

Cilia Provide a Platform for the Generation, Regulated Secretion, and Reception of Peptidergic Signals


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
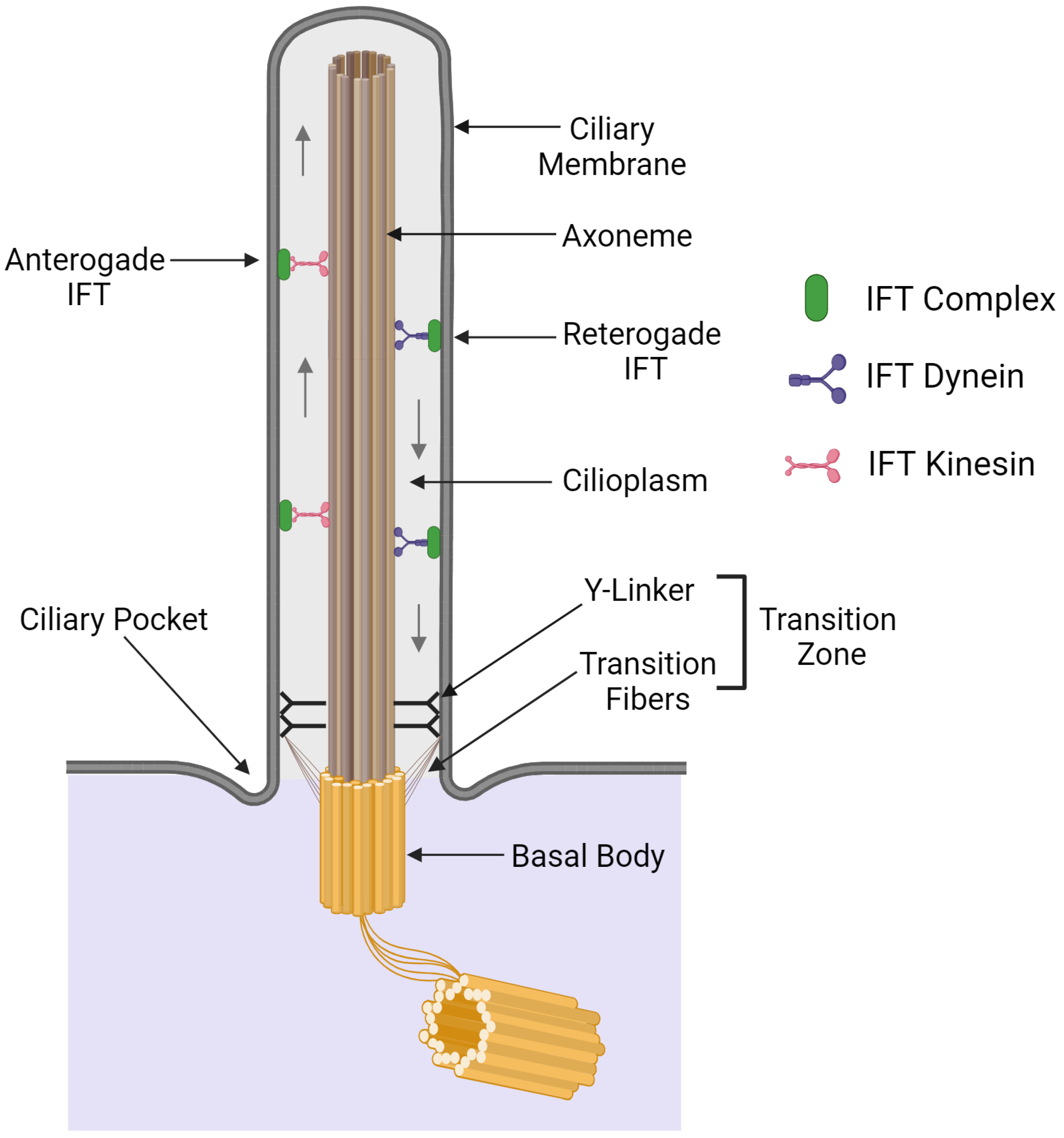
2. General Organization of Cilia
3. Cilia Serve as Specialized Signaling Organelles
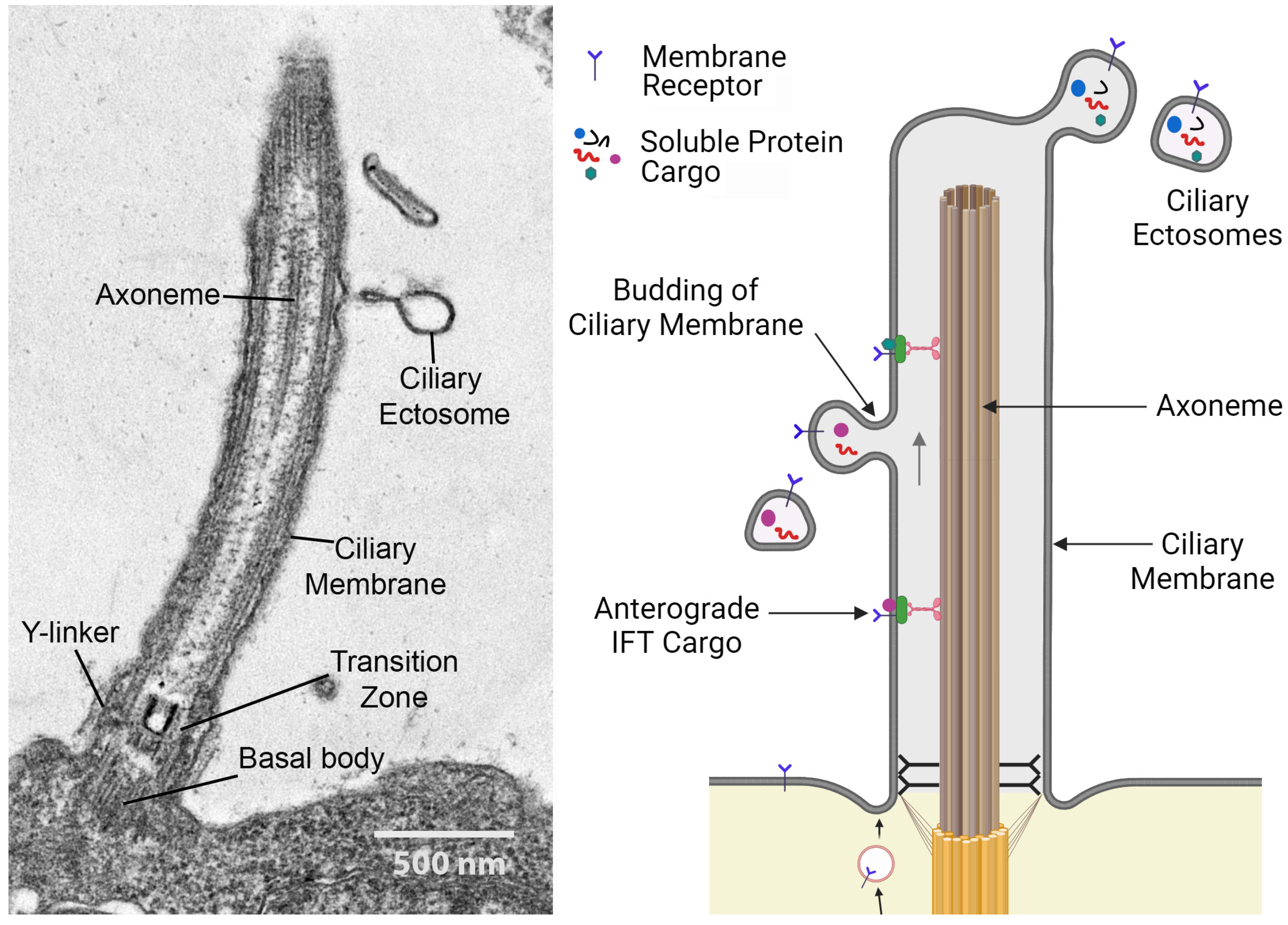
4. Cilia as Secretory Organelles
5. Cilia as a Platform for Peptidergic Signaling
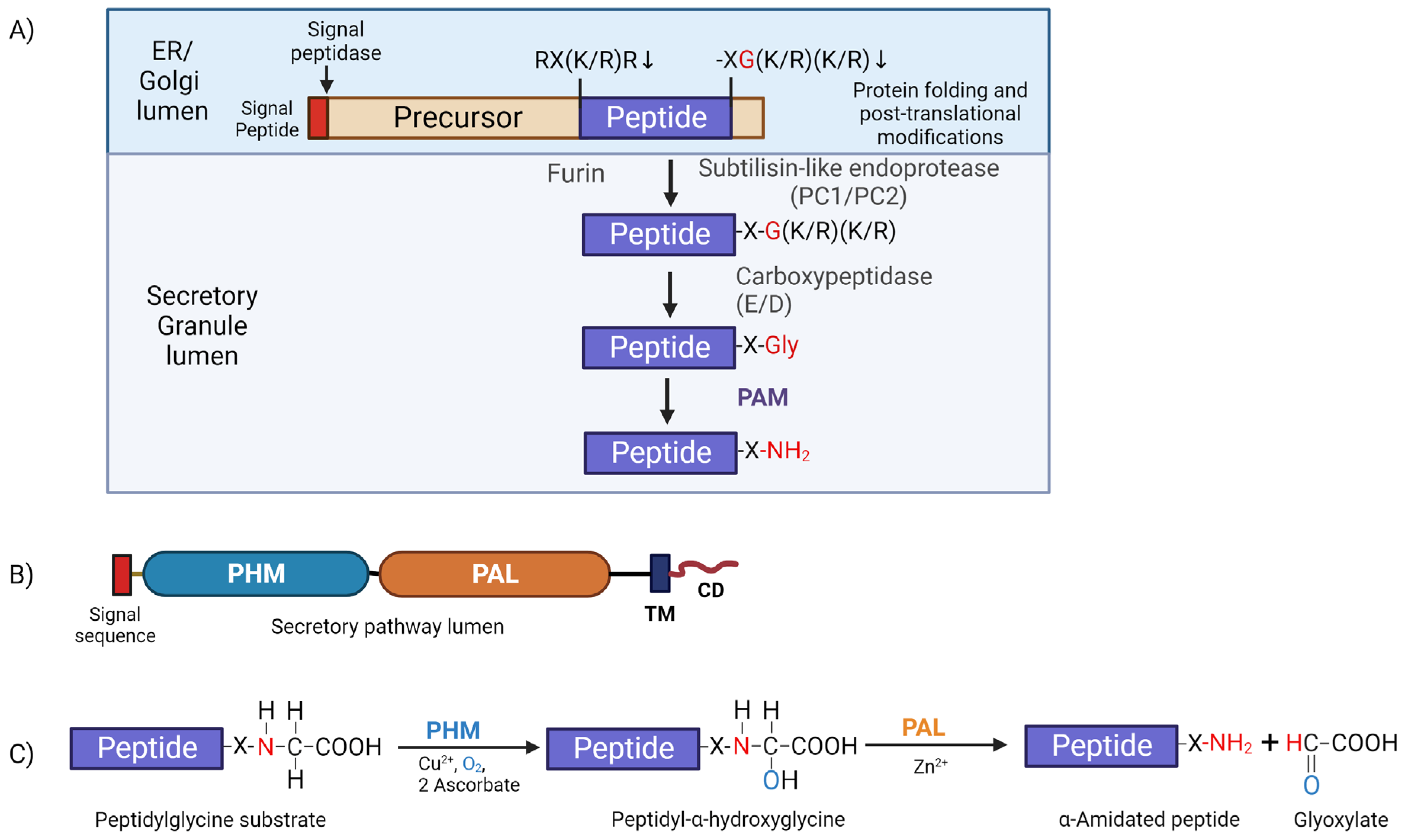
6. The Peptide Amidation Pathway
7. Regulated Processing and Secretion of Peptidergic Signals through Cilia
8. Chlamydomonas Has the Machinery Needed to Generate, Secrete, and Detect Peptidergic Signals
9. Ciliary Localization of the Peptide Amidating Enzyme (PAM)
10. Regulated Trafficking of PAM, Amidated Products, and a Prohormone Convertase into Ciliary Ectosomes
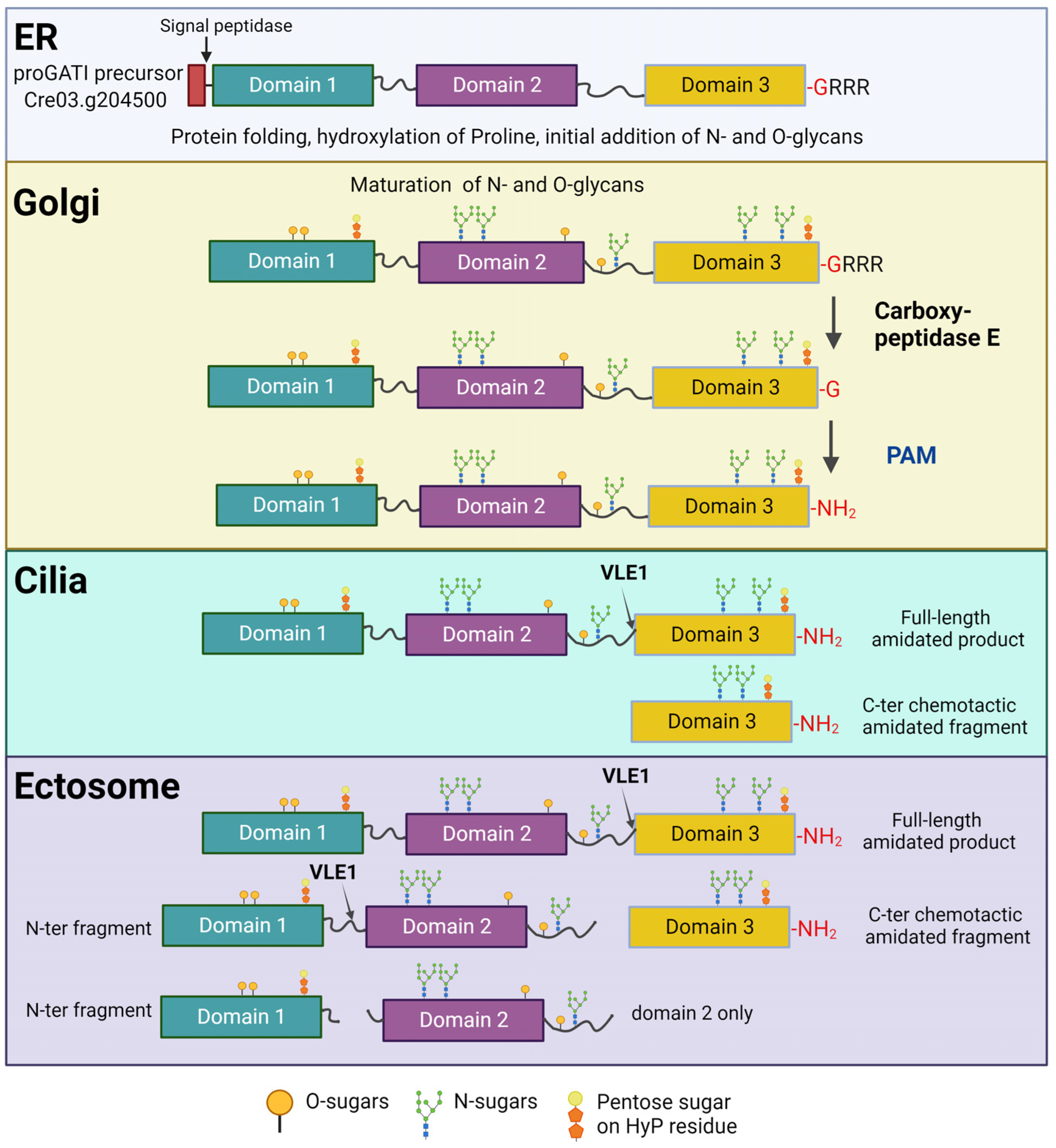
11. The proGATI Precursor Is Heavily Glycosylated
12. Endo-Proteolytic Cleavage of proGATI Is Triggered by Its Ectosomal Entry
13. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carvalho-Santos, Z.; Azimzadeh, J.; Pereira-Leal, J.B.; Bettencourt-Dias, M. Evolution: Tracing the origins of centrioles, cilia, and flagella. J. Cell Biol. 2011, 194, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Malicki, J.J.; Johnson, C.A. The cilium: Cellular antenna and central processing unit. Trends Cell Biol. 2017, 27, 126–140. [Google Scholar] [CrossRef]
- Wood, C.R.; Huang, K.; Diener, D.R.; Rosenbaum, J.L. The cilium secretes bioactive ectosomes. Curr. Biol. 2013, 23, 906–911. [Google Scholar] [CrossRef]
- Wang, J.; Nikonorova, I.A.; Silva, M.; Walsh, J.D.; Tilton, P.E.; Gu, A.; Akella, J.S.; Barr, M.M. Sensory cilia act as a specialized venue for regulated extracellular vesicle biogenesis and signaling. Curr. Biol. 2021, 31, 3943–3951.e3943. [Google Scholar] [CrossRef] [PubMed]
- Luxmi, R.; Kumar, D.; Mains, R.E.; King, S.M.; Eipper, B.A. Cilia-based peptidergic signaling. PLoS Biol. 2019, 17, e3000566. [Google Scholar] [CrossRef]
- Wang, J.; Silva, M.; Haas, L.A.; Morsci, N.S.; Nguyen, K.C.Q.; Hall, D.H.; Barr, M.M. C. elegans ciliated sensory neurons release extracellular vesicles that function in animal communication. Curr. Biol. 2014, 24, 519–525. [Google Scholar] [CrossRef]
- Wang, J.; Barr, M.M. Cell–cell communication via ciliary extracellular vesicles: Clues from model systems. Essays Biochem. 2018, 62, 205–213. [Google Scholar] [CrossRef]
- Cai, Y.; Fedeles, S.V.; Dong, K.; Anyatonwu, G.; Onoe, T.; Mitobe, M.; Gao, J.D.; Okuhara, D.; Tian, X.; Gallagher, A.R.; et al. Altered trafficking and stability of polycystins underlie polycystic kidney disease. J. Clin. Investig. 2014, 124, 5129–5144. [Google Scholar] [CrossRef]
