
Postnatal Dynamic Ciliary ARL13B and ADCY3 Localization in the Mouse Brain

 , , and
, , and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Genotyping
2.3. Diet and Feeding Manipulations
2.4. Fixation and Preparation of Slide Sections
2.5. Immunofluorescence
2.6. RNAScope In Situ Hybridization
2.7. Imaging
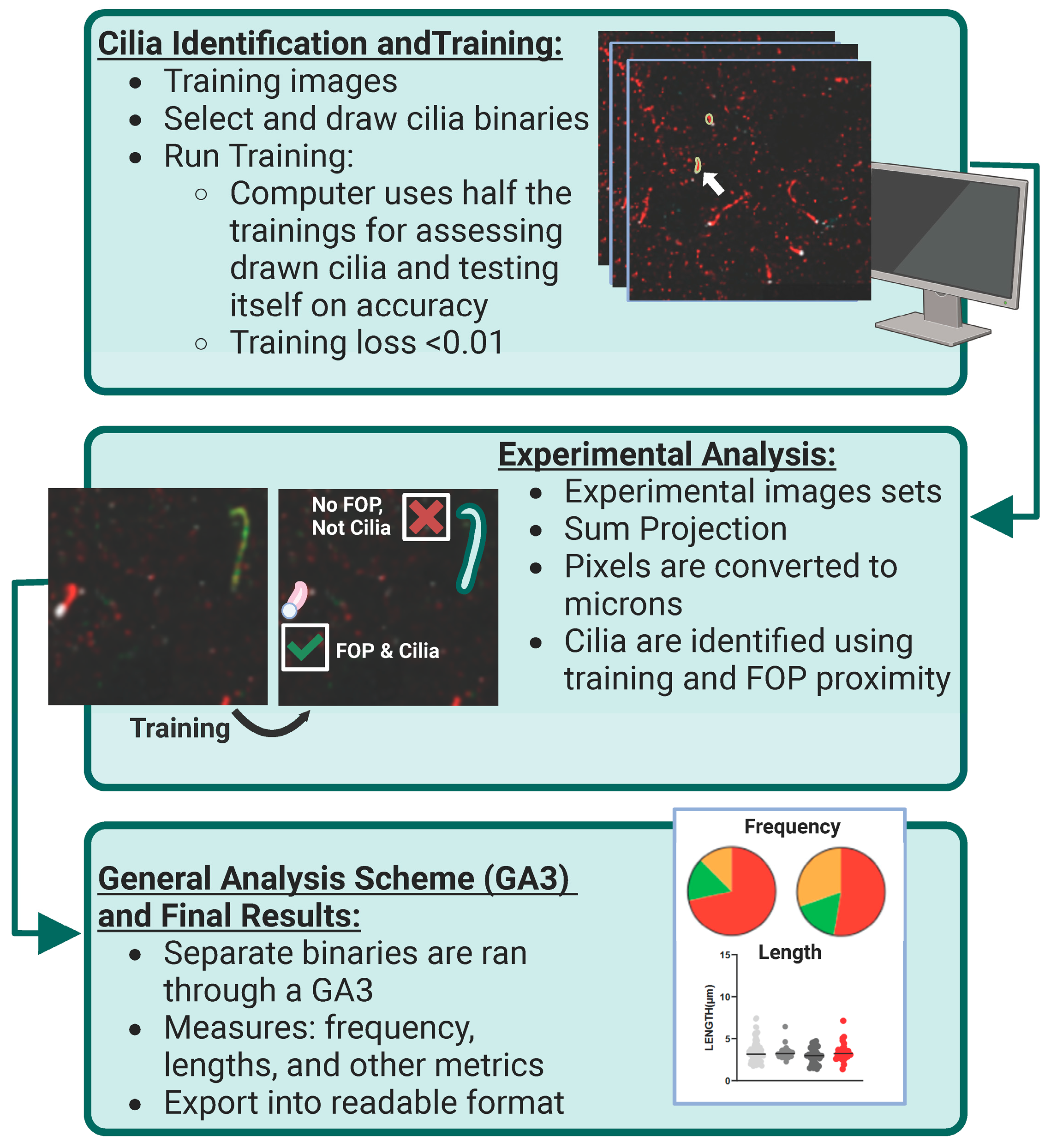
2.8. Image Data Analysis
2.9. Statistical Analysis
3. Results
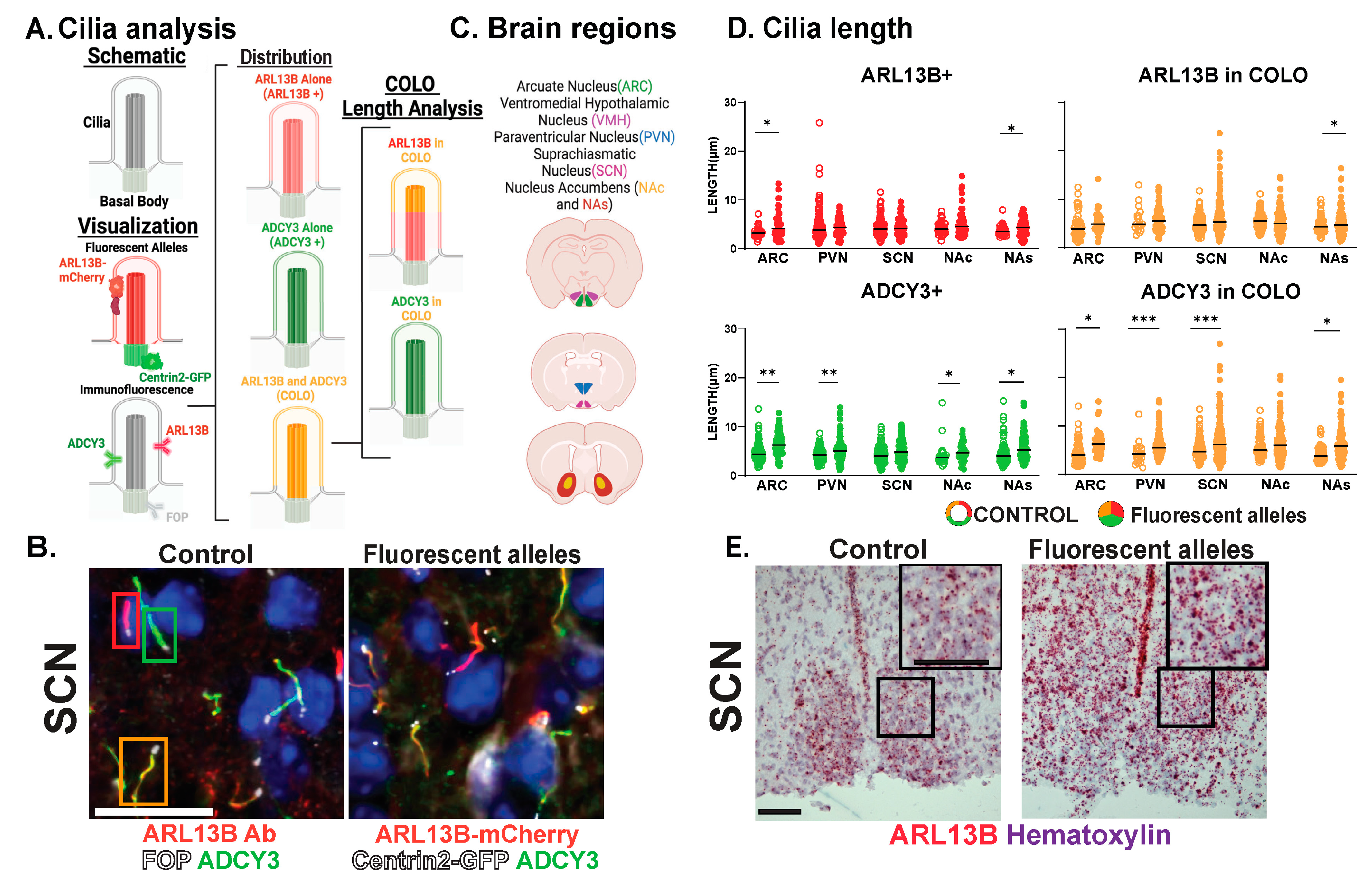
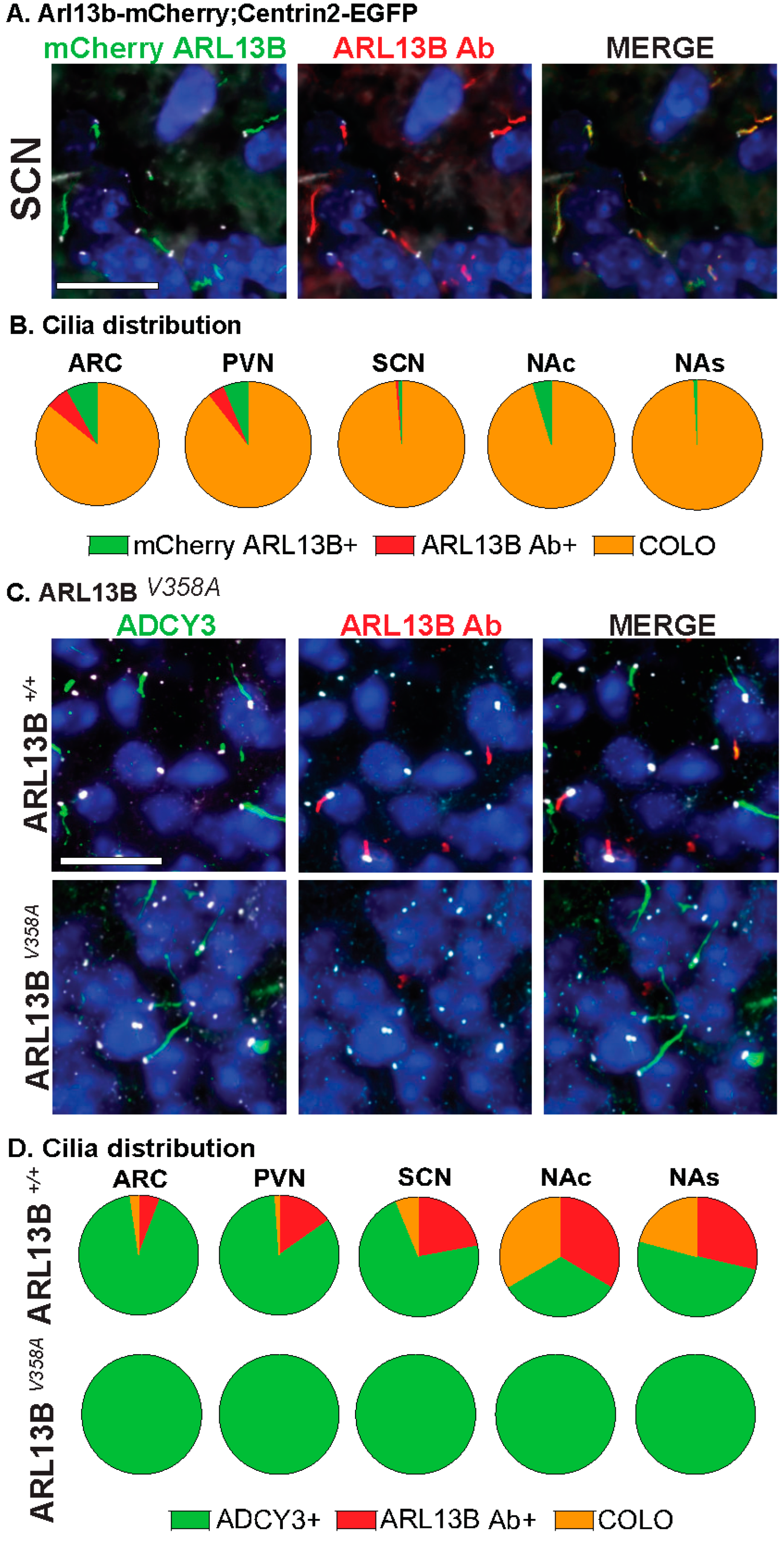
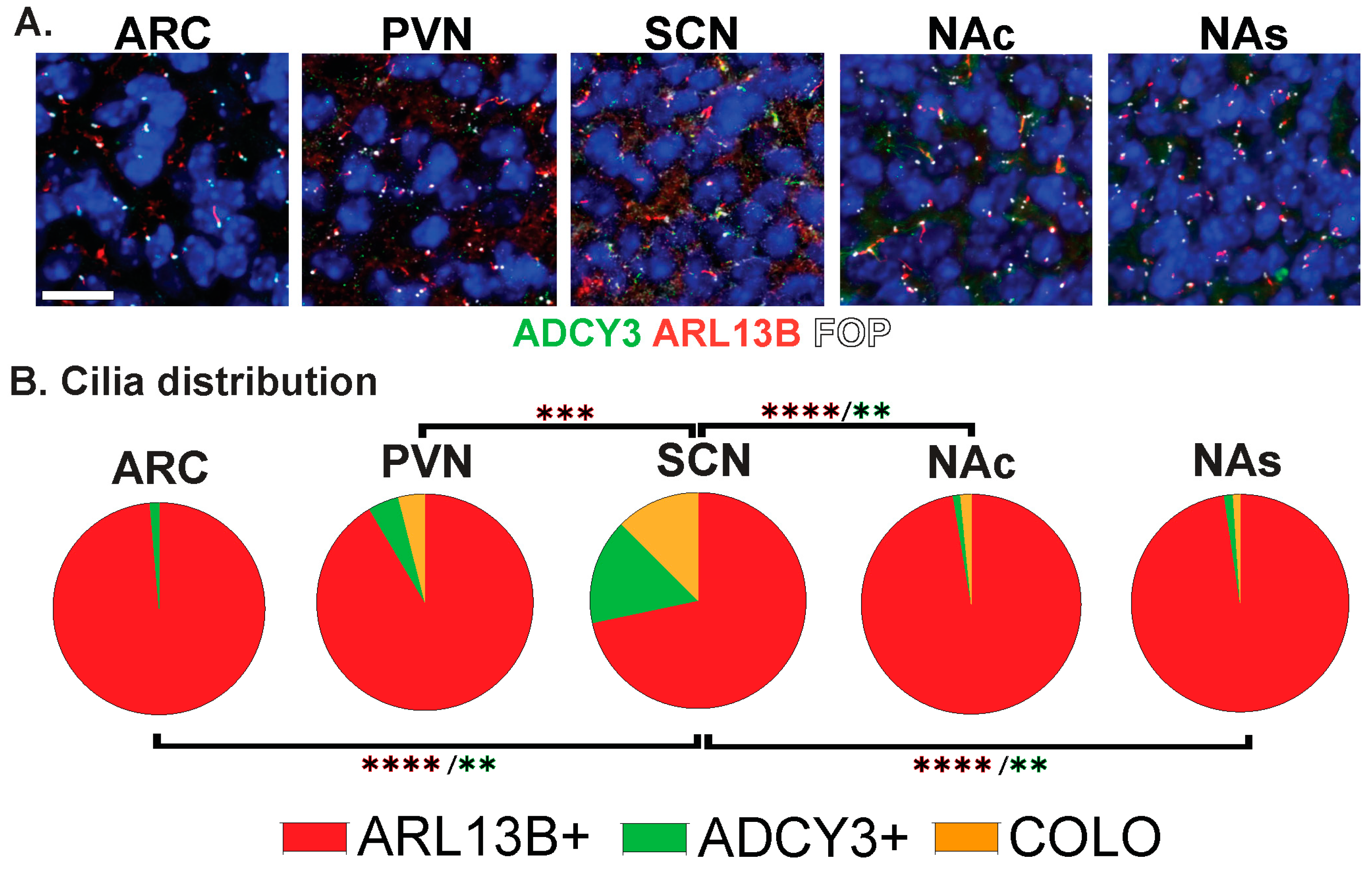
3.1. Approaches for Cilia Visualization and Large-Scale Analysis in the Brain
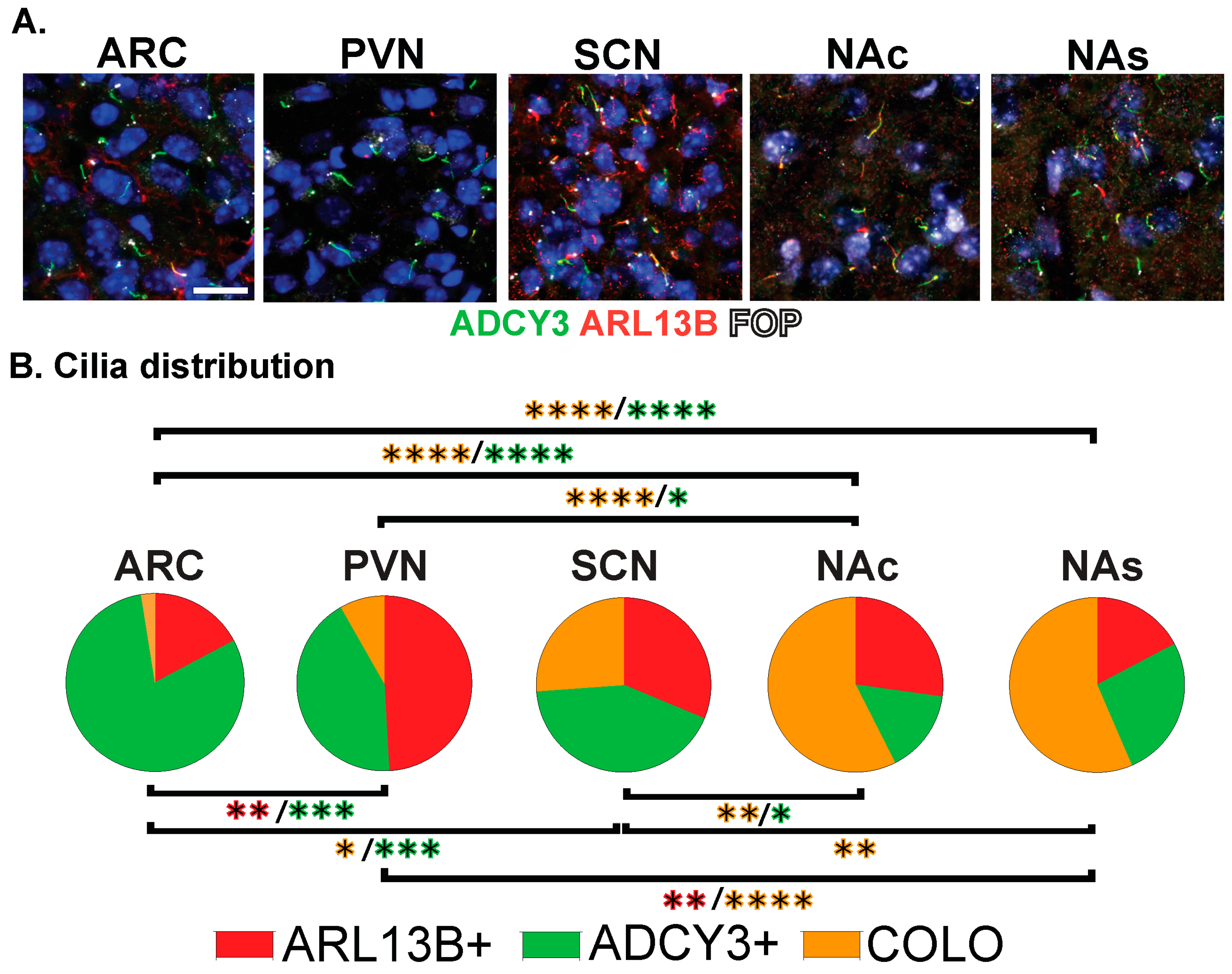
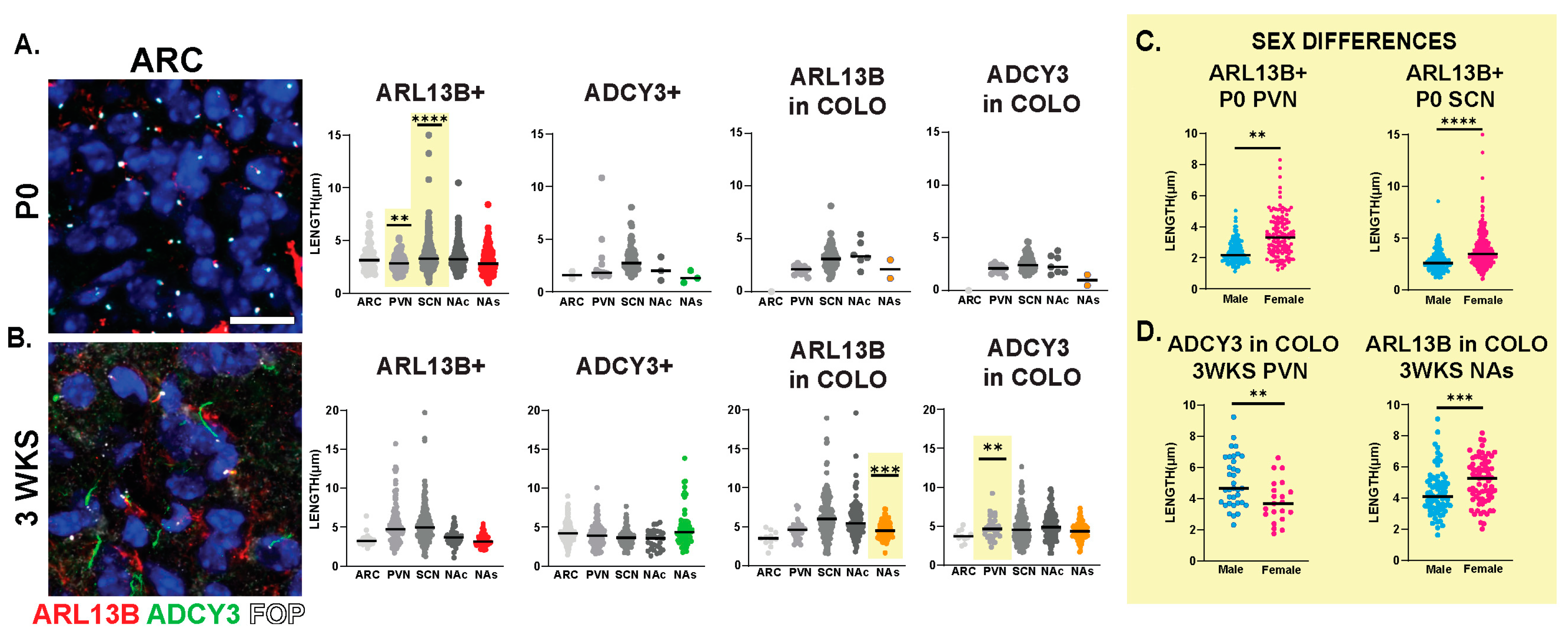
3.2. Neuroanatomical Region Dependent Cilia Signatures in the Adult and Perinatal (P0) Brain
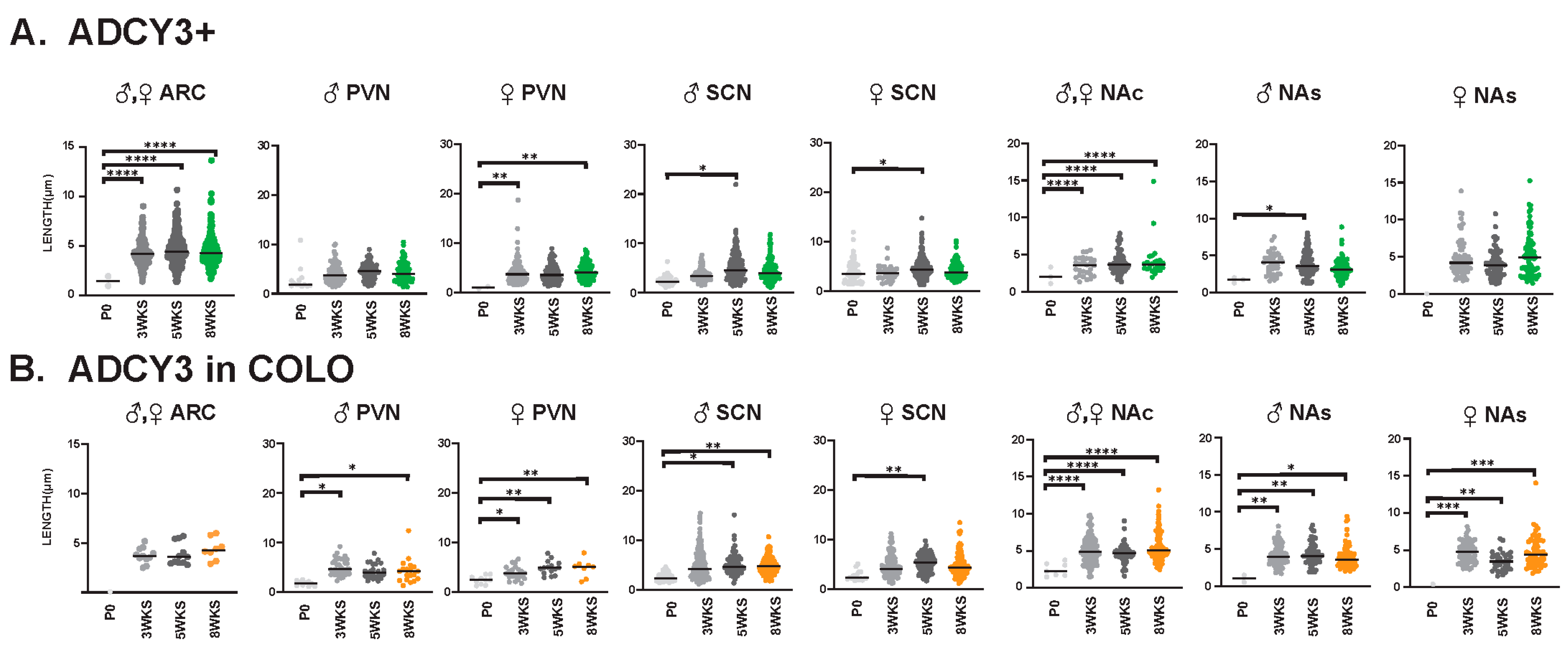
3.3. Sex-Dependent Cilia Length Differences
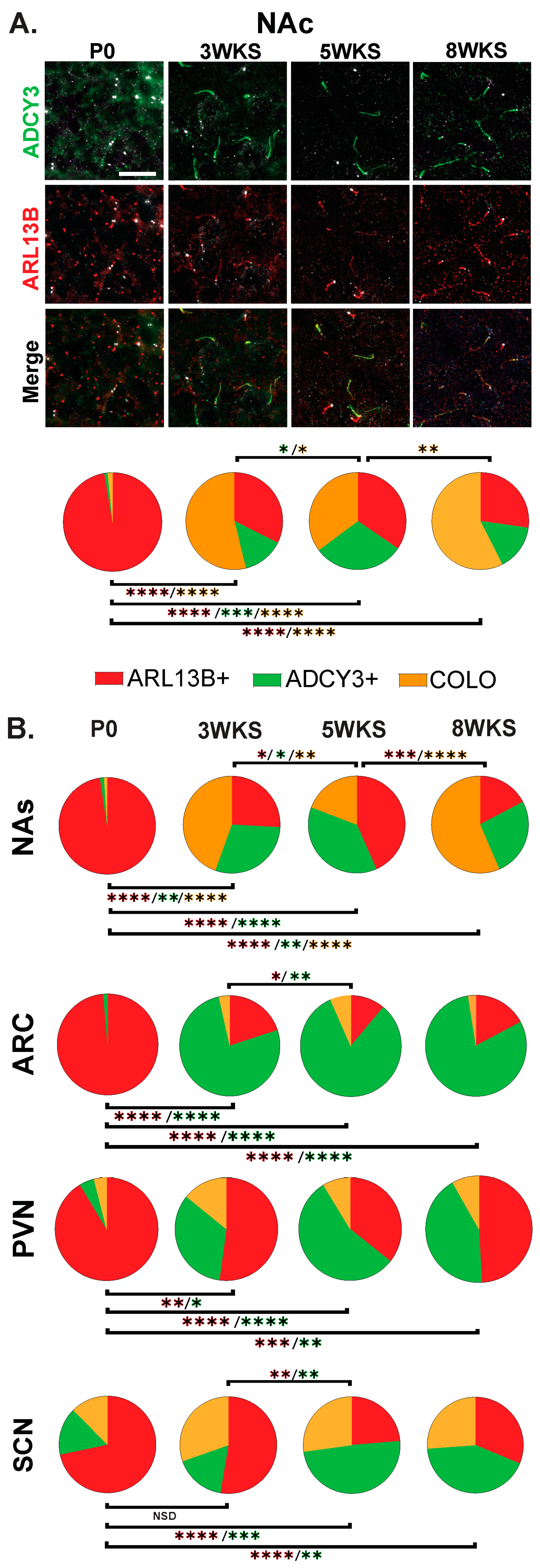
3.4. Cilia Signatures in the Brain at Different Postnatal Developmental Ages
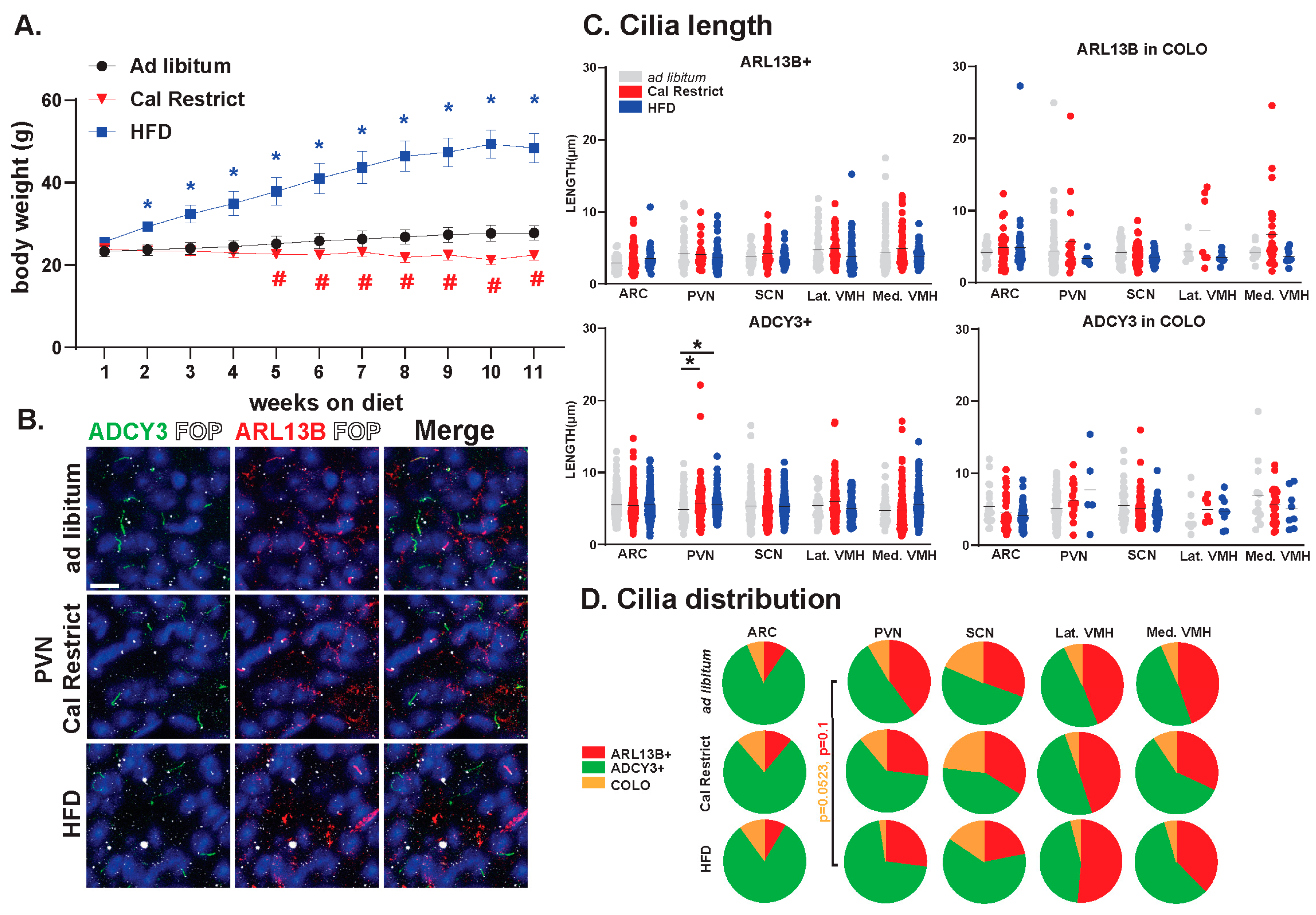
3.5. Impact of Energy Homeostasis Changes on Cilia Lengths in the Hypothalamus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reiter, J.F.; Leroux, M.R. Genes and molecular pathways underpinning ciliopathies. Nat. Rev. Mol. Cell Biol. 2017, 18, 533–547. [Google Scholar] [CrossRef] [PubMed]
- Singla, V.; Reiter, J.F. The primary cilium as the cell’s antenna: Signaling at a sensory organelle. Science 2006, 313, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Hamamoto, A.; Saito, Y. Analysis of ciliary status via G-protein-coupled receptors localized on primary cilia. Microscopy 2020, 69, 277–285. [Google Scholar] [CrossRef]
- Mykytyn, K.; Askwith, C. G-Protein-Coupled Receptor Signaling in Cilia. Cold Spring Harb. Perspect. Biol. 2017, 9, a028183. [Google Scholar] [CrossRef] [PubMed]
- Higginbotham, H.; Guo, J.; Yokota, Y.; Umberger, N.L.; Su, C.Y.; Li, J.; Verma, N.; Hirt, J.; Ghukasyan, V.; Caspary, T.; et al. Arl13b-regulated cilia activities are essential for polarized radial glial scaffold formation. Nat. Neurosci. 2013, 16, 1000–1007. [Google Scholar] [CrossRef]
