.; von Borstel, A.; Masinovic, S.; Varese, N.; Aui, P.M.; Wines, B.D.; et al. Subcutaneous immunotherapy for bee venom allergy induces epitope spreading and immunophenotypic changes in allergen-specific memory B cells. J. Allergy Clin. Immunol. 2024, 154, 1511–1522. [Google Scholar] [CrossRef]
- McKenzie, C.I.; Varese, N.; Aui, P.M.; Reinwald, S.; Wines, B.D.; Hogarth, P.M.; Thien, F.; Hew, M.; Rolland, J.M.; O’Hehir, R.E.; et al. RNA sequencing of single allergen-specific memory B cells after grass pollen immunotherapy: Two unique cell fates and CD29 as a biomarker for treatment effect. Allergy 2023, 78, 822–835. [Google Scholar] [CrossRef]
- Suprun, M.; Lee, A.S.E.; Getts, R.; Peck, S.; Sindher, S.B.; Nadeau, K.C.; Chinthrajah, R.S.; Galli, S.J.; Sampson, H.A. Baseline epitope-specific IgE profiles are predictive of sustained unresponsiveness or high threshold 1-year post oral immunotherapy in the POISED trial. J. Allergy Clin. Immunol. 2025, 155, 923–931.e2. [Google Scholar] [CrossRef]
- Vickery, B.P.; Scurlock, A.M.; Kulis, M.; Steele, P.H.; Kamilaris, J.; Berglund, J.P.; Burk, C.; Hiegel, A.; Carlisle, S.; Christie, L.; et al. Sustained unresponsiveness to peanut in subjects who have completed peanut oral immunotherapy. J. Allergy Clin. Immunol. 2014, 133, 468–475. [Google Scholar] [CrossRef]
