
Innate Lymphoid Cells and Their Role in the Immune Response to Infections

 and
and
Abstract
1. Introduction
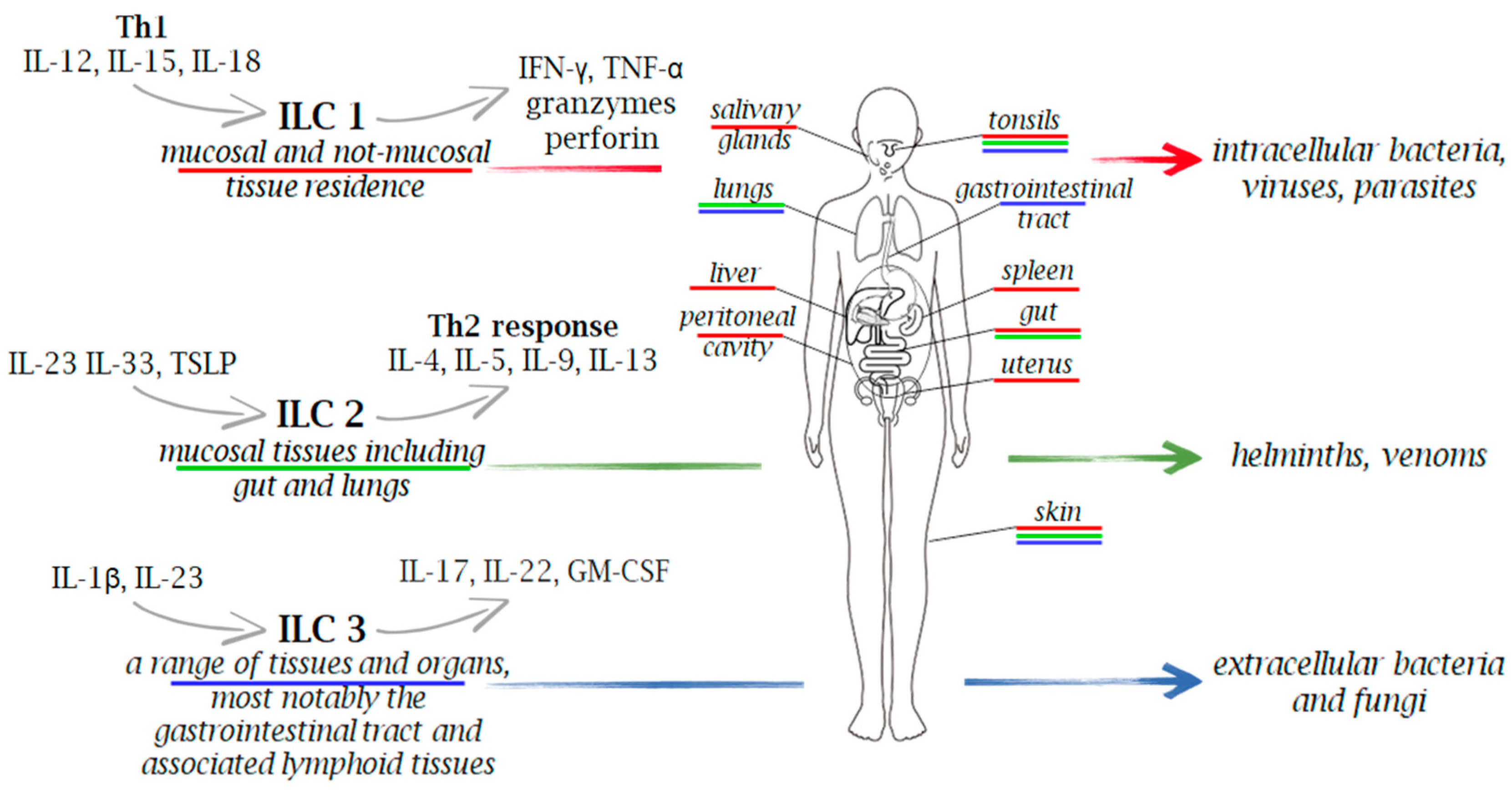
2. Innate Lymphoid Cells (ILCs) Functional Diversity

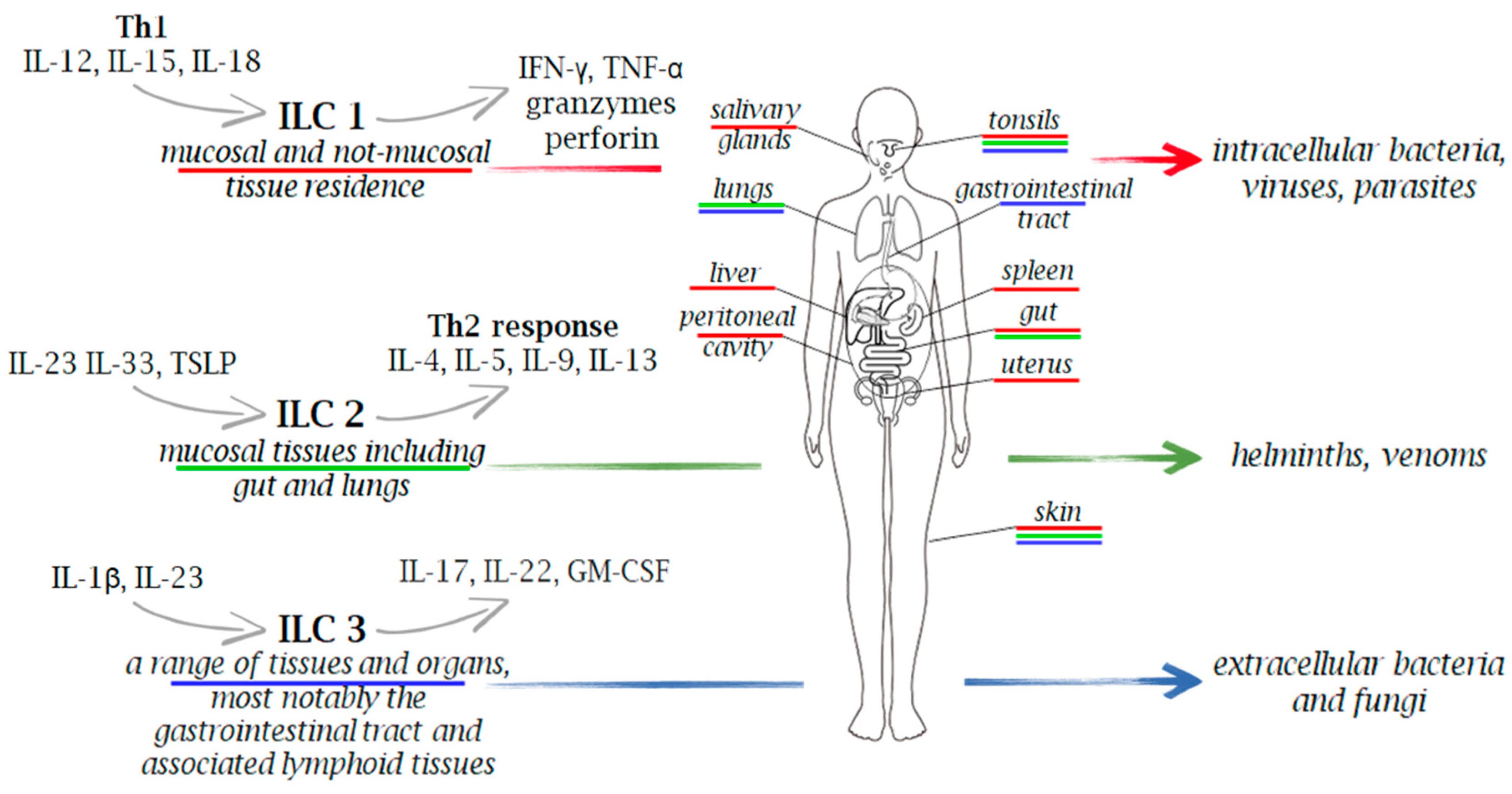{kind=link}
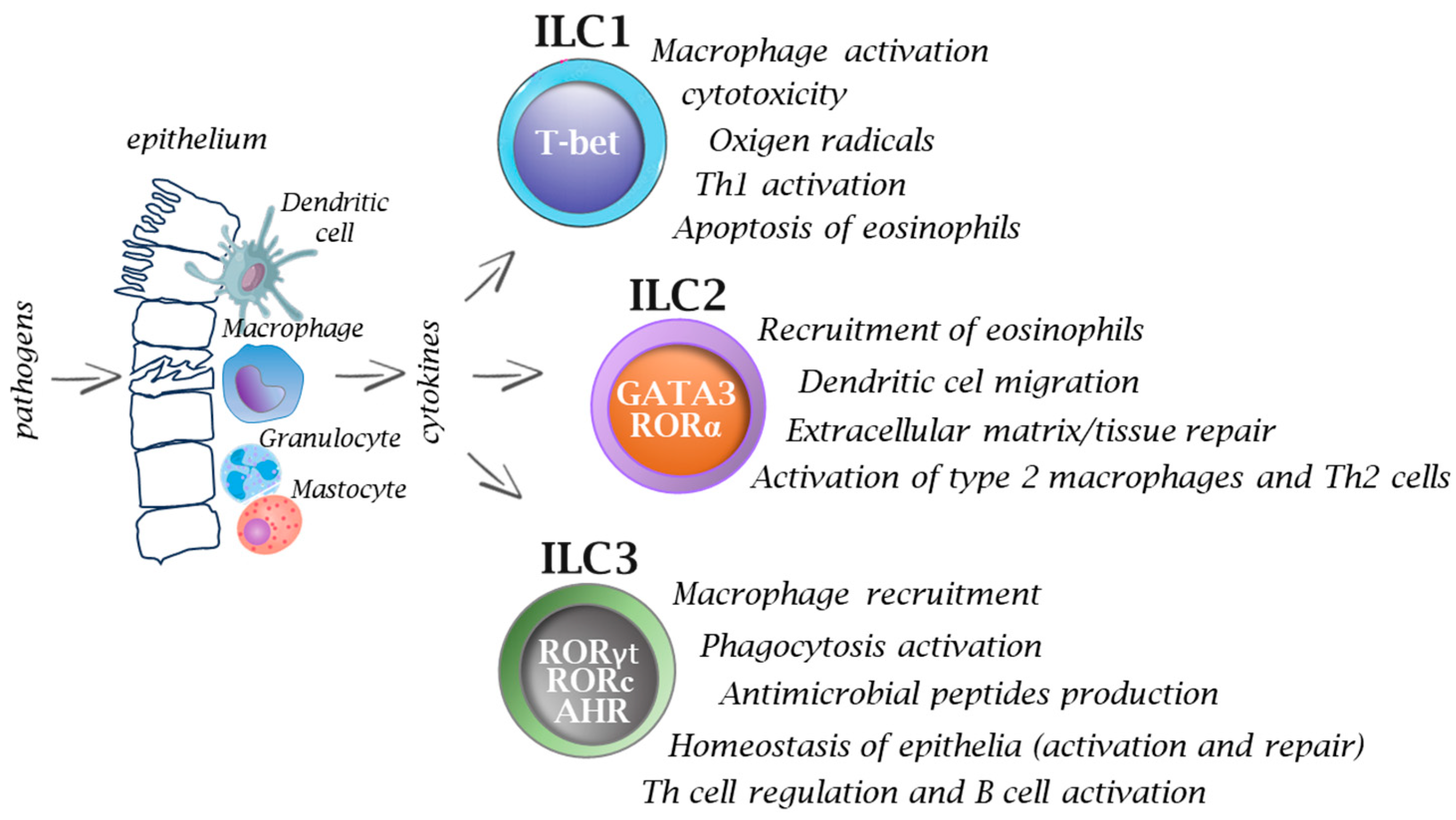{kind=link}
| ILC Subgroup | Phenotypical Markers | Transcription Factors | Cytokines | References |
|---|---|---|---|---|
| NK cells | CD56, NCR1, IL-12Rβ2 | T-bet, Eomes | IFN-γ, TNF-α | [3,5,6,8,11,12,14,15,18,22] |
| ILC1 | CD127,CD161, IL-1R, IL-12Rβ2, ICOS | T-bet, Eomes | IFN-γ, TNF-α | |
| ILC2 | CD127, CD161, ICOS, CRTH2, IL-1R, ST2, IL-17RB | GATA3 | IL-4, IL-5, IL-9, IL-13 | |
| ILC3 | CD117, CD127, CD161, NCR2, ICOS, NCR1, IL-1R, IL-23R | RORγt | IL-17, IL-22, GM-CSF | |
| LTi | CD117, CD127, IL-1R, IL-23R | TCF1 | IL-2, IL-5, IL-13 |
3. ILCs in Immunity to Intracellular and Extracellular Pathogens
3.1. Mycobacterium tuberculosis (M.tb)
3.2. Salmonella Typhimurium
3.3. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2)
3.4. Respiratory Syncytial Virus (RSV)
3.5. Chlamydia sp.
3.6. Toxoplasma gondii
3.7. Streptococcus pneumoniae
3.8. Klebsiella pneumoniae
3.9. Pseudomonas aeruginosa
3.10. Bordetella pertussis
3.11. Clostridium difficile
3.12. Helicobacter pylori
3.13. Candida albicans
4. Therapeutic Potential of ILCs in the Battle against Infections
5. Concluding Remarks
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tsymala, I.; Kuchler, K. Innate Lymphoid Cells—Underexplored Guardians of Immunity. PLoS Pathog. 2023, 19, e1011678. [Google Scholar] [CrossRef]
- Srivastava, R.K.; Sapra, L.; Bhardwaj, A.; Mishra, P.K.; Verma, B.; Baig, Z. Unravelling the Immunobiology of Innate Lymphoid Cells (ILCs): Implications in Health and Disease. Cytokine Growth Factor Rev. 2023, 74, 56–75. [Google Scholar] [CrossRef]
- Eberl, G.; Colonna, M.; Santo, J.P.D.; McKenzie, A.N.J. Innate Lymphoid Cells: A New Paradigm in Immunology. Science 2015, 348, aaa6566. [Google Scholar] [CrossRef]
- Thomas, C.M.; Peebles, R.S. Development and Function of Regulatory Innate Lymphoid Cells. Front. Immunol. 2022, 13, 1014774. [Google Scholar] [CrossRef]
- Jegatheeswaran, S.; Mathews, J.A.; Crome, S.Q. Searching for the Elusive Regulatory Innate Lymphoid Cell. J. Immunol. 2021, 207, 1949–1957. [Google Scholar] [CrossRef] [PubMed]
- Van Maele, L.; Carnoy, C.; Cayet, D.; Ivanov, S.; Porte, R.; Deruy, E.; Chabalgoity, J.A.; Renauld, J.C.; Eberl, G.; Benecke, A.G.; et al. Activation of Type 3 Innate Lymphoid Cells and Interleukin 22 Secretion in the Lungs during Streptococcus Pneumoniae Infection. J. Infect. Dis. 2014, 210, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Artis, D.; Colonna, M.; Diefenbach, A.; Di Santo, J.P.; Eberl, G.; Koyasu, S.; Locksley, R.M.; McKenzie, A.N.J.; Mebius, R.E.; et al. Innate Lymphoid Cells: 10 Years On. Cell 2018, 174, 1054–1066. [Google Scholar] [CrossRef] [PubMed]
- Scoville, S.D.; Mundy-Bosse, B.L.; Zhang, M.H.; Chen, L.; Zhang, X.; Keller, K.A.; Hughes, T.; Chen, L.; Cheng, S.; Bergin, S.M.; et al. A Progenitor Cell Expressing Transcription Factor RORγt Generates All Human Innate Lymphoid Cell Subsets. Immunity 2016, 44, 1140. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Cherrier, D.E.; Chea, S.; Vosshenrich, C.; Serafini, N.; Petit, M.; Liu, P.; Golub, R.; Di Santo, J.P. An Id2RFP-Reporter Mouse Redefines Innate Lymphoid Cell Precursor Potentials. Immunity 2019, 50, 1054–1068.e3. [Google Scholar] [CrossRef] [PubMed]
- Scoville, S.D.; Freud, A.G.; Caligiuri, M.A. Cellular Pathways in the Development of Human and Murine Innate Lymphoid Cells. Curr. Opin. Immunol. 2019, 56, 100. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Zhang, J.; Ji, Y.; Sun, H.; Gu, Z.; Sun, Q.; Bai, M.; Gong, J.; Tang, J.; Zhang, Y.; et al. Tissue Signals Imprint Aiolos Expression in ILC2s to Modulate Type 2 Immunity. Mucosal. Immunol. 2021, 14, 1306–1322. [Google Scholar] [CrossRef]
- Koh, J.; Kim, H.Y.; Lee, Y.; Park, I.K.; Kang, C.H.; Kim, Y.T.; Kim, J.E.; Choi, M.; Lee, W.W.; Jeon, Y.K.; et al. IL23-Producing Human Lung Cancer Cells Promote Tumor Growth via Conversion of Innate Lymphoid Cell 1 (ILC1) into ILC3. Clin. Cancer Res. 2019, 25, 4026–4037. [Google Scholar] [CrossRef]
- Bernink, J.H.; Krabbendam, L.; Germar, K.; de Jong, E.; Gronke, K.; Kofoed-Nielsen, M.; Munneke, J.M.; Hazenberg, M.D.; Villaudy, J.; Buskens, C.J.; et al. Interleukin-12 and -23 Control Plasticity of CD127(+) Group 1 and Group 3 Innate Lymphoid Cells in the Intestinal Lamina Propria. Immunity 2015, 43, 146–160. [Google Scholar] [CrossRef]
- Ohne, Y.; Silver, J.S.; Thompson-Snipes, L.A.; Collet, M.A.; Blanck, J.P.; Cantarel, B.L.; Copenhaver, A.M.; Humbles, A.A.; Liu, Y.J. IL-1 Is a Critical Regulator of Group 2 Innate Lymphoid Cell Function and Plasticity. Nat. Immunol. 2016, 17, 646–655. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Souza-Fonseca-Guimaraes, F.; Bald, T.; Ng, S.S.; Young, A.; Ngiow, S.F.; Rautela, J.; Straube, J.; Waddell, N.; Blake, S.J.; et al. Tumor Immunoevasion by the Conversion of Effector NK Cells into Type 1 Innate Lymphoid Cells. Nat. Immunol. 2017, 18, 1004–1015. [Google Scholar] [CrossRef] [PubMed]
- Krabbendam, L.; Bernink, J.H.; Spits, H. Innate Lymphoid Cells: From Helper to Killer. Curr. Opin. Immunol. 2021, 68, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Krabbendam, L.; Heesters, B.A.; Kradolfer, C.M.A.; Spits, H.; Bernink, J.H. Identification of Human Cytotoxic ILC3s. Eur. J. Immunol. 2021, 51, 811–823. [Google Scholar] [CrossRef] [PubMed]
