
A Bird’s-Eye View of Chromosomic Evolution in the Class Aves

 ,
,  ,
,  and
and 
Abstract
1. Avian Biology and Its Importance
2. Avian Evolution
3. Defining the Avian Karyotype
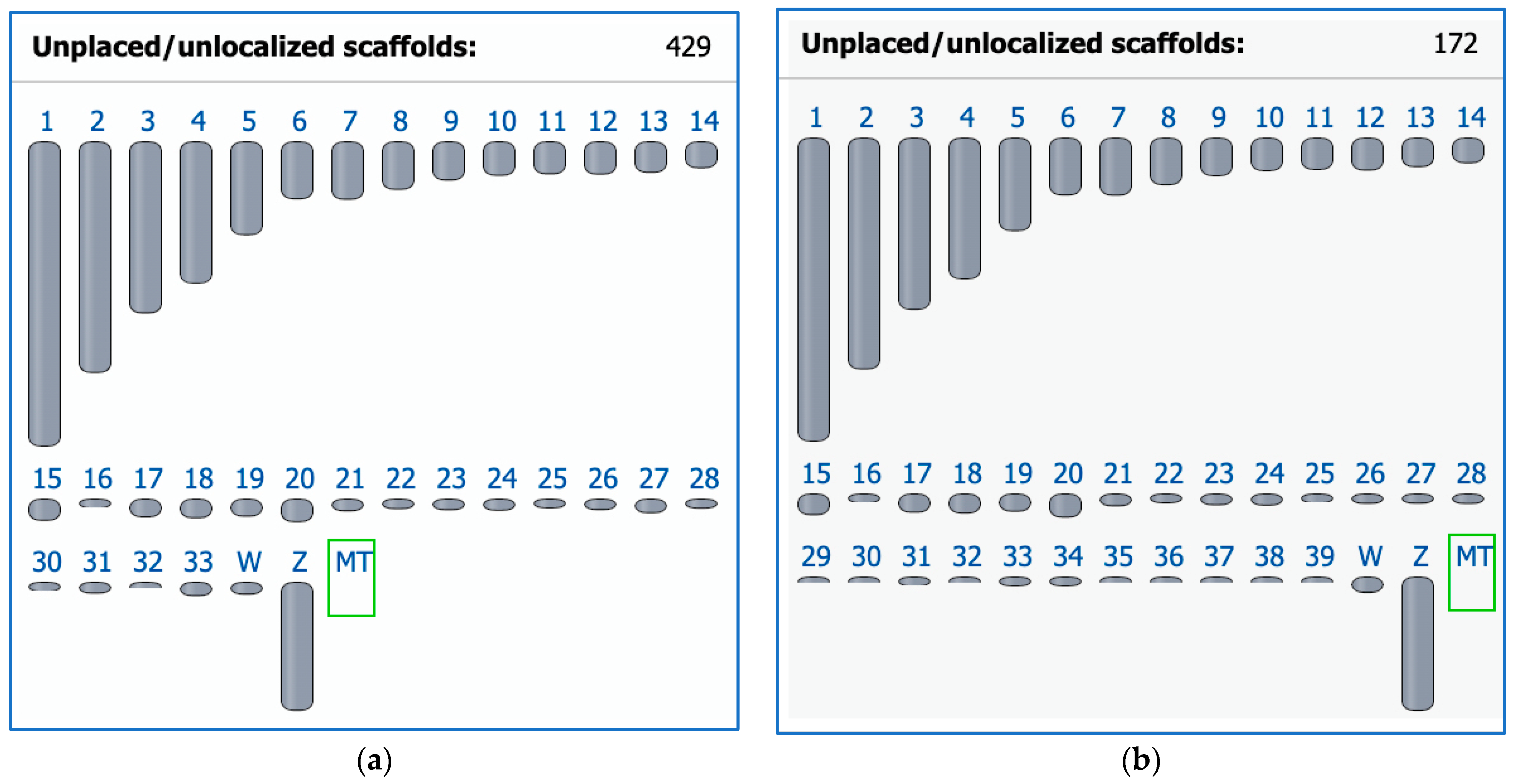
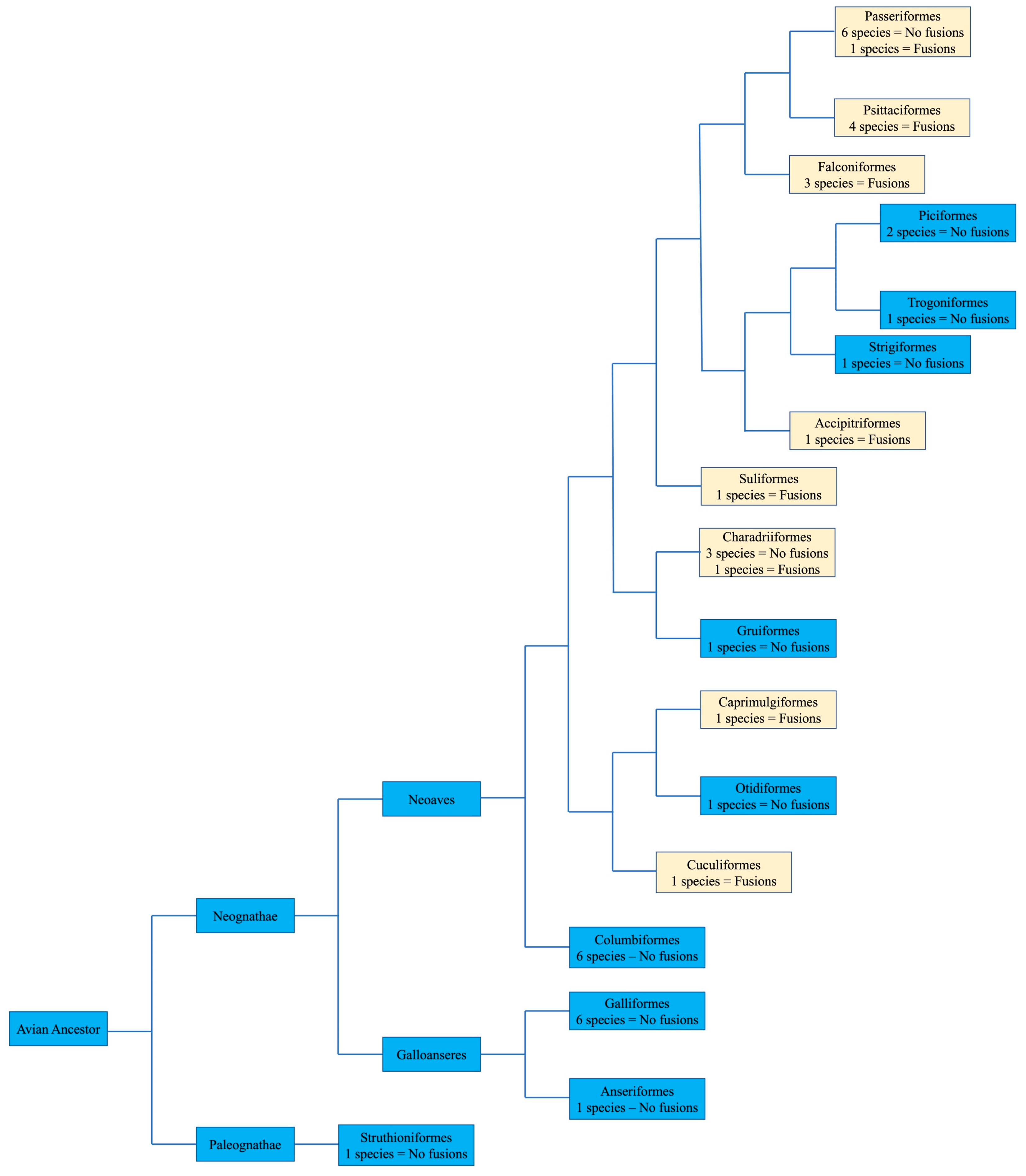
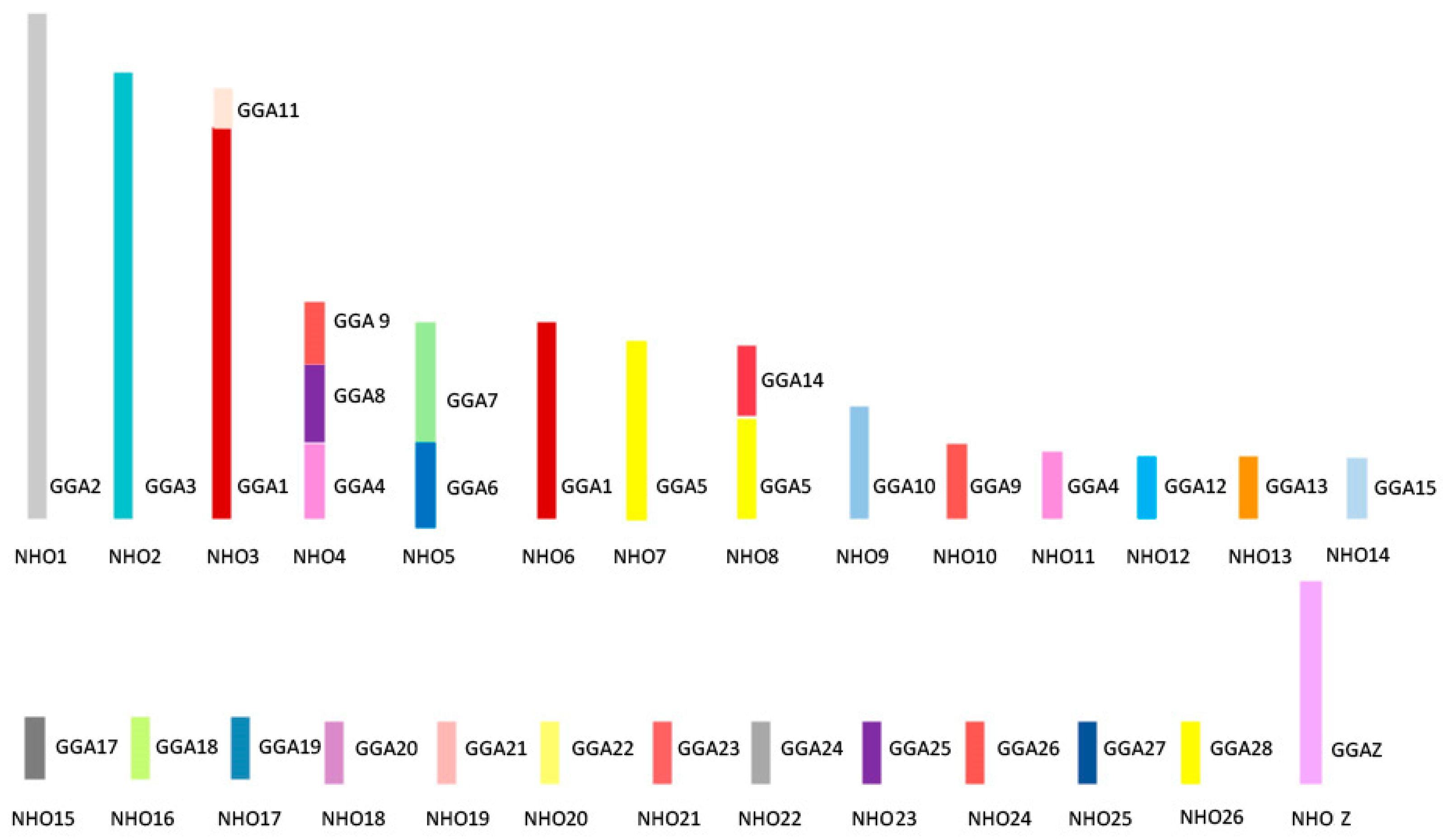
4. Avian Karyotypic Diversity
5. Sex Chromosomes in Birds
6. Chromosomal Rearrangements in Birds
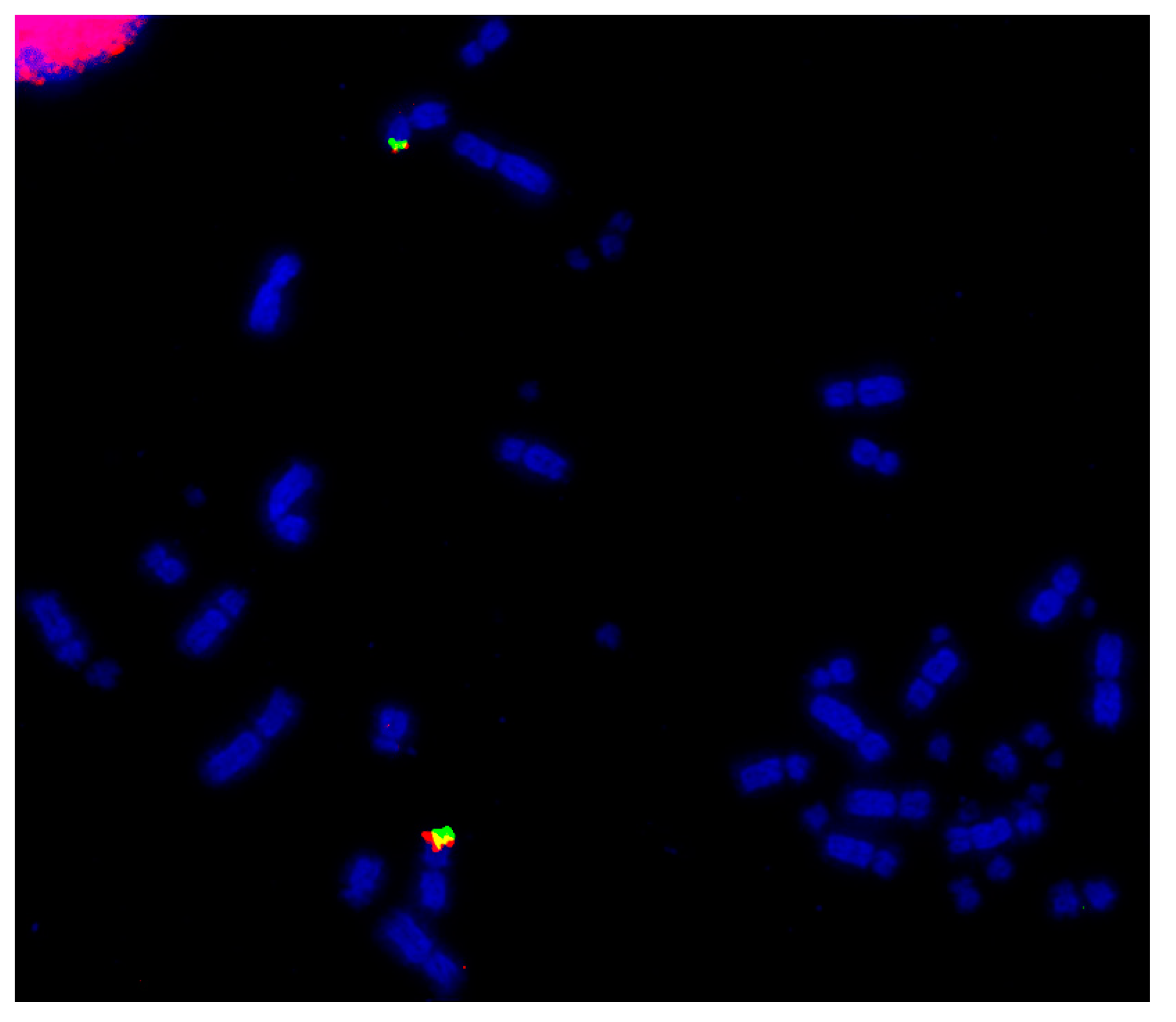
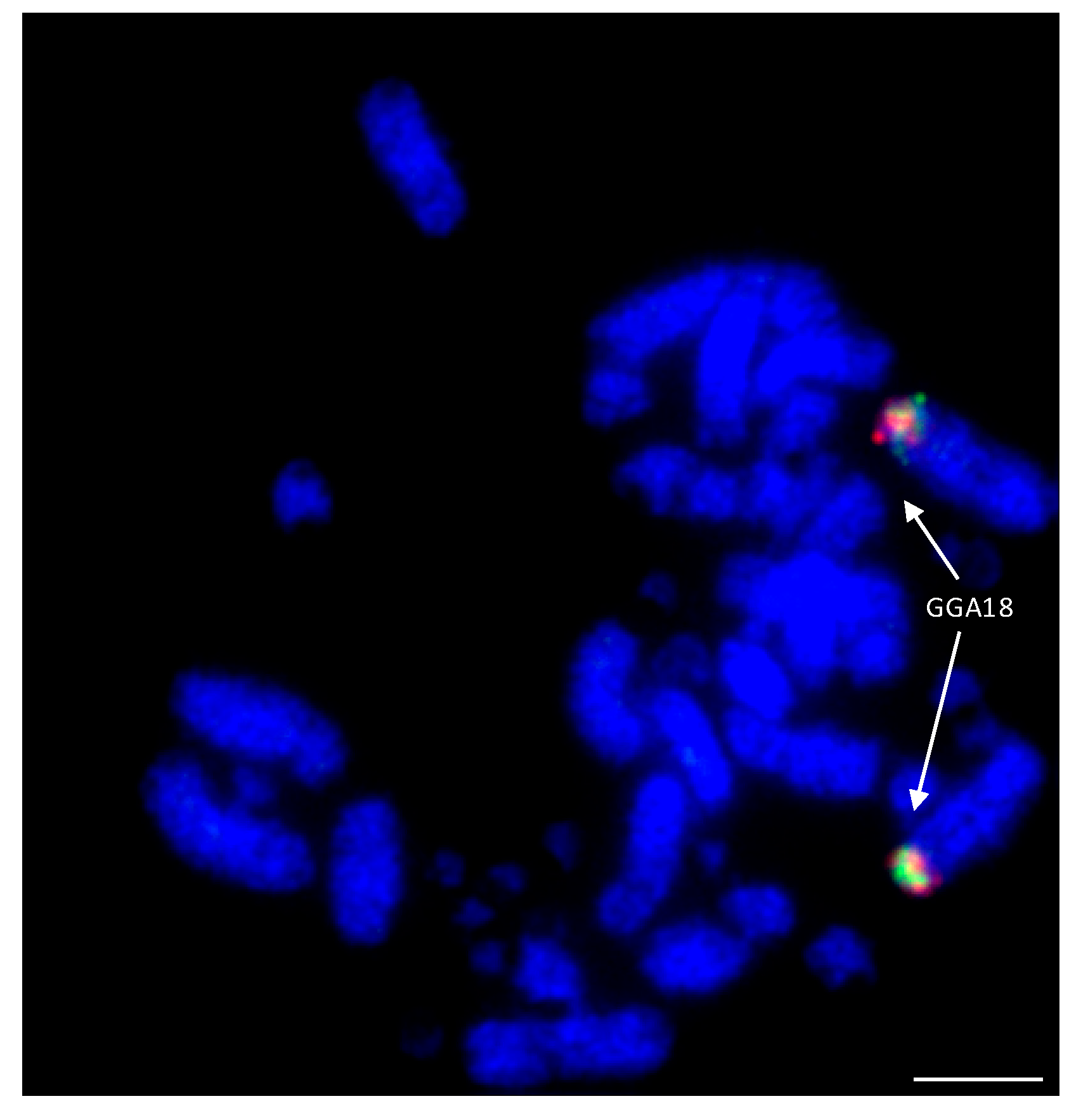
7. Closer Examination of Avian Microchromosomes
7.1. Species with No Apparent Interchromosomal Rearrangement between the Microchromosomes
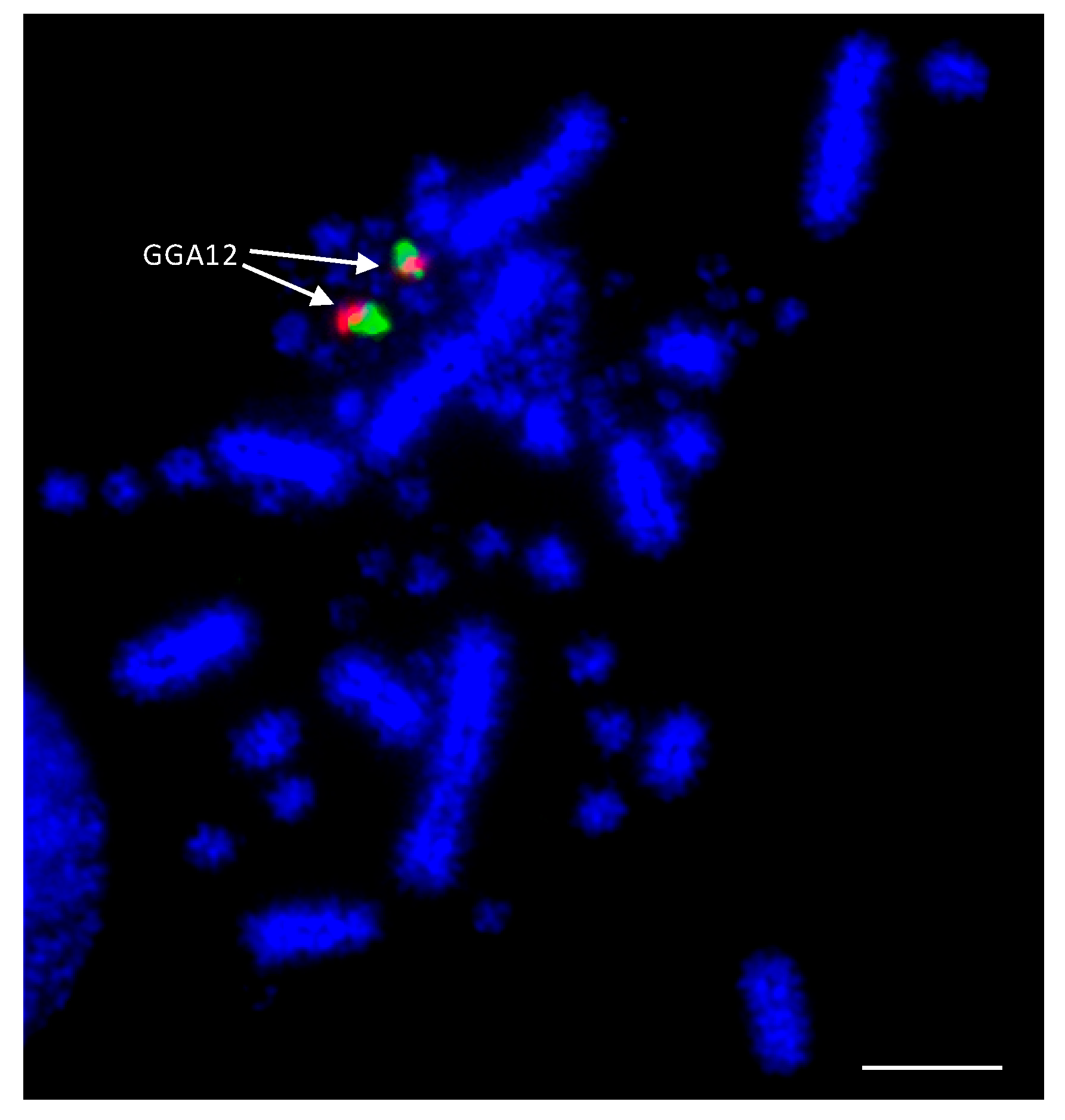
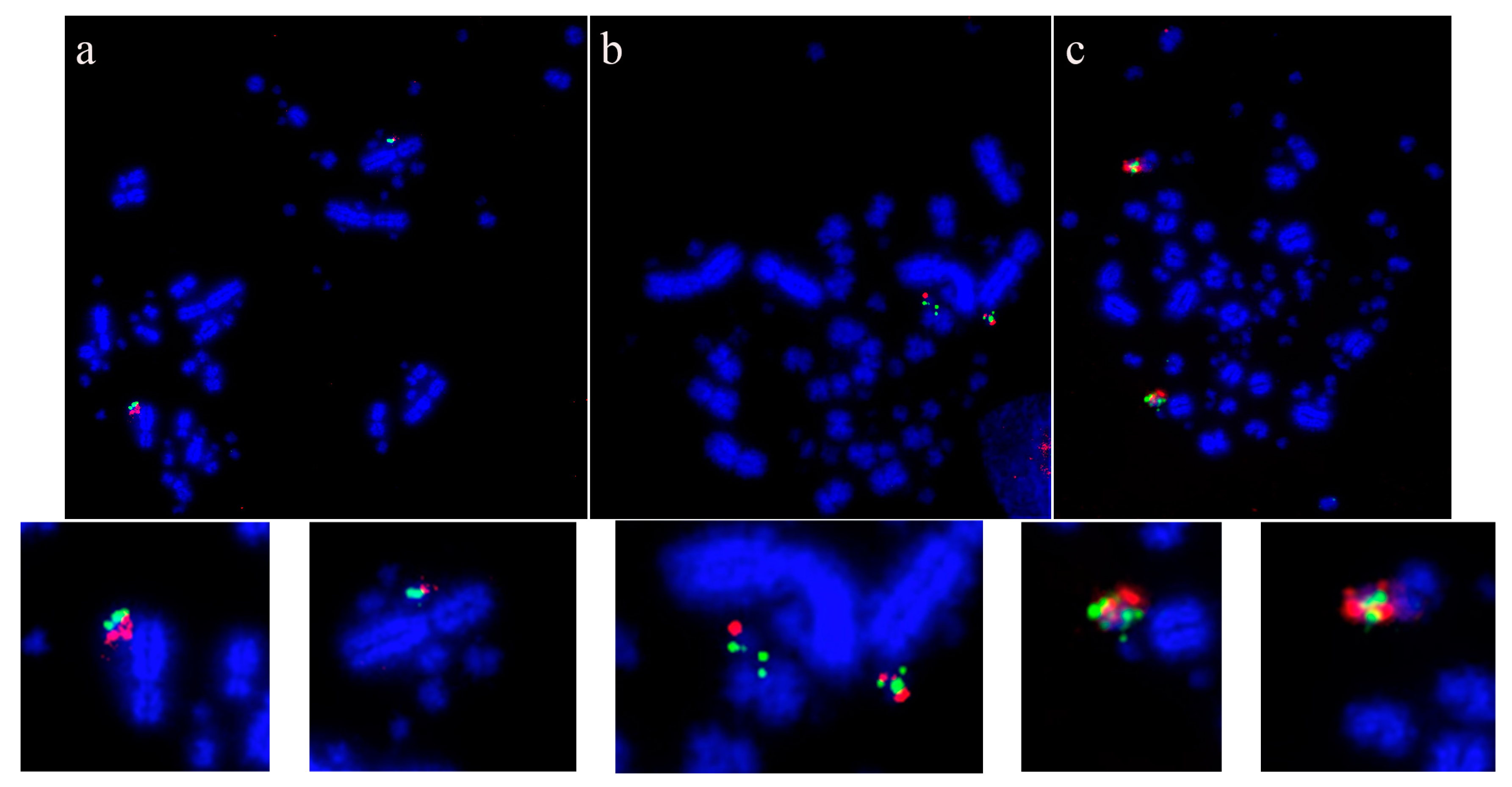
7.2. Species Demonstrating Rearrangement between Microchromosomes
7.2.1. Psittaciformes
7.2.2. Falconiformes
7.2.3. Ciconiiformes
7.2.4. Pelecaniformes
7.2.5. Caprimulgiformes, Cuculiformes, Suliformes, and Passeriformes
8. Microchromosomes and their Conservation, Whether Discrete or Fused
9. Dinosaurian Origins of the Signature Avian Karyotype
10. Why Do Some Avian Genomes Break the Rules?
11. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gill, F.; Donsker, D.; Rasmussen, P. (Eds.) IOC World Bird List (v12.2). 2022. Available online: https://www.worldbirdnames.org/new/ (accessed on 1 November 2023).
- Weir, J.T.; Schluter, D. The latitudinal gradient in recent speciation and extinction rates of birds and mammals. Science 2007, 315, 1574–1576. [Google Scholar] [CrossRef]
- Holmes, D.J.; Ottinger, M.A. Birds as long-lived animal models for the study of aging. Exp. Gerontol. 2003, 38, 1365–1375. [Google Scholar] [CrossRef]
- Farrar, V.S.; Flores, L.; Viernes, R.C.; Pereira, L.O.; Mushtari, S.; Calisi, R.M. Prolactin promotes parental responses and alters reproductive axis gene expression, but not courtship behaviors, in both sexes of a biparental bird. Horm. Behav. 2022, 144, 105217. [Google Scholar] [CrossRef]
- Mérő, T.O.; Žuljević, A.; Lengyel, S. The role of reed management and habitat quality on brood parasitism and chick survival of the brood parasitic common cuckoo. Ecol. Evol. 2023, 13, e9705. [Google Scholar] [CrossRef] [PubMed]
- Vanadzina, K.; Street, S.E.; Healy, S.D.; Laland, K.N.; Sheard, C. Global drivers of variation in cup nest size in passerine birds. J. Anim. Ecol. 2023, 92, 338–351. [Google Scholar] [CrossRef] [PubMed]
- Beacon, T.H.; Davie, J.R. The chicken model organism for epigenomic research. Genome 2021, 64, 476–489. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, R.E.; Romanov, M.N.; Kiazim, L.G.; Barrett, P.M.; Farré, M.; Damas, J.; Ferguson-Smith, M.; Valenzuela, N.; Larkin, D.M.; Griffin, D.K. Reconstruction of the diapsid ancestral genome permits chromosome evolution tracing in avian and non-avian dinosaurs. Nat. Commun. 2018, 9, 1883. [Google Scholar] [CrossRef] [PubMed]
- BirdLife International. We Have Lost Over 150 Bird Species Since 1500. 2017. Available online: http://www.datazone.birdlife.org/sowb/casestudy/we-have-lost-over-150-bird-species-since-1500 (accessed on 1 November 2023).
- Chiappe, L.M.; Dyke, G.J. The early evolutionary history of birds. J. Paleontol. Soc. Korea 2006, 22, 133–151. [Google Scholar]
- Dyke, G.; Kaiser, G.W. Living Dinosaurs the Evolutionary History of Modern Birds; Wiley Blackwell: Hoboken, NJ, USA, 2011. [Google Scholar]
- von Meyer, H. Archaeopteryx lithographica (Vogel-Feder) und Pterodactylus von Solnhofen. (Letter to Prof. Bronn of 30 September 1861). Neues Jahrb. Für Mineral. Geogn. Geol. Und Petrefaktenkunde 1861, 1861, 678–679. [Google Scholar]
- Mayr, G.; Pohl, B.; Hartman, S.; Peters, D.S. The tenth skeletal specimen of Archaeopteryx. Zool. J. Linn. Soc. 2007, 149, 97–116. [Google Scholar] [CrossRef]
- Clarke, J.A.; Tambussi, C.P.; Noriega, J.I.; Erickson, G.M.; Ketcham, R.A. Definitive fossil evidence for the extant avian radiation in the Cretaceous. Nature 2005, 433, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Witmer, L.M. The debate on avian ancestry: Phylogeny, function, and fossils. In Mesozoic Birds: Above the Heads of Dinosaurs; Chiappe, L.M., Witmer, L.M., Eds.; University of California Press: Berkeley, CA, USA, 2002; pp. 3–30. [Google Scholar]
- Jarvis, E.D.; Mirarab, S.; Aberer, A.J.; Li, B.; Houde, P.; Li, C.; Ho, S.Y.; Faircloth, B.C.; Nabholz, B.; Howard, J.T.; et al. Whole-genome analyses resolve early branches in the tree of life of modern birds. Science 2014, 346, 1320–1331. [Google Scholar] [CrossRef] [PubMed]
- Schulte, P.; Alegret, L.; Arenillas, I.; Arz, J.A.; Barton, P.J.; Bown, P.R.; Bralower, T.J.; Christeson, G.L.; Claeys, P.; Cockell, C.S.; et al. The Chicxulub asteroid impact and mass extinction at the Cretaceous-Paleogene boundary. Science 2010, 327, 1214–1218. [Google Scholar] [CrossRef] [PubMed]
- Longrich, N.R.; Tokaryk, T.; Field, D.J. Mass extinction of birds at the Cretaceous-Paleogene (K-Pg) boundary. Proc. Natl. Acad. Sci. USA 2011, 108, 15253–15257. [Google Scholar] [CrossRef] [PubMed]
- Degrandi, T.M.; Barcellos, S.A.; Costa, A.L.; Garnero, A.D.V.; Hass, I.; Gunski, R.J. Introducing the bird chromosome database: An overview of cytogenetic studies in birds. Cytogenet. Genome Res. 2020, 160, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Masabanda, J.S.; Burt, D.W.; O’Brien, P.C.; Vignal, A.; Fillon, V.; Walsh, P.S.; Cox, H.; Tempest, H.G.; Smith, J.; Habermann, F.; et al. Molecular cytogenetic definition of the chicken genome: The first complete avian karyotype. Genetics 2004, 166, 1367–1373. [Google Scholar] [CrossRef] [PubMed]
- Lithgow, P.E.; O’Connor, R.; Smith, D.; Fonseka, G.; Al Mutery, A.; Rathje, C.; Frodsham, R.; O’Brien, P.; Kasai, F.; Ferguson-Smith, M.A.; et al. Novel tools for characterising inter and intra chromosomal rearrangements in avian microchromosomes. Chromosome Res. 2014, 22, 85–97. [Google Scholar] [CrossRef]
- Griffin, D.K.; Robertson, L.B.; Tempest, H.G.; Vignal, A.; Fillon, V.; Crooijmans, R.P.; Groenen, M.A.; Deryusheva, S.; Gaginskaya, E.; Carré, W.; et al. Whole genome comparative studies between chicken and turkey and their implications for avian genome evolution. BMC Genom. 2008, 9, 168. [Google Scholar] [CrossRef]
- Skinner, B.M.; Robertson, L.B.; Tempest, H.G.; Langley, E.J.; Ioannou, D.; Fowler, K.E.; Crooijmans, R.P.; Hall, A.D.; Griffin, D.K.; Völker, M. Comparative genomics in chicken and Pekin duck using FISH mapping and microarray analysis. BMC Genom. 2009, 10, 357. [Google Scholar] [CrossRef]
- Fillon, V.; Vignoles, M.; Crooijmans, R.P.; Groenen, M.A.; Zoorob, R.; Vignal, A. FISH mapping of 57 BAC clones reveals strong conservation of synteny between Galliformes and Anseriformes. Anim. Genet. 2007, 38, 303–307. [Google Scholar] [CrossRef]
- Christidis, L. Aves. In Animal Cytogenetics. Volume 4: Chordata 3 B.; John, B., Kayano, H., Levan, A., Eds.; Gebrüder Borntraeger: Berlin, Germany, 1990. [Google Scholar]
- Bloom, S.E. A current list of chromosome numbers and variations for species of the avian subclass Carinatae. J. Hered. 1969, 60, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Ray-Chaudhuri, R. Cytotaxonomy and chromosome evolution in birds. In Cytotaxonomy and Vertebrate Evolution; Chiarelli, A.B., Capanna, E., Eds.; Academic Press: New York, NY, USA, 1973; pp. 425–483. [Google Scholar]