- Volz, A.K.; Frei, A.; Kretschmer, V.; de Jesus Domingues, A.M.; Ketting, R.F.; Ueffing, M.; Boldt, K.; Krämer-Albers, E.M.; May-Simera, H.L. Bardet-Biedl syndrome proteins modulate the release of bioactive extracellular vesicles. Nat. Commun. 2021, 12, 5671. [Google Scholar] [CrossRef] [PubMed]
- Steiner, D.F. On the discovery of precursor processing. Methods Mol. Biol. 2011, 768, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Molina, B.; Martin, M.G.; Lindberg, I. Chapter Two—PCSK1 variants and human obesity. In Progress in Molecular Biology and Translational Science; Tao, Y.-X., Ed.; Academic Press: Cambridge, MA, USA, 2016; Volume 140, pp. 47–74. [Google Scholar]
- Händel, M.; Schulz, S.; Stanarius, A.; Schreff, M.; Erdtmann-Vourliotis, M.; Schmidt, H.; Wolf, G.; Höllt, V. Selective targeting of somatostatin receptor 3 to neuronal cilia. Neuroscience 1999, 89, 909–926. [Google Scholar] [CrossRef]
- Loktev, A.V.; Jackson, P.K. Neuropeptide Y family receptors traffic via the Bardet-Biedl syndrome pathway to signal in neuronal primary cilia. Cell Rep. 2013, 5, 1316–1329. [Google Scholar] [CrossRef]
- Choi, H.; Shin, J.H.; Kim, E.S.; Park, S.J.; Bae, I.H.; Jo, Y.K.; Jeong, I.Y.; Kim, H.J.; Lee, Y.; Park, H.C.; et al. Primary cilia negatively regulate melanogenesis in melanocytes and pigmentation in a human skin model. PLoS ONE 2016, 11, e0168025. [Google Scholar] [CrossRef]
- Czyzyk, T.A.; Ning, Y.; Hsu, M.-S.; Peng, B.; Mains, R.E.; Eipper, B.A.; Pintar, J.E. Deletion of peptide amidation enzymatic activity leads to edema and embryonic lethality in the mouse. Dev. Biol. 2005, 287, 301–313. [Google Scholar] [CrossRef]
- Wheway, G.; Nazlamova, L.; Hancock, J.T. Signaling through the primary cilium. Front. Cell Dev. Biol. 2018, 6, 8. [Google Scholar] [CrossRef]
- Bradbury, A.F.; Finnie, M.D.; Smyth, D.G. Mechanism of C-terminal amide formation by pituitary enzymes. Nature 1982, 298, 686–688. [Google Scholar] [CrossRef]
- Kumar, D.; Mains, R.E.; Eipper, B.A. 60 YEARS OF POMC: From POMC and α-MSH to PAM, molecular oxygen, copper, and vitamin C. J. Mol. Endocrinol. 2016, 56, T63–T76. [Google Scholar] [CrossRef] [PubMed]
- Luxmi, R.; Mains, R.E.; King, S.M.; Eipper, B.A. Amino Acids|Peptidylglycine α-amidating monooxygenase (PAM). In Encyclopedia of Biological Chemistry III (Third Edition); Jez, J., Ed.; Elsevier: Oxford, UK, 2021; pp. 88–104. [Google Scholar]
- Delgado-Prudencio, G.; Possani, L.D.; Becerril, B.; Ortiz, E. The dual α-amidation system in scorpion venom glands. Toxins 2019, 11, 425. [Google Scholar] [CrossRef]