- Larkins, C.E.; Aviles, G.D.; East, M.P.; Kahn, R.A.; Caspary, T. Arl13b regulates ciliogenesis and the dynamic localization of Shh signaling proteins. Mol. Biol. Cell 2011, 22, 4694–4703. [Google Scholar] [CrossRef] [PubMed]
- Roy, K.; Jerman, S.; Jozsef, L.; McNamara, T.; Onyekaba, G.; Sun, Z.; Marin, E.P. Palmitoylation of the ciliary GTPase ARL13b is necessary for its stability and its role in cilia formation. J. Biol. Chem. 2017, 292, 17703–17717. [Google Scholar] [CrossRef]
- Toumba, M.; Fanis, P.; Vlachakis, D.; Neocleous, V.; Phylactou, L.A.; Skordis, N.; Mantzoros, C.S.; Pantelidou, M. Molecular modelling of novel ADCY3 variant predicts a molecular target for tackling obesity. Int. J. Mol. Med. 2022, 49, 10. [Google Scholar] [CrossRef]
- Blitzer, A.L.; Panagis, L.; Gusella, G.L.; Danias, J.; Mlodzik, M.; Iomini, C. Primary cilia dynamics instruct tissue patterning and repair of corneal endothelium. Proc. Natl. Acad. Sci. USA 2011, 108, 2819–2824. [Google Scholar] [CrossRef]
- Anvarian, Z.; Mykytyn, K.; Mukhopadhyay, S.; Pedersen, L.B.; Christensen, S.T. Cellular signalling by primary cilia in development, organ function and disease. Nat. Rev. Nephrol. 2019, 15, 199–219. [Google Scholar] [CrossRef]
- Corbit, K.C.; Aanstad, P.; Singla, V.; Norman, A.R.; Stainier, D.Y.; Reiter, J.F. Vertebrate Smoothened functions at the primary cilium. Nature 2005, 437, 1018–1021. [Google Scholar] [CrossRef] [PubMed]
- Haycraft, C.J.; Banizs, B.; Aydin-Son, Y.; Zhang, Q.; Michaud, E.J.; Yoder, B.K. Gli2 and Gli3 localize to cilia and require the intraflagellar transport protein polaris for processing and function. PLoS Genet. 2005, 1, e53. [Google Scholar] [CrossRef]
- Rohatgi, R.; Milenkovic, L.; Scott, M.P. Patched1 regulates hedgehog signaling at the primary cilium. Science 2007, 317, 372–376. [Google Scholar] [CrossRef]
- Bangs, F.; Anderson, K.V. Primary Cilia and Mammalian Hedgehog Signaling. Cold Spring Harb. Perspect. Biol. 2017, 9, a028175. [Google Scholar] [CrossRef]
- Arrojo, E.D.R.; Lev-Ram, V.; Tyagi, S.; Ramachandra, R.; Deerinck, T.; Bushong, E.; Phan, S.; Orphan, V.; Lechene, C.; Ellisman, M.H.; et al. Age Mosaicism across Multiple Scales in Adult Tissues. Cell Metab. 2019, 30, 343–351.e3. [Google Scholar] [CrossRef]
