Phenotypic Heterogeneity of Cancer Associated Fibroblasts in Cervical Cancer Progression: FAP as a Central Activation Marker


 , ,
, ,  ,
,  , and
, and
Abstract

1. Introduction
2. Materials and Methods
2.1. Cell Culture Conditions
2.2. Supernatant Collection of Tumor Cells and Stimulation of MSCs
2.3. Characterization of MSC-CAFs
2.4. Cytokine Profile of MSCs-CAFs
2.5. Cervical Cancer Patients
2.6. Automated Immunohistochemistry
2.7. Statistical Analysis
3. Results
3.1. Expression of CAF-Associated Proteins in MSC Cells Stimulated with Cervical Cancer Cell Supernatants
3.2. Cytokine Production from MSCs Stimulated with Cervical Cancer Cell Supernatants
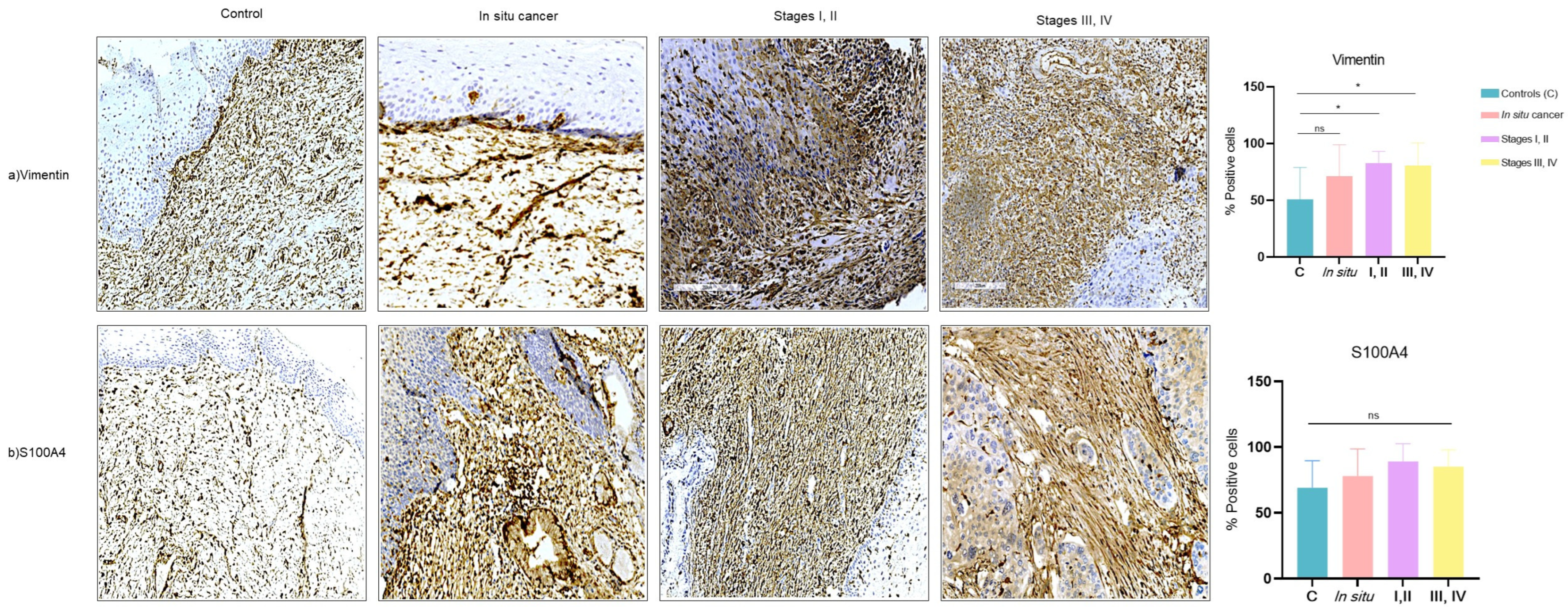
3.3. Expression of CAF-Associated Proteins (Vimentin, S100A4) in Tissues of Cervical Cancer
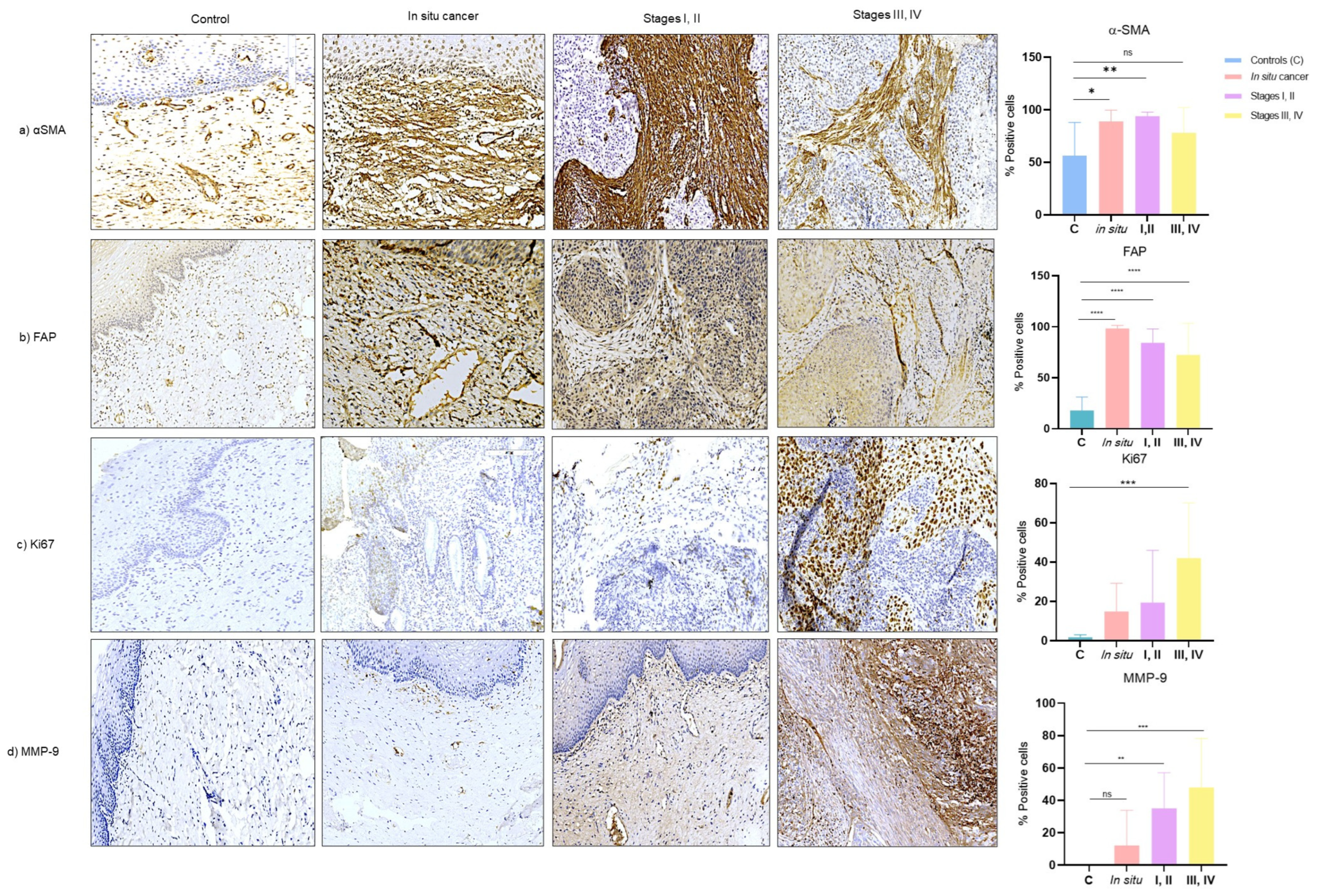
3.4. Expression of CAF-Associated Proteins (αSMA, FAP, Ki67, MMP9) in Tissues of Cervical Cancer
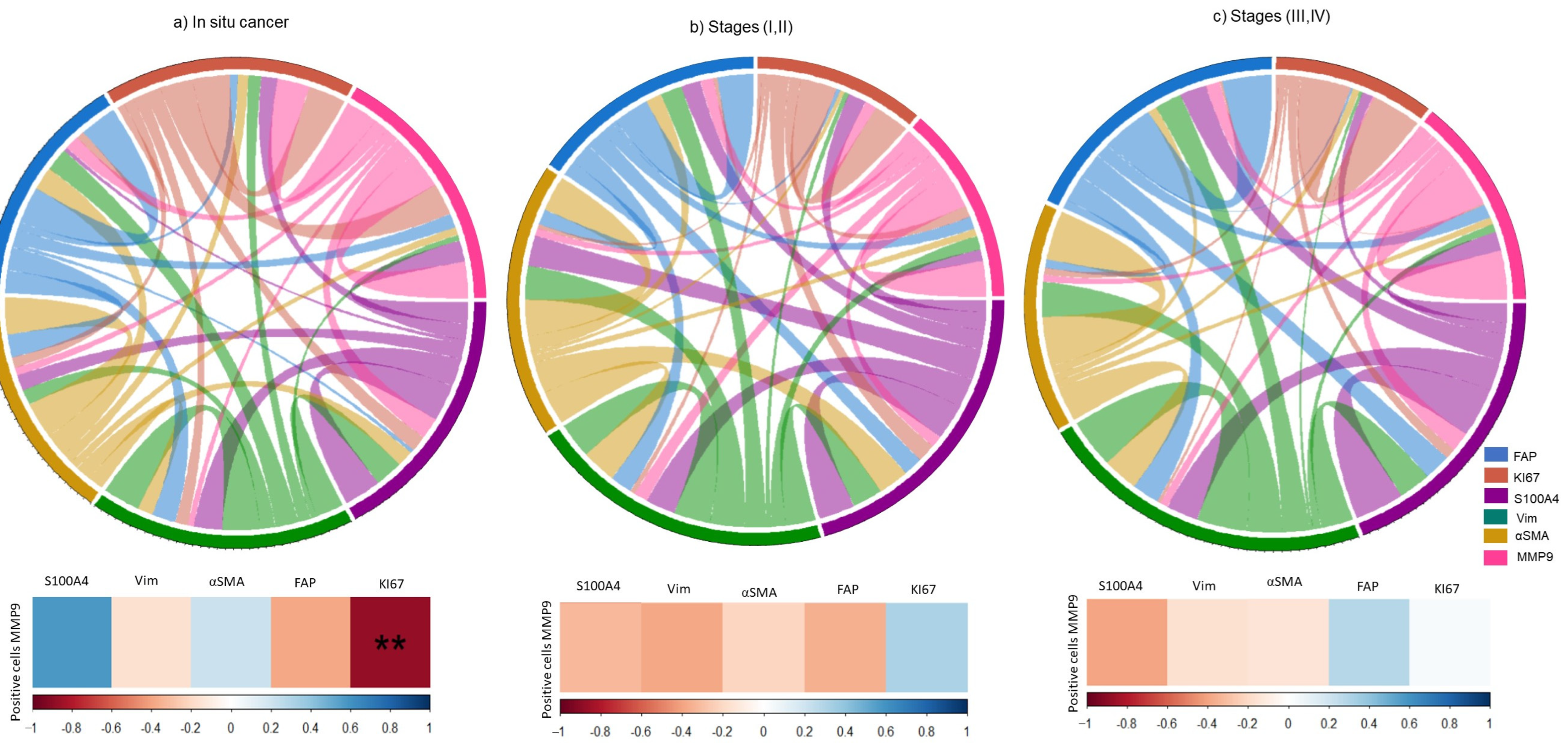
3.5. Association of CAFs related-proteins with Proliferation and Invasion in Tissues of Cervical Cancer
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Bhatla, N.; Aoki, D.; Sharma, D.N.; Sankaranarayanan, R. Cancer of the Cervix Uteri: 2021 Update. Int. J. Gynecol. Obstet. 2021, 155, 28–44. [Google Scholar] [CrossRef]
- Muñoz, N.; Bosch, F.X.; De Sanjosé, S.; Herrero, R.; Castellsagué, X.; Shah, K.V.; Snijders, P.J.F.; Meijer, C.J.L.M. Epidemiologic Classification of Human Papillomavirus Types Associated with Cervical Cancer. N. Engl. J. Med. 2003, 348, 518–527. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Hanahan, D.; Coussens, L.M. Accessories to the Crime: Functions of Cells Recruited to the Tumor Microenvironment. Cancer Cell 2012, 21, 309–322. [Google Scholar] [CrossRef]
- Wright, K.; Ly, T.; Kriet, M.; Czirok, A.; Thomas, S.M. Cancer-Associated Fibroblasts: Master Tumor Microenvironment Modifiers. Cancers 2023, 15, 1899. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Song, E. Turning Foes to Friends: Targeting Cancer-Associated Fibroblasts. Nat. Rev. Drug Discov. 2019, 18, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Hua, J. Interactions between Mesenchymal Stem Cells and the Immune System. Cell. Mol. Life Sci. 2017, 74, 2345–2360. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.-C.; Shyu, W.-C.; Lin, S.-Z. Mesenchymal Stem Cells. Cell Transplant. 2011, 20, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.-X.; Cao, Z.-B.; He, T.-T.; Huang, T.; Xiang, C.-L.; Liu, Y. TGFβ1 Is Essential for MSCs-CAFs Differentiation and Promotes HCT116 Cells Migration and Invasion via JAK/STAT3 Signaling. Onco Targets Ther. 2019, 12, 5323–5334. [Google Scholar] [CrossRef] [PubMed]
- Younesi, F.S.; Son, D.O.; Firmino, J.; Hinz, B. Myofibroblast Markers and Microscopy Detection Methods in Cell Culture and Histology. In Myofibroblasts; Hinz, B., Lagares, D., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2021; Volume 2299, pp. 17–47. ISBN 978-1-07-161381-8. [Google Scholar]
- Nurmik, M.; Ullmann, P.; Rodriguez, F.; Haan, S.; Letellier, E. In Search of Definitions: Cancer-associated Fibroblasts and Their Markers. Int. J. Cancer 2020, 146, 895–905. [Google Scholar] [CrossRef]
- Qi, J.; Sun, H.; Zhang, Y.; Wang, Z.; Xun, Z.; Li, Z.; Ding, X.; Bao, R.; Hong, L.; Jia, W.; et al. Single-Cell and Spatial Analysis Reveal Interaction of FAP+ Fibroblasts and SPP1+ Macrophages in Colorectal Cancer. Nat. Commun. 2022, 13, 1742. [Google Scholar] [CrossRef]
- Scheau, C.; Badarau, I.A.; Costache, R.; Caruntu, C.; Mihai, G.L.; Didilescu, A.C.; Constantin, C.; Neagu, M. The Role of Matrix Metalloproteinases in the Epithelial-Mesenchymal Transition of Hepatocellular Carcinoma. Anal. Cell. Pathol. 2019, 2019, 9423907. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Zhu, H.; Shu, J.; Gong, D.; Zheng, D.; Gao, J. Overexpression of TGF-Β1 and SDF-1 in Cervical Cancer-Associated Fibroblasts Promotes Cell Growth, Invasion and Migration. Arch. Gynecol. Obstet. 2022, 305, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.-J.; Yang, Y.; Wei, W.-F.; Wu, X.-G.; Yan, R.-M.; Zhou, C.-F.; Chen, X.-J.; Wu, S.; Wang, W.; Fan, L.-S. Tumor-Secreted Exosomal Wnt2B Activates Fibroblasts to Promote Cervical Cancer Progression. Oncogenesis 2021, 10, 30. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.L.; Wang, W.; Lan, X.L.; Zeng, Z.C.; Liang, Y.S.; Yan, Y.R.; Song, F.Y.; Wang, F.F.; Zhu, X.H.; Liao, W.J.; et al. CAFs Secreted Exosomes Promote Metastasis and Chemotherapy Resistance by Enhancing Cell Stemness and Epithelial-Mesenchymal Transition in Colorectal Cancer. Mol. Cancer 2019, 18, 91. [Google Scholar] [CrossRef] [PubMed]