- Patil, S.U.; Steinbrecher, J.; Calatroni, A.; Smith, N.; Ma, A.; Ruiter, B.; Virkud, Y.; Schneider, M.; Shreffler, W.G. Early decrease in basophil sensitivity to Ara h 2 precedes sustained unresponsiveness after peanut oral immunotherapy. J. Allergy Clin. Immunol. 2019, 144, 1310–1319.e4. [Google Scholar] [CrossRef]
- Suarez-Farinas, M.; Suprun, M.; Bahnson, H.T.; Raghunathan, R.; Getts, R.; duToit, G.; Lack, G.; Sampson, H.A. Evolution of epitope-specific IgE and IgG(4) antibodies in children enrolled in the LEAP trial. J. Allergy Clin. Immunol. 2021, 148, 835–842. [Google Scholar] [CrossRef]
- Du Toit, G.; Roberts, G.; Sayre, P.H.; Bahnson, H.T.; Radulovic, S.; Santos, A.F.; Brough, H.A.; Phippard, D.; Basting, M.; Feeney, M.; et al. Randomized trial of peanut consumption in infants at risk for peanut allergy. N. Engl. J. Med. 2015, 372, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, G.; Sayre, P.H.; Roberts, G.; Sever, M.L.; Lawson, K.; Bahnson, H.T.; Brough, H.A.; Santos, A.F.; Harris, K.M.; Radulovic, S.; et al. Effect of Avoidance on Peanut Allergy after Early Peanut Consumption. N. Engl. J. Med. 2016, 374, 1435–1443. [Google Scholar] [CrossRef] [PubMed]
- Brandtzaeg, P. Molecular and cellular aspects of the secretory immunoglobulin system. APMIS 1995, 103, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Turk, A.; Lichtenstein, L.M.; Norman, P.S. Nasal secretory antibody to inhalant allergens in allergic and non-allergic patients. Immunology 1970, 19, 85–95. [Google Scholar]