- Rethacker, L.; Boy, M.; Bisio, V.; Roussin, F.; Denizeau, J.; Vincent-Salomon, A.; Borcoman, E.; Sedlik, C.; Piaggio, E.; Toubert, A.; et al. Innate Lymphoid Cells: NK and Cytotoxic ILC3 Subsets Infiltrate Metastatic Breast Cancer Lymph Nodes. Oncoimmunology 2022, 11, 2057396. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.J.; Sciumè, G.; Amarnath, S. Twenty-One Flavors of Type 1 Innate Lymphoid Cells with PD-1 (Programmed Cell Death-1 Receptor) Sprinkles. Discov. Immunol. 2023, 2, kyad003. [Google Scholar] [CrossRef]
- Llaó-Cid, L.; Roessner, P.M.; Chapaprieta, V.; Öztürk, S.; Roider, T.; Bordas, M.; Izcue, A.; Colomer, D.; Dietrich, S.; Stilgenbauer, S.; et al. EOMES Is Essential for Antitumor Activity of CD8+ T Cells in Chronic Lymphocytic Leukemia. Leukemia 2021, 35, 3152–3162. [Google Scholar] [CrossRef] [PubMed]
- Jacquelot, N.; Seillet, C.; Wang, M.; Pizzolla, A.; Liao, Y.; Hediyeh-zadeh, S.; Grisaru-Tal, S.; Louis, C.; Huang, Q.; Schreuder, J.; et al. Blockade of the Co-Inhibitory Molecule PD-1 Unleashes ILC2-Dependent Anti-Tumor Immunity in Melanoma. Nat. Immunol. 2021, 22, 851. [Google Scholar] [CrossRef]
- Oliphant, C.J.; Hwang, Y.Y.; Walker, J.A.; Salimi, M.; Wong, S.H.; Brewer, J.M.; Englezakis, A.; Barlow, J.L.; Hams, E.; Scanlon, S.T.; et al. MHCII-Mediated Dialog between Group 2 Innate Lymphoid Cells and CD4(+) T Cells Potentiates Type 2 Immunity and Promotes Parasitic Helminth Expulsion. Immunity 2014, 41, 283–295. [Google Scholar] [CrossRef]
- Grigg, J.B.; Shanmugavadivu, A.; Regen, T.; Parkhurst, C.N.; Ahmed, A.; Joseph, A.M.; Mazzucco, M.; Gronke, K.; Diefenbach, A.; Eberl, G.; et al. Antigen-Presenting Innate Lymphoid Cells Orchestrate Neuroinflammation. Nature 2021, 600, 707–712. [Google Scholar] [CrossRef]
- Yang, Y.; Li, Y.; Xu, Y.; Zhang, H.; Diao, Y.; Chen, S.; Cui, Y.; Corrigan, C.J.; Wang, W.; Ying, S. Group 2 Innate Lymphoid Cells Can Engulf and Destroy Bacteria. Cell Mol. Immunol. 2021, 18, 2569–2571. [Google Scholar] [CrossRef]
- Wang, S.; Xia, P.; Chen, Y.; Qu, Y.; Xiong, Z.; Ye, B.; Du, Y.; Tian, Y.; Yin, Z.; Xu, Z.; et al. Regulatory Innate Lymphoid Cells Control Innate Intestinal Inflammation. Cell 2017, 171, 201–216.e18. [Google Scholar] [CrossRef] [PubMed]
- Ardain, A.; Domingo-Gonzalez, R.; Das, S.; Kazer, S.W.; Howard, N.C.; Singh, A.; Ahmed, M.; Nhamoyebonde, S.; Rangel-Moreno, J.; Ogongo, P.; et al. Group 3 Innate Lymphoid Cells Mediate Early Protective Immunity against Tuberculosis. Nature 2019, 570, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Mackley, E.C.; Houston, S.; Marriott, C.L.; Halford, E.E.; Lucas, B.; Cerovic, V.; Filbey, K.J.; Maizels, R.M.; Hepworth, M.R.; Sonnenberg, G.F.; et al. CCR7-Dependent Trafficking of RORγ+ ILCs Creates a Unique Microenvironment within Mucosal Draining Lymph Nodes. Nat. Commun. 2015, 6, 5862. [Google Scholar] [CrossRef] [PubMed]
- Schulz-Kuhnt, A.; Greif, V.; Hildner, K.; Knipfer, L.; Döbrönti, M.; Zirlik, S.; Fuchs, F.; Atreya, R.; Zundler, S.; López-Posadas, R.; et al. ILC2 Lung-Homing in Cystic Fibrosis Patients: Functional Involvement of CCR6 and Impact on Respiratory Failure. Front. Immunol. 2020, 11, 691. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, G.; Fan, X.; Dikiy, S.; Lee, S.Y.; Rudensky, A.Y. Tissue Residency of Innate Lymphoid Cells in Lymphoid and Nonlymphoid Organs. Science 2015, 350, 981–985. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Mao, K.; Chen, X.; Sun, M.A.; Kawabe, T.; Li, W.; Usher, N.; Zhu, J.; Urban, J.F.; Paul, W.E.; et al. S1P-Dependent Interorgan Trafficking of Group 2 Innate Lymphoid Cells Supports Host Defense. Science 2018, 359, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Serafini, N.; Jarade, A.; Surace, L.; Goncalves, P.; Sismeiro, O.; Varet, H.; Legendre, R.; Coppee, J.Y.; Disson, O.; Durum, S.K.; et al. Trained ILC3 Responses Promote Intestinal Defense. Science 2022, 375, 859–863. [Google Scholar] [CrossRef] [PubMed]
- Hatfield, J.K.; Brown, M.A. Group 3 Innate Lymphoid Cells Accumulate and Exhibit Disease-Induced Activation in the Meninges in EAE. Cell Immunol. 2015, 297, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Gonzalez, I.; Mathä, L.; Steer, C.A.; Ghaedi, M.; Poon, G.F.T.; Takei, F. Allergen-Experienced Group 2 Innate Lymphoid Cells Acquire Memory-like Properties and Enhance Allergic Lung Inflammation. Immunity 2016, 45, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Peng, H.; Cong, J.; Wang, X.; Lian, Z.; Wei, H.; Sun, R.; Tian, Z. Memory Formation and Long-Term Maintenance of IL-7Rα+ ILC1s via a Lymph Node-Liver Axis. Nat. Commun. 2018, 9, 4854. [Google Scholar] [CrossRef] [PubMed]
- Withers, D.R.; Gaspal, F.M.; Mackley, E.C.; Marriott, C.L.; Ross, E.A.; Desanti, G.E.; Roberts, N.A.; White, A.J.; Flores-Langarica, A.; McConnell, F.M.; et al. Cutting Edge: Lymphoid Tissue Inducer Cells Maintain Memory CD4 T Cells within Secondary Lymphoid Tissue. J. Immunol. 2012, 189, 2094–2098. [Google Scholar] [CrossRef] [PubMed]
- Paust, S.; Gill, H.S.; Wang, B.Z.; Flynn, M.P.; Moseman, E.A.; Senman, B.; Szczepanik, M.; Telenti, A.; Askenase, P.W.; Compans, R.W.; et al. Critical Role for the Chemokine Receptor CXCR6 in NK Cell-Mediated Antigen-Specific Memory of Haptens and Viruses. Nat. Immunol. 2010, 11, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- de Bree, L.C.J.; Koeken, V.A.C.M.; Joosten, L.A.B.; Aaby, P.; Benn, C.S.; van Crevel, R.; Netea, M.G. Non-Specific Effects of Vaccines: Current Evidence and Potential Implications. Semin. Immunol. 2018, 39, 35–43. [Google Scholar] [CrossRef]
- Balz, K.; Trassl, L.; Härtel, V.; Nelson, P.P.; Skevaki, C. Virus-Induced T Cell-Mediated Heterologous Immunity and Vaccine Development. Front. Immunol. 2020, 11, 513. [Google Scholar] [CrossRef]
- Silva, M.T.; Silva Pestana, N.T. The in Vivo Extracellular Life of Facultative Intracellular Bacterial Parasites: Role in Pathogenesis. Immunobiology 2013, 218, 325–337. [Google Scholar] [CrossRef]