- Richards, K.E.; Zeleniak, A.E.; Fishel, M.L.; Wu, J.; Littlepage, L.E.; Hill, R. Cancer-Associated Fibroblast Exosomes Regulate Survival and Proliferation of Pancreatic Cancer Cells. Oncogene 2017, 36, 1770–1778. [Google Scholar] [CrossRef] [PubMed]
- Nagura, M.; Matsumura, N.; Baba, T.; Murakami, R.; Kharma, B.; Hamanishi, J.; Yamaguchi, K.; Abiko, K.; Koshiyama, M.; Mandai, M.; et al. Invasion of Uterine Cervical Squamous Cell Carcinoma Cells Is Facilitated by Locoregional Interaction with Cancer-Associated Fibroblasts via Activating Transforming Growth Factor-Beta. Gynecol. Oncol. 2015, 136, 104–111. [Google Scholar] [CrossRef]
- Mishra, P.J.; Mishra, P.J.; Humeniuk, R.; Medina, D.J.; Alexe, G.; Mesirov, J.P.; Ganesan, S.; Glod, J.W.; Banerjee, D. Carcinoma-Associated Fibroblast–Like Differentiation of Human Mesenchymal Stem Cells. Cancer Res. 2008, 68, 4331–4339. [Google Scholar] [CrossRef]
- Tan, H.-X.; Xiao, Z.-G.; Huang, T.; Fang, Z.-X.; Liu, Y.; Huang, Z.-C. CXCR4/TGF-Β1 Mediated Self-Differentiation of Human Mesenchymal Stem Cells to Carcinoma-Associated Fibroblasts and Promoted Colorectal Carcinoma Development. Cancer Biol. Ther. 2020, 21, 248–257. [Google Scholar] [CrossRef]
- McAndrews, K.M.; Chen, Y.; Darpolor, J.K.; Zheng, X.; Yang, S.; Carstens, J.L.; Li, B.; Wang, H.; Miyake, T.; Correa De Sampaio, P.; et al. Identification of Functional Heterogeneity of Carcinoma-Associated Fibroblasts with Distinct IL6-Mediated Therapy Resistance in Pancreatic Cancer. Cancer Discov. 2022, 12, 1580–1597. [Google Scholar] [CrossRef]
- Carrero, Y.N.; Callejas, D.E.; Mosquera, J.A. In Situ Immunopathological Events in Human Cervical Intraepithelial Neoplasia and Cervical Cancer: Review. Transl. Oncol. 2021, 14, 101058. [Google Scholar] [CrossRef] [PubMed]