- Smeekens, J.M.; Baloh, C.; Lim, N.; Larson, D.; Qin, T.; Wheatley, L.; Kim, E.H.; Jones, S.M.; Burks, A.W.; Kulis, M.D. Peanut-Specific IgG4 and IgA in Saliva Are Modulated by Peanut Oral Immunotherapy. J. Allergy Clin. Immunol. Pract. 2022, 10, 3270–3275. [Google Scholar] [CrossRef]
- El Ansari, Y.S.; Kanagaratham, C.; Burton, O.T.; Santos, J.V.; Hollister, B.A.; Lewis, O.L.; Renz, H.; Oettgen, H.C. Allergen-Specific IgA Antibodies Block IgE-Mediated Activation of Mast Cells and Basophils. Front. Immunol. 2022, 13, 881655. [Google Scholar] [CrossRef]
- Jarvinen, K.M.; Westfall, J.E.; Seppo, M.S.; James, A.K.; Tsuang, A.J.; Feustel, P.J.; Sampson, H.A.; Berin, C. Role of maternal elimination diets and human milk IgA in the development of cow’s milk allergy in the infants. Clin. Exp. Allergy 2014, 44, 69–78. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, E.; Gertie, J.A.; Joseph, J.; Xu, L.; Pinker, E.Y.; Waizman, D.A.; Catanzaro, J.; Hamza, K.H.; Lahl, K.; et al. Divergent T follicular helper cell requirement for IgA and IgE production to peanut during allergic sensitization. Sci. Immunol. 2020, 5, eaay2754. [Google Scholar] [CrossRef]
