
Circulating miRNAs in Women with Polycystic Ovary Syndrome: A Longitudinal Cohort Study


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Participants
2.2. Anthropometric and Clinical Characteristics at FU
2.3. Serum MiRNA Isolation and Analysis
2.4. Statistical Analysis
3. Results
3.1. Clinical Phenotype at FU in Control and PCOS Subjects
3.2. Longitudinal Changes in MiRNA Levels
3.3. MiRNA Profile Associated with a Shift in Metabolic Profile
3.4. BL Circulating MiRNAs Are Associated with PCOS
3.5. Androgen Levels and LH/FSH Ratio Correlate with MiRNA Levels at BL
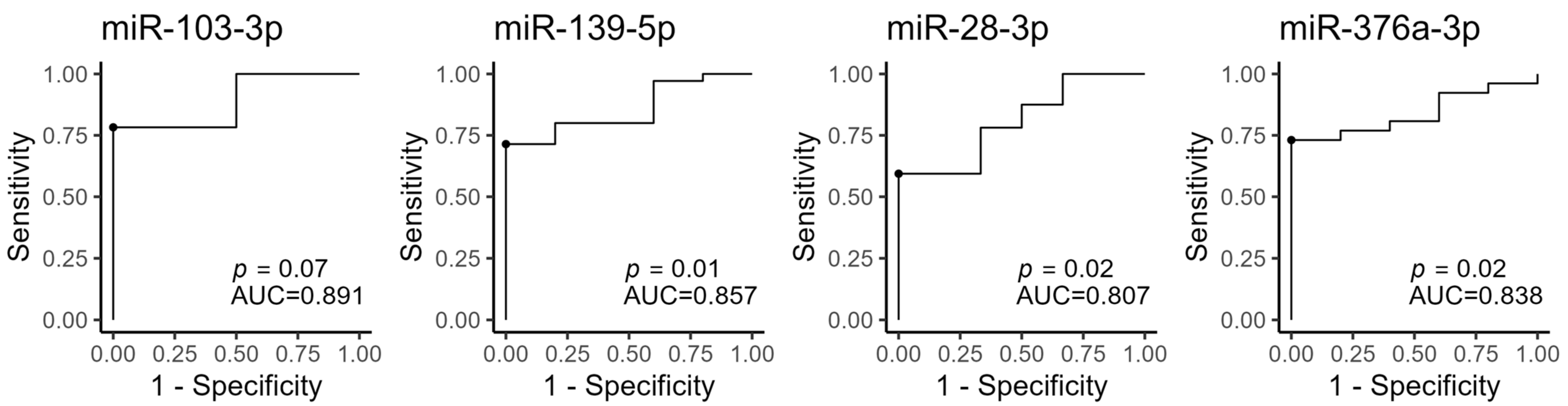
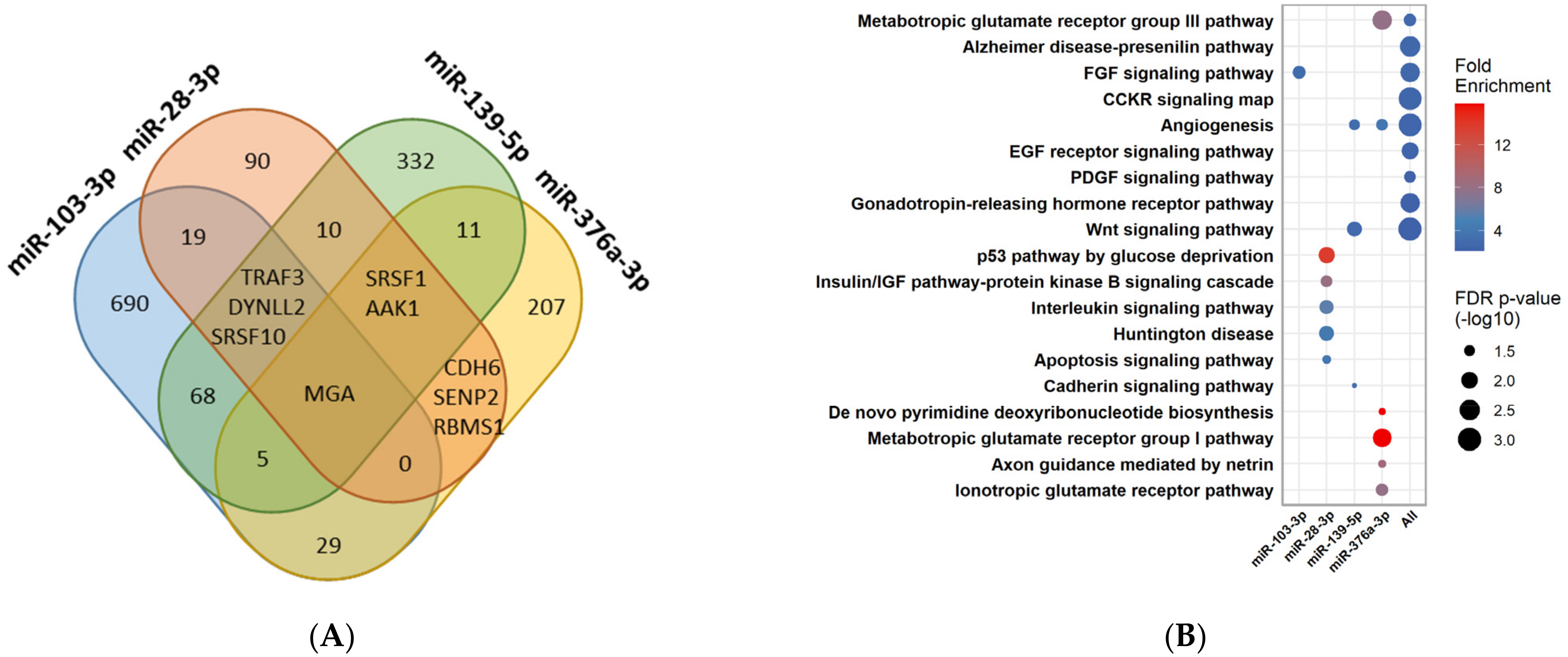
3.6. Pathway Enrichment Analysis for the Four MiRNAs Different between PCOS and Control Women during FU
3.7. Pathway Enrichment Analysis for the MiRNAs Significantly Changed in PCOS Women during FU
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Teede, H.J.; Misso, M.L.; Costello, M.F.; Dokras, A.; Laven, J.; Moran, L.; Piltonen, T.; Norman, R.J. Pcos Network International. Recommendations from the international evidence-based guideline for the assessment and management of polycystic ovary syndrome. Clin. Endocrinol. 2018, 89, 251–268. [Google Scholar] [CrossRef] [PubMed]
- Diamanti-Kandarakis, E.; Dunaif, A. Insulin resistance and the polycystic ovary syndrome revisited: An update on mechanisms and implications. Endocr. Rev. 2012, 33, 981–1030. [Google Scholar] [CrossRef] [PubMed]
- Glintborg, D.; Rubin, K.H.; Nybo, M.; Abrahamsen, B.; Andersen, M. Cardiovascular disease in a nationwide population of Danish women with polycystic ovary syndrome. Cardiovasc. Diabetol. 2018, 17, 37. [Google Scholar] [CrossRef]
- Rubin, K.H.; Glintborg, D.; Nybo, M.; Abrahamsen, B.; Andersen, M. Development and Risk Factors of Type 2 Diabetes in a Nationwide Population of Women with Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2017, 102, 3848–3857. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Vienberg, S.; Geiger, J.; Madsen, S.; Dalgaard, L.T. MicroRNAs in metabolism. Acta Physiol. 2017, 219, 346–361. [Google Scholar] [CrossRef] [PubMed]
- Lynn, F.C. Meta-regulation: microRNA regulation of glucose and lipid metabolism. Trends Endocrinol. Metab. 2009, 20, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Sebastiani, G.; Guay, C.; Latreille, M. Circulating Noncoding RNAs as Candidate Biomarkers of Endocrine and Metabolic Diseases. Int. J. Endocrinol. 2018, 2018, 9514927. [Google Scholar] [CrossRef]