- Lai, D.M.; Shu, Q.; Fan, J. The Origin and Role of Innate Lymphoid Cells in the Lung. Mil. Med. Res. 2016, 3, 25. [Google Scholar] [CrossRef]
- Gasteiger, G.; D’osualdo, A.; Schubert, D.A.; Weber, A.; Bruscia, E.M.; Hartl, D. Cellular Innate Immunity: An Old Game with New Players. J. Innate Immun. 2017, 9, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Zook, E.C.; Kee, B.L. Development of Innate Lymphoid Cells. Nat. Immunol. 2016, 17, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, B. The Development and Diversity of ILCs, NK Cells and Their Relevance in Health and Diseases. Adv. Exp. Med. Biol. 2017, 1024, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Schaerli, P.; Willimann, K.; Lang, A.B.; Lipp, M.; Loetscher, P.; Moser, B. CXC Chemokine Receptor 5 Expression Defines Follicular Homing T Cells with B Cell Helper Function. J. Exp. Med. 2000, 192, 1553–1562. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.W.; Hill, D.G.; Jones, S.A. Understanding Immune Cells in Tertiary Lymphoid Organ Development: It Is All Starting to Come Together. Front. Immunol. 2016, 7, 401. [Google Scholar] [CrossRef] [PubMed]
- Corral, D.; Charton, A.; Krauss, M.Z.; Blanquart, E.; Levillain, F.; Lefrançais, E.; Sneperger, T.; Vahlas, Z.; Girard, J.P.; Eberl, G.; et al. ILC Precursors Differentiate into Metabolically Distinct ILC1-like Cells during Mycobacterium Tuberculosis Infection. Cell Rep. 2022, 39, 110715. [Google Scholar] [CrossRef]
- Tripathi, D.; Radhakrishnan, R.K.; Thandi, R.S.; Paidipally, P.; Devalraju, K.P.; Neela, V.S.K.; McAllister, M.K.; Samten, B.; Valluri, V.L.; Vankayalapati, R. IL-22 Produced by Type 3 Innate Lymphoid Cells (ILC3s) Reduces the Mortality of Type 2 Diabetes Mellitus (T2DM) Mice Infected with Mycobacterium Tuberculosis. PLoS Pathog. 2019, 15, e1008140. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.M.; Stavropoulos, E.; Redford, P.S.; Beebe, A.M.; Bancroft, G.J.; Young, D.B.; O’Garra, A. Blockade of IL-10 Signaling during Bacillus Calmette-Guérin Vaccination Enhances and Sustains Th1, Th17, and Innate Lymphoid IFN-γ and IL-17 Responses and Increases Protection to Mycobacterium Tuberculosis Infection. J. Immunol. 2012, 189, 4079–4087. [Google Scholar] [CrossRef]
- Khader, S.A.; Bell, G.K.; Pearl, J.E.; Fountain, J.J.; Rangel-Moreno, J.; Cilley, G.E.; Shen, F.; Eaton, S.M.; Gaffen, S.L.; Swain, S.L.; et al. IL-23 and IL-17 in the Establishment of Protective Pulmonary CD4+ T Cell Responses after Vaccination and during Mycobacterium Tuberculosis Challenge. Nat. Immunol. 2007, 8, 369–377. [Google Scholar] [CrossRef]
- Cowan, J.; Pandey, S.; Filion, L.G.; Angel, J.B.; Kumar, A.; Cameron, D.W. Comparison of Interferon-γ-, Interleukin (IL)-17- and IL-22-Expressing CD4 T Cells, IL-22-Expressing Granulocytes and Proinflammatory Cytokines during Latent and Active Tuberculosis Infection. Clin. Exp. Immunol. 2012, 167, 317. [Google Scholar] [CrossRef]
- Khader, S.A.; Pearl, J.E.; Sakamoto, K.; Gilmartin, L.; Bell, G.K.; Jelley-Gibbs, D.M.; Ghilardi, N.; deSauvage, F.; Cooper, A.M. IL-23 Compensates for the Absence of IL-12p70 and Is Essential for the IL-17 Response during Tuberculosis but Is Dispensable for Protection and Antigen-Specific IFN-Gamma Responses If IL-12p70 Is Available. J. Immunol. 2005, 175, 788–795. [Google Scholar] [CrossRef] [PubMed]
- De Melo, M.G.M.; Mesquita, E.D.D.; Oliveira, M.M.; Da Silva-Monteiro, C.; Silveira, A.K.A.; Malaquias, T.S.; Dutra, T.C.P.; Galliez, R.M.; Kritski, A.L.; Silva, E.C. Imbalance of NET and Alpha-1-Antitrypsin in Tuberculosis Patients Is Related With Hyper Inflammation and Severe Lung Tissue Damage. Front. Immunol. 2019, 9, 3147. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Maniakis-Grivas, G.; Singh, U.K.; Asher, R.M.; Mauri, F.; Elkington, P.T.; Friedland, J.S. Interleukin-17 Regulates Matrix Metalloproteinase Activity in Human Pulmonary Tuberculosis. J. Pathol. 2018, 244, 311. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Chen, X.; Liu, X.; Qiu, W.; Liu, Y.; Jiang, W.; Zheng, Y.; Mou, Y.; Xu, W.; Li, X.; et al. Innate Lymphoid Cells Exhibited IL-17-Expressing Phenotype in Active Tuberculosis Disease. BMC Pulm. Med. 2021, 21, 318. [Google Scholar] [CrossRef] [PubMed]
- Fàbrega, A.; Vila, J. Salmonella Enterica Serovar Typhimurium Skills to Succeed in the Host: Virulence and Regulation. Clin. Microbiol. Rev. 2013, 26, 308–341. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Chávez, F.; Bäumler, A.J. The Pyromaniac Inside You: Salmonella Metabolism in the Host Gut. Annu. Rev. Microbiol. 2015, 69, 31–48. [Google Scholar] [CrossRef]
- Zhang, S.; Kingsley, R.A.; Santos, R.L.; Andrews-Polymenis, H.; Raffatellu, M.; Figueiredo, J.; Nunes, J.; Tsolis, R.M.; Adams, L.G.; Bäumler, A.J. Molecular Pathogenesis of Salmonella Enterica Serotype Typhimurium-Induced Diarrhea. Infect. Immun. 2003, 71, 1–12. [Google Scholar] [CrossRef]
- Stecher, B.; Hapfelmeier, S.; Müller, C.; Kremer, M.; Stallmach, T.; Hardt, W.D. Flagella and Chemotaxis Are Required for Efficient Induction of Salmonella Enterica Serovar Typhimurium Colitis in Streptomycin-Pretreated Mice. Infect. Immun. 2004, 72, 4138. [Google Scholar] [CrossRef]
- Galán, J.E. Salmonella Typhimurium and Inflammation: A Pathogen-Centric Affair. Nat. Rev. Microbiol. 2021, 19, 716. [Google Scholar] [CrossRef] [PubMed]
- Lapaque, N.; Walzer, T.; Méresse, S.; Vivier, E.; Trowsdale, J. Interactions between Human NK Cells and Macrophages in Response to Salmonella Infection. J. Immunol. 2009, 182, 4339–4348. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Yu, J.; Hu, B.; Lu, C.; Liu, X.; Gao, X.; Li, W.; Zhou, L.; Wang, J.; Wang, D.; et al. Runx3 Mediates Resistance to Intracellular Bacterial Infection by Promoting IL12 Signaling in Group 1 ILC and NCR+ILC3. Front. Immunol. 2018, 9, 2101. [Google Scholar] [CrossRef]
