
Molecular Fingerprint of Endocannabinoid Signaling in the Developing Paraventricular Nucleus of the Hypothalamus as Revealed by Single-Cell RNA-Seq and In Situ Hybridization

 , ,
, , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 2-AG | 2-arachidonoylglycerol |
| 3V | Third ventricle |
| Abhd4/6/12 (ABHD4/6/12) | α/β-hydrolase domain containing 4/6/12 |
| AEA | Anandamide |
| AHN | Anterior hypothalamic nucleus |
| ATP | Adenosine triphosphate |
| Avp (AVP) | Arginine vasopressin |
| Cnr1/2 (CB1R) | Cannabinoid receptor type 1/2 |
| Cox2 (COX2) | Cyclooxygenase-2 |
| Crh (CRH) | Corticotrophin-releasing hormone |
| Dagla/b (DAGLα/β) | Sn1-specific diacylglycerol lipase alpha/beta |
| E | Embryonic |
| Faah (FAAH) | Fatty-acid amide hydrolase |
| Gde1 (GDE1) | Glycerophosphodiester Phosphodiesterase 1 |
| Gpr55 (GPR55) | G protein-coupled receptor 55 |
| HPA | Hypothalamic–pituitary–adrenal |
| Lox (LOX) | Lipoxygenase |
| MS | Medial septum |
| Mgll (MGLL) | Monoacylglycerol lipase |
| Napepld (NAPE-PLD) | N-acyl phosphatidylethanolamine phospholipase D |
| OB | Olfactory bulb |
| OT | Olfactory tract |
| P | Postnatal |
| Ptpn22 (PTPN22) | Protein tyrosine phosphatase non-receptor type 22 |
| PVN | Paraventricular nucleus |
| SCN | Suprachiasmatic nucleus |
| Sst (SST) | Somatostatin |
| Trh (TRH) | Thyrotropin-releasing hormone |
| Oxt (OXT) | Oxytocin |
| UMAP | Uniform Manifold Approximation and Projection |
| UpSet | Visualization of Intersecting Sets |
References
- Fong, H.; Zheng, J.; Kurrasch, D. The structural and functional complexity of the integrative hypothalamus. Science 2023, 382, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Li, J.; Tang, K. The Paraventricular Nucleus of the Hypothalamus: Development, Function, and Human Diseases. Endocrinology 2018, 159, 3458–3472. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shlomo, A.; Melmed, S. Pituitary Somatostatin Receptor Signaling. Trends Endocrinol. Metab. TEM 2010, 21, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Sun, L.; You, J.; Peng, H.; Yan, H.; Wang, J.; Sun, F.; Cui, M.; Wang, S.; Zhang, Z.; et al. Role and mechanism of PVN-sympathetic-adipose circuit in depression and insulin resistance induced by chronic stress. EMBO Rep. 2023, 24, e57176. [Google Scholar] [CrossRef]
- Li, Y.-J.; Du, W.-J.; Liu, R.; Zan, G.-Y.; Ye, B.-L.; Li, Q.; Sheng, Z.-H.; Yuan, Y.-W.; Song, Y.-J.; Liu, J.-G.; et al. Paraventricular nucleus-central amygdala oxytocinergic projection modulates pain-related anxiety-like behaviors in mice. CNS Neurosci. Ther. 2023, 29, 3493–3506. [Google Scholar] [CrossRef]
- Fenselau, H.; Campbell, J.N.; Verstegen, A.M.J.; Madara, J.C.; Xu, J.; Shah, B.P.; Resch, J.M.; Yang, Z.; Mandelblat-Cerf, Y.; Livneh, Y.; et al. A rapidly acting glutamatergic ARC→PVH satiety circuit postsynaptically regulated by α-MSH. Nat. Neurosci. 2017, 20, 42–51. [Google Scholar] [CrossRef]