- Hess, A.L.; Larsen, L.H.; Udesen, P.B.; Sanz, Y.; Larsen, T.M.; Dalgaard, L.T. Levels of Circulating miR-122 are Associated with Weight Loss and Metabolic Syndrome. Obesity 2020, 28, 493–501. [Google Scholar] [CrossRef]
- Ortega, F.J.; Mercader, J.M.; Moreno-Navarrete, J.M.; Rovira, O.; Guerra, E.; Esteve, E.; Xifra, G.; Martinez, C.; Ricart, W.; Rieusset, J.; et al. Profiling of circulating microRNAs reveals common microRNAs linked to type 2 diabetes that change with insulin sensitization. Diabetes Care 2014, 37, 1375–1383. [Google Scholar] [CrossRef]
- Bae, Y.U.; Kim, Y.; Lee, H.; Kim, H.; Jeon, J.S.; Noh, H.; Han, D.C.; Ryu, S.; Kwon, S.H. Bariatric Surgery Alters microRNA Content of Circulating Exosomes in Patients with Obesity. Obesity 2019, 27, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Willeit, P.; Skroblin, P.; Moschen, A.R.; Yin, X.; Kaudewitz, D.; Zampetaki, A.; Barwari, T.; Whitehead, M.; Ramirez, C.M.; Goedeke, L.; et al. Circulating MicroRNA-122 Is Associated with the Risk of New-Onset Metabolic Syndrome and Type 2 Diabetes. Diabetes 2017, 66, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Karolina, D.S.; Tavintharan, S.; Armugam, A.; Sepramaniam, S.; Pek, S.L.; Wong, M.T.; Lim, S.C.; Sum, C.F.; Jeyaseelan, K. Circulating miRNA profiles in patients with metabolic syndrome. J. Clin. Endocrinol. Metab. 2012, 97, E2271–E2276. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, A.E.; Udesen, P.B.; Wissing, M.L.; Englund, A.L.; Dalgaard, L.T. MicroRNAs related to androgen metabolism and polycystic ovary syndrome. Chem. Biol. Interact. 2016, 259 Pt A, 8–16. [Google Scholar] [CrossRef]
- Sorensen, A.E.; Wissing, M.L.; Salo, S.; Englund, A.L.; Dalgaard, L.T. MicroRNAs Related to Polycystic Ovary Syndrome (PCOS). Genes 2014, 5, 684–708. [Google Scholar] [CrossRef]
- Eisenberg, I.; Nahmias, N.; Persky, M.N.; Greenfield, C.; Goldman-Wohl, D.; Hurwitz, A.; Haimov-Kochman, R.; Yagel, S.; Imbar, T. Elevated circulating micro-ribonucleic acid (miRNA)-200b and miRNA-429 levels in anovulatory women. Fertil. Steril. 2017, 107, 269–275. [Google Scholar] [CrossRef]
- Murri, M.; Insenser, M.; Fernandez-Duran, E. San-Millan, J.L. Luque-Ramirez, M. Escobar-Morreale, H.F. Non-targeted profiling of circulating microRNAs in women with polycystic ovary syndrome (PCOS): Effects of obesity and sex hormones. Metabolism 2018, 86, 49–60. [Google Scholar] [CrossRef]