- Shields, G.F. Comparative avian cytogenetics: A review. Condor 1982, 84, 45–58. [Google Scholar] [CrossRef]
- de Boer, L.E.M. New developments in vertebrate cytotaxonomy VIII. A current list of references on avian karyology. Genetica 1984, 65, 3–6. [Google Scholar] [CrossRef]
- Capanna, E.; Civitelli, M.; Martinico, E.I. I cromosomi degli uccelli. Citotassonomia ed evoluzione cariotipica. Avocetta 1987, 11, 101–143. [Google Scholar]
- Ellegren, H. Evolutionary stasis: The stable chromosomes of birds. Trends Ecol. Evol. 2010, 25, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Nie, W.; O’Brien, P.C.; Ng, B.L.; Fu, B.; Volobouev, V.; Carter, N.P.; Ferguson-Smith, M.A.; Yang, F. Avian comparative genomics: Reciprocal chromosome painting between domestic chicken (Gallus gallus) and the stone curlew (Burhinus oedicnemus, Charadriiformes)—An atypical species with low diploid number. Chromosome Res. 2009, 17, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Bian, X.; Li, Q. Studies on The Karyotypes of Birds V. The 20 species of Climber birds. (Aves). Zool. Res. 1989, 10, 309–317. [Google Scholar]
- Castro, M.S.; Recco-Pimentel, S.M.; Rocha, G.T. Karyotypic characterization of Ramphastidae (Piciformes, Aves). Genet. Mol. Biol. 2002, 25, 147–150. [Google Scholar] [CrossRef]
- McQueen, H.A.; Siriaco, G.; Bird, A.P. Chicken microchromosomes are hyperacetylated, early replicating, and gene rich. Genome Res. 1998, 8, 621–630. [Google Scholar] [CrossRef]
- Smith, J.; Paton, I.R.; Bruley, C.K.; Windsor, D.; Burke, D.; Ponce de Leon, F.A.; Burt, D.W. Integration of the genetic and physical maps of the chicken macrochromosomes. Anim. Genet. 2000, 31, 20–27. [Google Scholar] [CrossRef]
- Habermann, F.A.; Cremer, M.; Walter, J.; Kreth, G.; von Hase, J.; Bauer, K.; Wienberg, J.; Cremer, C.; Cremer, T.; Solovei, I. Arrangements of macro- and microchromosomes in chicken cells. Chromosome Res. 2001, 9, 569–584. [Google Scholar] [CrossRef] [PubMed]
- Burt, D.W. Origin and evolution of avian microchromosomes. Cytogenet. Genome Res. 2002, 96, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Backström, N.; Forstmeier, W.; Schielzeth, H.; Mellenius, H.; Nam, K.; Bolund, E.; Webster, M.T.; Ost, T.; Schneider, M.; Kempenaers, B.; et al. The recombination landscape of the zebra finch Taeniopygia guttata genome. Genome Res. 2010, 20, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Schield, D.R.; Card, D.C.; Hales, N.R.; Perry, B.W.; Pasquesi, G.M.; Blackmon, H.; Adams, R.H.; Corbin, A.B.; Smith, C.F.; Ramesh, B.; et al. The origins and evolution of chromosomes, dosage compensation, and mechanisms underlying venom regulation in snakes. Genome Res. 2019, 29, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Schield, D.R.; Pasquesi, G.I.M.; Perry, B.W.; Adams, R.H.; Nikolakis, Z.L.; Westfall, A.K.; Orton, R.W.; Meik, J.M.; Mackessy, S.P.; Castoe, T.A. Snake recombination landscapes are concentrated in functional regions despite PRDM9. Mol. Biol. Evol. 2020, 37, 1272–1294. [Google Scholar] [CrossRef]
- Nakatani, Y.; Takeda, H.; Kohara, Y.; Morishita, S. Reconstruction of the vertebrate ancestral genome reveals dynamic genome reorganization in early vertebrates. Genome Res. 2007, 17, 1254–1265. [Google Scholar] [CrossRef]
- Graves, J.A.M.; Shetty, S. Sex from W to Z: Evolution of vertebrate sex chromosomes and sex determining genes. J. Exp. Zool. 2001, 290, 449–462. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, J.; Yang, W.; An, N.; Zhang, P.; Zhang, G.; Zhou, Q. Temporal genomic evolution of bird sex chromosomes. BMC Evol. Biol. 2014, 14, 250. [Google Scholar] [CrossRef]
- Schmid, M.; Enderle, E.; Schindler, D.; Schemp, W. Chromosome banding and DNA replication patterns in bird karyotypes. Cytogenet. Cell Genet. 1989, 52, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Solari, A.J. Sex Chromosomes and Sex Determination in Vertebrates; CRC Press: Boca Raton, FL, USA, 1993. [Google Scholar]
- Nanda, I.; Schmid, M. Conservation of avian Z chromosomes as revealed by comparative mapping of the Z-linked aldolase B gene. Cytogenet. Genome Res. 2002, 96, 176–178. [Google Scholar] [CrossRef] [PubMed]
- Schartl, M.; Schmid, M.; Nanda, I. Dynamics of vertebrate sex chromosome evolution: From equal size to giants and dwarfs. Chromosoma 2016, 125, 553–571. [Google Scholar] [CrossRef]
- Shetty, S.; Griffin, D.K.; Graves, J.A.M. Comparative painting reveals strong chromosome homology over 80 million years of bird evolution. Chromosome Res. 1999, 7, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Takagi, N.; Itoh, M.; Sasaki, M. Chromosome studies in four species of Ratitae (Aves). Chromosoma 1972, 36, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Ansari, H.A.; Takagi, N.; Sasaki, M. Morphological differentiation of sex chromosomes in three species of ratite birds. Cytogenet. Cell Genet. 1988, 47, 185–188. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Z.; Li, J.; Xu, L.; Liu, J.; Feng, S.; Guo, C.; Chen, S.; Ren, Z.; Rao, J.; et al. A new emu genome illuminates the evolution of genome configuration and nuclear architecture of avian chromosomes. Genome Res. 2021, 31, 497–511. [Google Scholar] [CrossRef] [PubMed]
- Deakin, J.E.; Ezaz, T. Tracing the evolution of amniote chromosomes. Chromosoma 2014, 123, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Nanda, I.; Shan, Z.; Schartl, M.; Burt, D.W.; Koehler, M.; Nothwang, H.; Grützner, F.; Paton, I.R.; Windsor, D.; Dunn, I.; et al. 300 million years of conserved synteny between chicken Z and human chromosome 9. Nat. Genet. 1999, 21, 258–259. [Google Scholar] [CrossRef]
- Bellott, D.W.; Skaletsky, H.; Pyntikova, T.; Mardis, E.R.; Graves, T.; Kremitzki, C.; Brown, L.G.; Rozen, S.; Warren, W.C.; Wilson, R.K.; et al. Convergent evolution of chicken Z and human X chromosomes by expansion and gene acquisition. Nature 2010, 466, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Ross, M.T.; Grafham, D.V.; Coffey, A.J.; Scherer, S.; McLay, K.; Muzny, D.; Platzer, M.; Howell, G.R.; Burrows, C.; Bird, C.P.; et al. The DNA sequence of the human X chromosome. Nature 2005, 434, 325–337. [Google Scholar] [CrossRef]
- Graves, J.A. Avian sex, sex chromosomes, and dosage compensation in the age of genomics. Chromosome Res. 2014, 22, 45–57. [Google Scholar] [CrossRef]
- Smith, C.A.; Roeszler, K.N.; Ohnesorg, T.; Cummins, D.M.; Farlie, P.G.; Doran, T.J.; Sinclair, A.H. The avian Z-linked gene DMRT1 is required for male sex determination in the chicken. Nature 2009, 461, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Bellott, D.W.; Page, D.C.; Clark, A.G. Identification of avian W-linked contigs by short-read sequencing. BMC Genom. 2012, 14, 183. [Google Scholar] [CrossRef] [PubMed]
- Gunski, R.J.; Cañedo, A.D.; Garnero, A.D.V.; Ledesma, M.A.; Coria, N.; Montalti, D.; Degrandi, T.M. Multiple sex chromosome system in penguins (Pygoscelis, Spheniscidae). Comp. Cytogenet. 2017, 11, 541–552. [Google Scholar] [CrossRef]
- Sigeman, H.; Ponnikas, S.; Hansson, B. Whole-genome analysis across 10 songbird families within Sylvioidea reveals a novel autosome–sex chromosome fusion. Biol. Lett. 2020, 16, 20200082. [Google Scholar] [CrossRef]