- Krashes, M.J.; Shah, B.P.; Koda, S.; Lowell, B.B. Rapid versus Delayed Stimulation of Feeding by the Endogenously Released AgRP Neuron Mediators GABA, NPY, and AgRP. Cell Metab. 2013, 18, 588–595. [Google Scholar] [CrossRef]
- Cowley, M.A.; Smart, J.L.; Rubinstein, M.; Cerdán, M.G.; Diano, S.; Horvath, T.L.; Cone, R.D.; Low, M.J. Leptin activates anorexigenic POMC neurons through a neural network in the arcuate nucleus. Nature 2001, 411, 480–484. [Google Scholar] [CrossRef]
- Caron, A.; Dungan Lemko, H.M.; Castorena, C.M.; Fujikawa, T.; Lee, S.; Lord, C.C.; Ahmed, N.; Lee, C.E.; Holland, W.L.; Liu, C.; et al. POMC neurons expressing leptin receptors coordinate metabolic responses to fasting via suppression of leptin levels. eLife 2018, 7, e33710. [Google Scholar] [CrossRef]
- Kondoh, K.; Lu, Z.; Ye, X.; Olson, D.P.; Lowell, B.B.; Buck, L.B. A specific area of olfactory cortex involved in stress hormone responses to predator odours. Nature 2016, 532, 103–106. [Google Scholar] [CrossRef]
- Kaouane, N.; Ada, S.; Hausleitner, M.; Haubensak, W. Dorsal Bed Nucleus of the Stria Terminalis-Subcortical Output Circuits Encode Positive Bias in Pavlovian Fear and Reward. Front. Neural Circuits 2021, 15, 772512. [Google Scholar] [CrossRef]
- Decavel, C.; Van Den Pol, A.N. GABA: A dominant neurotransmitter in the hypothalamus. J. Comp. Neurol. 1990, 302, 1019–1037. [Google Scholar] [CrossRef] [PubMed]
- Mendonça, M.M.; Santana, J.S.; da Cruz, K.R.; Ianzer, D.; Ghedini, P.C.; Nalivaiko, E.; Fontes, M.A.P.; Ferreira, R.N.; Pedrino, G.R.; Colugnati, D.B.; et al. Involvement of GABAergic and Adrenergic Neurotransmissions on Paraventricular Nucleus of Hypothalamus in the Control of Cardiac Function. Front. Physiol. 2018, 9, 670. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-F.; Jackson, K.L.; Stern, J.E.; Rabeler, B.; Patel, K.P. Interaction between glutamate and GABA systems in the integration of sympathetic outflow by the paraventricular nucleus of the hypothalamus. Am. J. Physiol.-Heart Circ. Physiol. 2006, 291, H2847–H2856. [Google Scholar] [CrossRef]
- Romanov, R.A.; Tretiakov, E.O.; Kastriti, M.E.; Zupancic, M.; Häring, M.; Korchynska, S.; Popadin, K.; Benevento, M.; Rebernik, P.; Lallemend, F.; et al. Molecular design of hypothalamus development. Nature 2020, 582, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, L.A.; Lolait, S.J.; Brownstein, M.J.; Young, A.C.; Bonner, T.I. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 1990, 346, 561–564. [Google Scholar] [CrossRef]
- Mechoulam, R.; Ben-Shabat, S.; Hanus, L.; Ligumsky, M.; Kaminski, N.E.; Schatz, A.R.; Gopher, A.; Almog, S.; Martin, B.R.; Compton, D.R. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem. Pharmacol. 1995, 50, 83–90. [Google Scholar] [CrossRef]
- Stella, N.; Schweitzer, P.; Piomelli, D. A second endogenous cannabinoid that modulates long-term potentiation. Nature 1997, 388, 773–778. [Google Scholar] [CrossRef]