- Klose, C.S.N.; Kiss, E.A.; Schwierzeck, V.; Ebert, K.; Hoyler, T.; D’Hargues, Y.; Göppert, N.; Croxford, A.L.; Waisman, A.; Tanriver, Y.; et al. A T-Bet Gradient Controls the Fate and Function of CCR6−RORγt+ Innate Lymphoid Cells. Nature 2013, 494, 261–265. [Google Scholar] [CrossRef]
- Castleman, M.J.; Dillon, S.M.; Purba, C.; Cogswell, A.C.; McCarter, M.; Barker, E.; Wilson, C. Enteric Bacteria Induce IFNγ and Granzyme B from Human Colonic Group 1 Innate Lymphoid Cells. Gut Microbes 2020, 12, 1667723. [Google Scholar] [CrossRef]
- Kästele, V.; Mayer, J.; Lee, E.S.; Papazian, N.; Cole, J.J.; Cerovic, V.; Belz, G.; Tomura, M.; Eberl, G.; Goodyear, C.; et al. Intestinal-Derived ILCs Migrating in Lymph Increase IFNγ Production in Response to Salmonella Typhimurium Infection. Mucosal. Immunol. 2021, 14, 717–727. [Google Scholar] [CrossRef]
- Kulesza, J.; Kulesza, E.; Koziński, P.; Karpik, W.; Broncel, M.; Fol, M. BCG and SARS-CoV-2—What Have We Learned? Vaccines 2022, 10, 1641. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F. Aging, Male Sex, Obesity, and Metabolic Inflammation Create the Perfect Storm for COVID-19. Diabetes 2020, 69, 1857–1863. [Google Scholar] [CrossRef] [PubMed]
- Alkhouli, M.; Nanjundappa, A.; Annie, F.; Bates, M.C.; Bhatt, D.L. Sex Differences in Case Fatality Rate of COVID-19: Insights From a Multinational Registry. Mayo Clin. Proc. 2020, 95, 1613–1620. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, N.J.; Wang, Y.; Manickas-Hill, Z.; Carbone, C.; Dauphin, A.; Boribong, B.P.; Loiselle, M.; Davis, J.; Leonard, M.M.; Kuri-Cervantes, L.; et al. Innate Lymphoid Cells and COVID-19 Severity in SARS-CoV-2 Infection. eLife 2022, 11, 74681. [Google Scholar] [CrossRef] [PubMed]
- Darboe, A.; Nielsen, C.M.; Wolf, A.S.; Wildfire, J.; Danso, E.; Sonko, B.; Bottomley, C.; Moore, S.E.; Riley, E.M.; Goodier, M.R. Age-Related Dynamics of Circulating Innate Lymphoid Cells in an African Population. Front. Immunol. 2020, 11, 594107. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.S.; Artis, D. Group 2 Innate Lymphoid Cells in Health and Disease. Cold Spring Harb. Perspect. Biol. 2015, 7, a016337. [Google Scholar] [CrossRef] [PubMed]
- Lo Presti, E.; De Gaetano, A.; Pioggia, G.; Gangemi, S. Comprehensive Analysis of the ILCs and Unconventional T Cells in Virus Infection: Profiling and Dynamics Associated with COVID-19 Disease for a Future Monitoring System and Therapeutic Opportunities. Cells 2022, 11, 542. [Google Scholar] [CrossRef] [PubMed]
- Kindermann, M.; Knipfer, L.; Obermeyer, S.; Müller, U.; Alber, G.; Bogdan, C.; Schleicher, U.; Neurath, M.F.; Wirtz, S. Group 2 Innate Lymphoid Cells (ILC2) Suppress Beneficial Type 1 Immune Responses During Pulmonary Cryptococcosis. Front. Immunol. 2020, 11, 209. [Google Scholar] [CrossRef] [PubMed]
- Shannon, J.P.; Vrba, S.M.; Reynoso, G.V.; Wynne-Jones, E.; Kamenyeva, O.; Malo, C.S.; Cherry, C.R.; McManus, D.T.; Hickman, H.D. Group 1 Innate Lymphoid-Cell-Derived Interferon-γ Maintains Anti-Viral Vigilance in the Mucosal Epithelium. Immunity 2021, 54, 276–290.e5. [Google Scholar] [CrossRef] [PubMed]
- Vashist, N.; Trittel, S.; Ebensen, T.; Chambers, B.J.; Guzmán, C.A.; Riese, P. Influenza-Activated ILC1s Contribute to Antiviral Immunity Partially Influenced by Differential GITR Expression. Front. Immunol. 2018, 8, 22. [Google Scholar] [CrossRef] [PubMed]
- Monticelli, L.A.; Sonnenberg, G.F.; Abt, M.C.; Alenghat, T.; Ziegler, C.G.K.; Doering, T.A.; Angelosanto, J.M.; Laidlaw, B.J.; Yang, C.Y.; Sathaliyawala, T.; et al. Innate Lymphoid Cells Promote Lung Tissue Homeostasis Following Acute Influenza Virus Infection. Nat. Immunol. 2011, 12, 1045. [Google Scholar] [CrossRef]
- Califano, D.; Furuya, Y.; Roberts, S.; Avram, D.; McKenzie, A.N.J.; Metzger, D.W. IFN-γ Increases Susceptibility to Influenza A Infection through Suppression of Group II Innate Lymphoid Cells. Mucosal. Immunol. 2018, 11, 209–219. [Google Scholar] [CrossRef]
- Gomez-Cadena, A.; Spehner, L.; Kroemer, M.; Ben Khelil, M.; Bouiller, K.; Verdeil, G.; Trabanelli, S.; Borg, C.; Loyon, R.; Jandus, C. Severe COVID-19 Patients Exhibit an ILC2 NKG2D+ Population in Their Impaired ILC Compartment. Cell Mol. Immunol. 2021, 18, 484. [Google Scholar] [CrossRef]
- García, M.; Kokkinou, E.; Carrasco García, A.; Parrot, T.; Palma Medina, L.M.; Maleki, K.T.; Christ, W.; Varnaitė, R.; Filipovic, I.; Ljunggren, H.G.; et al. Innate Lymphoid Cell Composition Associates with COVID-19 Disease Severity. Clin. Transl. Immunol. 2020, 9, e1224. [Google Scholar] [CrossRef]
- Fonseca, W.; Lukacs, N.W.; Elesela, S.; Malinczak, C.A. Role of ILC2 in Viral-Induced Lung Pathogenesis. Front. Immunol. 2021, 12, 675169. [Google Scholar] [CrossRef] [PubMed]
- Jouan, Y.; Guillon, A.; Gonzalez, L.; Perez, Y.; Boisseau, C.; Ehrmann, S.; Ferreira, M.; Daix, T.; Jeannet, R.; François, B.; et al. Phenotypical and Functional Alteration of Unconventional T Cells in Severe COVID-19 Patients. J. Exp. Med. 2020, 217, e20200872. [Google Scholar] [CrossRef] [PubMed]
- McNamara, P.S.; Smyth, R.L. The Pathogenesis of Respiratory Syncytial Virus Disease in Childhood. Br. Med. Bull. 2002, 61, 13–28. [Google Scholar] [CrossRef]
- Russell, C.D.; Unger, S.A.; Walton, M.; Schwarze, J. The Human Immune Response to Respiratory Syncytial Virus Infection. Clin. Microbiol. Rev. 2017, 30, 481–502. [Google Scholar] [CrossRef]
- Bloomfield, A.; DeVincenzo, J.P.; Ambrose, C.S.; Krilov, L.R. RSV and Non-RSV Illness Hospitalization in RSV Immunoprophylaxis Recipients: A Systematic Literature Review. J. Clin. Virol. 2020, 129, 104339. [Google Scholar] [CrossRef] [PubMed]