- Murri, M.; Insenser, M.; Fernandez-Duran, E. San-Millan, J.L. Escobar-Morreale, H.F. Effects of polycystic ovary syndrome (PCOS), sex hormones, and obesity on circulating miRNA-21, miRNA-27b, miRNA-103, and miRNA-155 expression. J. Clin. Endocrinol. Metab. 2013, 98, E1835–E1844. [Google Scholar] [CrossRef]
- Jiang, L.; Huang, J.; Chen, Y.; Yang, Y.; Li, R.; Li, Y.; Chen, X.; Yang, D. Identification of several circulating microRNAs from a genome-wide circulating microRNA expression profile as potential biomarkers for impaired glucose metabolism in polycystic ovarian syndrome. Endocrine 2016, 53, 280–290. [Google Scholar] [CrossRef]
- Wander, P.L.; Enquobahrie, D.A.; Bammler, T.K.; MacDonald, J.W. Srinouanprachanh, S.; Kaleru, T.; Khakpour, D.; Trikudanathan, S. Associations of plasma miRNAs with waist circumference and insulin resistance among women with polycystic ovary syndrome—Pilot study. Mol. Cell Endocrinol. 2022, 554, 111723. [Google Scholar] [CrossRef]
- Sorensen, A.E.; Wissing, M.L.; Englund, A.L.; Dalgaard, L.T. MicroRNA Species in Follicular Fluid Associating With Polycystic Ovary Syndrome and Related Intermediary Phenotypes. J. Clin. Endocrinol. Metab. 2016, 101, 1579–1589. [Google Scholar] [CrossRef]
- Sang, Q.; Yao, Z.; Wang, H.; Feng, R.; Wang, H.; Zhao, X.; Xing, Q.; Jin, L.; He, L.; Wu, L.; et al. Identification of microRNAs in human follicular fluid: Characterization of microRNAs that govern steroidogenesis in vitro and are associated with polycystic ovary syndrome in vivo. J. Clin. Endocrinol. Metab. 2013, 98, 3068–3079. [Google Scholar] [CrossRef]
- Jiang, L.; Huang, J.; Li, L.; Chen, Y.; Chen, X.; Zhao, X.; Yang, D. MicroRNA-93 promotes ovarian granulosa cells proliferation through targeting CDKN1A in polycystic ovarian syndrome. J. Clin. Endocrinol. Metab. 2015, 100, E729–E738. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Heneidi, S.; Lee, J.M.; Layman, L.C.; Stepp, D.W.; Gamboa, G.M.; Chen, B.S.; Chazenbalk, G.; Azziz, R. miRNA-93 inhibits GLUT4 and is overexpressed in adipose tissue of Polycystic Ovary Syndrome patients and women with insulin resistance. Diabetes 2013, 62, 2278–2286. [Google Scholar] [CrossRef] [PubMed]
- Sathyapalan, T.; David, R.; Gooderham, N.J.; Atkin, S.L. Increased expression of circulating miRNA-93 in women with polycystic ovary syndrome may represent a novel, non-invasive biomarker for diagnosis. Sci. Rep. 2015, 5, 16890. [Google Scholar] [CrossRef]