- Kano, M.; Ohno-Shosaku, T.; Hashimotodani, Y.; Uchigashima, M.; Watanabe, M. Endocannabinoid-mediated control of synaptic transmission. Physiol. Rev. 2009, 89, 309–380. [Google Scholar] [CrossRef]
- Cardinal, P.; Bellocchio, L.; Guzmán-Quevedo, O.; André, C.; Clark, S.; Elie, M.; Leste-Lasserre, T.; Gonzales, D.; Cannich, A.; Marsicano, G.; et al. Cannabinoid type 1 (CB1) receptors on Sim1-expressing neurons regulate energy expenditure in male mice. Endocrinology 2015, 156, 411–418. [Google Scholar] [CrossRef]
- Mazier, W.; Saucisse, N.; Simon, V.; Cannich, A.; Marsicano, G.; Massa, F.; Cota, D. mTORC1 and CB1 receptor signaling regulate excitatory glutamatergic inputs onto the hypothalamic paraventricular nucleus in response to energy availability. Mol. Metab. 2019, 28, 151–159. [Google Scholar] [CrossRef]
- Cruz-Martínez, A.M.; Tejas-Juárez, J.G.; Mancilla-Díaz, J.M.; Florán-Garduño, B.; López-Alonso, V.E.; Escartín-Pérez, R.E. CB1 receptors in the paraventricular nucleus of the hypothalamus modulate the release of 5-HT and GABA to stimulate food intake in rats. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2018, 28, 1247–1259. [Google Scholar] [CrossRef]
- Petrie, G.N.; Balsevich, G.; Füzesi, T.; Aukema, R.J.; Driever, W.P.F.; van der Stelt, M.; Bains, J.S.; Hill, M.N. Disruption of tonic endocannabinoid signalling triggers cellular, behavioural and neuroendocrine responses consistent with a stress response. Br. J. Pharmacol. 2023, 180, 3146–3159. [Google Scholar] [CrossRef] [PubMed]
- Gorzalka, B.B.; Hill, M.N. Integration of endocannabinoid signaling into the neural network regulating stress-induced activation of the hypothalamic-pituitary-adrenal axis. Curr. Top. Behav. Neurosci. 2009, 1, 289–306. [Google Scholar] [PubMed]
- Tanimura, A.; Yamazaki, M.; Hashimotodani, Y.; Uchigashima, M.; Kawata, S.; Abe, M.; Kita, Y.; Hashimoto, K.; Shimizu, T.; Watanabe, M.; et al. The endocannabinoid 2-arachidonoylglycerol produced by diacylglycerol lipase alpha mediates retrograde suppression of synaptic transmission. Neuron 2010, 65, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, M.; Araque, A. Endocannabinoids Mediate Neuron-Astrocyte Communication. Neuron 2008, 57, 883–893. [Google Scholar] [CrossRef]
- Navarrete, M.; Araque, A. Endocannabinoids Potentiate Synaptic Transmission through Stimulation of Astrocytes. Neuron 2010, 68, 113–126. [Google Scholar] [CrossRef]
- Ivell, R.; Richter, D. Structure and comparison of the oxytocin and vasopressin genes from rat. Proc. Natl. Acad. Sci. USA 1984, 81, 2006–2010. [Google Scholar] [CrossRef]
- Kim, D.W.; Washington, P.W.; Wang, Z.Q.; Lin, S.H.; Sun, C.; Ismail, B.T.; Wang, H.; Jiang, L.; Blackshaw, S. The cellular and molecular landscape of hypothalamic patterning and differentiation from embryonic to late postnatal development. Nat. Commun. 2020, 11, 4360. [Google Scholar] [CrossRef]
- Munro, S.; Thomas, K.L.; Abu-Shaar, M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993, 365, 61–65. [Google Scholar] [CrossRef]