- Binns, E.; Tuckerman, J.; Licciardi, P.V.; Wurzel, D. Respiratory Syncytial Virus, Recurrent Wheeze and Asthma: A Narrative Review of Pathophysiology, Prevention and Future Directions. J. Paediatr. Child Health 2022, 58, 1741–1746. [Google Scholar] [CrossRef]
- Norlander, A.E.; Peebles, R.S. Innate Type 2 Responses to Respiratory Syncytial Virus Infection. Viruses 2020, 12, 521. [Google Scholar] [CrossRef]
- Rossi, G.A.; Ballarini, S.; Salvati, P.; Sacco, O.; Colin, A.A. Alarmins and Innate Lymphoid Cells 2 Activation: A Common Pathogenetic Link Connecting Respiratory Syncytial Virus Bronchiolitis and Later Wheezing/Asthma? Pediatr. Allergy Immunol. 2022, 33, e13803. [Google Scholar] [CrossRef] [PubMed]
- Vu, L.D.; Siefker, D.; Jones, T.L.; You, D.; Taylor, R.; DeVincenzo, J.; Cormier, S.A. Elevated Levels of Type 2 Respiratory Innate Lymphoid Cells in Human Infants with Severe Respiratory Syncytial Virus Bronchiolitis. Am. J. Respir. Crit. Care Med. 2019, 200, 1414–1423. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Wang, S.; Wang, D.; Wang, J.; Zheng, R.; Jiang, X.; Liu, B. Group 2 Innate Lymphoid Cells Mediate the Activation of CD4+ T Cells and Aggravate Th1/Th2 Imbalance via MHC II Molecules during Respiratory Syncytial Virus Infection. Int. Immunopharmacol. 2022, 113, 109306. [Google Scholar] [CrossRef]
- Elwell, C.; Mirrashidi, K.; Engel, J. Chlamydia Cell Biology and Pathogenesis. Nat. Rev. Microbiol. 2016, 14, 385–400. [Google Scholar] [CrossRef]
- Peng, L.; Chen, J.L.; Wang, D. Progress and Perspectives in Point of Care Testing for Urogenital Chlamydia Trachomatis Infection: A Review. Med. Sci. Monit. 2020, 26, e920873. [Google Scholar] [CrossRef]
- Miyashita, N. Atypical Pneumonia: Pathophysiology, Diagnosis, and Treatment. Respir. Investig. 2022, 60, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Sturd, N.; Rucks, E.A. Chlamydia Trachomatis. Trends Microbiol. 2023, 31, 535–536. [Google Scholar] [CrossRef] [PubMed]
- Saka, H.A.; Damiani, M.T. Editorial: New Insights in Chlamydia: Host Interactions and Pathogenesis. Front. Cell. Infect. Microbiol. 2023, 13, 1251582. [Google Scholar] [CrossRef] [PubMed]
- Turman, B.J.; Darville, T.; O’Connell, C.M. Plasmid-Mediated Virulence in Chlamydia. Front. Cell. Infect. Microbiol. 2023, 13, 1251135. [Google Scholar] [CrossRef]
- He, Y.; Xu, H.; Song, C.; Koprivsek, J.J.; Arulanandam, B.; Yang, H.; Tao, L.; Zhong, G. Adoptive Transfer of Group 3-Like Innate Lymphoid Cells Restores Mouse Colon Resistance to Colonization of a Gamma Interferon-Susceptible Chlamydia Muridarum Mutant. Infect. Immun. 2021, 89, e00533-20. [Google Scholar] [CrossRef] [PubMed]
- Barth, S.; Kirschnek, S.; Ortmann, N.; Tanriver, Y.; Häcker, G. The Reaction of Innate Lymphoid Cells in the Mouse Female Genital Tract to Chlamydial Infection. Infect. Immun. 2021, 89, e0080020. [Google Scholar] [CrossRef] [PubMed]
- Korchagina, A.A.; Koroleva, E.; Tumanov, A.V. Innate Lymphoid Cell Plasticity in Mucosal Infections. Microorganisms 2023, 11, 461. [Google Scholar] [CrossRef]
- Aguirre, A.A.; Longcore, T.; Barbieri, M.; Dabritz, H.; Hill, D.; Klein, P.N.; Lepczyk, C.; Lilly, E.L.; McLeod, R.; Milcarsky, J.; et al. The One Health Approach to Toxoplasmosis: Epidemiology, Control, and Prevention Strategies. Ecohealth 2019, 16, 378–390. [Google Scholar] [CrossRef]
- de Barros, R.A.M.; Torrecilhas, A.C.; Marciano, M.A.M.; Mazuz, M.L.; Pereira-Chioccola, V.L.; Fux, B. Toxoplasmosis in Human and Animals Around the World. Diagnosis and Perspectives in the One Health Approach. Acta Trop. 2022, 231, 106432. [Google Scholar] [CrossRef]
- Al-Malki, E.S. Toxoplasmosis: Stages of the Protozoan Life Cycle and Risk Assessment in Humans and Animals for an Enhanced Awareness and an Improved Socio-Economic Status. Saudi J. Biol. Sci. 2021, 28, 962–969. [Google Scholar] [CrossRef]
- Nayeri, T.; Sarvi, S.; Daryani, A. Toxoplasmosis: Targeting Neurotransmitter Systems in Psychiatric Disorders. Metab. Brain Dis. 2022, 37, 123–146. [Google Scholar] [CrossRef]
- Spits, H.; Bernink, J.H.; Lanier, L. NK Cells and Type 1 Innate Lymphoid Cells: Partners in Host Defense. Nat. Immunol. 2016, 17, 758–764. [Google Scholar] [CrossRef] [PubMed]
- López-Yglesias, A.H.; Burger, E.; Camanzo, E.; Martin, A.T.; Araujo, A.M.; Kwok, S.F.; Yarovinsky, F. T-Bet-Dependent ILC1- and NK Cell-Derived IFN-γ Mediates CDC1-Dependent Host Resistance against Toxoplasma Gondii. PLoS Pathog. 2021, 17, 1008299. [Google Scholar] [CrossRef] [PubMed]
- Snyder, L.M.; Belmares-Ortega, J.; Doherty, C.M.; Denkers, E.Y. Impact of MyD88, Microbiota, and Location on Type 1 and Type 3 Innate Lymphoid Cells during Toxoplasma Gondii Infection. Immunohorizons 2022, 6, 660–670. [Google Scholar] [CrossRef] [PubMed]
- Steffen, J.; Ehrentraut, S.; Bank, U.; Biswas, A.; Figueiredo, C.A.; Hölsken, O.; Düsedau, H.P.; Dovhan, V.; Knop, L.; Thode, J.; et al. Type 1 Innate Lymphoid Cells Regulate the Onset of Toxoplasma Gondii-Induced Neuroinflammation. Cell Rep. 2022, 38, 110564. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Patel, S.; Wang, Q.; Andhey, P.; Zaitsev, K.; Porter, S.; Hershey, M.; Bern, M.; Plougastel-Douglas, B.; Collins, P.; et al. Toxoplasma Gondii Infection Drives Conversion of NK Cells into ILC1-like Cells. eLife 2019, 8, e47605. [Google Scholar] [CrossRef] [PubMed]
- Moro, K.; Yamada, T.; Tanabe, M.; Takeuchi, T.; Ikawa, T.; Kawamoto, H.; Furusawa, J.I.; Ohtani, M.; Fujii, H.; Koyasu, S. Innate Production of TH2 Cytokines by Adipose Tissue-Associated c-Kit+Sca-1+ Lymphoid Cells. Nature 2009, 463, 540–544. [Google Scholar] [CrossRef] [PubMed]