- Huang, Z.; Furo, I.O.; Liu, J.; Peona, V.; Gomes, A.J.B.; Cen, W.; Huang, H.; Zhang, Y.; Chen, D.; Xue, T.; et al. Recurrent chromosome reshuffling and the evolution of neo-sex chromosomes in parrots. Nat. Commun. 2022, 13, 944. [Google Scholar] [CrossRef]
- Kretschmer, R.; Gunski, R.J.; Garnero, A.d.V.; de Freitas, T.R.O.; Toma, G.A.; Cioffi, M.d.B.; Oliveira, E.H.C.d.; O’Connor, R.E.; Griffin, D.K. Chromosomal analysis in Crotophaga ani (Aves, Cuculiformes) reveals extensive genomic reorganization and an unusual Z-autosome Robertsonian translocation. Cells 2021, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, R.E.; Kiazim, L.; Skinner, B.; Fonseka, G.; Joseph, S.; Jennings, R.; Larkin, D.M.; Griffin, D.K. Patterns of microchromosome organization remain highly conserved throughout avian evolution. Chromosoma 2019, 128, 21–29. [Google Scholar] [CrossRef]
- Ferguson-Smith, M.A.; Trifonov, V. Mammalian karyotype evolution. Nat. Rev. Genet. 2007, 8, 950–962. [Google Scholar] [CrossRef]
- Graphodatsky, A.S.; Trifonov, V.A.; Stanyon, R. The genome diversity and karyotype evolution of mammals. Mol. Cytogenet. 2011, 4, 22. [Google Scholar] [CrossRef]
- Damas, J.; Corbo, M.; Lewin, H.A. Vertebrate Chromosome Evolution. Annu. Rev. Anim. Biosci. 2021, 9, 1–27. [Google Scholar] [CrossRef]
- Kretschmer, R.; Gunski, R.J.; Garnero, A.D.V.; Furo, I.O.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Molecular cytogenetic characterization of multiple intrachromosomal rearrangements in two representatives of the genus Turdus (Turdidae, Passeriformes). PLoS ONE 2014, 9, e103338. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; Furo, I.O.; Gunski, R.J.; Garnero, A.D.V.; Pereira, J.C.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C.; de Freitas, T.R.O. Comparative chromosome painting in Columbidae (Columbiformes) reinforces divergence in Passerea and Columbea. Chromosome Res. 2018, 26, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Kiazim, L.G.; O’Connor, R.E.; Larkin, D.M.; Romanov, M.N.; Narushin, V.G.; Brazhnik, E.A.; Griffin, D.K. Comparative mapping of the macrochromosomes of eight avian species provides further insight into their phylogenetic relationships and avian karyotype evolution. Cells 2021, 10, 362. [Google Scholar] [CrossRef] [PubMed]
- Choudhuri, J.V.; Schleiermacher, C.; Kurtz, S.; Giegerich, R. GenAlyzer: Interactive visualization of sequence similarities between entire genomes. Bioinformatics 2004, 20, 1964–1965. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.; Morisson, M.; Faraut, T.; Bardes, S.; Fève, K.; Labarthe, E.; Fillon, V.; Huang, Y.; Li, N.; Vignal, A. A duck RH panel and its potential for assisting NGS genome assembly. BMC Genom. 2012, 13, 513. [Google Scholar] [CrossRef] [PubMed]
- Raudsepp, T.; Houck, M.L.; O’Brien, P.C.; Ferguson-Smith, M.A.; Ryder, O.A.; Chowdhary, B.P. Cytogenetic analysis of California condor (Gymnogyps californianus) chromosomes: Comparison with chicken (Gallus gallus) macrochromosomes. Cytogenet. Genome Res. 2002, 98, 54–60. [Google Scholar] [CrossRef]
- Shibusawa, M.; Nishida-Umehara, C.; Masabanda, J.; Griffin, D.K.; Isobe, T.; Matsuda, Y. Chromosome rearrangements between chicken and guinea fowl defined by comparative chromosome painting and FISH mapping of DNA clones. Cytogenet. Genome Res. 2002, 98, 225–230. [Google Scholar] [CrossRef]
- Itoh, Y.; Arnold, A.P. Chromosomal polymorphism and comparative painting analysis in the zebra finch. Chromosome Res. 2005, 13, 47–56. [Google Scholar] [CrossRef]
- Nanda, I.; Benisch, P.; Fetting, D.; Haaf, T.; Schmid, M. Synteny conservation of chicken macrochromosomes 1–10 in different avian lineages revealed by cross-species chromosome painting. Cytogenet. Genome Res. 2011, 132, 165–181. [Google Scholar] [CrossRef]
- Nishida, C.; Ishijima, J.; Kosaka, A.; Tanabe, H.; Habermann, F.A.; Griffin, D.K.; Matsuda, Y. Characterization of chromosome structures of Falconinae (Falconidae, Falconiformes, Aves) by chromosome painting and delineation of chromosome rearrangements during their differentiation. Chromosome Res. 2008, 16, 171–181. [Google Scholar] [CrossRef]
- Nishida-Umehara, C.; Tsuda, Y.; Ishijima, J.; Ando, J.; Fujiwara, A.; Matsuda, Y.; Griffin, D.K. The molecular basis of chromosome orthologies and sex chromosomal differentiation in palaeognathous birds. Chromosome Res. 2007, 15, 721–734. [Google Scholar] [CrossRef] [PubMed]
- Guttenbach, M.; Nanda, I.; Feichtinger, W.; Masabanda, J.S.; Griffin, D.K.; Schmid, M. Comparative chromosome painting of chicken autosomal paints 1-9 in nine different bird species. Cytogenet. Genome Res. 2003, 103, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Kasai, F.; Garcia, C.; Arruga, M.V.; Ferguson-Smith, M.A. Chromosome homology between chicken (Gallus gallus domesticus) and the red-legged partridge (Alectoris rufa): Evidence of the occurrence of a neocentromere during evolution. Cytogenet. Genome Res. 2003, 102, 326–330. [Google Scholar] [CrossRef]
- Shibusawa, M.; Nishibori, M.; Nishida-Umehara, C.; Tsudzuki, M.; Masabanda, J.; Griffin, D.K.; Matsuda, Y. Karyotypic evolution in the Galliformes: An examination of the process of karyotypic evolution by comparison of the molecular cytogenetic findings with the molecular phylogeny. Cytogenet. Genome Res. 2004, 106, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Shibusawa, M.; Nishida-Umehara, C.; Tsudzuki, M.; Masabanda, J.; Griffin, D.K.; Matsuda, Y. A comparative karyological study of the blue-breasted quail (Coturnix chinensis, Phasianidae) and California quail (Callipepla californica, Odontophoridae). Cytogenet. Genome Res. 2004, 106, 82–90. [Google Scholar] [CrossRef]
- Islam, F.B.; Uno, Y.; Nunome, M.; Nishimura, O.; Tarui, H.; Agata, K.; Matsuda, Y. Comparison of the chromosome structures between the chicken and three anserid species, the domestic duck (Anas platyrhynchos), Muscovy duck (Cairina moschata), and Chinese goose (Anser cygnoides), and the delineation of their karyotype evolution by comparative chromosome mapping. J. Poult. Sci. 2014, 51, 1–13. [Google Scholar] [CrossRef]
- Rodrigues, B.S.; de Assis, M.D.F.L.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Chromosomal studies on Coscoroba coscoroba reinforce the Coscoroba–Cereopsis clade. Biol. J. Linn. Soc. 2014, 111, 274–279. [Google Scholar] [CrossRef]
- Uno, Y.; Nishida, C.; Hata, A.; Ishishita, S.; Matsuda, Y. Molecular cytogenetic characterization of repetitive sequences comprising centromeric heterochromatin in three Anseriformes species. PLoS ONE 2019, 14, e0214028. [Google Scholar] [CrossRef]
- Degrandi, T.M.; Furo, I.O.; de Oliveira, E.H.C.; Costa, A.L.; Ferguson-Smith, M.A.; O’Brien, P.C.M.; Pereira, J.C.; Garnero, A.D.V.; Gunski, R.J.; Artoni, R.F. Comparative chromosome painting in hummingbirds (Trochilidae). Genet. Mol. Biol. 2020, 43, e20200162. [Google Scholar] [CrossRef]
- dos Santos, M.S.; Kretschmer, R.; Furo, I.O.; Gunski, R.J.; Garnero, A.D.V.; Valeri, M.P.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Chromosomal evolution and phylogenetic considerations in cuckoos (Aves, Cuculiformes, Cuculidae). PLoS ONE 2020, 15, e0232509. [Google Scholar] [CrossRef]