- DeMars, K.M.; Ross, M.R.; Starr, A.; McIntyre, J.C. Neuronal primary cilia integrate peripheral signals with metabolic drives. Front. Physiol. 2023, 14, 1150232. [Google Scholar] [CrossRef]
- Tu, H.Q.; Li, S.; Xu, Y.L.; Zhang, Y.C.; Li, P.Y.; Liang, L.Y.; Song, G.P.; Jian, X.X.; Wu, M.; Song, Z.Q.; et al. Rhythmic cilia changes support SCN neuron coherence in circadian clock. Science 2023, 380, 972–979. [Google Scholar] [CrossRef]
- Domire, J.S.; Mykytyn, K. Markers for neuronal cilia. Methods Cell Biol. 2009, 91, 111–121. [Google Scholar] [CrossRef]
- Shin, J.; Prescott, M.; Mair, J.; Campbell, R.E. Roles for primary cilia in gonadotrophin-releasing hormone neurones in the mouse. J. Neuroendocrinol. 2014, 26, 18–25. [Google Scholar] [CrossRef]
- Green, J.A.; Mykytyn, K. Neuronal primary cilia: An underappreciated signaling and sensory organelle in the brain. Neuropsychopharmacology 2014, 39, 244–245. [Google Scholar] [CrossRef]
- Mirzadeh, Z.; Kusne, Y.; Duran-Moreno, M.; Cabrales, E.; Gil-Perotin, S.; Ortiz, C.; Chen, B.; Garcia-Verdugo, J.M.; Sanai, N.; Alvarez-Buylla, A. Bi- and uniciliated ependymal cells define continuous floor-plate-derived tanycytic territories. Nat. Commun. 2017, 8, 13759. [Google Scholar] [CrossRef]
- Eichele, G.; Bodenschatz, E.; Ditte, Z.; Gunther, A.K.; Kapoor, S.; Wang, Y.; Westendorf, C. Cilia-driven flows in the brain third ventricle. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20190154. [Google Scholar] [CrossRef]
- Sheu, S.H.; Upadhyayula, S.; Dupuy, V.; Pang, S.; Deng, F.; Wan, J.; Walpita, D.; Pasolli, H.A.; Houser, J.; Sanchez-Martinez, S.; et al. A serotonergic axon-cilium synapse drives nuclear signaling to alter chromatin accessibility. Cell 2022, 185, 3390–3407.e18. [Google Scholar] [CrossRef]
- Wu, J.Y.; Cho, S.J.; Descant, K.; Li, P.H.; Shapson-Coe, A.; Januszewski, M.; Berger, D.R.; Meyer, C.; Casingal, C.; Huda, A.; et al. Mapping of neuronal and glial primary cilia contactome and connectome in the human cerebral cortex. Neuron 2023, 112, 41–55.e3. [Google Scholar] [CrossRef]