- Fallon, P.G.; Ballantyne, S.J.; Mangan, N.E.; Barlow, J.L.; Dasvarma, A.; Hewett, D.R.; McIlgorm, A.; Jolin, H.E.; McKenzie, A.N.J. Identification of an Interleukin (IL)-25-Dependent Cell Population That Provides IL-4, IL-5, and IL-13 at the Onset of Helminth Expulsion. J. Exp. Med. 2006, 203, 1105–1116. [Google Scholar] [CrossRef]
- Shimokawa, C.; Kanaya, T.; Hachisuka, M.; Ishiwata, K.; Hisaeda, H.; Kurashima, Y.; Kiyono, H.; Yoshimoto, T.; Kaisho, T.; Ohno, H. Mast Cells Are Crucial for Induction of Group 2 Innate Lymphoid Cells and Clearance of Helminth Infections. Immunity 2017, 46, 863–874.e4. [Google Scholar] [CrossRef]
- Mukai, K.; Karasuyama, H.; Kabashima, K.; Kubo, M.; Galli, S.J. Differences in the Importance of Mast Cells, Basophils, IgE, and IgG versus That of CD4+ T Cells and ILC2 Cells in Primary and Secondary Immunity to Strongyloides Venezuelensis. Infect. Immun. 2017, 85, e00053-17. [Google Scholar] [CrossRef]
- Angkasekwinai, P.; Sodthawon, W.; Jeerawattanawart, S.; Hansakon, A.; Pattanapanyasat, K.; Wang, Y.H. ILC2s Activated by IL-25 Promote Antigen-Specific Th2 and Th9 Functions That Contribute to the Control of Trichinella Spiralis Infection. PLoS ONE 2017, 12, e0184684. [Google Scholar] [CrossRef] [PubMed]
- Spencer, S.P.; Wilhelm, C.; Yang, Q.; Hall, J.A.; Bouladoux, N.; Boyd, A.; Nutman, T.B.; Urban, J.F.; Wang, J.; Ramalingam, T.R.; et al. Adaptation of Innate Lymphoid Cells to a Micronutrient Deficiency Promotes Type 2 Barrier Immunity. Science 2014, 343, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Nausch, N.; Appleby, L.J.; Sparks, A.M.; Midzi, N.; Mduluza, T.; Mutapi, F. Group 2 Innate Lymphoid Cell Proportions Are Diminished in Young Helminth Infected Children and Restored by Curative Anti-Helminthic Treatment. PLoS Negl. Trop. Dis. 2015, 9, e0003627. [Google Scholar] [CrossRef]
- Ivanova, D.L.; Denton, S.L.; Fettel, K.D.; Sondgeroth, K.S.; Gutierrez, J.M.; Bangoura, B.; Dunay, I.R.; Gigley, J.P. Innate Lymphoid Cells in Protection, Pathology, and Adaptive Immunity during Apicomplexan Infection. Front. Immunol. 2019, 10, 402816. [Google Scholar] [CrossRef]
- Mills, K.H.G. IL-17 and IL-17-Producing Cells in Protection versus Pathology. Nat. Rev. Immunol. 2023, 23, 38–54. [Google Scholar] [CrossRef] [PubMed]
- Passos, S.T.; Silver, J.S.; O’Hara, A.C.; Sehy, D.; Stumhofer, J.S.; Hunter, C.A. IL-6 Promotes NK Cell Production of IL-17 during Toxoplasmosis. J. Immunol. 2010, 184, 1776–1783. [Google Scholar] [CrossRef]
- Subramanian, K.; Henriques-Normark, B.; Normark, S. Emerging Concepts in the Pathogenesis of the Streptococcus Pneumoniae: From Nasopharyngeal Colonizer to Intracellular Pathogen. Cell Microbiol. 2019, 21, e13077. [Google Scholar] [CrossRef]
- Gray, J.; Oehrle, K.; Worthen, G.; Alenghat, T.; Whitsett, J.; Deshmukh, H. Intestinal Commensal Bacteria Mediate Lung Mucosal Immunity and Promote Resistance of Newborn Mice to Infection. Sci. Transl. Med. 2017, 9, eaaf9412. [Google Scholar] [CrossRef]
- Hassane, M.; Jouan, Y.; Creusat, F.; Soulard, D.; Boisseau, C.; Gonzalez, L.; Patin, E.C.; Heuzé-Vourc’h, N.; Sirard, J.C.; Faveeuw, C.; et al. Interleukin-7 Protects against Bacterial Respiratory Infection by Promoting IL-17A-Producing Innate T-Cell Response. Mucosal. Immunol. 2020, 13, 128–139. [Google Scholar] [CrossRef]
- Saluzzo, S.; Gorki, A.D.; Rana, B.M.J.; Martins, R.; Scanlon, S.; Starkl, P.; Lakovits, K.; Hladik, A.; Korosec, A.; Sharif, O.; et al. First-Breath-Induced Type 2 Pathways Shape the Lung Immune Environment. Cell Rep. 2017, 18, 1893–1905. [Google Scholar] [CrossRef]
- Rocha, V.F.D.; Barbosa, M.S.; Leal, H.F.; Silva, G.E.O.; Sales, N.M.M.D.; de Souza Santos Monteiro, A.; Azevedo, J.; Malheiros, A.R.X.; Ataide, L.A.; Moreira, B.M.; et al. Prolonged Outbreak of Carbapenem and Colistin-Resistant Klebsiella Pneumoniae at a Large Tertiary Hospital in Brazil. Front. Microbiol. 2022, 13, 831770. [Google Scholar] [CrossRef]
- Xiong, H.; Keith, J.W.; Samilo, D.W.; Carter, R.A.; Leiner, I.M.; Pamer, E.G. Innate Lymphocyte/Ly6C(Hi) Monocyte Crosstalk Promotes Klebsiella Pneumoniae Clearance. Cell 2016, 165, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Eddens, T.; Trevejo-Nunez, G.; Way, E.E.; Elsegeiny, W.; Ricks, D.M.; Garg, A.V.; Erb, C.J.; Bo, M.; Wang, T.; et al. IL-17 Receptor Signaling in the Lung Epithelium Is Required for Mucosal Chemokine Gradients and Pulmonary Host Defense against K. Pneumoniae. Cell Host Microbe 2016, 20, 596. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Hatano, S.; Yamada, H.; Iwakura, Y.; Yoshikai, Y. Two Types of Interleukin 17A–Producing Γδ T Cells in Protection Against Pulmonary Infection With Klebsiella Pneumoniae. J. Infect. Dis. 2016, 214, 1752–1761. [Google Scholar] [CrossRef] [PubMed]
- Iwanaga, N.; Sandquist, I.; Wanek, A.; McCombs, J.; Song, K.; Kolls, J.K. Host Immunology and Rational Immunotherapy for Carbapenem-Resistant Klebsiella Pneumoniae Infection. JCI Insight 2020, 5, e135591. [Google Scholar] [CrossRef] [PubMed]
- Diggle, S.P.; Whiteley, M. Microbe Profile: Pseudomonas Aeruginosa: Opportunistic Pathogen and Lab Rat. Microbiology 2020, 166, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Villeret, B.; Ghinnagow, R.; Kheir, S.; Born-Bony, M.; Kolls, J.K.; Garcia-Verdugo, I.; Sallenave, J.M. Pseudomonas Aeruginosa Lung Infection Subverts Lymphocytic Responses through IL-23 and IL-22 Post-Transcriptional Regulation. Int. J. Mol. Sci. 2022, 23, 8427. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Huang, X.; Wang, Q.; Yao, D.; Lu, W. Virulence Factors of Pseudomonas Aeruginosa and Antivirulence Strategies to Combat Its Drug Resistance. Front. Cell. Infect. Microbiol. 2022, 12, 926758. [Google Scholar] [CrossRef]