- Liu, E.G.; Zhang, B.; Martin, V.; Anthonypillai, J.; Kraft, M.; Grishin, A.; Grishina, G.; Catanzaro, J.R.; Chinthrajah, S.; Sindher, T.; et al. Food-specific immunoglobulin A does not correlate with natural tolerance to peanut or egg allergens. Sci. Transl. Med. 2022, 14, eabq0599. [Google Scholar] [CrossRef]
- Kulis, M.D.; Patil, S.U.; Wambre, E.; Vickery, B.P. Immune mechanisms of oral immunotherapy. J. Allergy Clin. Immunol. 2018, 141, 491–498. [Google Scholar] [CrossRef]
- Burton, O.T.; Logsdon, S.L.; Zhou, J.S.; Medina-Tamayo, J.; Abdel-Gadir, A.; Noval Rivas, M.; Koleoglou, K.J.; Chatila, T.A.; Schneider, L.C.; Rachid, R.; et al. Oral immunotherapy induces IgG antibodies that act through FcgammaRIIb to suppress IgE-mediated hypersensitivity. J. Allergy Clin. Immunol. 2014, 134, 1310–1317.e6. [Google Scholar] [CrossRef] [PubMed]
- James, L.K.; Shamji, M.H.; Walker, S.M.; Wilson, D.R.; Wachholz, P.A.; Francis, J.N.; Jacobson, M.R.; Kimber, I.; Till, S.J.; Durham, S.R. Long-term tolerance after allergen immunotherapy is accompanied by selective persistence of blocking antibodies. J. Allergy Clin. Immunol. 2011, 127, 509–516.e5. [Google Scholar] [CrossRef] [PubMed]
- Nouri-Aria, K.T.; Wachholz, P.A.; Francis, J.N.; Jacobson, M.R.; Walker, S.M.; Wilcock, L.K.; Staple, S.Q.; Aalberse, R.C.; Till, S.J.; Durham, S.R. Grass pollen immunotherapy induces mucosal and peripheral IL-10 responses and blocking IgG activity. J. Immunol. 2004, 172, 3252–3259. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.F.; James, L.K.; Bahnson, H.T.; Shamji, M.H.; Couto-Francisco, N.C.; Islam, S.; Houghton, S.; Clark, A.T.; Stephens, A.; Turcanu, V.; et al. IgG4 inhibits peanut-induced basophil and mast cell activation in peanut-tolerant children sensitized to peanut major allergens. J. Allergy Clin. Immunol. 2015, 135, 1249–1256. [Google Scholar] [CrossRef]
- Keswani, T.; LaHood, N.A.; Marini-Rapoport, O.; Karmakar, B.; Andrieux, L.; Reese, B.; Sneed, S.L.; Pedersen, L.C.; Mueller, G.A.; Patil, S.U. Neutralizing IgG(4) antibodies are a biomarker of sustained efficacy after peanut oral immunotherapy. J. Allergy Clin. Immunol. 2024, 153, 1611–1620.e7. [Google Scholar] [CrossRef]
- Strobl, M.R.; Demir, H.; Sanchez Acosta, G.; Drescher, A.; Kitzmuller, C.; Mobs, C.; Pfutzner, W.; Bohle, B. The role of IgG(1) and IgG(4) as dominant IgE-blocking antibodies shifts during allergen immunotherapy. J. Allergy Clin. Immunol. 2023, 151, 1371–1378.e5. [Google Scholar] [CrossRef]
- Smith, S.A.; Khan, Y.W.; Shrem, R.A.; Hemler, J.A.; Doyle, J.E.; Daniel, J.; Zhang, J.; Pena-Amelunxen, G.; Aglas, L.; Hamilton, R.G.; et al. Antigenic determinants underlying IgE-mediated anaphylaxis to peanut. J. Allergy Clin. Immunol. 2025, 155, 1595–1606.e10. [Google Scholar] [CrossRef]