- Ul-Hasan, S.; Burgess, D.M.; Gajewiak, J.; Li, Q.; Hu, H.; Yandell, M.; Olivera, B.M.; Bandyopadhyay, P.K. Characterization of the peptidylglycine α-amidating monooxygenase (PAM) from the venom ducts of neogastropods, Conus bullatus and Conus geographus. Toxicon 2013, 74, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Kreil, G. Structure and multistep activation of the precursors of peptides from honeybee venom glands and frog skin. Curr. Top. Cell Regul. 1984, 24, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Varoqueaux, F.; Williams, E.A.; Grandemange, S.; Truscello, L.; Kamm, K.; Schierwater, B.; Jékely, G.; Fasshauer, D. High cell diversity and complex peptidergic signaling underlie placozoan behavior. Curr. Biol. 2018, 28, 3495–3501.e3492. [Google Scholar] [CrossRef]
- Taghert, P.H.; Nitabach, M.N. Peptide neuromodulation in invertebrate model systems. Neuron 2012, 76, 82–97. [Google Scholar] [CrossRef]
- Elphick, M.R.; Thorndyke, M.C. Molecular characterisation of SALMFamide neuropeptides in sea urchins. J. Exp. Biol. 2005, 208, 4273–4282. [Google Scholar] [CrossRef] [PubMed]
- Conzelmann, M.; Williams, E.A.; Krug, K.; Franz-Wachtel, M.; Macek, B.; Jékely, G. The neuropeptide complement of the marine annelid Platynereis dumerilii. BMC Genom. 2013, 14, 906. [Google Scholar] [CrossRef]
- Kumar, D.; Blaby-Haas, C.E.; Merchant, S.S.; Mains, R.E.; King, S.M.; Eipper, B.A. Early eukaryotic origins for cilia-associated bioactive peptide-amidating activity. J. Cell Sci. 2016, 129, 943–956. [Google Scholar] [CrossRef] [PubMed]
- Luxmi, R.; Mains, R.E.; Eipper, B.A.; King, S.M. Regulated processing and secretion of a peptide precursor in cilia. Proc. Natl. Acad. Sci. USA 2022, 119, e2206098119. [Google Scholar] [CrossRef]
- Marshall, R.B.W. Cilia; Cold Spring Harb. Lab.: Cold Spring Harbor, NY, USA, 2017. [Google Scholar]
- Vincensini, L.; Blisnick, T.; Bastin, P. 1001 model organisms to study cilia and flagella. Biol. Cell 2011, 103, 109–130. [Google Scholar] [CrossRef]
- Feistel, K.; Blum, M. Three types of cilia including a novel 9+4 axoneme on the notochordal plate of the rabbit embryo. Dev. Dyn. 2006, 235, 3348–3358. [Google Scholar] [CrossRef] [PubMed]
- Gluenz, E.; Höög, J.L.; Smith, A.E.; Dawe, H.R.; Shaw, M.K.; Gull, K. Beyond 9+0: Noncanonical axoneme structures characterize sensory cilia from protists to humans. FASEB J. 2010, 24, 3117–3121. [Google Scholar] [CrossRef]
- Prensier, G.; Vivier, E.; Goldstein, S.; Schrével, J. Motile flagellum with a “3 + 0” ultrastructure. Science 1980, 207, 1493–1494. [Google Scholar] [CrossRef]
- Cavalier-Smith, T.; Chao, E.E.Y. Phylogeny of choanozoa, apusozoa, and other protozoa and early eukaryote megaevolution. J. Mol. Evol. 2003, 56, 540–563. [Google Scholar] [CrossRef] [PubMed]
- King, S.M. Axonemal dynein arms. Cold Spring Harb. Perspect. Biol. 2016, 8, a028100. [Google Scholar] [CrossRef] [PubMed]