- Ott, C.M.; Torres, R.; Kuan, T.S.; Kuan, A.; Buchanan, J.; Elabbady, L.; Seshamani, S.; Bodor, A.L.; Collman, F.; Bock, D.D.; et al. Nanometer-scale views of visual cortex reveal anatomical features of primary cilia poised to detect synaptic spillover. bioRxiv 2023. [Google Scholar] [CrossRef]
- Park, S.M.; Jang, H.J.; Lee, J.H. Roles of Primary Cilia in the Developing Brain. Front. Cell Neurosci. 2019, 13, 218. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Stearns, T. FOP is a centriolar satellite protein involved in ciliogenesis. PLoS ONE 2013, 8, e58589. [Google Scholar] [CrossRef] [PubMed]
- Bangs, F.K.; Schrode, N.; Hadjantonakis, A.K.; Anderson, K.V. Lineage specificity of primary cilia in the mouse embryo. Nat. Cell Biol. 2015, 17, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Mykytyn, K.; Mullins, R.F.; Andrews, M.; Chiang, A.P.; Swiderski, R.E.; Yang, B.; Braun, T.; Casavant, T.; Stone, E.M.; Sheffield, V.C. Bardet-Biedl syndrome type 4 (BBS4)-null mice implicate Bbs4 in flagella formation but not global cilia assembly. Proc. Natl. Acad. Sci. USA 2004, 101, 8664–8669. [Google Scholar] [CrossRef]
- Gigante, E.D.; Taylor, M.R.; Ivanova, A.A.; Kahn, R.A.; Caspary, T. ARL13B regulates Sonic hedgehog signaling from outside primary cilia. Elife 2020, 9, e50434. [Google Scholar] [CrossRef]
- Terry, T.T.; Gigante, E.D.; Alexandre, C.M.; Brewer, K.M.; Engle, S.E.; Yue, X.; Berbari, N.F.; Vaisse, C.; Caspary, T. Ciliary ARL13B prevents obesity in mice. bioRxiv 2023. [Google Scholar] [CrossRef]
- Wolterink-Donselaar, I.G.; Meerding, J.M.; Fernandes, C. A method for gender determination in newborn dark pigmented mice. Lab Anim. 2009, 38, 35–38. [Google Scholar] [CrossRef]
- Papaioannou, V.E.; Behringer, R.R. Sex Genotyping Mice by Polymerase Chain Reaction. Cold Spring Harb. Protoc. 2024, 2024, pdb-prot108062. [Google Scholar] [CrossRef] [PubMed]
- Higginbotham, H.; Bielas, S.; Tanaka, T.; Gleeson, J.G. Transgenic mouse line with green-fluorescent protein-labeled Centrin 2 allows visualization of the centrosome in living cells. Transgenic Res. 2004, 13, 155–164. [Google Scholar] [CrossRef]