- Lauckner, J.E.; Jensen, J.B.; Chen, H.-Y.; Lu, H.-C.; Hille, B.; Mackie, K. GPR55 is a cannabinoid receptor that increases intracellular calcium and inhibits M current. Proc. Natl. Acad. Sci. USA 2008, 105, 2699–2704. [Google Scholar] [CrossRef] [PubMed]
- Grabon, W.; Ruiz, A.; Gasmi, N.; Degletagne, C.; Georges, B.; Belmeguenai, A.; Bodennec, J.; Rheims, S.; Marcy, G.; Bezin, L. CB2 expression in mouse brain: From mapping to regulation in microglia under inflammatory conditions. J. Neuroinflamm. 2024, 21, 206. [Google Scholar] [CrossRef] [PubMed]
- Cherif, H.; Argaw, A.; Cécyre, B.; Bouchard, A.; Gagnon, J.; Javadi, P.; Desgent, S.; Mackie, K.; Bouchard, J.-F. Role of GPR55 during Axon Growth and Target Innervation. eNeuro 2015, 2, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Bisogno, T.; Howell, F.; Williams, G.; Minassi, A.; Cascio, M.G.; Ligresti, A.; Matias, I.; Schiano-Moriello, A.; Paul, P.; Williams, E.-J.; et al. Cloning of the first sn1-DAG lipases points to the spatial and temporal regulation of endocannabinoid signaling in the brain. J. Cell Biol. 2003, 163, 463–468. [Google Scholar] [CrossRef]
- Dinh, T.P.; Carpenter, D.; Leslie, F.M.; Freund, T.F.; Katona, I.; Sensi, S.L.; Kathuria, S.; Piomelli, D. Brain monoglyceride lipase participating in endocannabinoid inactivation. Proc. Natl. Acad. Sci. USA 2002, 99, 10819–10824. [Google Scholar] [CrossRef]
- Oudin, M.J.; Hobbs, C.; Doherty, P. DAGL-dependent endocannabinoid signalling: Roles in axonal pathfinding, synaptic plasticity and adult neurogenesis. Eur. J. Neurosci. 2011, 34, 1634–1646. [Google Scholar] [CrossRef]
- Okamoto, Y.; Morishita, J.; Tsuboi, K.; Tonai, T.; Ueda, N. Molecular Characterization of a Phospholipase D Generating Anandamide and Its Congeners. J. Biol. Chem. 2004, 279, 5298–5305. [Google Scholar] [CrossRef]
- Leishman, E.; Mackie, K.; Luquet, S.; Bradshaw, H.B. Lipidomics profile of a NAPE-PLD KO mouse provides evidence of a broader role of this enzyme in lipid metabolism in the brain. Biochim. Biophys. Acta 2016, 1861, 491–500. [Google Scholar] [CrossRef]
- Simon, G.M.; Cravatt, B.F. Anandamide Biosynthesis Catalyzed by the Phosphodiesterase GDE1 and Detection of Glycerophospho-N-acyl Ethanolamine Precursors in Mouse Brain. J. Biol. Chem. 2008, 283, 9341–9349. [Google Scholar] [CrossRef]
- Deutsch, D.G.; Chin, S.A. Enzymatic synthesis and degradation of anandamide, a cannabinoid receptor agonist. Biochem. Pharmacol. 1993, 46, 791–796. [Google Scholar] [CrossRef]
- Coiret, G.; Ster, J.; Grewe, B.; Wendling, F.; Helmchen, F.; Gerber, U.; Benquet, P. Neuron to astrocyte communication via cannabinoid receptors is necessary for sustained epileptiform activity in rat hippocampus. PLoS ONE 2012, 7, e37320. [Google Scholar] [CrossRef] [PubMed]
- Eraso-Pichot, A.; Pouvreau, S.; Olivera-Pinto, A.; Gomez-Sotres, P.; Skupio, U.; Marsicano, G. Endocannabinoid signaling in astrocytes. Glia 2023, 71, 44–59. [Google Scholar] [CrossRef]