- Lee, L. Mechanisms of mammalian ciliary motility: Insights from primary ciliary dyskinesia genetics. Gene 2011, 473, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, K.; Chen, D.; Nishida, T.; Misaki, K.; Yonemura, S.; Hamada, H. Absence of radial spokes in mouse node cilia is required for rotational movement but confers ultrastructural instability as a trade-off. Dev. Cell 2015, 35, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Djenoune, L.; Mahamdeh, M.; Truong, T.V.; Nguyen, C.T.; Fraser, S.E.; Brueckner, M.; Howard, J.; Yuan, S. Cilia function as calcium-mediated mechanosensors that instruct left-right asymmetry. Science 2023, 379, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Kiesel, P.; Alvarez Viar, G.; Tsoy, N.; Maraspini, R.; Gorilak, P.; Varga, V.; Honigmann, A.; Pigino, G. The molecular structure of mammalian primary cilia revealed by cryo-electron tomography. Nat. Struct. Mol. Biol. 2020, 27, 1115–1124. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Marshall, W.F. Intraflagellar transport and ciliary dynamics. Cold Spring Harb. Perspect. Biol. 2017, 9, a021998. [Google Scholar] [CrossRef] [PubMed]
- Pigino, G. Intraflagellar transport. Curr. Biol. 2021, 31, R530–R536. [Google Scholar] [CrossRef]
- Pazour, G.J.; Dickert, B.L.; Vucica, Y.; Seeley, E.S.; Rosenbaum, J.L.; Witman, G.B.; Cole, D.G. Chlamydomonas IFT88 and Its mouse homologue, polycystic kidney disease gene Tg737, are required for assembly of cilia and flagella. J. Cell Biol. 2000, 151, 709–718. [Google Scholar] [CrossRef]
- Rosenbaum, J.L.; Witman, G.B. Intraflagellar transport. Nat. Rev. Mol. Cell Biol. 2002, 3, 813–825. [Google Scholar] [CrossRef]
- Reiter, J.F.; Blacque, O.E.; Leroux, M.R. The base of the cilium: Roles for transition fibres and the transition zone in ciliary formation, maintenance and compartmentalization. EMBO Rep. 2012, 13, 608–618. [Google Scholar] [CrossRef] [PubMed]
- Weiss, R.L.; Goodenough, D.A.; Goodenough, U.W. Membrane particle arrays associated with the basal body and with contractile vacuole secretion in Chlamydomonas. J. Cell Biol. 1977, 72, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Garcia, G.; Raleigh, D.R.; Reiter, J.F. How the ciliary membrane is organized inside-out to communicate outside-in. Curr. Biol. 2018, 28, R421–R434. [Google Scholar] [CrossRef] [PubMed]
- Reiter, J.F.; Leroux, M.R. Genes and molecular pathways underpinning ciliopathies. Nat. Rev. Mol. Cell Biol. 2017, 18, 533–547. [Google Scholar] [CrossRef] [PubMed]
- Vertii, A.; Bright, A.; Delaval, B.; Hehnly, H.; Doxsey, S. New frontiers: Discovering cilia-independent functions of cilia proteins. EMBO Rep. 2015, 16, 1275–1287. [Google Scholar] [CrossRef] [PubMed]
- Jensen, V.L.; Leroux, M.R. Gates for soluble and membrane proteins, and two trafficking systems (IFT and LIFT), establish a dynamic ciliary signaling compartment. Curr. Opin. Cell Biol. 2017, 47, 83–91. [Google Scholar] [CrossRef]
- Amack, J.D. Structures and functions of cilia during vertebrate embryo development. Mol. Reprod. Dev. 2022, 89, 579–596. [Google Scholar] [CrossRef]