- Pedraza-Brindis, E.J.; Sánchez-Reyes, K.; Hernández-Flores, G.; Bravo-Cuellar, A.; Jave-Suárez, L.F.; Aguilar-Lemarroy, A.; Gómez-Lomelí, P.; López-López, B.A.; Ortiz-Lazareno, P.C. Culture Supernatants of Cervical Cancer Cells Induce an M2 Phenotypic Profile in THP-1 Macrophages. Cell. Immunol. 2016, 310, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Tjiong, M.Y.; Van Der Vange, N.; Ter Schegget, J.S.; Burger, M.P.M.; Ten Kate, F.W.J.; Out, T.A. Cytokines in Cervicovaginal Washing Fluid From Patients with Cervical Neoplasia. Cytokine 2001, 14, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Qi, F.; Zhao, F.; Li, G.; Shao, S.; Zhang, X.; Yuan, L.; Feng, Y. Cancer-Associated Fibroblasts Enhance Tumor-Associated Macrophages Enrichment and Suppress NK Cells Function in Colorectal Cancer. Cell Death Dis. 2019, 10, 273. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Sakakura, K.; Kudo, T.; Toyoda, M.; Kaira, K.; Oyama, T.; Chikamatsu, K. Cancer-Associated Fibroblasts Promote an Immunosuppressive Microenvironment through the Induction and Accumulation of Protumoral Macrophages. Oncotarget 2017, 8, 8633–8647. [Google Scholar] [CrossRef]
- Borriello, L.; Nakata, R.; Sheard, M.A.; Fernandez, G.E.; Sposto, R.; Malvar, J.; Blavier, L.; Shimada, H.; Asgharzadeh, S.; Seeger, R.C.; et al. Cancer-Associated Fibroblasts Share Characteristics and Protumorigenic Activity with Mesenchymal Stromal Cells. Cancer Res. 2017, 77, 5142–5157. [Google Scholar] [CrossRef] [PubMed]
- Hellner, K.; Mar, J.; Fang, F.; Quackenbush, J.; Münger, K. HPV16 E7 Oncogene Expression in Normal Human Epithelial Cells Causes Molecular Changes Indicative of an Epithelial to Mesenchymal Transition. Virology 2009, 391, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Watson, R.A.; Thomas, M.; Banks, L.; Roberts, S. Activity of the Human Papillomavirus E6 PDZ-Binding Motif Correlates with an Enhanced Morphological Transformation of Immortalized Human Keratinocytes. J. Cell Sci. 2003, 116, 4925–4934. [Google Scholar] [CrossRef]
- Öhlund, D.; Handly-Santana, A.; Biffi, G.; Elyada, E.; Almeida, A.S.; Ponz-Sarvise, M.; Corbo, V.; Oni, T.E.; Hearn, S.A.; Lee, E.J.; et al. Distinct Populations of Inflammatory Fibroblasts and Myofibroblasts in Pancreatic Cancer. J. Exp. Med. 2017, 214, 579–596. [Google Scholar] [CrossRef]