- Hebert-Chatelain, E.; Desprez, T.; Serrat, R.; Bellocchio, L.; Soria-Gomez, E.; Busquets-Garcia, A.; Pagano Zottola, A.C.; Delamarre, A.; Cannich, A.; Vincent, P.; et al. A cannabinoid link between mitochondria and memory. Nature 2016, 539, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Akdemir, E.S.; Huang, A.Y.-S.; Deneen, B. Astrocytogenesis: Where, when, and how. F1000Research 2020, 9, F1000 Faculty Rev–233. [Google Scholar] [CrossRef]
- Rothstein, J.D.; Martin, L.; Levey, A.I.; Dykes-Hoberg, M.; Jin, L.; Wu, D.; Nash, N.; Kuncl, R.W. Localization of neuronal and glial glutamate transporters. Neuron 1994, 13, 713–725. [Google Scholar] [CrossRef]
- Jacque, C.M.; Vinner, C.; Kujas, M.; Raoul, M.; Racadot, J.; Baumann, N.A. Determination of glial fibrillary acidic protein (GFAP) in human brain tumors. J. Neurol. Sci. 1978, 35, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Cahoy, J.D.; Emery, B.; Kaushal, A.; Foo, L.C.; Zamanian, J.L.; Christopherson, K.S.; Xing, Y.; Lubischer, J.L.; Krieg, P.A.; Krupenko, S.A.; et al. A Transcriptome Database for Astrocytes, Neurons, and Oligodendrocytes: A New Resource for Understanding Brain Development and Function. J. Neurosci. 2008, 28, 264–278. [Google Scholar] [CrossRef]
- Bosier, B.; Bellocchio, L.; Metna-Laurent, M.; Soria-Gomez, E.; Matias, I.; Hebert-Chatelain, E.; Cannich, A.; Maitre, M.; Leste-Lasserre, T.; Cardinal, P.; et al. Astroglial CB1 cannabinoid receptors regulate leptin signaling in mouse brain astrocytes. Mol. Metab. 2013, 2, 393–404. [Google Scholar] [CrossRef]
- Jimenez-Blasco, D.; Busquets-Garcia, A.; Hebert-Chatelain, E.; Serrat, R.; Vicente-Gutierrez, C.; Ioannidou, C.; Gómez-Sotres, P.; Lopez-Fabuel, I.; Resch-Beusher, M.; Resel, E.; et al. Glucose metabolism links astroglial mitochondria to cannabinoid effects. Nature 2020, 583, 603–608. [Google Scholar] [CrossRef]
- Dudek, K.A.; Paton, S.E.J.; Binder, L.B.; Collignon, A.; Dion-Albert, L.; Cadoret, A.; Lebel, M.; Lavoie, O.; Bouchard, J.; Kaufmann, F.N.; et al. Astrocytic cannabinoid receptor 1 promotes resilience by dampening stress-induced blood-brain barrier alterations. Nat. Neurosci. 2025, 28, 766–782. [Google Scholar] [CrossRef]
- Moro Chao, D.H.; Kirchner, M.K.; Pham, C.; Foppen, E.; Denis, R.G.; Castel, J.; Morel, C.; Montalban, E.; Hassouna, R.; Bui, L.-C.; et al. Hypothalamic astrocytes control systemic glucose metabolism and energy balance. Cell Metab. 2022, 34, 1532–1547.e6. [Google Scholar] [CrossRef] [PubMed]
- Mulder, J.; Aguado, T.; Keimpema, E.; Barabás, K.; Ballester Rosado, C.J.; Nguyen, L.; Monory, K.; Marsicano, G.; Di Marzo, V.; Hurd, Y.L.; et al. Endocannabinoid signaling controls pyramidal cell specification and long-range axon patterning. Proc. Natl. Acad. Sci. USA 2008, 105, 8760–8765. [Google Scholar] [CrossRef] [PubMed]
- Begbie, J.; Doherty, P.; Graham, A. Cannabinoid receptor, CB1, expression follows neuronal differentiation in the early chick embryo. J. Anat. 2004, 205, 213–218. [Google Scholar] [CrossRef]