- Mykytyn, K.; Askwith, C. G-protein-coupled receptor signaling in cilia. Cold Spring Harb. Perspect. Biol. 2017, 9, a028183. [Google Scholar] [CrossRef]
- Nager, A.R.; Goldstein, J.S.; Herranz-Pérez, V.; Portran, D.; Ye, F.; Garcia-Verdugo, J.M.; Nachury, M.V. An actin network dispatches ciliary GPCRs into extracellular vesicles to modulate signaling. Cell 2017, 168, 252–263.e214. [Google Scholar] [CrossRef]
- Pala, R.; Alomari, N.; Nauli, S.M. Primary cilium-dependent signaling mechanisms. Int. J. Mol. Sci. 2017, 18, 2272. [Google Scholar] [CrossRef]
- Mao, S.; Shah, A.S.; Moninger, T.O.; Ostedgaard, L.S.; Lu, L.; Tang, X.X.; Thornell, I.M.; Reznikov, L.R.; Ernst, S.E.; Karp, P.H.; et al. Motile cilia of human airway epithelia contain hedgehog signaling components that mediate noncanonical hedgehog signaling. Proc. Natl. Acad. Sci. USA 2018, 115, 1370–1375. [Google Scholar] [CrossRef] [PubMed]
- Tözser, J.; Earwood, R.; Kato, A.; Brown, J.; Tanaka, K.; Didier, R.; Megraw, T.L.; Blum, M.; Kato, Y. TGF-β signaling regulates the differentiation of motile cilia. Cell Rep. 2015, 11, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.N.; Kaiser, F.; Klausen, C.; Stüven, B.; Chong, R.; Bönigk, W.; Mick, D.U.; Möglich, A.; Jurisch-Yaksi, N.; Schmidt, F.I.; et al. Nanobody-directed targeting of optogenetic tools to study signaling in the primary cilium. eLife 2020, 9, e57907. [Google Scholar] [CrossRef] [PubMed]
- Wachten, D.; Mick, D.U. Signal transduction in primary cilia—Analyzing and manipulating GPCR and second messenger signaling. Pharmacol. Ther. 2021, 224, 107836. [Google Scholar] [CrossRef] [PubMed]
- Vinay, L.; Belleannée, C. EV duty vehicles: Features and functions of ciliary extracellular vesicles. Front. Genet. 2022, 13, 916233. [Google Scholar] [CrossRef] [PubMed]
- Woith, E.; Fuhrmann, G.; Melzig, M.F. Extracellular vesicles—Connecting kingdoms. Int. J. Mol. Sci. 2019, 20, 5695. [Google Scholar] [CrossRef]
- Mantel, P.-Y.; Marti, M. The role of extracellular vesicles in Plasmodium and other protozoan parasites. Cell. Microbiol. 2014, 16, 344–354. [Google Scholar] [CrossRef]
- Long, H.; Zhang, F.; Xu, N.; Liu, G.; Diener, D.R.; Rosenbaum, J.L.; Huang, K. Comparative analysis of ciliary membranes and ectosomes. Curr. Biol. 2016, 26, 3327–3335. [Google Scholar] [CrossRef]
- Skotland, T.; Sagini, K.; Sandvig, K.; Llorente, A. An emerging focus on lipids in extracellular vesicles. Adv. Drug Deliv. Rev. 2020, 159, 308–321. [Google Scholar] [CrossRef] [PubMed]
- Abels, E.R.; Breakefield, X.O. Introduction to extracellular vesicles: Biogenesis, RNA cargo selection, content, release, and uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Malkin, E.Z.; Bratman, S.V. Bioactive DNA from extracellular vesicles and particles. Cell Death Dis. 2020, 11, 584. [Google Scholar] [CrossRef] [PubMed]