- Yang, X.; Lin, Y.; Shi, Y.; Li, B.; Liu, W.; Yin, W.; Dang, Y.; Chu, Y.; Fan, J.; He, R. FAP Promotes Immunosuppression by Cancer-Associated Fibroblasts in the Tumor Microenvironment via STAT3–CCL2 Signaling. Cancer Res. 2016, 76, 4124–4135. [Google Scholar] [CrossRef]
- Costa, A.; Kieffer, Y.; Scholer-Dahirel, A.; Pelon, F.; Bourachot, B.; Cardon, M.; Sirven, P.; Magagna, I.; Fuhrmann, L.; Bernard, C.; et al. Fibroblast Heterogeneity and Immunosuppressive Environment in Human Breast Cancer. Cancer Cell 2018, 33, 463–479. [Google Scholar] [CrossRef]
- Pelon, F.; Bourachot, B.; Kieffer, Y.; Magagna, I.; Mermet-Meillon, F.; Bonnet, I.; Costa, A.; Givel, A.-M.; Attieh, Y.; Barbazan, J.; et al. Cancer-Associated Fibroblast Heterogeneity in Axillary Lymph Nodes Drives Metastases in Breast Cancer through Complementary Mechanisms. Nat. Commun. 2020, 11, 404. [Google Scholar] [CrossRef]
- Shi, J.; Hou, Z.; Yan, J.; Qiu, W.; Liang, L.; Meng, M.; Li, L.; Wang, X.; Xie, Y.; Jiang, L.; et al. The Prognostic Significance of Fibroblast Activation Protein-α in Human Lung Adenocarcinoma. Ann. Transl. Med. 2020, 8, 224. [Google Scholar] [CrossRef]
- Mhawech-Fauceglia, P.; Yan, L.; Sharifian, M.; Ren, X.; Liu, S.; Kim, G.; Gayther, S.A.; Pejovic, T.; Lawrenson, K. Stromal Expression of Fibroblast Activation Protein Alpha (FAP) Predicts Platinum Resistance and Shorter Recurrence in Patients with Epithelial Ovarian Cancer. Cancer Microenviron. 2015, 8, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wu, Q.; Liu, Z.; Luo, X.; Fan, Y.; Liu, Y.; Zhang, Y.; Hua, S.; Fu, Q.; Zhao, M.; et al. Downregulation of FAP Suppresses Cell Proliferation and Metastasis through PTEN/PI3K/AKT and Ras-ERK Signaling in Oral Squamous Cell Carcinoma. Cell Death Dis. 2014, 5, e1155. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.M.; Jung, J.; Aziz, N.; Kissil, J.L.; Puré, E. Targeting Fibroblast Activation Protein Inhibits Tumor Stromagenesis and Growth in Mice. J. Clin. Investig. 2009, 119, 3613–3625. [Google Scholar] [CrossRef] [PubMed]