- Naji, M.; Aleyasin, A.; Nekoonam, S.; Arefian, E.; Mahdian, R.; Amidi, F. Differential Expression of miR-93 and miR-21 in Granulosa Cells and Follicular Fluid of Polycystic Ovary Syndrome Associating with Different Phenotypes. Sci. Rep. 2017, 7, 14671. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Li, W.; Wu, M.; Cao, S. Ciculating miRNA-21 as a Biomarker Predicts Polycystic Ovary Syndrome (PCOS) in Patients. Clin. Lab. 2015, 61, 1009–1015. [Google Scholar] [CrossRef]
- Vitale, S.G.; Fulghesu, A.M.; Mikus, M.; Watrowski, R.; D’Alterio, M.N.; Lin, L.T.; Shah, M.; Reyes-Munoz, E.; Sathyapalan, T.; Angioni, S. The Translational Role of miRNA in Polycystic Ovary Syndrome: From Bench to Bedside-A Systematic Literature Review. Biomedicines 2022, 10, 1816. [Google Scholar] [CrossRef] [PubMed]
- Fauser, B.C.; Tarlatzis, B.C.; Rebar, R.W.; Legro, R.S.; Balen, A.H.; Lobo, R.; Carmina, E.; Chang, J.; Yildiz, B.O.; Laven, J.S.; et al. Consensus on women’s health aspects of polycystic ovary syndrome (PCOS): The Amsterdam ESHRE/ASRM-Sponsored 3rd PCOS Consensus Workshop Group. Fertil. Steril. 2012, 97, 28–38. [Google Scholar] [CrossRef]
- Aziz, M.; Naver, K.V.; Wissing, M.L.; Mikkelsen, A.L.; Nilas, L.; Skouby, S.O. Polycystic ovary syndrome: Infertility, cardiovascular, metabolic and obstetrical risks, laboratory and clinical outcomes -- the PICOLO Study. Gynecol. Endocrinol. 2012, 28, 253–258. [Google Scholar] [CrossRef]
- Wissing, M.L.; Bjerge, M.R.; Olesen, A.I.; Hoest, T.; Mikkelsen, A.L. Impact of PCOS on early embryo cleavage kinetics. Reprod. Biomed. Online 2013, 28, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, A.E.; Udesen, P.B.; Maciag, G.; Geiger, J.; Saliani, N.; Januszewski, A.S.; Jiang, G.; Ma, R.C.; Hardikar, A.A.; Wissing, M.L.M.; et al. Dalgaard. Hyperandrogenism and Metabolic Syndrome Are Associated With Changes in Serum-Derived microRNAs in Women With Polycystic Ovary Syndrome. Front. Med. 2019, 6, 242. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef]
- Vermeulen, A.; Verdonck, L.; Kaufman, J.M. A critical evaluation of simple methods for the estimation of free testosterone in serum. J. Clin. Endocrinol. Metab. 1999, 84, 3666–3672. [Google Scholar] [CrossRef]
- Kolde, R. Pheatmap. (2019). Available online: https://cran.r-project.org/package=pheatmap (accessed on 20 February 2023).
- Wei, T.; Simko, V. R Package Corrplot: Visualization of a Correlation Matrix. (2017). Available online: https://github.com/taiyun/corrplot (accessed on 20 February 2023).