- Bayes, H.K.; Ritchie, N.D.; Evans, T.J. Interleukin-17 Is Required for Control of Chronic Lung Infection Caused by Pseudomonas Aeruginosa. Infect. Immun. 2016, 84, 3507–3516. [Google Scholar] [CrossRef]
- Santos, D.R.; Nguyen, A.; Stokes, J.; Pavlov, N.; Abramson, T. Inhibition of Innate Lymphoid Cell Activation by Interleukin-23 Secreting Dendritic Cells During Bordetella Pertussis Infections in Mice. J. Immunol. 2017, 198, 131.10. [Google Scholar] [CrossRef]
- Raeven, R.H.M.; Brummelman, J.; Van Der Maas, L.; Tilstra, W.; Pennings, J.L.A.; Han, W.G.H.; Van Els, C.A.C.M.; Van Riet, E.; Kersten, G.F.A.; Metz, B. Immunological Signatures after Bordetella Pertussis Infection Demonstrate Importance of Pulmonary Innate Immune Cells. PLoS ONE 2016, 11, e0164027. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Guzmán, C.A.; Walker, M.J. The Virulence Factors of Bordetella Pertussis: A Matter of Control. FEMS Microbiol. Rev. 2001, 25, 309–333. [Google Scholar] [CrossRef] [PubMed]
- Byrne, P.; McGuirk, P.; Todryk, S.; Mills, K.H.G. Depletion of NK Cells Results in Disseminating Lethal Infection with Bordetella Pertussis Associated with a Reduction of Antigen-Specific Th1 and Enhancement of Th2, but Not Tr1 Cells. Eur. J. Immunol. 2004, 34, 2579–2588. [Google Scholar] [CrossRef] [PubMed]
- Kroes, M.M.; Mariman, R.; Hijdra, D.; Hamstra, H.J.; van Boxtel, K.J.W.M.; van Putten, J.P.M.; de Wit, J.; Pinelli, E. Activation of Human NK Cells by Bordetella Pertussis Requires Inflammasome Activation in Macrophages. Front. Immunol. 2019, 10, 2030. [Google Scholar] [CrossRef] [PubMed]
- Rupnik, M.; Wilcox, M.H.; Gerding, D.N. Clostridium Difficile Infection: New Developments in Epidemiology and Pathogenesis. Nat. Rev. Microbiol. 2009, 7, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Weintraub, A.; Fang, H.; Nord, C.E. Antimicrobial Resistance in Clostridium Difficile. Int. J. Antimicrob. Agents 2009, 34, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Dubberke, E.R.; Reske, K.A.; Srivastava, A.; Sadhu, J.; Gatti, R.; Young, R.M.; Rakes, L.C.; Dieckgraefe, B.; DiPersio, J.; Fraser, V.J. Clostridium Difficile-Associated Disease in Allogeneic Hematopoietic Stem-Cell Transplant Recipients: Risk Associations, Protective Associations, and Outcomes. Clin. Transplant. 2010, 24, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Smits, W.K.; Lyras, D.; Lacy, D.B.; Wilcox, M.H.; Kuijper, E.J. Clostridium Difficile Infection. Nat. Rev. Dis. Primers 2016, 2, 16020. [Google Scholar] [CrossRef]
- Abt, M.C.; Lewis, B.B.; Caballero, S.; Xiong, H.; Carter, R.A.; Susac, B.; Ling, L.; Leiner, I.; Pamer, E.G. Innate Immune Defenses Mediated by Two ILC Subsets Are Critical for Protection against Acute Clostridium Difficile Infection. Cell Host Microbe 2015, 18, 27–37. [Google Scholar] [CrossRef]
- Geiger, T.L.; Abt, M.C.; Gasteiger, G.; Firth, M.A.; O’Connor, M.H.; Geary, C.D.; O’Sullivan, T.E.; van den Brink, M.R.; Pamer, E.G.; Hanash, A.M.; et al. Nfil3 Is Crucial for Development of Innate Lymphoid Cells and Host Protection against Intestinal Pathogens. J. Exp. Med. 2014, 211, 1723–1731. [Google Scholar] [CrossRef]
- Baj, J.; Forma, A.; Sitarz, M.; Portincasa, P.; Garruti, G.; Krasowska, D.; Maciejewski, R. Helicobacter Pylori Virulence Factors-Mechanisms of Bacterial Pathogenicity in the Gastric Microenvironment. Cells 2020, 10, 27. [Google Scholar] [CrossRef]
- Sukri, A.; Hanafiah, A.; Mohamad Zin, N.; Kosai, N.R. Epidemiology and Role of Helicobacter Pylori Virulence Factors in Gastric Cancer Carcinogenesis. APMIS 2020, 128, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Bostick, J.W.; Wang, Y.; Shen, Z.; Ge, Y.; Brown, J.; Chen, Z.; Ming, E.; Mohamadzadeh, M.; Fox, J.G.; Zhou, L. Dichotomous Regulation of Group 3 Innate Lymphoid Cells by Nongastric Helicobacter Species. Proc. Natl. Acad. Sci. USA 2019, 116, 24760–24769. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Jiang, X.X.; Zhang, L.F.; Liu, X.M.; Hu, T.Z.; Xia, X.J.; Li, M.; Xu, C.X. Group 2 Innate Lymphoid Cells Are Involved in Skewed Type 2 Immunity of Gastric Diseases Induced by Helicobacter Pylori Infection. Mediat. Inflamm. 2017, 2017, 4927964. [Google Scholar] [CrossRef] [PubMed]
- Satoh-Takayama, N.; Kato, T.; Motomura, Y.; Kageyama, T.; Taguchi-Atarashi, N.; Kinoshita-Daitoku, R.; Kuroda, E.; Di Santo, J.P.; Mimuro, H.; Moro, K.; et al. Bacteria-Induced Group 2 Innate Lymphoid Cells in the Stomach Provide Immune Protection through Induction of IgA. Immunity 2020, 52, 635–649.e4. [Google Scholar] [CrossRef] [PubMed]
- Ohno, H.; Satoh-Takayama, N. Stomach Microbiota, Helicobacter Pylori, and Group 2 Innate Lymphoid Cells. Exp. Mol. Med. 2020, 52, 1377–1382. [Google Scholar] [CrossRef]
- Sorini, C.; Tripathi, K.P.; Wu, S.; Higdon, S.M.; Wang, J.; Cheng, L.; Banerjee, S.; Reinhardt, A.; Kreslavsky, T.; Thorell, A.; et al. Metagenomic and Single-Cell RNA-Seq Survey of the Helicobacter Pylori-Infected Stomach in Asymptomatic Individuals. JCI Insight 2023, 8, e161042. [Google Scholar] [CrossRef]
- Calderone, R.A.; Fonzi, W.A. Virulence Factors of Candida Albicans. Trends Microbiol. 2001, 9, 327–335. [Google Scholar] [CrossRef]
- Talapko, J.; Juzbašić, M.; Matijević, T.; Pustijanac, E.; Bekić, S.; Kotris, I.; Škrlec, I. Candida Albicans- The Virulence Factors and Clinical Manifestations of Infection. J. Fungi 2021, 7, 79. [Google Scholar] [CrossRef]
- Conti, H.R.; Shen, F.; Nayyar, N.; Stocum, E.; Sun, J.N.; Lindemann, M.J.; Ho, A.W.; Hai, J.H.; Yu, J.J.; Jung, J.W.; et al. Th17 Cells and IL-17 Receptor Signaling Are Essential for Mucosal Host Defense against Oral Candidiasis. J. Exp. Med. 2009, 206, 299–311. [Google Scholar] [CrossRef]