- Derjusheva, S.; Kurganova, A.; Habermann, F.; Gaginskaya, E. High chromosome conservation detected by comparative chromosome painting in chicken, pigeon and passerine birds. Chromosome Res. 2004, 12, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; Furo, I.d.O.; Gomes, A.J.B.; Kiazim, L.G.; Gunski, R.J.; Garnero, A.d.V.; Pereira, J.C.; Ferguson-Smith, M.A.; de Oliveira, E.H.C.; Griffin, D.K.; et al. A comprehensive cytogenetic analysis of several members of the family Columbidae (Aves, Columbiformes). Genes 2020, 11, 632. [Google Scholar] [CrossRef] [PubMed]
- Furo, I.O.; Kretschmer, R.; O’Brien, P.C.M.; Pereira, J.C.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Phylogenetic analysis and karyotype evolution in two species of core Gruiformes: Aramides cajaneus and Psophia viridis. Genes 2020, 11, 307. [Google Scholar] [CrossRef] [PubMed]
- Furo, I.O.; Kretschmer, R.; O’Brien, P.C.M.; Pereira, J.C.C.; Gunski, R.J.; Garnero, A.D.V.; O’Connor, R.E.; Griffin, D.K.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Cytotaxonomy of Gallinula melanops (Gruiformes, Rallidae): Karyotype evolution and phylogenetic inference. Genet. Mol. Biol. 2021, 44, e20200241. [Google Scholar] [CrossRef] [PubMed]
- Hansmann, T.; Nanda, I.; Volobouev, V.; Yang, F.; Schartl, M.; Haaf, T.; Schmid, M. Cross-species chromosome painting corroborates microchromosome fusion during karyotype evolution of birds. Cytogenet. Genome Res. 2009, 126, 281–304. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; Gunski, R.J.; Garnero, A.D.V.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Freitas, T.R.O.; de Oliveira, E.H.C. Chromosome painting in Vanellus chilensis: Detection of a fusion common to clade Charadrii (Charadriiformes). Cytogenet. Genome Res. 2015, 146, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; Souza, M.S.; Barcellos, S.A.; Degrandi, T.M.; Pereira, J.C.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Gunski, R.J.; Garnero, A.D.V.; de Oliveira, E.H.C.; et al. Novel insights into chromosome evolution of Charadriiformes: Extensive genomic reshuffling in the wattled jacana (Jacana jacana, Charadriiformes, Jacanidae). Genet. Mol. Biol. 2020, 43, e20190236. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, M.L.S.; Nagamachi, C.Y.; Ribas, T.F.A.; Diniz, C.G.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Yang, F.; Pieczarka, J.C. Chromosomal painting of the sandpiper (Actitis macularius) detects several fissions for the Scolopacidae family (Charadriiformes). BMC Ecol. Evol. 2021, 21, 8. [Google Scholar] [CrossRef]
- Pinheiro, M.L.S.; Nagamachi, C.Y.; Ribas, T.F.A.; Diniz, C.G.; O Brien, P.C.M.; Ferguson-Smith, M.A.; Yang, F.; Pieczarka, J.C. Chromosomal painting in Charadrius collaris Vieillot, 1818 and Vanellus chilensis Molina, 1782 and an analysis of chromosomal signatures in Charadriiformes. PLoS ONE 2022, 17, e0272836. [Google Scholar] [CrossRef]
- Furo, I.O.; Monte, A.A.; dos Santos, M.S.; Tagliarini, M.M.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Cytotaxonomy of Eurypyga helias (Gruiformes, Eurypygidae): First karyotypic description and phylogenetic proximity with Rynochetidae. PLoS ONE 2015, 10, e0143982. [Google Scholar] [CrossRef]
- Seligmann, I.C.A.; Furo, I.O.; dos Santos, M.S.; Tagliarini, M.M.; Araujo, C.C.D.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Comparative chromosome painting in two Brazilian stork species with different diploid numbers. Cytogenet. Genome Res. 2019, 159, 32–38. [Google Scholar] [CrossRef]
- Wang, J.; Su, W.; Hu, Y.; Li, S.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Yang, F.; Nie, W. Comparative chromosome maps between the stone curlew and three ciconiiform species (the grey heron, little egret and crested ibis). BMC Ecol. Evol. 2022, 22, 23. [Google Scholar] [CrossRef] [PubMed]
- Seligmann, I.C.A.; Furo, I.O.; dos Santos, M.S.; Gunski, R.J.; Garnero, A.D.V.; Silva, F.A.O.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Kretschmer, R.; de Oliveira, E.H.C. Comparative chromosome painting in three Pelecaniformes species (Aves): Exploring the role of macro and microchromosome fusions in karyotypic evolution. PLoS ONE 2023, 18, e0294776. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, M.S.; Furo, I.O.; Tagliarini, M.M.; Kretschmer, R.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. The karyotype of the hoatzin (Opisthocomus hoazin)—A phylogenetic enigma of the Neornithes. Cytogenet. Genome Res. 2018, 156, 158–164. [Google Scholar] [CrossRef]
- de Oliveira, E.H.C.; Habermann, F.A.; Lacerda, O.; Sbalqueiro, I.J.; Wienberg, J.; Müller, S. Chromosome reshuffling in birds of prey: The karyotype of the world’s largest eagle (Harpy eagle, Harpia harpyja) compared to that of the chicken (Gallus gallus). Chromosoma 2005, 114, 338–343. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, E.H.C.; Tagliarini, M.M.; Rissino, J.D.; Pieczarka, J.C.; Nagamachi, C.Y.; O’Brien, P.C.M.; Ferguson-Smith, M.A. Reciprocal chromosome painting between white hawk (Leucopternis albicollis) and chicken reveals extensive fusions and fissions during karyotype evolution of Accipitridae (Aves, Falconiformes). Chromosome Res. 2010, 18, 349–355. [Google Scholar] [CrossRef]
- de Oliveira, E.H.C.; Tagliarini, M.M.; dos Santos, M.S.; O’Brien, P.C.M.; Ferguson-Smith, M.A. Chromosome painting in three species of Buteoninae: A cytogenetic signature reinforces the monophyly of South American species. PLoS ONE 2013, 8, e70071. [Google Scholar] [CrossRef]
- Nanda, I.; Karl, E.; Volobouev, V.; Griffin, D.K.; Schartl, M.; Schmid, M. Extensive gross genomic rearrangements between chicken and Old World vultures (Falconiformes: Accipitridae). Cytogenet. Genome Res. 2006, 112, 286–295. [Google Scholar] [CrossRef]
- Tagliarini, M.M.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Maintenance of syntenic groups between Cathartidae and Gallus gallus indicates symplesiomorphic karyotypes in new world vultures. Genet. Mol. Biol. 2011, 34, 80–83. [Google Scholar] [CrossRef]
- Nishida, C.; Ishijima, J.; Ishishita, S.; Yamada, K.; Griffin, D.K.; Yamazaki, T.; Matsuda, Y. Karyotype reorganization with conserved genomic compartmentalization in dot-shaped microchromosomes in the Japanese mountain hawk-eagle (Nisaetus nipalensis orientalis, Accipitridae). Cytogenet. Genome Res. 2013, 141, 284–294. [Google Scholar] [CrossRef]
- Nishida, C.; Ishishita, S.; Yamada, K.; Griffin, D.K.; Matsuda, Y. Dynamic chromosome reorganization in the osprey (Pandion haliaetus, Pandionidae, Falconiformes): Relationship between chromosome size and the chromosomal distribution of centromeric repetitive DNA sequences. Cytogenet. Genome Res. 2014, 142, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Nie, W.; O’Brien, P.C.; Fu, B.; Wang, J.; Su, W.; He, K.; Bed’Hom, B.; Volobouev, V.; Ferguson-Smith, M.A.; Dobigny, G.; et al. Multidirectional chromosome painting substantiates the occurrence of extensive genomic reshuffling within Accipitriformes. BMC Evol. Biol. 2015, 15, 205. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.A.; Furo, I.O.; O’Brien, P.C.M.; Pereira, J.; O’Connor, R.E.; Griffin, D.; Ferguson-Smith, M.; de Oliveira, E.H.C. Comparative chromosome painting in Spizaetus tyrannus and Gallus gallus with the use of macro- and microchromosome probes. PLoS ONE 2021, 18, e0259905. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, E.H.C.; de Moura, S.P.; dos Anjos, L.J.S.; Nagamachi, C.Y.; Pieczarka, J.C.; O’Brien, P.C.M.; Ferguson-Smith, M.A. Comparative chromosome painting between chicken and spectacled owl (Pulsatrix perspicillata): Implications for chromosomal evolution in the Strigidae (Aves, Strigiformes). Cytogenet. Genome Res. 2008, 122, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Degrandi, T.M.; Garnero, A.D.V.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Kretschmer, R.; de Oliveira, E.H.C.; Gunski, R.J. Chromosome painting in Trogon s. surrucura (Aves, Trogoniformes) reveals a karyotype derived by chromosomal fissions, fusions, and inversions. Cytogenet. Genome Res. 2017, 151, 208–215. [Google Scholar] [CrossRef]