- Gao, Y.; Vasilyev, D.V.; Goncalves, M.B.; Howell, F.V.; Hobbs, C.; Reisenberg, M.; Shen, R.; Zhang, M.-Y.; Strassle, B.W.; Lu, P.; et al. Loss of Retrograde Endocannabinoid Signaling and Reduced Adult Neurogenesis in Diacylglycerol Lipase Knock-out Mice. J. Neurosci. 2010, 30, 2017–2024. [Google Scholar] [CrossRef]
- Zaia, C.T.B.V.; Uchôa, E.T.; Santos, A.A.T.D.; Ribeiro, R.C.d.A.; Batista, A.C.S.; Crespigio, J.; Utida, L.; Moura, G.B.; Brownlow, M.L.; Garnica-Siqueira, M.C.; et al. Vasoactive intestinal peptide promotes hypophagia and metabolic changes: Role of paraventricular hypothalamic nucleus and nitric oxide. Brain Res. Bull. 2022, 189, 102–110. [Google Scholar] [CrossRef]
- Abtahi, S.; Mirza, A.; Howell, E.; Currie, P.J. Ghrelin enhances food intake and carbohydrate oxidation in a nitric oxide dependent manner. Gen. Comp. Endocrinol. 2017, 250, 9–14. [Google Scholar] [CrossRef]
- Zhang, K.; Mayhan, W.G.; Patel, K.P. Nitric oxide within the paraventricular nucleus mediates changes in renal sympathetic nerve activity. Am. J. Physiol. 1997, 273, R864–R872. [Google Scholar] [CrossRef] [PubMed]
- McBryde, F.D.; Liu, B.H.; Roloff, E.V.; Kasparov, S.; Paton, J.F.R. Hypothalamic paraventricular nucleus neuronal nitric oxide synthase activity is a major determinant of renal sympathetic discharge in conscious Wistar rats. Exp. Physiol. 2018, 103, 419–428. [Google Scholar] [CrossRef]
- Yamaguchi, N.; Okada, S.; Usui, D.; Yokotani, K. Nitric oxide synthase isozymes in spinally projecting PVN neurons are involved in CRF-induced sympathetic activation. Auton. Neurosci. 2009, 148, 83–89. [Google Scholar] [CrossRef]
- Savinainen, J.R.; Saario, S.M.; Laitinen, J.T. The serine hydrolases MAGL, ABHD6 and ABHD12 as guardians of 2-arachidonoylglycerol signalling through cannabinoid receptors. Acta Physiol. Oxf. Engl. 2012, 204, 267–276. [Google Scholar] [CrossRef]
- Marrs, W.R.; Blankman, J.L.; Horne, E.A.; Thomazeau, A.; Lin, Y.H.; Coy, J.; Bodor, A.L.; Muccioli, G.G.; Hu, S.S.-J.; Woodruff, G.; et al. The serine hydrolase ABHD6 controls the accumulation and efficacy of 2-AG at cannabinoid receptors. Nat. Neurosci. 2010, 13, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Blankman, J.L.; Simon, G.M.; Cravatt, B.F. A Comprehensive Profile of Brain Enzymes that Hydrolyze the Endocannabinoid 2-Arachidonoylglycerol. Chem. Biol. 2007, 14, 1347–1356. [Google Scholar] [CrossRef] [PubMed]
- Maccarrone, M. Metabolism of the Endocannabinoid Anandamide: Open Questions after 25 Years. Front. Mol. Neurosci. 2017, 10, 166. [Google Scholar] [CrossRef] [PubMed]
- Simard, M.; Archambault, A.-S.; Lavoie, J.-P.C.; Dumais, É.; Di Marzo, V.; Flamand, N. Biosynthesis and metabolism of endocannabinoids and their congeners from the monoacylglycerol and N-acyl-ethanolamine families. Biochem. Pharmacol. 2022, 205, 115261. [Google Scholar] [CrossRef]