- Schmidt, S.; Tramsen, L.; Rais, B.; Ullrich, E.; Lehrnbecher, T. Natural Killer Cells as a Therapeutic Tool for Infectious Diseases—Current Status and Future Perspectives. Oncotarget 2018, 9, 20891. [Google Scholar] [CrossRef]
- Gladiator, A.; Wangler, N.; Trautwein-Weidner, K.; LeibundGut-Landmann, S. Cutting Edge: IL-17-Secreting Innate Lymphoid Cells Are Essential for Host Defense against Fungal Infection. J. Immunol. 2013, 190, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Sparber, F.; Dolowschiak, T.; Mertens, S.; Lauener, L.; Clausen, B.E.; Joller, N.; Stoitzner, P.; Tussiwand, R.; LeibundGut-Landmann, S. Langerin+ DCs Regulate Innate IL-17 Production in the Oral Mucosa during Candida Albicans-Mediated Infection. PLoS Pathog. 2018, 14, e1007069. [Google Scholar] [CrossRef] [PubMed]
- Conti, H.R.; Peterson, A.C.; Brane, L.; Huppler, A.R.; Herna’ndez-Santos, N.; Whibley, N.; Garg, A.V.; Simpson-Abelson, M.R.; Gibson, G.A.; Mamo, A.J.; et al. Oral-Resident Natural Th17 Cells and Γδ T Cells Control Opportunistic Candida Albicans Infections. J. Exp. Med. 2014, 211, 2075–2084. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, S.; Canaday, L.M.; Ochayon, D.E.; Rangel, K.M.; Ali, A.; Gyurova, I.E.; Krishnamurthy, D.; Fletcher, J.S.; Reighard, S.D.; Cox, A.; et al. The Promise and Peril of Natural Killer Cell Therapies in Pulmonary Infection. Immunity 2020, 52, 887–889. [Google Scholar] [CrossRef] [PubMed]
- Björkström, N.K.; Strunz, B.; Ljunggren, H.G. Natural Killer Cells in Antiviral Immunity. Nat. Rev. Immunol. 2022, 22, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Cobb, L.M.; Verneris, M.R. Therapeutic Manipulation of Innate Lymphoid Cells. JCI Insight 2021, 6, e146006. [Google Scholar] [CrossRef]
- Buonocore, S.; Ahern, P.P.; Uhlig, H.H.; Ivanov, I.I.; Littman, D.R.; Maloy, K.J.; Powrie, F. Innate Lymphoid Cells Drive Interleukin-23-Dependent Innate Intestinal Pathology. Nature 2010, 464, 1371–1375. [Google Scholar] [CrossRef] [PubMed]
- Kaser, A. Not All Monoclonals Are Created Equal—Lessons from Failed Drug Trials in Crohn’s Disease. Best Prac. Res. Clin. Gastroenterol. 2014, 28, 437–449. [Google Scholar] [CrossRef]
- Li, J.; Doty, A.L.; Iqbal, A.; Glover, S.C. The Differential Frequency of Lineage(-)CRTH2(-)CD45(+)NKp44(-)CD117(-)CD127(+)ILC Subset in the Inflamed Terminal Ileum of Patients with Crohn’s Disease. Cell. Immunol. 2016, 304–305, 63–68. [Google Scholar] [CrossRef]
- Chevalier, G.; Laveissière, A.; Desachy, G.; Barnich, N.; Sivignon, A.; Maresca, M.; Nicoletti, C.; Di Pasquale, E.; Martinez-Medina, M.; Simpson, K.W.; et al. Blockage of Bacterial FimH Prevents Mucosal Inflammation Associated with Crohn’s Disease. Microbiome 2021, 9, 176. [Google Scholar] [CrossRef] [PubMed]
- Reinisch, W.; Hébuterne, X.; Buisson, A.; Schreiber, S.; Desreumaux, P.; Primas, C.; Paillarse, J.M.; Chevalier, G.; Bonny, C. Safety, Pharmacokinetic, and Pharmacodynamic Study of Sibofimloc, a Novel FimH Blocker in Patients with Active Crohn’s Disease. J. Gastroenterol. Hepatol. 2022, 37, 832–840. [Google Scholar] [CrossRef]
- Vadstrup, K.; Bendtsen, F. Anti-NKG2D MAb: A New Treatment for Crohn’s Disease? Int. J. Mol. Sci. 2017, 18, 1997. [Google Scholar] [CrossRef] [PubMed]
- Eken, A.; Yetkin, M.F.; Vural, A.; Okus, F.Z.; Erdem, S.; Azizoglu, Z.B.; Haliloglu, Y.; Cakir, M.; Turkoglu, E.M.; Kilic, O.; et al. Fingolimod Alters Tissue Distribution and Cytokine Production of Human and Murine Innate Lymphoid Cells. Front. Immunol. 2019, 10, 217. [Google Scholar] [CrossRef] [PubMed]
- Notarangelo, L.D.; Giliani, S.; Mazza, C.; Mazza, C.; Mella, P.; Savoldi, G.; Rodriguezpérez, C.; Mazzolari, E.; Fiorini, M.; Duse, M.; et al. Of Genes and Phenotypes: The Immunological and Molecular Spectrum of Combined Immune Deficiency. Defects of the Gamma(c)-JAK3 Signaling Pathway as a Model. Immunol. Rev. 2000, 178, 39–48. [Google Scholar] [CrossRef]
- Jost, S.; Altfeld, M. Control of Human Viral Infections by Natural Killer Cells. Annu. Rev. Immunol. 2013, 31, 163–194. [Google Scholar] [CrossRef]
- Park, S.J.; Hughes, M.A.; Burdick, M.; Strieter, R.M.; Mehrad, B. Early NK Cell-Derived Interferon-Gamma Is Essential to Host Defense in Neutropenic Invasive Aspergillosis. J. Immunol. 2009, 182, 4306. [Google Scholar] [CrossRef]
- Viegas, N.; Andzinski, L.; Wu, C.F.; Komoll, R.M.; Gekara, N.; Dittmar, K.E.; Weiss, S.; Jablonska, J. IFN-γ Production by CD27+ NK Cells Exacerbates Listeria Monocytogenes Infection in Mice by Inhibiting Granulocyte Mobilization. Eur. J. Immunol. 2013, 43, 2626–2637. [Google Scholar] [CrossRef]
- Newton, D.W.; Runnels, H.A.; Kearns, R.J. Enhanced Splenic Bacterial Clearance and Neutrophilia in Anti-NK1.1-Treated Mice Infected with Pseudomonas Aeruginosa. Nat. Immun. 1992, 11, 335–344. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fol, M.; Karpik, W.; Zablotni, A.; Kulesza, J.; Kulesza, E.; Godkowicz, M.; Druszczynska, M. Innate Lymphoid Cells and Their Role in the Immune Response to Infections. Cells 2024, 13, 335. https://doi.org/10.3390/cells13040335
Fol M, Karpik W, Zablotni A, Kulesza J, Kulesza E, Godkowicz M, Druszczynska M. Innate Lymphoid Cells and Their Role in the Immune Response to Infections. Cells. 2024; 13(4):335. https://doi.org/10.3390/cells13040335
Chicago/Turabian StyleFol, Marek, Wojciech Karpik, Agnieszka Zablotni, Jakub Kulesza, Ewelina Kulesza, Magdalena Godkowicz, and Magdalena Druszczynska. 2024. "Innate Lymphoid Cells and Their Role in the Immune Response to Infections" Cells 13, no. 4: 335. https://doi.org/10.3390/cells13040335
APA StyleFol, M., Karpik, W., Zablotni, A., Kulesza, J., Kulesza, E., Godkowicz, M., & Druszczynska, M. (2024). Innate Lymphoid Cells and Their Role in the Immune Response to Infections. Cells, 13(4), 335. https://doi.org/10.3390/cells13040335