- Kretschmer, R.; Furo, I.O.; Cioffi, M.B.; Gunski, R.J.; Garnero, A.D.V.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Freitas, T.R.O.; de Oliveira, E.H.C. Extensive chromosomal fissions and repetitive DNA accumulation shaped the atypical karyotypes of two Ramphastidae (Aves: Piciformes) species. Biol. J. Linn. Soc. 2020, 130, 839–849. [Google Scholar] [CrossRef]
- Joseph, S.; O’Connor, R.E.; Al Mutery, A.F.; Watson, M.; Larkin, D.M.; Griffin, D.K. Chromosome level genome assembly and comparative genomics between three Falcon species reveals an unusual pattern of genome organisation. Diversity 2018, 10, 113. [Google Scholar] [CrossRef]
- Nanda, I.; Karl, E.; Griffin, D.K.; Schartl, M.; Schmid, M. Chromosome repatterning in three representative parrots (Psittaciformes) inferred from comparative chromosome painting. Cytogenet. Genome Res. 2007, 117, 43–53. [Google Scholar] [CrossRef]
- Seabury, C.M.; Dowd, S.E.; Seabury, P.M.; Raudsepp, T.; Brightsmith, D.J.; Liboriussen, P.; Halley, Y.; Fisher, C.A.; Owens, E.; Viswanathan, G.; et al. A multi-platform draft de novo genome assembly and comparative analysis for the Scarlet Macaw (Ara macao). PLoS ONE 2013, 8, e62415. [Google Scholar] [CrossRef]
- Furo, I.O.; Kretschmer, R.; O’Brien, P.C.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Chromosomal diversity and karyotype evolution in South American macaws (Psittaciformes, Psittacidae). PLoS ONE 2015, 10, e0130157. [Google Scholar] [CrossRef]
- Furo, I.O.; Kretschmer, R.; O’Brien, P.C.M.; Pereira, J.C.; Garnero, A.D.V.; Gunski, R.J.; Ferguson-Smith, M.A.; De Oliveira, E.H.C. Chromosome painting in Neotropical long- and short-tailed parrots (Aves, Psittaciformes): Phylogeny and proposal for a putative ancestral karyotype for tribe Arini. Genes 2018, 9, 491. [Google Scholar] [CrossRef] [PubMed]
- Furo, I.O.; Kretschmer, R.; O’Brien, P.C.M.; Pereira, J.; Garnero, A.D.V.; Gunski, R.J.; O’Connor, R.E.; Griffin, D.K.; Gomes, A.J.B.; Ferguson-Smith, M.A.; et al. Chromosomal evolution in the phylogenetic context: A remarkable karyotype reorganization in neotropical parrot Myiopsitta monachus (Psittacidae). Front. Genet. 2020, 11, 721. [Google Scholar] [CrossRef]
- Seibold-Torres, C.; Owens, E.; Chowdhary, R.; Ferguson-Smith, M.A.; Tizard, I.; Raudsepp, T. Comparative cytogenetics of the Congo African grey parrot (Psittacus erithacus). Cytogenet. Genome Res. 2015, 147, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; de Oliveira, E.H.C.; dos Santos, M.S.; Furo, I.O.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Garnero, A.D.V.; Gunski, R.J. Chromosome mapping of the large elaenia (Elaenia spectabilis): Evidence for a cytogenetic signature for passeriform birds? Biol. J. Linn. Soc. 2015, 115, 391–398. [Google Scholar] [CrossRef]
- Kretschmer, R.; de Lima, V.L.C.; de Souza, M.S.; Costa, A.L.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C.; Gunski, R.J.; Garnero, A.D.V. Multidirectional chromosome painting in Synallaxis frontalis (Passeriformes, Furnariidae) reveals high chromosomal reorganization, involving fissions and inversions. Comp. Cytogenet. 2018, 12, 97–110. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, M.S.; Kretschmer, R.; Silva, F.A.O.; Ledesma, M.A.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Garnero, A.D.V.; de Oliveira, E.H.C.; Gunski, R.J. Intrachromosomal rearrangements in two representatives of the genus Saltator (Thraupidae, Passeriformes) and the occurrence of heteromorphic Z chromosomes. Genetica 2015, 143, 535–543. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, M.S.; Kretschmer, R.; Frankl-Vilches, C.; Bakker, A.; Gahr, M.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Comparative cytogenetics between two important songbird, models: The zebra finch and the canary. PLoS ONE 2017, 12, e0170997. [Google Scholar] [CrossRef] [PubMed]
- Ribas, T.F.A.; Nagamachi, C.Y.; Aleixo, A.; Pinheiro, M.L.S.; O´Brien, P.C.M.; Ferguson-Smith, M.A.; Yang, F.; Suarez, P.; Pieczarka, J.C. Chromosome painting in Glyphorynchus spirurus (Vieillot, 1819) detects a new fission in Passeriformes. PLoS ONE 2018, 13, e0202040. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, B.S.; Kretschmer, R.; Gunski, R.J.; Garnero, A.D.V.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Chromosome painting in tyrant flycatchers confirms a set of inversions shared by Oscines and Suboscines (Aves, Passeriformes). Cytogenet. Genome Res. 2017, 153, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Bülau, S.E.; Kretschmer, R.; Gunski, R.J.; Garnero, A.D.V.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C.; de Freitas, T.R.O. Chromosomal polymorphism and comparative chromosome painting in the rufous-collared sparrow (Zonotrichia capensis). Genet. Mol. Biol. 2018, 41, 799–805. [Google Scholar] [CrossRef]
- Barcellos, S.A.; Kretschmer, R.; de Souza, M.S.; Costa, A.L.; Degrandi, T.M.; Lopes, C.F.; Ferguson-Smith, M.A.; Pereira, J.; de Oliveira, E.H.C.; Gunski, R.J.; et al. Comparative analyses of three swallow species (Aves, Passeriformes, Hirundinidae): Insights on karyotype evolution and genomic organization. Genet. Mol. Biol. 2020, 43, e20190232. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, T.D.; Kretschmer, R.; Bertocchi, N.A.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Garnero, A.D.V.; de Oliveira, E.H.C.; Gunski, R.J. The molecular cytogenetic characterization of Conopophaga lineata indicates a common chromosome rearrangement in the Parvorder Furnariida (Aves, Passeriformes). Genet. Mol. Biol. 2020, 43, e20200018. [Google Scholar] [CrossRef] [PubMed]
- International Chicken Genome Sequencing Consortium. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 2004, 432, 695–716. [Google Scholar] [CrossRef]
- Griffin, D.K.; Masabanda, J.S. Microchromosomal probes derived from individual chromosome microdissection were not successfully sequenced and since degraded in the freezer. 2006; unpublished results. [Google Scholar]
- Damas, J.; O’Connor, R.; Farré, M.; Lenis, V.P.E.; Martell, H.J.; Mandawala, A.; Fowler, K.; Joseph, S.; Swain, M.T.; Griffin, D.K.; et al. Upgrading short-read animal genome assemblies to chromosome level using comparative genomics and a universal probe set. Genome Res. 2017, 27, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Xu, Z.; Bai, H.; Huang, Y.; Kang, N.; Ding, X.; Liu, J.; Luo, H.; Yang, C.; Chen, W.; et al. Evolutionary analysis of a complete chicken genome. Proc. Natl. Acad. Sci. USA 2023, 120, e2216641120. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, Y.; Nishida-Umehara, C.; Tarui, H.; Kuroiwa, A.; Yamada, K.; Isobe, T.; Ando, J.; Fujiwara, A.; Hirao, Y.; Nishimura, O.; et al. Highly conserved linkage homology between birds and turtles: Bird and turtle chromosomes are precise counterparts of each other. Chromosome Res. 2005, 13, 601–615. [Google Scholar] [CrossRef] [PubMed]