- Croote, D.; Wong, J.J.W.; Pecalvel, C.; Leveque, E.; Casanovas, N.; Kamphuis, J.B.J.; Creeks, P.; Romero, J.; Sohail, S.; Bedinger, D.; et al. Widespread monoclonal IgE antibody convergence to an immunodominant, proanaphylactic Ara h 2 epitope in peanut allergy. J. Allergy Clin. Immunol. 2024, 153, 182–192.e7. [Google Scholar] [CrossRef]
- Sackesen, C.; Suarez-Farinas, M.; Silva, R.; Lin, J.; Schmidt, S.; Getts, R.; Gimenez, G.; Yilmaz, E.A.; Cavkaytar, O.; Buyuktiryaki, B.; et al. A new Luminex-based peptide assay to identify reactivity to baked, fermented, and whole milk. Allergy 2019, 74, 327–336. [Google Scholar] [CrossRef]
- Suprun, M.; Getts, R.; Raghunathan, R.; Grishina, G.; Witmer, M.; Gimenez, G.; Sampson, H.A.; Suarez-Farinas, M. Novel Bead-Based Epitope Assay is a sensitive and reliable tool for profiling epitope-specific antibody repertoire in food allergy. Sci. Rep. 2019, 9, 18425. [Google Scholar] [CrossRef]
- Ho, H.E.; Arditi, Z.; Radigan, L.; Grishina, G.; Zhang, L.; Chun, Y.; Lo, T.; Wang, J.; Sicherer, S.; Bunyavanich, S. Saliva antibody profiles are associated with reaction threshold and severity of peanut allergic reactions. J. Allergy Clin. Immunol. 2024, 154, 690–697.e4. [Google Scholar] [CrossRef] [PubMed]
- Landhuis, E. IgGenix: Engineering antibodies to fight food allergies. Nat. Biotechnol. 2021. [Google Scholar] [CrossRef] [PubMed]