- Li, X.; Yang, S.; Deepak, V.; Chinipardaz, Z.; Yang, S. Identification of Cilia in Different Mouse Tissues. Cells 2021, 10, 1623. [Google Scholar] [CrossRef]
- Brewer, K.M.; Engle, S.E.; Bansal, R.; Brewer, K.K.; Jasso, K.R.; McIntyre, J.C.; Vaisse, C.; Reiter, J.F.; Berbari, N.F. Physiological Condition-Dependent Changes in Ciliary GPCR Localization in the Brain. eNeuro 2023, 10. [Google Scholar] [CrossRef] [PubMed]
- Engle, S.E.; Antonellis, P.J.; Whitehouse, L.S.; Bansal, R.; Emond, M.R.; Jontes, J.D.; Kesterson, R.A.; Mykytyn, K.; Berbari, N.F. A CreER mouse to study melanin concentrating hormone signaling in the developing brain. Genesis 2018, 56, e23217. [Google Scholar] [CrossRef]
- Bansal, R.; Engle, S.E.; Kamba, T.K.; Brewer, K.M.; Lewis, W.R.; Berbari, N.F. Artificial Intelligence Approaches to Assessing Primary Cilia. J. Vis. Exp. 2021, 171, e62521. [Google Scholar] [CrossRef]
- Brewer, K.M.; Brewer, K.K.; Richardson, N.C.; Berbari, N.F. Neuronal cilia in energy homeostasis. Front. Cell Dev. Biol. 2022, 10, 1082141. [Google Scholar] [CrossRef]
- Pickel, L.; Sung, H.K. Feeding Rhythms and the Circadian Regulation of Metabolism. Front. Nutr. 2020, 7, 39. [Google Scholar] [CrossRef]
- Everett, T.; Ten Eyck, T.W.; Wu, C.H.; Shelowitz, A.L.; Stansbury, S.M.; Firek, A.; Setlow, B.; McIntyre, J.C. Cilia loss on distinct neuron populations differentially alters cocaine-induced locomotion and reward. J. Psychopharmacol. 2023, 27, 2698811231219058. [Google Scholar] [CrossRef]
- Sun, J.S.; Yang, D.J.; Kinyua, A.W.; Yoon, S.G.; Seong, J.K.; Kim, J.; Moon, S.J.; Shin, D.M.; Choi, Y.H.; Kim, K.W. Ventromedial hypothalamic primary cilia control energy and skeletal homeostasis. J. Clin. Investig. 2021, 131, e138107. [Google Scholar] [CrossRef]
- Yang, D.; Wu, X.; Wang, W.; Zhou, Y.; Wang, Z. Ciliary Type III Adenylyl Cyclase in the VMH Is Crucial for High-Fat Diet-Induced Obesity Mediated by Autophagy. Adv. Sci. 2022, 9, e2102568. [Google Scholar] [CrossRef] [PubMed]