- Kasai, F.; O’Brien, P.C.; Ferguson-Smith, M.A. Reassessment of genome size in turtle and crocodile based on chromosome measurement by flow karyotyping: Close similarity to chicken. Biol. Lett. 2012, 8, 631–635. [Google Scholar] [CrossRef]
- Chowdhary, B.P.; Raudsepp, T.; Frönicke, L.; Scherthan, H. Emerging patterns of comparative genome organization in some mammalian species as revealed by Zoo-FISH. Genome Res. 1998, 8, 577–589. [Google Scholar] [CrossRef]
- Dodgson, J.B.; Delany, M.E.; Cheng, H.H. Poultry genome sequences: Progress and outstanding challenges. Cytogenet. Genome Res. 2011, 134, 19–26. [Google Scholar] [CrossRef]
- O’Connor, R.E.; Farré, M.; Joseph, S.; Damas, J.; Kiazim, L.; Jennings, R.; Bennett, S.; Slack, E.A.; Allanson, E.; Larkin, D.M.; et al. Chromosome-level assembly reveals extensive rearrangement in saker falcon and budgerigar, but not ostrich, genomes. Genome Biol. 2018, 19, 171. [Google Scholar] [CrossRef]
- Cheng, Y.; Burt, D.W. Chicken genomics. Int. J. Dev. Biol. 2018, 62, 265–271. [Google Scholar] [CrossRef]
- Kretschmer, R.; de Souza, M.S.; Furo, I.d.O.; Romanov, M.N.; Gunski, R.J.; Garnero, A.d.V.; de Freitas, T.R.O.; de Oliveira, E.H.C.; O’Connor, R.E.; Griffin, D.K. Interspecies chromosome mapping in Caprimulgiformes, Piciformes, Suliformes, and Trogoniformes (Aves): Cytogenomic insight into microchromosome organization and karyotype evolution in birds. Cells 2021, 10, 826. [Google Scholar] [CrossRef]
- de Souza, M.S.; Barcellos, S.A.; dos Santos, M.d.S.; Gunski, R.J.; Garnero, A.d.V.; de Oliveira, E.H.C.; O’Connor, R.E.; Griffin, D.K.; Kretschmer, R. Microchromosome BAC-FISH reveals different patterns of genome organization in three Charadriiformes species. Animals 2022, 12, 3052. [Google Scholar] [CrossRef]
- Kretschmer, R.; Franz, I.; de Souza, M.S.; Garnero, A.D.V.; Gunski, R.J.; de Oliveira, E.H.C.; O’Connor, R.E.; Griffin, D.K.; de Freitas, T.R.O. Cytogenetic evidence clarifies the phylogeny of the family Rhynchocyclidae (Aves: Passeriformes). Cells 2021, 10, 2650. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; Rodrigues, B.S.; Barcellos, S.A.; Costa, A.L.; Cioffi, M.d.B.; Garnero, A.d.V.; Gunski, R.J.; de Oliveira, E.H.C.; Griffin, D.K. Karyotype evolution and genomic organization of repetitive DNAs in the Saffron finch, Sicalis flaveola (Passeriformes, Aves). Animals 2021, 11, 1456. [Google Scholar] [CrossRef] [PubMed]
- Ribas, T.F.A.; Pieczarka, J.C.; Griffin, D.K.; Kiazim, L.G.; Nagamachi, C.Y.; O´Brien, P.C.M.; Ferguson-Smith, M.A.; Yang, F.; Aleixo, A.; O’Connor, R.E. Analysis of multiple chromosomal rearrangements in the genome of Willisornis vidua using BAC-FISH and chromosome painting on a supposed conserved karyotype. BMC Ecol. Evol. 2021, 21, 34. [Google Scholar] [CrossRef]
- de Souza, M.S.; Barcellos, S.A.; Tura, V.; Bobrowski, V.L.; Garnero, A.D.V.; Gunski, R.J.; Griffin, D.K.; Kretschmer, R. Highly Conserved Microchromosomal Organization in Passeriformes Birds Revealed via BAC-FISH Analysis. Birds 2023, 4, 236–244. [Google Scholar] [CrossRef]
- Pala, I.; Hasselquist, D.; Bensch, S.; Hansson, B. Patterns of molecular evolution of an avian neo-sex chromosome. Mol. Biol. Evol. 2012, 29, 3741–3754. [Google Scholar] [CrossRef]
- Pala, I.; Naurin, S.; Stervander, M.; Hasselquist, D.; Bensch, S.; Hansson, B. Evidence of a neo-sex chromosome in birds. Heredity 2012, 108, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Sigeman, H.; Ponnikas, S.; Chauhan, P.; Dierickx, E.; Brooke, M.d.L.; Hansson, B. Repeated sex chromosome evolution in vertebrates supported by expanded avian sex chromosomes. Proc. R. Soc. B 2019, 286, 20192051. [Google Scholar] [CrossRef]
- Ellegren, H. The evolutionary genomics of birds. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 239–259. [Google Scholar] [CrossRef]
- Romanov, M.N.; Farré, M.; Lithgow, P.E.; Fowler, K.E.; Skinner, B.M.; O’Connor, R.; Fonseka, G.; Backström, N.; Matsuda, Y.; Nishida, C.; et al. Reconstruction of gross avian genome structure, organization and evolution suggests that the chicken lineage most closely resembles the dinosaur avian ancestor. BMC Genom. 2014, 15, 1060. [Google Scholar] [CrossRef] [PubMed]
- Maslova, A.; Zlotina, A.; Kosyakova, N.; Sidorova, M.; Krasikova, A. Three-dimensional architecture of tandem repeats in chicken interphase nucleus. Chromosome Res. 2015, 23, 625–639. [Google Scholar] [CrossRef] [PubMed]
- Perry, B.W.; Schield, D.R.; Adams, R.H.; Castoe, T.A. Microchromosomes exhibit distinct features of vertebrate chromosome structure and function with underappreciated ramifications for genome evolution. Mol. Biol. Evol. 2021, 38, 904–910. [Google Scholar] [CrossRef]
- Waters, P.D.; Patel, H.R.; Ruiz-Herrera, A.; Álvarez-González, L.; Lister, N.C.; Simakov, O.; Ezaz, T.; Kaur, P.; Frere, C.; Grützner, F.; et al. Microchromosomes are building blocks of bird, reptile, and mammal chromosomes. Proc. Natl. Acad. Sci. USA 2021, 118, e2112494118. [Google Scholar] [CrossRef]
- Tanabe, H.; Müller, S.; Neusser, M.; von Hase, J.; Calcagno, E.; Cremer, M.; Solovei, I.; Cremer, C.; Cremer, T. Evolutionary conservation of chromosome territory arrangements in cell nuclei from higher primates. Proc. Natl. Acad. Sci. USA 2002, 99, 4424–4429. [Google Scholar] [CrossRef]
- Tanabe, H.; Küpper, K.; Ishida, T.; Neusser, M.; Mizusawa, H. Inter- and intra-specific gene-density-correlated radial chromosome territory arrangements are conserved in Old World monkeys. Cytogenet. Genome Res. 2005, 108, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Srikulnath, K.; Ahmad, S.F.; Singchat, W.; Panthum, T. Why Do Some Vertebrates Have Microchromosomes? Cells 2021, 10, 2182. [Google Scholar] [CrossRef]
- Mengden, G.A.; Stock, A.D. Chromosomal evolution in Serpentes: A comparison of G and C chromosome banding patterns of some Colubrid and Boid genera. Chromosoma 1980, 79, 53–64. [Google Scholar] [CrossRef]
- Olmo, E. Trends in the evolution of reptilian chromosomes. Integr. Comp. Biol. 2008, 48, 486–493. [Google Scholar] [CrossRef]
- Srikulnath, K.; Thapana, W.; Muangmai, N. Role of chromosome changes in Crocodylus evolution and diversity. Genom. Inform. 2015, 13, 102. [Google Scholar] [CrossRef]
- Cohen, M.M.; Gans, C. The chromosomes of the order Crocodilia. Cytogenetics 1970, 9, 81–105. [Google Scholar] [CrossRef]
- Pokorná, M.; Giovannotti, M.; Kratochvíl, L.; Caputo, V.; Olmo, E.; Ferguson-Smith, M.A.; Rens, W. Conservation of chromosomes syntenic with avian autosomes in squamate reptiles revealed by comparative chromosome painting. Chromosoma 2012, 121, 409–418. [Google Scholar] [CrossRef]
- King, M. Species Evolution: The Role of Chromosome Change; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Gregory, T.R. A bird’s-eye view of the C-value enigma: Genome size, cell size, and metabolic rate in the class aves. Evolution 2002, 56, 121–130. [Google Scholar] [CrossRef]