- Puré, E.; Blomberg, R. Pro-Tumorigenic Roles of Fibroblast Activation Protein in Cancer: Back to the Basics. Oncogene 2018, 37, 4343–4357. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Yang, P.; Sun, Y.; Song, S.; Chen, Z. Fibroblast Activation Protein-Based Theranostics in Pancreatic Cancer. Front. Oncol. 2022, 12, 969731. [Google Scholar] [CrossRef]
- Tulley, S.; Chen, W.-T. Transcriptional Regulation of Seprase in Invasive Melanoma Cells by Transforming Growth Factor-β Signaling. J. Biol. Chem. 2014, 289, 15280–15296. [Google Scholar] [CrossRef]
- Popiel-Kopaczyk, A.; Grzegrzolka, J.; Piotrowska, A.; Olbromski, M.; Smolarz, B.; Romanowicz, H.; Rusak, A.; Mrozowska, M.; Dziegiel, P.; Podhorska-Okolow, M.; et al. The Expression of Testin, Ki-67 and P16 in Cervical Cancer Diagnostics. Curr. Issues Mol. Biol. 2023, 45, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, A.; Chander, R.V.; Srinivasan, C.; Vengadassalapathy, S. Prevalence of Angiogenesis, Proliferation, and Apoptosis Markers of Cervical Cancer and Their Correlation with Clinicopathological Parameters. J. Oncol. 2020, 2020, 8541415. [Google Scholar] [CrossRef] [PubMed]
- Joseph, C.; Alsaleem, M.; Orah, N.; Narasimha, P.L.; Miligy, I.M.; Kurozumi, S.; Ellis, I.O.; Mongan, N.P.; Green, A.R.; Rakha, E.A. Elevated MMP9 Expression in Breast Cancer Is a Predictor of Shorter Patient Survival. Breast Cancer Res. Treat. 2020, 182, 267–282. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Li, W.-J.; Weng, J.-J.; Chen, Z.-J.; Wen, Y.-Y.; Deng, T.; Le, H.-B.; Zhang, Y.-K.; Zhang, B.-J. Cancer-associated Fibroblasts, Matrix Metalloproteinase-9 and Lymphatic Vessel Density Are Associated with Progression from Adenocarcinoma In Situ to Invasive Adenocarcinoma of the Lung. Oncol. Lett. 2020, 20, 130. [Google Scholar] [CrossRef]
- Stuelten, C.H.; Byfield, S.D.; Arany, P.R.; Karpova, T.S.; Stetler-Stevenson, W.G.; Roberts, A.B. Breast Cancer Cells Induce Stromal Fibroblasts to Express MMP-9 via Secretion of TNF-α and TGF-β. J. Cell Sci. 2005, 118, 2143–2153. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Groups | Patients with Confirmed Diagnosis | ||||
|---|---|---|---|---|---|
| n = 10 42–72 years | n = 31 25–85 years | ||||
| Histopathological diagnosis | % | Clinical stage | % | Histological types | % |