- Simon, G.M.; Cravatt, B.F. Characterization of mice lacking candidate N-acyl ethanolamine biosynthetic enzymes provides evidence for multiple pathways that contribute to endocannabinoid production in vivo. Mol. Biosyst. 2010, 6, 1411–1418. [Google Scholar] [CrossRef]
- Buczynski, M.W.; Parsons, L.H. Quantification of brain endocannabinoid levels: Methods, interpretations and pitfalls. Br. J. Pharmacol. 2010, 160, 423–442. [Google Scholar] [CrossRef]
- Ohno-Shosaku, T.; Kano, M. Endocannabinoid-mediated retrograde modulation of synaptic transmission. Curr. Opin. Neurobiol. 2014, 29, 1–8. [Google Scholar] [CrossRef]
- Savinainen, J.R.; Järvinen, T.; Laine, K.; Laitinen, J.T. Despite substantial degradation, 2-arachidonoylglycerol is a potent full efficacy agonist mediating CB1 receptor-dependent G-protein activation in rat cerebellar membranes. Br. J. Pharmacol. 2001, 134, 664–672. [Google Scholar] [CrossRef]
- Hill, M.N.; Tasker, J.G. Endocannabinoid Signaling, Glucocorticoid-Mediated Negative Feedback and Regulation of the HPA Axis. Neuroscience 2012, 204, 5–16. [Google Scholar] [CrossRef]
- Chapman, C.D.; Dono, L.M.; French, M.C.; Weinberg, Z.Y.; Schuette, L.M.; Currie, P.J. Paraventricular nucleus anandamide signaling alters eating and substrate oxidation. Neuroreport 2012, 23, 425–429. [Google Scholar] [CrossRef]
- Abruzzese, F.; Greco, M.; Perlino, E.; Doonan, S.; Marra, E. Lack of correlation between mRNA expression and enzymatic activity of the aspartate aminotransferase isoenzymes in various tissues of the rat. FEBS Lett. 1995, 366, 170–172. [Google Scholar] [CrossRef] [PubMed]
- Walter, L.; Dinh, T.; Stella, N. ATP induces a rapid and pronounced increase in 2-arachidonoylglycerol production by astrocytes, a response limited by monoacylglycerol lipase. J. Neurosci. 2004, 24, 8068–8074. [Google Scholar] [CrossRef]
- Virshup, I.; Rybakov, S.; Theis, F.J.; Angerer, P.; Wolf, F.A. anndata: Access and store annotated datamatrices. J. Open Source Softw. 2024, 9, 4371. [Google Scholar] [CrossRef]
- Blischak, J.; Carbonetto, P.; Stephens, M. Creating and Sharing Reproducible Research Code the Workflowr Way. F1000Research 2019, 8, 1749. [Google Scholar] [CrossRef]
- Hao, Y.; Hao, S.; Andersen-Nissen, E.; Mauck, W.M., 3rd; Zheng, S.; Butler, A.; Lee, M.J.; Wilk, A.J.; Darby, C.; Zager, M.; et al. Integrated analysis of multimodal single-cell data. Cell 2021, 184, 3573–3587. [Google Scholar] [CrossRef]
- McInnes, L.; Healy, J.; Saul, N.; Großberger, L. UMAP: Uniform Manifold Approximation and Projection. J. Open Source Softw. 2018, 3, 861. [Google Scholar] [CrossRef]
- Kobak, D.; Linderman, G.C. Initialization is critical for preserving global data structure in both t-SNE and UMAP. Nat. Biotechnol. 2021, 39, 156–157. [Google Scholar] [CrossRef] [PubMed]
- Zupančič, M.; Keimpema, E.; Tretiakov, E.O.; Eder, S.J.; Lev, I.; Englmaier, L.; Bhandari, P.; Fietz, S.A.; Härtig, W.; Renaux, E.; et al. Concerted transcriptional regulation of the morphogenesis of hypothalamic neurons by ONECUT3. Nat. Commun. 2024, 15, 8631. [Google Scholar] [CrossRef]