- Amintas, S.; Vendrely, V.; Dupin, C.; Buscail, L.; Laurent, C.; Bournet, B.; Merlio, J.-P.; Bedel, A.; Moreau-Gaudry, F.; Boutin, J.; et al. Next-generation cancer biomarkers: Extracellular vesicle DNA as a circulating surrogate of tumor DNA. Front. Cell Dev. Biol. 2021, 8, 622048. [Google Scholar] [CrossRef] [PubMed]
- Luxmi, R.; King, S.M. Cilia-derived vesicles: An ancient route for intercellular communication. Semin. Cell Dev. Biol. 2022, 129, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Desai, P.B.; Stuck, M.W.; Lv, B.; Pazour, G.J. Ubiquitin links smoothened to intraflagellar transport to regulate Hedgehog signaling. J. Cell Biol. 2020, 219, e201912104. [Google Scholar] [CrossRef] [PubMed]
- Shinde, S.R.; Nager, A.R.; Nachury, M.V. Ubiquitin chains earmark GPCRs for BBSome-mediated removal from cilia. J. Cell Biol. 2020, 219, e202003020. [Google Scholar] [CrossRef] [PubMed]
- Ojeda Naharros, I.; Nachury, M.V. Shedding of ciliary vesicles at a glance. J. Cell Sci. 2022, 135, jcs246553. [Google Scholar] [CrossRef] [PubMed]
- Brailov, I.; Bancila, M.; Brisorgueil, M.J.; Miquel, M.C.; Hamon, M.; Vergé, D. Localization of 5-HT(6) receptors at the plasma membrane of neuronal cilia in the rat brain. Brain Res. 2000, 872, 271–275. [Google Scholar] [CrossRef]
- Abdul-Majeed, S.; Nauli, S.M. Dopamine receptor type 5 in the primary cilia has dual chemo- and mechano-sensory roles. Hypertension 2011, 58, 325–331. [Google Scholar] [CrossRef]
- Domire, J.S.; Green, J.A.; Lee, K.G.; Johnson, A.D.; Askwith, C.C.; Mykytyn, K. Dopamine receptor 1 localizes to neuronal cilia in a dynamic process that requires the Bardet-Biedl syndrome proteins. Cell Mol. Life Sci. 2011, 68, 2951–2960. [Google Scholar] [CrossRef]
- Wang, Y.; Bernard, A.; Comblain, F.; Yue, X.; Paillart, C.; Zhang, S.; Reiter, J.F.; Vaisse, C. Melanocortin 4 receptor signals at the neuronal primary cilium to control food intake and body weight. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef]
- Berbari, N.F.; Malarkey, E.B.; Yazdi, S.M.; McNair, A.D.; Kippe, J.M.; Croyle, M.J.; Kraft, T.W.; Yoder, B.K. Hippocampal and cortical primary cilia are required for aversive memory in mice. PLoS ONE 2014, 9, e106576. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Wen, X.; Ratti, N.; Loktev, A.; Rangell, L.; Scales, S.J.; Jackson, P.K. The ciliary G-protein-coupled receptor Gpr161 negatively regulates the sonic hedgehog pathway via cAMP signaling. Cell 2013, 152, 210–223. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Wen, X.; Scales, S.J. The orphan G protein-coupled receptor Gpr175 (Tpra40) enhances hedgehog signaling by modulating cAMP levels. J. Biol. Chem. 2015, 290, 29663–29675. [Google Scholar] [CrossRef]