- Bernard, A.; Ojeda Naharros, I.; Yue, X.; Mifsud, F.; Blake, A.; Bourgain-Guglielmetti, F.; Ciprin, J.; Zhang, S.; McDaid, E.; Kim, K.; et al. MRAP2 regulates energy homeostasis by promoting primary cilia localization of MC4R. JCI Insight 2023, 8, e155900. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.F.; Lin, Z.; Wu, Y.; Searby, C.; Thedens, D.R.; Richerson, G.B.; Usachev, Y.M.; Grobe, J.L.; Sheffield, V.C.; Rahmouni, K. The BBSome in POMC and AgRP Neurons Is Necessary for Body Weight Regulation and Sorting of Metabolic Receptors. Diabetes 2019, 68, 1591–1603. [Google Scholar] [CrossRef] [PubMed]
- Kang, G.M.; Han, Y.M.; Ko, H.W.; Kim, J.; Oh, B.C.; Kwon, I.; Kim, M.S. Leptin Elongates Hypothalamic Neuronal Cilia via Transcriptional Regulation and Actin Destabilization. J. Biol. Chem. 2015, 290, 18146–18155. [Google Scholar] [CrossRef]
- Wang, Y.; Bernard, A.; Comblain, F.; Yue, X.; Paillart, C.; Zhang, S.; Reiter, J.F.; Vaisse, C. Melanocortin 4 receptor signals at the neuronal primary cilium to control food intake and body weight. J. Clin. Investig. 2021, 131, e142064. [Google Scholar] [CrossRef] [PubMed]
- Sterpka, A.; Yang, J.; Strobel, M.; Zhou, Y.; Pauplis, C.; Chen, X. Diverged morphology changes of astrocytic and neuronal primary cilia under reactive insults. Mol. Brain 2020, 13, 28. [Google Scholar] [CrossRef] [PubMed]
- Ott, C.M.; Constable, S.; Nguyen, T.M.; White, K.; Lee, W.A.; Lippincott-Schwartz, J.; Mukhopadhyay, S. Permanent deconstruction of intracellular primary cilia in differentiating granule cell neurons. bioRxiv 2023. [Google Scholar] [CrossRef]
- Katidou, M.; Grosmaitre, X.; Lin, J.; Mombaerts, P. G-protein coupled receptors Mc4r and Drd1a can serve as surrogate odorant receptors in mouse olfactory sensory neurons. Mol. Cell Neurosci. 2018, 88, 138–147. [Google Scholar] [CrossRef]
- Barbeito, P.; Garcia-Gonzalo, F.R. HTR6 and SSTR3 targeting to primary cilia. Biochem. Soc. Trans. 2021, 49, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Loktev, A.V.; Jackson, P.K. Neuropeptide Y family receptors traffic via the Bardet-Biedl syndrome pathway to signal in neuronal primary cilia. Cell Rep. 2013, 5, 1316–1329. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; LeBel, R.P.; Storm, D.R.; Chen, X. Type 3 adenylyl cyclase: A key enzyme mediating the cAMP signaling in neuronal cilia. Int. J. Physiol. Pathophysiol. Pharmacol. 2016, 8, 95–108. [Google Scholar] [PubMed]