- Andrews, C.B.; Mackenzie, S.A.; Gregory, T.R. Genome size and wing parameters in passerine birds. Proc. Biol. Sci. 2009, 276, 55–61. [Google Scholar] [CrossRef]
- Janes, D.E.; Organ, C.L.; Fujita, M.K.; Shedlock, A.M.; Edwards, S.V. Genome evolution in Reptilia, the sister group of mammals. Annu. Rev. Genomics Hum. Genet. 2010, 11, 239–264. [Google Scholar] [CrossRef] [PubMed]
- Organ, C.L.; Shedlock, A.M.; Meade, A.; Pagel, M.; Edwards, S.V. Origin of avian genome size and structure in non-avian dinosaurs. Nature 2007, 446, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Organ, C.L.; Shedlock, A.M. Palaeogenomics of pterosaurs and the evolution of small genome size in flying vertebrates. Biol. Lett. 2009, 5, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.L.; Hughes, M.K. Small genomes for better flyers. Nature 1995, 377, 391. [Google Scholar] [CrossRef]
- Garner, J.P.; Taylor, G.K.; Thomas, A.L.R. On the origins of birds: The sequence of character acquisition in the evolution of avian flight. Proc. Biol. Sci. 1999, 266, 1259. [Google Scholar] [CrossRef]
- Ledesma, M.A.; de Freitas, T.R.O.; da Silva, J.; da Silva, F.; Gunski, R.J. Descripción cariotípica de Spheniscus magellanicus (Spheniscidae). El Hornero 2003, 18, 61–64. [Google Scholar] [CrossRef]
- Eichler, E.E.; Sankoff, D. Structural dynamics of eukaryotic chromosome evolution. Science 2003, 301, 793–797. [Google Scholar] [CrossRef] [PubMed]
- Farré, M.; Narayan, J.; Slavov, G.T.; Damas, J.; Auvil, L.; Li, C.; Jarvis, E.D.; Burt, D.W.; Griffin, D.K.; Larkin, D.M. Novel Insights into Chromosome Evolution in Birds, Archosaurs, and Reptiles. Genome Biol. Evol. 2016, 8, 2442–2451. [Google Scholar] [CrossRef] [PubMed]
- Jetz, W.; Thomas, G.H.; Joy, J.B.; Hartmann, K.; Mooers, A.O. The global diversity of birds in space and time. Nature 2012, 491, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Larkin, D.M.; Pape, G.; Donthu, R.; Auvil, L.; Welge, M.; Lewin, H.A. Breakpoint regions and homologous synteny blocks in chromosomes have different evolutionary histories. Genome Res. 2009, 19, 770–777. [Google Scholar] [CrossRef] [PubMed]
- Donthu, R.; Lewin, H.A.; Larkin, D.M. SyntenyTracker: A tool for defining homologous synteny blocks using radiation hybrid maps and whole-genome sequence. BMC Res. Notes 2009, 2, 148. [Google Scholar] [CrossRef] [PubMed]
- Farré, M.; Kim, J.; Proskuryakova, A.A.; Zhang, Y.; Kulemzina, A.I.; Li, Q.; Zhou, Y.; Xiong, Y.; Johnson, J.L.; Perelman, P.L.; et al. Evolution of gene regulation in ruminants differs between evolutionary breakpoint regions and homologous synteny blocks. Genome Res. 2019, 29, 576–589. [Google Scholar] [CrossRef] [PubMed]
- Damas, J.; Corbo, M.; Kim, J.; Turner-Maier, J.; Farré, M.; Larkin, D.M.; Ryder, O.A.; Steiner, C.; Houck, M.L.; Hall, S.; et al. Evolution of the ancestral mammalian karyotype and syntenic regions. Proc. Natl. Acad. Sci. USA 2022, 119, e2209139119. [Google Scholar] [CrossRef]
- Ruiz-Herrera, A.; Castresana, J.; Robinson, T.J. Is mammalian chromosomal evolution driven by regions of genome fragility? Gen. Biol. 2006, 7, R115. [Google Scholar] [CrossRef]
- Lewin, H.A.; Larkin, D.M.; Pontius, J.; O’Brien, S.J. Every genome sequence needs a good map. Genome Res. 2009, 19, 1925–1928. [Google Scholar] [CrossRef]
- Kim, J.; Farré, M.; Auvil, L.; Capitanu, B.; Larkin, D.M.; Ma, J.; Lewin, H.A. Reconstruction and evolutionary history of eutherian chromosomes. Proc. Natl. Acad. Sci. USA 2017, 114, E5379–E5388. [Google Scholar] [CrossRef]
- Capilla, L.; Sánchez-Guillén, R.A.; Farré, M.; Paytuví-Gallart, A.; Malinverni, R.; Ventura, J.; Larkin, D.M.; Ruiz-Herrera, A. Mammalian Comparative Genomics Reveals Genetic and Epigenetic Features Associated with Genome Reshuffling in Rodentia. Gen. Biol. Evol. 2016, 8, 3703–3717. [Google Scholar] [CrossRef][Green Version]
- Murphy, W.J.; Larkin, D.M.; Everts-van der Wind, A.; Bourque, G.; Tesler, G.; Auvil, L.; Beever, J.E.; Chowdhary, B.P.; Galibert, F.; Gatzke, L.; et al. Dynamics of mammalian chromosome evolution inferred from multispecies comparative maps. Science 2005, 309, 613–617. [Google Scholar] [CrossRef]
- Lerner, H.R.; Fleischer, R.C. Prospects for the use of next-generation sequencing methods in ornithology. Auk 2010, 127, 4–15. [Google Scholar] [CrossRef]
- Delmore, K.E.; Liedvogel, M. Avian population genomics taking off: Latest findings and future prospects. In Statistical Population Genomics; Dutheil, J.Y., Ed.; Methods in Molecular Biology; Humana: New York, NY, USA, 2020; Volume 2090, pp. 413–433. [Google Scholar] [CrossRef]
- Bravo, G.A.; Schmitt, C.J.; Edwards, S.V. What have we learned from the first 500 avian genomes? Annu. Rev. Ecol. Evol. Syst. 2021, 52, 611–639. [Google Scholar] [CrossRef]
- Raza, S.H.A.; Hassanin, A.A.; Pant, S.D.; Bing, S.; Sitohy, M.Z.; Abdelnour, S.A.; Alotaibi, M.A.; Al-Hazani, T.M.; Abd El-Aziz, A.H.; Cheng, G.; et al. Potentials, prospects and applications of genome editing technologies in livestock production. Saudi J. Biol. Sci. 2022, 29, 1928–1935. [Google Scholar] [CrossRef]
- Campagna, L.; Toews, D.P. The genomics of adaptation in birds. Curr. Biol. 2022, 32, R1173–R1186. [Google Scholar] [CrossRef] [PubMed]
- Ruvinskiy, D.; Larkin, D.M.; Farré, M.A. Near Chromosome Assembly of the Dromedary Camel Genome. Front. Genet. 2019, 10, 32. [Google Scholar] [CrossRef] [PubMed]
- Farré, M.; Li, Q.; Zhou, Y.; Damas, J.; Chemnick, L.G.; Kim, J.; Ryder, O.A.; Ma, J.; Zhang, G.; Larkin, D.M.; et al. A near-chromosome-scale genome assembly of the gemsbok (Oryx gazella): An iconic antelope of the Kalahari desert. GigaScience 2019, 8, giy162. [Google Scholar] [CrossRef]
- Farré, M.; Li, Q.; Darolti, I.; Zhou, Y.; Damas, J.; Proskuryakova, A.A.; Kulemzina, A.I.; Chemnick, L.G.; Kim, J.; Ryder, O.A.; et al. An integrated chromosome-scale genome assembly of the Masai giraffe (Giraffa camelopardalis tippelskirchi). GigaScience 2019, 8, giz090. [Google Scholar] [CrossRef] [PubMed]
- Secomandi, S.; Gallo, G.R.; Sozzoni, M.; Iannucci, A.; Galati, E.; Abueg, L.; Balacco, J.; Caprioli, M.; Chow, W.; Ciofi, C.; et al. A chromosome-level reference genome and pangenome for barn swallow population genomics. Cell Rep. 2023, 42, 111992. [Google Scholar] [CrossRef]
- Formenti, G.; Chiara, M.; Poveda, L.; Francoijs, K.J.; Bonisoli-Alquati, A.; Canova, L.; Gianfranceschi, L.; Horner, D.S.; Saino, N. SMRT long reads and Direct Label and Stain optical maps allow the generation of a high-quality genome assembly for the European barn swallow (Hirundo rustica rustica). Gigascience 2019, 8, giy142. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Jiang, X.; Li, B.; Wu, J.; Shen, J.; Xu, Z.; Zhou, X.; Hou, M.; Huang, Z.; Ou, X.; et al. A high-quality genome assembly highlights the evolutionary history of the great bustard (Otis tarda, Otidiformes). Commun. Biol. 2023, 6, 746. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, D.C. Optical Mapping: New Applications, Advances and Challenges. Gigascience. 2014. Available online: https://academic.oup.com/gigascience/pages/optical_mapping_new_applications_advances_challenges (accessed on 25 January 2024).