- Herzog, H. Neuropeptide Y and energy homeostasis: Insights from Y receptor knockout models. Eur. J. Pharmacol. 2003, 480, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Tereshko, L.; Gao, Y.; Cary, B.A.; Turrigiano, G.G.; Sengupta, P. Ciliary neuropeptidergic signaling dynamically regulates excitatory synapses in postnatal neocortical pyramidal neurons. eLife 2021, 10, e65427. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, C.; Stone, K.L.; Francone, V.P.; Mains, R.E.; Eipper, B.A. Secretory granule to the nucleus: Role of a multiply phosphorylated intrinsically unstructured domain. J. Biol. Chem. 2009, 284, 25723–25734. [Google Scholar] [CrossRef]
- Bäck, N.; Luxmi, R.; Powers, K.G.; Mains, R.E.; Eipper, B.A. Peptidylglycine α-amidating monooxygenase is required for atrial secretory granule formation. Proc. Natl. Acad. Sci. USA 2020, 117, 17820–17831. [Google Scholar] [CrossRef]
- Halban, P.A. Structural domains and molecular lifestyles of insulin and its precursors in the pancreatic beta cell. Diabetologia 1991, 34, 767–778. [Google Scholar] [CrossRef]
- Luxmi, R.; Blaby-Haas, C.; Kumar, D.; Rauniyar, N.; King, S.M.; Mains, R.E.; Eipper, B.A. Proteases shape the Chlamydomonas Secretome: Comparison to classical neuropeptide processing machinery. Proteomes 2018, 6, 36. [Google Scholar] [CrossRef]
- Spudich, J.L.; Sager, R. Regulation of the Chlamydomonas cell cycle by light and dark. J. Cell Biol. 1980, 85, 136–145. [Google Scholar] [CrossRef]
- Goodenough, U.; Lin, H.; Lee, J.-H. Sex determination in Chlamydomonas. Semin. Cell Dev. Biol. 2007, 18, 350–361. [Google Scholar] [CrossRef]
- Elrick, M.M.; Samson, W.K.; Corbett, J.A.; Salvatori, A.S.; Stein, L.M.; Kolar, G.R.; Naatz, A.; Yosten, G.L.C. Neuronostatin acts via GPR107 to increase cAMP-independent PKA phosphorylation and proglucagon mRNA accumulation in pancreatic α-cells. Am. J. Physiol—Regul. Integr. Comp. Physiol. 2016, 310, R143–R155. [Google Scholar] [CrossRef]
- Edgar, A. Human GPR107 and murine Gpr108 are members of the LUSTR family of proteins found in both plants and animals, having similar topology to G-protein coupled receptors. DNA Seq. 2007, 18, 235–241. [Google Scholar] [CrossRef]
- Huang, K.; Diener, D.R.; Mitchell, A.; Pazour, G.J.; Witman, G.B.; Rosenbaum, J.L. Function and dynamics of PKD2 in Chlamydomonas reinhardtii flagella. J. Cell Biol. 2007, 179, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Strenkert, D.; Patel-King, R.S.; Leonard, M.T.; Merchant, S.S.; Mains, R.E.; King, S.M.; Eipper, B.A. A bioactive peptide amidating enzyme is required for ciliogenesis. Elife 2017, 6, e25728. [Google Scholar] [CrossRef]
- Kumar, D.; Thomason, R.T.; Yankova, M.; Gitlin, J.D.; Mains, R.E.; Eipper, B.A.; King, S.M. Microvillar and ciliary defects in zebrafish lacking an actin-binding bioactive peptide amidating enzyme. Sci. Rep. 2018, 8, 4547. [Google Scholar] [CrossRef]