- Higginbotham, H.; Eom, T.Y.; Mariani, L.E.; Bachleda, A.; Hirt, J.; Gukassyan, V.; Cusack, C.L.; Lai, C.; Caspary, T.; Anton, E.S. Arl13b in primary cilia regulates the migration and placement of interneurons in the developing cerebral cortex. Dev. Cell 2012, 23, 925–938. [Google Scholar] [CrossRef] [PubMed]
- Bakalyar, H.A.; Reed, R.R. Identification of a specialized adenylyl cyclase that may mediate odorant detection. Science 1990, 250, 1403–1406. [Google Scholar] [CrossRef]
- Bishop, G.A.; Berbari, N.F.; Lewis, J.; Mykytyn, K. Type III adenylyl cyclase localizes to primary cilia throughout the adult mouse brain. J. Comp. Neurol. 2007, 505, 562–571. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, Y.; Zheng, Y.; Zheng, M.; Sun, N.; Yang, X.; Gao, Y. Progress in Research on Brain Development and Function of Mice During Weaning. Curr. Protein Pept. Sci. 2019, 20, 705–712. [Google Scholar] [CrossRef]
- Yamaguchi, T. Neural circuit mechanisms of sex and fighting in male mice. Neurosci. Res. 2022, 174, 1–8. [Google Scholar] [CrossRef]
- Xie, Y.; Dorsky, R.I. Development of the hypothalamus: Conservation, modification and innovation. Development 2017, 144, 1588–1599. [Google Scholar] [CrossRef]
- Huppe-Gourgues, F.; O’Donnell, P. Periadolescent changes of D(2) -AMPA interactions in the rat nucleus accumbens. Synapse 2012, 66, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Huppe-Gourgues, F.; O’Donnell, P. D(1)-NMDA receptor interactions in the rat nucleus accumbens change during adolescence. Synapse 2012, 66, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Kasanetz, F.; Manzoni, O.J. Maturation of excitatory synaptic transmission of the rat nucleus accumbens from juvenile to adult. J. Neurophysiol. 2009, 101, 2516–2527. [Google Scholar] [CrossRef] [PubMed]
- Matthiesen, M.; Khlaifia, A.; Steininger, C.F.D., Jr.; Dadabhoy, M.; Mumtaz, U.; Arruda-Carvalho, M. Maturation of nucleus accumbens synaptic transmission signals a critical period for the rescue of social deficits in a mouse model of autism spectrum disorder. Mol. Brain 2023, 16, 46. [Google Scholar] [CrossRef] [PubMed]