- LaHood, N.A.; Min, J.; Keswani, T.; Richardson, C.M.; Amoako, K.; Zhou, J.; Marini-Rapoport, O.; Bernard, H.; Hazebrouck, S.; Shreffler, W.G.; et al. Immunotherapy-induced neutralizing antibodies disrupt allergen binding and sustain allergen tolerance in peanut allergy. J. Clin. Investig. 2023, 133, e164501. [Google Scholar] [CrossRef] [PubMed]
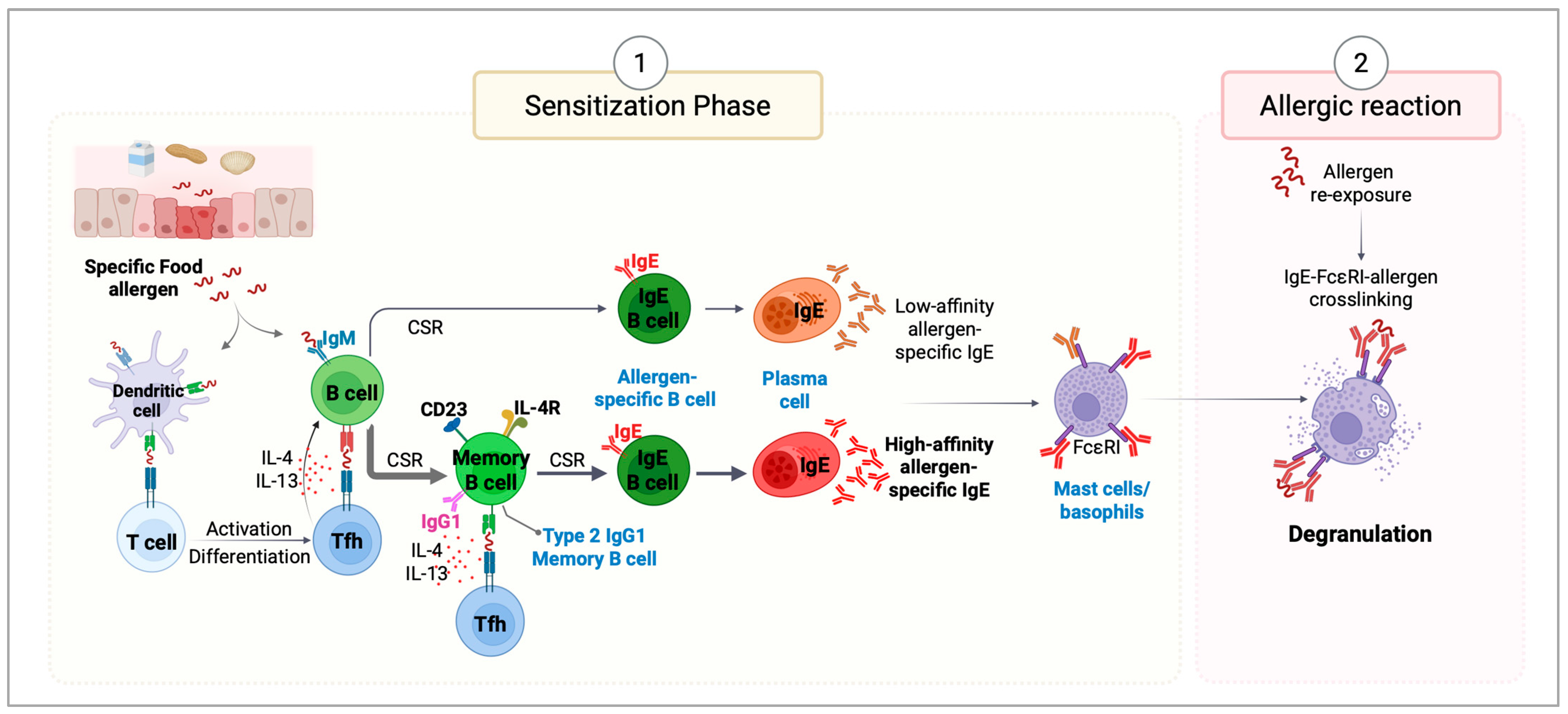
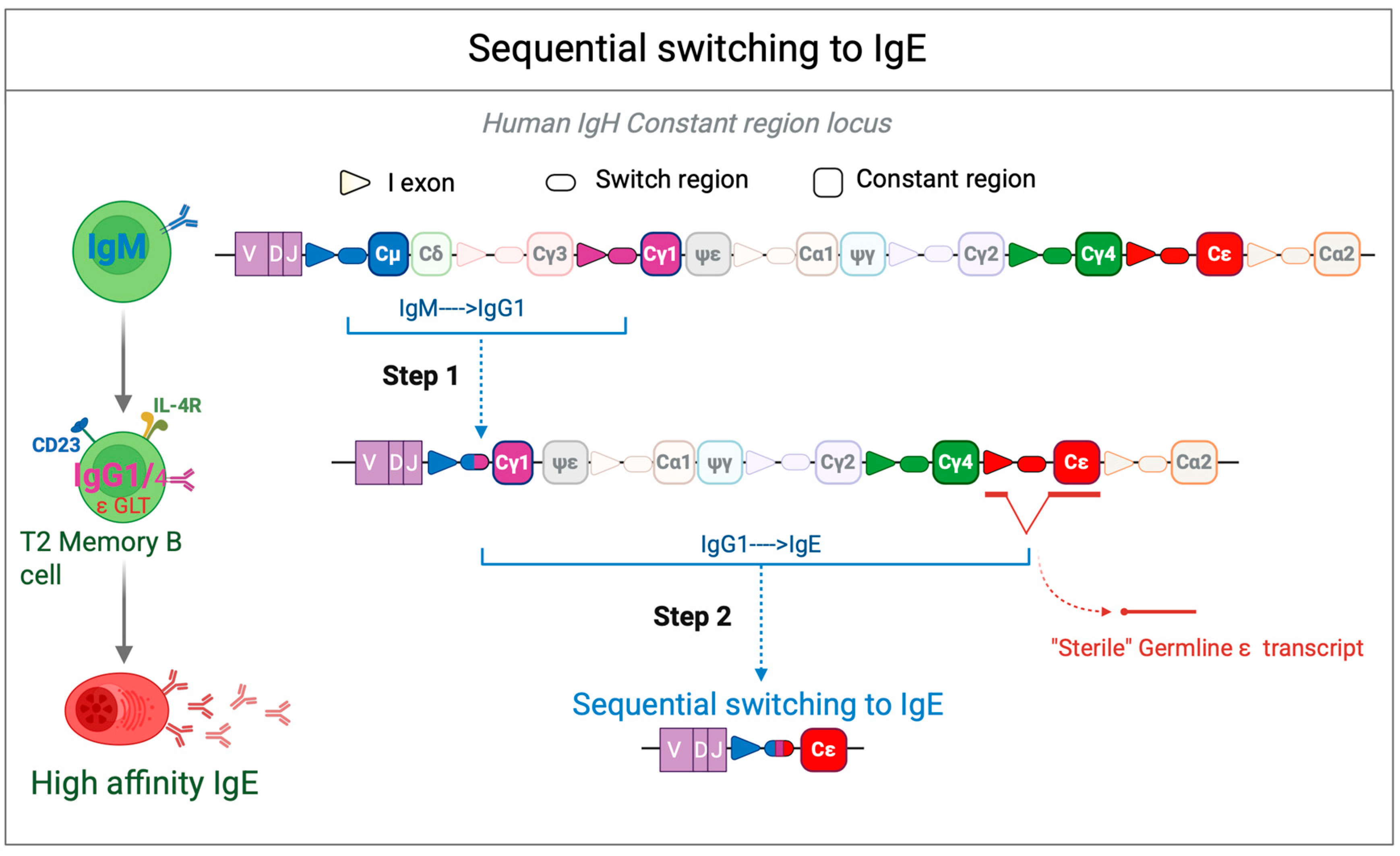
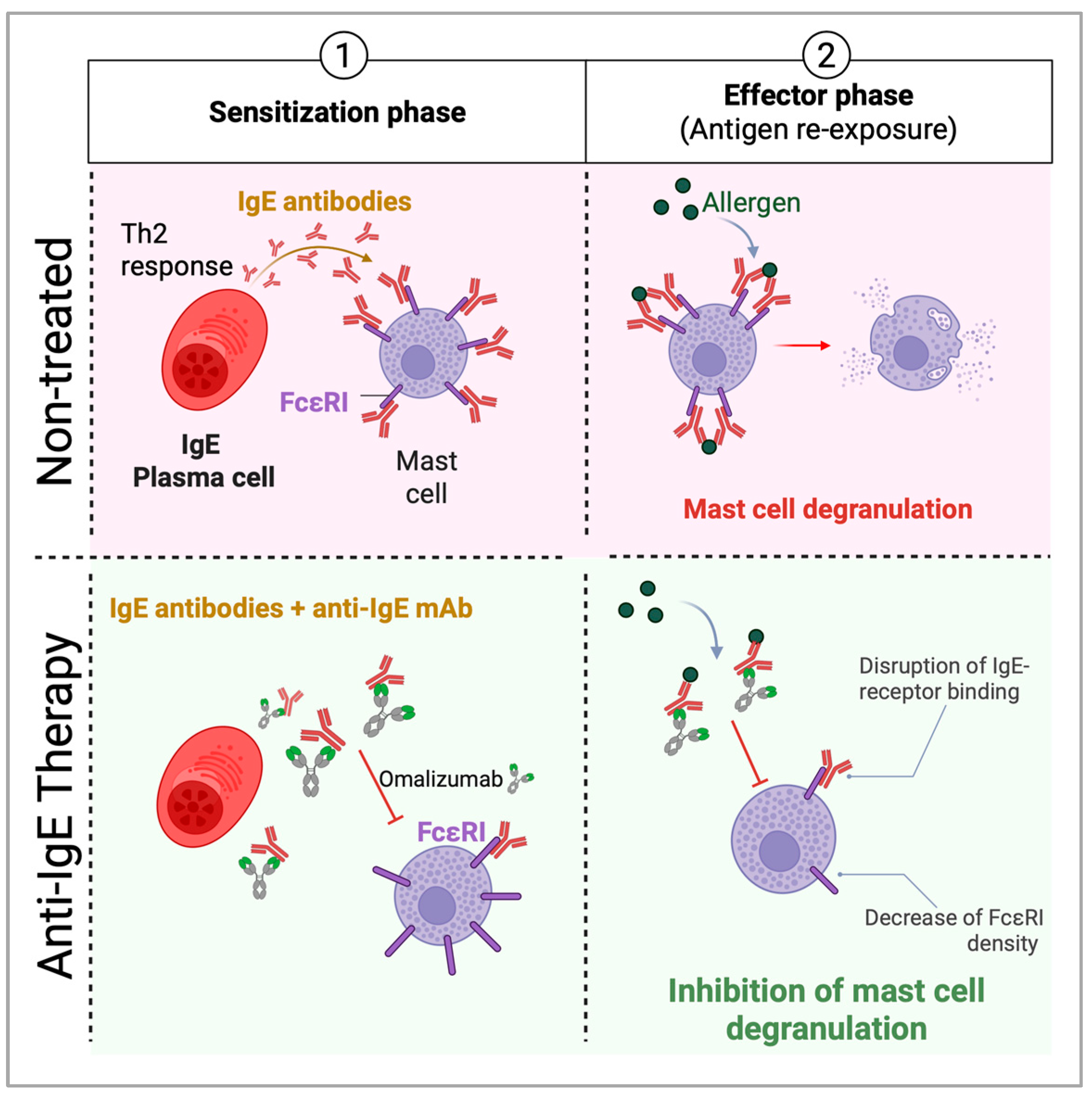
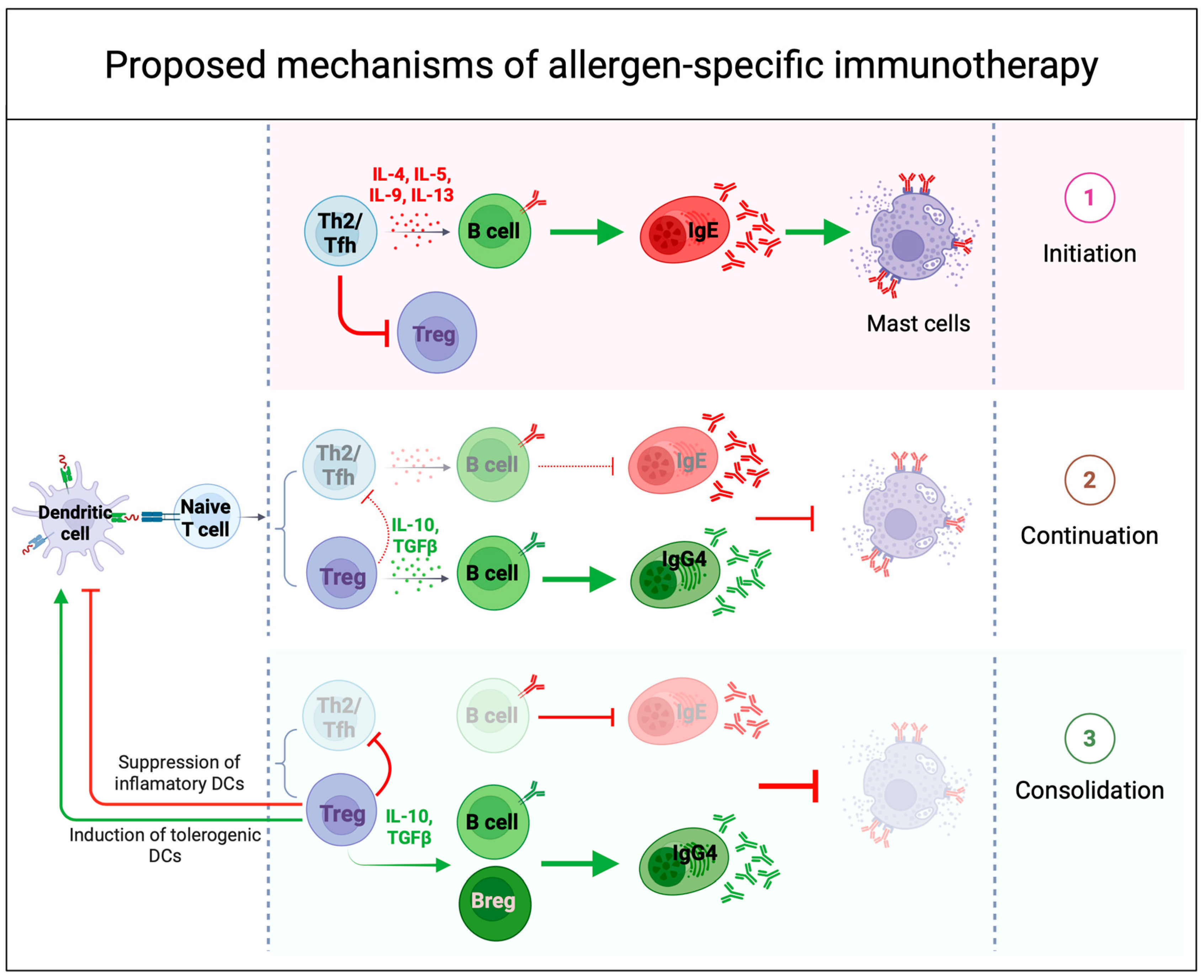
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Carmona, Y.; Curotto de Lafaille, M.A. Advances in Food Allergy Immunotherapy: Current Strategies and Role of Antibodies Isotypes. Cells 2025, 14, 900. https://doi.org/10.3390/cells14120900
Garcia-Carmona Y, Curotto de Lafaille MA. Advances in Food Allergy Immunotherapy: Current Strategies and Role of Antibodies Isotypes. Cells. 2025; 14(12):900. https://doi.org/10.3390/cells14120900
Chicago/Turabian StyleGarcia-Carmona, Yolanda, and Maria A. Curotto de Lafaille. 2025. "Advances in Food Allergy Immunotherapy: Current Strategies and Role of Antibodies Isotypes" Cells 14, no. 12: 900. https://doi.org/10.3390/cells14120900
APA StyleGarcia-Carmona, Y., & Curotto de Lafaille, M. A. (2025). Advances in Food Allergy Immunotherapy: Current Strategies and Role of Antibodies Isotypes. Cells, 14(12), 900. https://doi.org/10.3390/cells14120900