- Kumar, D.; Mains, R.E.; Eipper, B.A.; King, S.M. Ciliary and cytoskeletal functions of an ancient monooxygenase essential for bioactive amidated peptide synthesis. Cell. Mol. Life Sci. 2019, 76, 2329–2348. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Hayakawa, E.; Koizumi, O.; Fujisawa, T. Neuropeptides and their functions in Hydra. Acta Biol. Hung. 2008, 59, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Palma, M.S. Insect venom peptides. In Handbook of Biologically Active Peptides; Kastin, A.J., Ed.; Academic Press: Burlington, MA, USA, 2006; pp. 389–396. [Google Scholar]
- Szabó, R.; Láng, O.; Láng, J.; Illyés, E.; Kőhidai, L.; Hudecz, F. Effect of SXWS/WSXWS peptides on chemotaxis and adhesion of the macrophage-like cell line J774. J. Mol. Recognit. 2015, 28, 253–260. [Google Scholar] [CrossRef]
- Bollig, K.; Lamshöft, M.; Schweimer, K.; Marner, F.J.; Budzikiewicz, H.; Waffenschmidt, S. Structural analysis of linear hydroxyproline-bound O-glycans of Chlamydomonas reinhardtii—Conservation of the inner core in Chlamydomonas and land plants. Carbohydr. Res. 2007, 342, 2557–2566. [Google Scholar] [CrossRef]
- Tan, L.; Leykam, J.F.; Kieliszewski, M.J. Glycosylation motifs that direct arabinogalactan addition to arabinogalactan-proteins. Plant Physiol. 2003, 132, 1362–1369. [Google Scholar] [CrossRef] [PubMed]
- Madsen, T.D.; Hansen, L.H.; Hintze, J.; Ye, Z.; Jebari, S.; Andersen, D.B.; Joshi, H.J.; Ju, T.; Goetze, J.P.; Martin, C.; et al. An atlas of O-linked glycosylation on peptide hormones reveals diverse biological roles. Nat. Commun. 2020, 11, 4033. [Google Scholar] [CrossRef] [PubMed]
- Rouillé, Y.; Duguay, S.J.; Lund, K.; Furuta, M.; Gong, Q.; Lipkind, G.; Oliva, A.A., Jr.; Chan, S.J.; Steiner, D.F. Proteolytic processing mechanisms in the biosynthesis of neuroendocrine peptides: The subtilisin-like proprotein convertases. Front. Neuroendocrinol. 1995, 16, 322–361. [Google Scholar] [CrossRef] [PubMed]
- Sakato-Antoku, M.; King, S.M. Developmental changes in ciliary composition during gametogenesis in Chlamydomonas. Mol. Biol. Cell 2022, 33, br10. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luxmi, R.; King, S.M. Cilia Provide a Platform for the Generation, Regulated Secretion, and Reception of Peptidergic Signals. Cells 2024, 13, 303. https://doi.org/10.3390/cells13040303
Luxmi R, King SM. Cilia Provide a Platform for the Generation, Regulated Secretion, and Reception of Peptidergic Signals. Cells. 2024; 13(4):303. https://doi.org/10.3390/cells13040303
Chicago/Turabian StyleLuxmi, Raj, and Stephen M. King. 2024. "Cilia Provide a Platform for the Generation, Regulated Secretion, and Reception of Peptidergic Signals" Cells 13, no. 4: 303. https://doi.org/10.3390/cells13040303
APA StyleLuxmi, R., & King, S. M. (2024). Cilia Provide a Platform for the Generation, Regulated Secretion, and Reception of Peptidergic Signals. Cells, 13(4), 303. https://doi.org/10.3390/cells13040303