- Sutton, J.T.; Helmkampf, M.; Steiner, C.C.; Bellinger, M.R.; Korlach, J.; Hall, R.; Baybayan, P.; Muehling, J.; Gu, J.; Kingan, S.; et al. A high-quality, long-read de novo genome assembly to aid conservation of Hawaii’s last remaining crow species. Genes 2018, 9, 393. [Google Scholar] [CrossRef]
- Peona, V.; Weissensteiner, M.H.; Suh, A. How complete are “complete” genome assemblies?—An avian perspective. Mol. Ecol. Resour. 2018, 18, 1188–1195. [Google Scholar] [CrossRef]
- Huttener, R.; Thorrez, L.; Veld, T.I.T.; Granvik, M.; Van Lommel, L.; Waelkens, E.; Derua, R.; Lemaire, K.; Goyvaerts, L.; De Coster, S.; et al. Sequencing refractory regions in bird genomes are hotspots for accelerated protein evolution. BMC Ecol. Evol. 2021, 21, 176. [Google Scholar] [CrossRef] [PubMed]
- Korlach, J.; Gedman, G.; Kingan, S.B.; Chin, C.S.; Howard, J.T.; Audet, J.N.; Cantin, L.; Jarvis, E.D. De novo PacBio long-read and phased avian genome assemblies correct and add to reference genes generated with intermediate and short reads. Gigascience 2017, 6, gix085. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Minias, P.; Dunn, P.O. Long-read genome assemblies reveal extraordinary variation in the number and structure of MHC loci in birds. Genome Biol. Evol. 2021, 13, evaa270. [Google Scholar] [CrossRef]
- Wang, J.; Xi, Y.; Ma, S.; Qi, J.; Li, J.; Zhang, R.; Han, C.; Li, L.; Wang, J.; Liu, H. Single-molecule long-read sequencing reveals the potential impact of posttranscriptional regulation on gene dosage effects on the avian Z chromosome. BMC Genom. 2022, 23, 122. [Google Scholar] [CrossRef]
- Zhang, J.; Li, C.; Zhou, Q.; Zhang, G. Improving the ostrich genome assembly using optical mapping data. Gigascience 2015, 4, 24. [Google Scholar] [CrossRef]
- Lundberg, M.; Mackintosh, A.; Petri, A.; Bensch, S. Inversions maintain differences between migratory phenotypes of a songbird. Nat. Commun. 2023, 14, 452. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
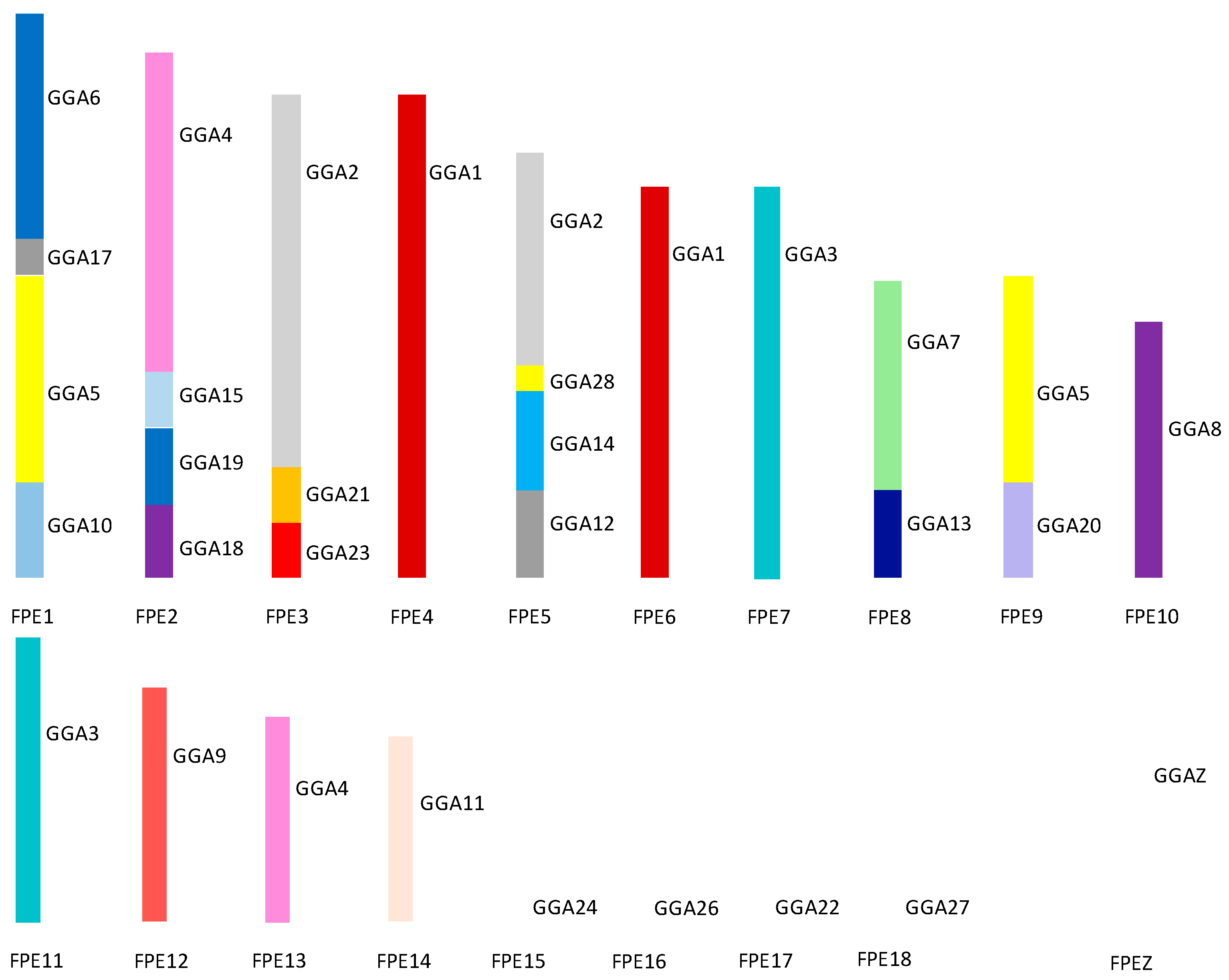{kind=link}
| Order | No. Species Examined | 2n | Fusions | Fissions | References |
|---|---|---|---|---|---|
| Struthioniformes | 1 | 80 | No | No | [78] |
| Rheiformes | 2 | 80 | No | No | [78,79] |
| Casuariiformes | 2 | 80 to 92 | No | No | [49,78] |
| Tinamiformes | 1 | 80 | No | No | [78] |
| Galliformes | 15 | 66 to 82 | Yes | Yes | [79,80,81,82] |
| Anseriformes | 8 | 80 to 98 | Yes | No | [76,79,83,84,85] |
| Apodiformes | 3 | 78 to 80 | Yes | No | [86] |
| Cuculiformes | 2 | 76 to 90 | Yes | Yes | [87] |
| Columbiformes | 11 | 76 to 86 | Yes | Yes | [69,79,88,89] |
| Gruiformes | 5 | 78 to 92 | Yes | Yes | [76,90,91] |
| Charadriiformes | 6 | 42 to 92 | Yes | Yes | [32,92,93,94,95,96] |
| Eurypygiformes | 1 | 80 | Yes | Yes | [97] |
| Ciconiiformes | 2 | 56 to 72 | Yes | No | [98] |
| Pelecaniformes | 6 | 62 to 74 | Yes | Yes | [99,100] |
| Opisthoconiformes | 1 | 80 | Yes | Yes | [101] |
| Accipitriformes | 15 | 58 to 80 | Yes | Yes | [73,102,103,104,105,106,107,108,109,110] |
| Strigiformes | 3 | 76 to 80 | Yes | Yes | [79,92,111] |
| Trogoniformes | 1 | 82 | Yes | Yes | [112] |
| Piciformes | 2 | 112 | Yes | Yes | [113] |
| Falconiformes | 3 | 40 to 52 | Yes | Yes | [77,114] |
| Psittaciformes | 10 | 48 to 72 | Yes | Yes | [115,116,117,118,119,120] |
| Passeriformes | 26 | 60 to 82 | Yes | Yes | [68,76,79,88,121,122,123,124,125,126,127,128,129] |
| Total | 126 |
| Order | Common Name | Species Name | References |
|---|---|---|---|
| Anseriformes | Greylag goose | Anser anser | [79] |
| Galliformes | Chinese bamboo-partridge | Bambusicola thoracica | [81] |
| Galliformes | Chinese quail | Coturnix chinensis | [81] |
| Galliformes | Common peafowl | Pavo cristatus | [81] |
| Galliformes | Chicken | Gallus gallus | [88] |
| Galliformes | Japanese quail | Coturnix japonica | [79,81] |
| Columbiformes | African collared dove | Streptopelia roseogrisea | [79] |
| Order | No. Species Examined | 2n | Fusions | References |
|---|---|---|---|---|
| Struthioniformes | 1 | 80 | No | [64] |
| Galliformes | 6 | 78 to 80 | No | [64] |
| Anseriformes | 1 | 80 | No | [64] |
| Caprimulgiformes | 1 | 74 | Yes | [140] |
| Otidiformes | 1 | 76 | No | [64] |
| Cuculiformes | 1 | 74 | Yes | [63] |
| Columbiformes | 6 | 76 to 86 | No | [64,89] |
| Gruiformes | 1 | 92 | No | [91] |
| Charadriiformes | 4 | 78 to 96 | Yes | [64,141] |
| Suliformes | 1 | 74 | Yes | [140] |
| Accipitriformes | 1 | 66 | Yes | [110] |
| Strigiformes | 1 | 80 | No | [64] |
| Trogoniformes | 1 | 82 | No | [140] |
| Piciformes | 2 | 112 | No | [140] |
| Falconiformes | 3 | 50 to 52 | Yes | [114,138] |
| Psittaciformes | 4 | 48 to 72 | Yes | [64,119,138] |
| Passeriformes | 7 | 60 to 80 | Yes | [64,142,143,144,145] |
| Total | 42 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Connor, R.E.; Kretschmer, R.; Romanov, M.N.; Griffin, D.K. A Bird’s-Eye View of Chromosomic Evolution in the Class Aves. Cells 2024, 13, 310. https://doi.org/10.3390/cells13040310
O’Connor RE, Kretschmer R, Romanov MN, Griffin DK. A Bird’s-Eye View of Chromosomic Evolution in the Class Aves. Cells. 2024; 13(4):310. https://doi.org/10.3390/cells13040310
Chicago/Turabian StyleO’Connor, Rebecca E., Rafael Kretschmer, Michael N. Romanov, and Darren K. Griffin. 2024. "A Bird’s-Eye View of Chromosomic Evolution in the Class Aves" Cells 13, no. 4: 310. https://doi.org/10.3390/cells13040310
APA StyleO’Connor, R. E., Kretschmer, R., Romanov, M. N., & Griffin, D. K. (2024). A Bird’s-Eye View of Chromosomic Evolution in the Class Aves. Cells, 13(4), 310. https://doi.org/10.3390/cells13040310