- Simmons, D.M.; Swanson, L.W. Comparison of the spatial distribution of seven types of neuroendocrine neurons in the rat paraventricular nucleus: Toward a global 3D model. J. Comp. Neurol. 2009, 516, 423–441. [Google Scholar] [CrossRef]
- Romanov, R.A.; Harkany, T. Neuronal heterogeneity in the paraventricular nucleus of the hypothalamus as revealed by single-cell RNA-seq. Curr. Opin. Endocr. Metab. Res. 2022, 25, 100366. [Google Scholar] [CrossRef]
- Keimpema, E.; Di Marzo, V.; Harkany, T. Biological basis of cannabinoid medicines. Science 2021, 374, 1449–1450. [Google Scholar] [CrossRef] [PubMed]
- Maccarrone, M.; Guzmán, M.; Mackie, K.; Doherty, P.; Harkany, T. Programming of neural cells by (endo)cannabinoids: From physiological rules to emerging therapies. Nat. Rev. Neurosci. 2014, 15, 786–801. [Google Scholar] [CrossRef] [PubMed]
- Conway, J.R.; Lex, A.; Gehlenborg, N. UpSetR: An R package for the visualization of intersecting sets and their properties. Bioinformatics 2017, 33, 2938–2940. [Google Scholar] [CrossRef] [PubMed]
- Kanatani, S.; Kreutzmann, J.C.; Li, Y.; West, Z.; Larsen, L.L.; Nikou, D.V.; Eidhof, I.; Walton, A.; Zhang, S.; Rodríguez-Kirby, L.R.; et al. Whole-brain spatial transcriptional analysis at cellular resolution. Science 2024, 386, 907–915. [Google Scholar] [CrossRef]
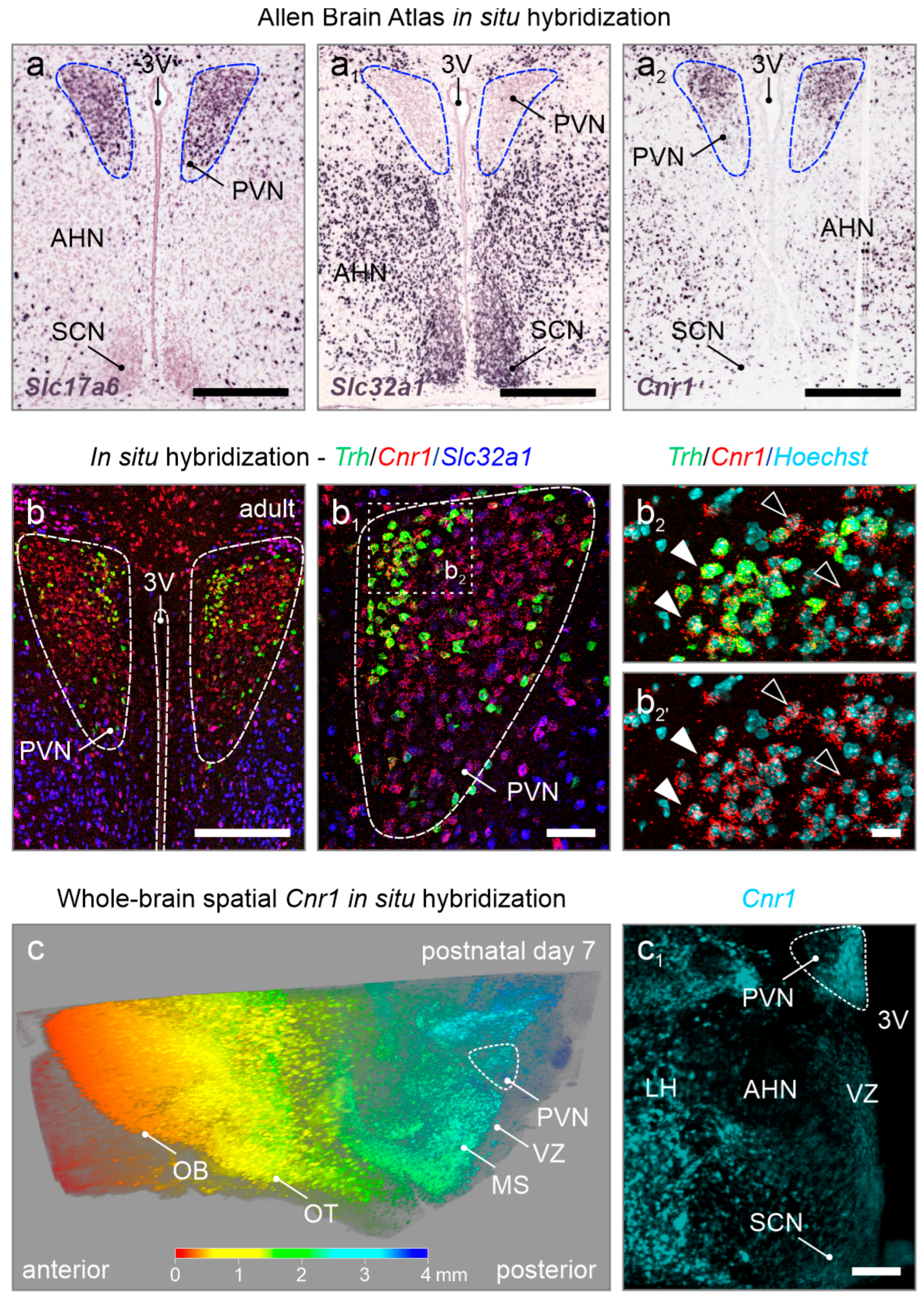

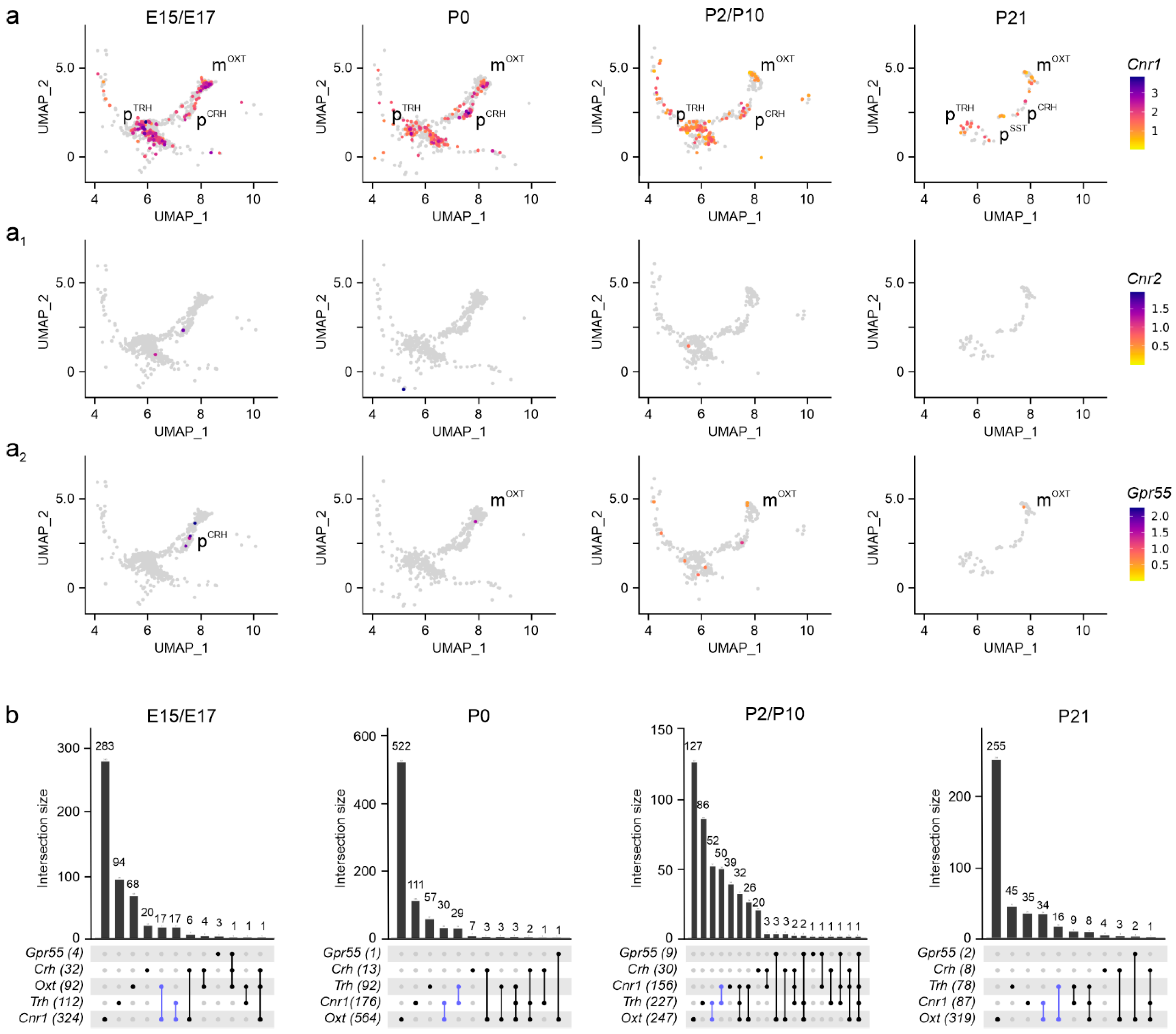
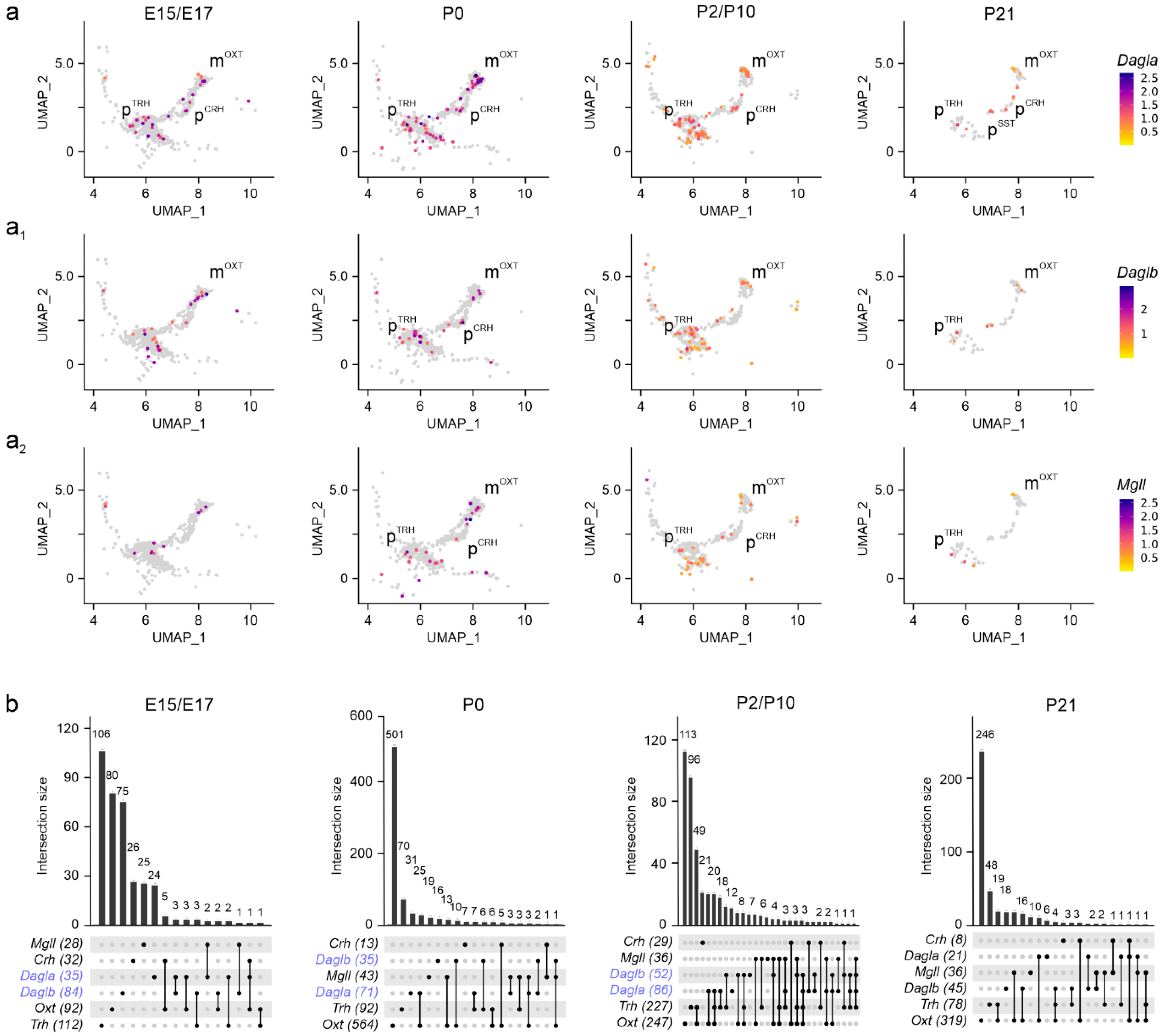
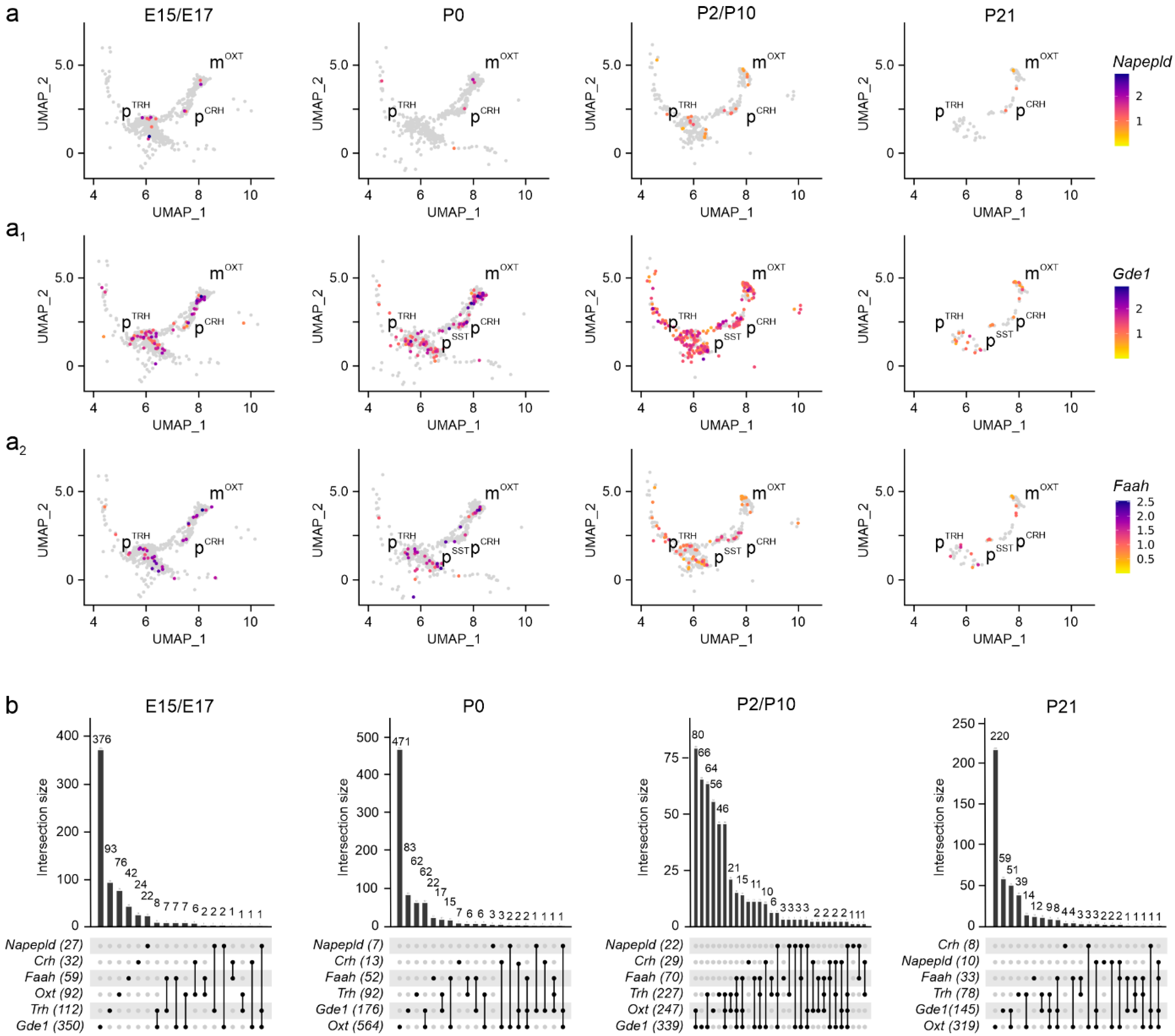

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tretiakov, E.O.; Hevesi, Z.; Böröczky, C.; Alpár, A.; Harkany, T.; Keimpema, E. Molecular Fingerprint of Endocannabinoid Signaling in the Developing Paraventricular Nucleus of the Hypothalamus as Revealed by Single-Cell RNA-Seq and In Situ Hybridization. Cells 2025, 14, 788. https://doi.org/10.3390/cells14110788
Tretiakov EO, Hevesi Z, Böröczky C, Alpár A, Harkany T, Keimpema E. Molecular Fingerprint of Endocannabinoid Signaling in the Developing Paraventricular Nucleus of the Hypothalamus as Revealed by Single-Cell RNA-Seq and In Situ Hybridization. Cells. 2025; 14(11):788. https://doi.org/10.3390/cells14110788
Chicago/Turabian StyleTretiakov, Evgenii O., Zsófia Hevesi, Csenge Böröczky, Alán Alpár, Tibor Harkany, and Erik Keimpema. 2025. "Molecular Fingerprint of Endocannabinoid Signaling in the Developing Paraventricular Nucleus of the Hypothalamus as Revealed by Single-Cell RNA-Seq and In Situ Hybridization" Cells 14, no. 11: 788. https://doi.org/10.3390/cells14110788
APA StyleTretiakov, E. O., Hevesi, Z., Böröczky, C., Alpár, A., Harkany, T., & Keimpema, E. (2025). Molecular Fingerprint of Endocannabinoid Signaling in the Developing Paraventricular Nucleus of the Hypothalamus as Revealed by Single-Cell RNA-Seq and In Situ Hybridization. Cells, 14(11), 788. https://doi.org/10.3390/cells14110788