| Cystic chronic cervicitis | 7(70%) | In situ cancer | 9 (29%) | Squamous cell carcinoma | 15 (68.18%) |
| Active chronic cervicitis with ulceration | 1 (10%) | Early stages (I, II) | 11 (35.48%) | Adenocarcinoma | 5 (22.72%) |
| Chronic acute cervicitis with ulceration | 1 (10%) | Mixed carcinoma (adenosquamous) | 1 (4.54%) | ||
| Chronic mild cervicitis | 1 (10%) | Late stages (III, IV) | 11 (35.48%) | Neuroendocrine carcinoma | 1 (4.54%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bueno-Urquiza, L.J.; Godínez-Rubí, M.; Villegas-Pineda, J.C.; Vega-Magaña, A.N.; Jave-Suárez, L.F.; Puebla-Mora, A.G.; Aguirre-Sandoval, G.E.; Martínez-Silva, M.G.; Ramírez-de-Arellano, A.; Pereira-Suárez, A.L. Phenotypic Heterogeneity of Cancer Associated Fibroblasts in Cervical Cancer Progression: FAP as a Central Activation Marker. Cells 2024, 13, 560. https://doi.org/10.3390/cells13070560
Bueno-Urquiza LJ, Godínez-Rubí M, Villegas-Pineda JC, Vega-Magaña AN, Jave-Suárez LF, Puebla-Mora AG, Aguirre-Sandoval GE, Martínez-Silva MG, Ramírez-de-Arellano A, Pereira-Suárez AL. Phenotypic Heterogeneity of Cancer Associated Fibroblasts in Cervical Cancer Progression: FAP as a Central Activation Marker. Cells. 2024; 13(7):560. https://doi.org/10.3390/cells13070560
Chicago/Turabian StyleBueno-Urquiza, Lesly Jazmin, Marisol Godínez-Rubí, Julio César Villegas-Pineda, Alejandra Natali Vega-Magaña, Luis Felipe Jave-Suárez, Ana Graciela Puebla-Mora, Gloria Estefanía Aguirre-Sandoval, María Guadalupe Martínez-Silva, Adrián Ramírez-de-Arellano, and Ana Laura Pereira-Suárez. 2024. "Phenotypic Heterogeneity of Cancer Associated Fibroblasts in Cervical Cancer Progression: FAP as a Central Activation Marker" Cells 13, no. 7: 560. https://doi.org/10.3390/cells13070560
APA StyleBueno-Urquiza, L. J., Godínez-Rubí, M., Villegas-Pineda, J. C., Vega-Magaña, A. N., Jave-Suárez, L. F., Puebla-Mora, A. G., Aguirre-Sandoval, G. E., Martínez-Silva, M. G., Ramírez-de-Arellano, A., & Pereira-Suárez, A. L. (2024). Phenotypic Heterogeneity of Cancer Associated Fibroblasts in Cervical Cancer Progression: FAP as a Central Activation Marker. Cells, 13(7), 560. https://doi.org/10.3390/cells13070560