- Nelson, L.H.; Warden, S.; Lenz, K.M. Sex differences in microglial phagocytosis in the neonatal hippocampus. Brain Behav. Immun. 2017, 64, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Alcantara, I.C.; Tapia, A.P.M.; Aponte, Y.; Krashes, M.J. Acts of appetite: Neural circuits governing the appetitive, consummatory, and terminating phases of feeding. Nat. Metab. 2022, 4, 836–847. [Google Scholar] [CrossRef] [PubMed]
- Dunphy-Doherty, F.; O’Mahony, S.M.; Peterson, V.L.; O’Sullivan, O.; Crispie, F.; Cotter, P.D.; Wigmore, P.; King, M.V.; Cryan, J.F.; Fone, K.C.F. Post-weaning social isolation of rats leads to long-term disruption of the gut microbiota-immune-brain axis. Brain Behav. Immun. 2018, 68, 261–273. [Google Scholar] [CrossRef]
- Xiong, Y.; Hong, H.; Liu, C.; Zhang, Y.Q. Social isolation and the brain: Effects and mechanisms. Mol. Psychiatry 2023, 28, 191–201. [Google Scholar] [CrossRef]
- Karigo, T.; Deutsch, D. Flexibility of neural circuits regulating mating behaviors in mice and flies. Front. Neural Circuits 2022, 16, 949781. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brewer, K.K.; Brewer, K.M.; Terry, T.T.; Caspary, T.; Vaisse, C.; Berbari, N.F. Postnatal Dynamic Ciliary ARL13B and ADCY3 Localization in the Mouse Brain. Cells 2024, 13, 259. https://doi.org/10.3390/cells13030259
Brewer KK, Brewer KM, Terry TT, Caspary T, Vaisse C, Berbari NF. Postnatal Dynamic Ciliary ARL13B and ADCY3 Localization in the Mouse Brain. Cells. 2024; 13(3):259. https://doi.org/10.3390/cells13030259
Chicago/Turabian StyleBrewer, Katlyn K., Kathryn M. Brewer, Tiffany T. Terry, Tamara Caspary, Christian Vaisse, and Nicolas F. Berbari. 2024. "Postnatal Dynamic Ciliary ARL13B and ADCY3 Localization in the Mouse Brain" Cells 13, no. 3: 259. https://doi.org/10.3390/cells13030259
APA StyleBrewer, K. K., Brewer, K. M., Terry, T. T., Caspary, T., Vaisse, C., & Berbari, N. F. (2024). Postnatal Dynamic Ciliary ARL13B and ADCY3 Localization in the Mouse Brain. Cells, 13(3), 259. https://doi.org/10.3390/cells13030259