- Thiele, C.; Hirschfield, G. Cutpointr: Improved Estimation and Validation of Optimal Cutpoints in R. J. Stat. Softw. 2021, 98, 1–27. [Google Scholar] [CrossRef]
- Legro, R.S.; Kunselman, A.R.; Dodson, W.C.; Dunaif, A. Prevalence and predictors of risk for type 2 diabetes mellitus and impaired glucose tolerance in polycystic ovary syndrome: A prospective, controlled study in 254 affected women. J. Clin. Endocrinol. Metab. 1999, 84, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Kiconco, S.; Tay, C.T.; Rassie, K.L.; Azziz, R.; Teede, H.J.; Joham, A.E. Natural history of polycystic ovary syndrome: A systematic review of cardiometabolic outcomes from longitudinal cohort studies. Clin. Endocrinol. 2022, 96, 475–498. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.; Brannstrom, M.; Landin-Wilhelmsen, K.; Dahlgren, E. Reproductive Hormone Levels and Anthropometry in Postmenopausal Women with Polycystic Ovary Syndrome (PCOS): A 21-Year Follow-Up Study of Women Diagnosed with PCOS around 50 Years Ago and Their Age-Matched Controls. J. Clin. Endocrinol. Metab. 2011, 96, 2178–2185. [Google Scholar] [CrossRef]
- Pinola, P.; Piltonen, T.T.; Puurunen, J.; Vanky, E.; Sundstrom-Poromaa, I.; Stener-Victorin, E.; Ruokonen, A.; Puukka, K.; Tapanainen, J.S.; Morin-Papunen, L.C. Androgen Profile Through Life in Women With Polycystic Ovary Syndrome: A Nordic Multicenter Collaboration Study. J. Clin. Endocrinol. Metab. 2015, 100, 3400–3407. [Google Scholar] [CrossRef]
- Schneider, C.; Setty, M.; Holmes, A.B.; Maute, R.L.; Leslie, C.S.; Mussolin, L.; Rosolen, A.; Dalla-Favera, R.; Basso, K. MicroRNA 28 controls cell proliferation and is down-regulated in B-cell lymphomas. Proc. Natl. Acad. Sci. USA 2014, 111, 8185–8190. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhao, H.; Li, T.; Gao, X.; Gao, Q.; Tang, R.; Zhang, J.; Chen, Z.J. Association study of gene LPP in women with polycystic ovary syndrome. PLoS ONE 2012, 7, e46370. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Lucena, R.; Rangel-Zuniga, O.A.; Alcala-Diaz, J.F.; Lopez-Moreno, J.; Roncero-Ramos, I.; Molina-Abril, H.; Yubero-Serrano, E.M.; Caballero-Villarraso, J. Delgado-Lista, J.; Castano, J.P.; et al. Circulating miRNAs as Predictive Biomarkers of Type 2 Diabetes Mellitus Development in Coronary Heart Disease Patients from the CORDIOPREV Study. Mol. Ther. Nucleic. Acids 2018, 12, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Zampetaki, A.; Kiechl, S.; Drozdov, I.; Willeit, P.; Mayr, U.; Prokopi, M.; Mayr, A.; Weger, S.; Oberhollenzer, F.; Bonora, E.; et al. Plasma microRNA profiling reveals loss of endothelial miR-126 and other microRNAs in type 2 diabetes. Circ. Res. 2010, 107, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Mikaeili, S.; Rashidi, B.H.; Safa, M.; Najafi, A.; Sobhani, A.; Asadi, E.; Abbasi, M. Altered FoxO3 expression and apoptosis in granulosa cells of women with polycystic ovary syndrome. Arch. Gynecol. Obs. 2016, 294, 185–192. [Google Scholar] [CrossRef]
- Bork-Jensen, J.; Thuesen, A.C.; Bang-Bertelsen, C.H.; Grunnet, L.G.; Pociot, F.; Beck-Nielsen, H.; Ozanne, S.E.; Poulsen, P.; Vaag, A. Genetic versus Non-Genetic Regulation of miR-103, miR-143 and miR-483-3p Expression in Adipose Tissue and Their Metabolic Implications-A Twin Study. Genes 2014, 5, 508–517. [Google Scholar] [CrossRef]
- Trajkovski, M.; Hausser, J.; Soutschek, J.; Bhat, B.; Akin, A.; Zavolan, M.; Heim, M.H.; Stoffel, M. MicroRNAs 103 and 107 regulate insulin sensitivity. Nature 2011, 474, 649–653. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Huang, L.; Wei, F.; Li, G.; Huang, B.; Li, J.; Cao, C. Up-regulation of miR-139-5p protects diabetic mice from liver tissue damage and oxidative stress through inhibiting Notch signaling pathway. Acta Biochim. Biophys. Sin. 2020, 52, 390–400. [Google Scholar] [CrossRef]
- Li, M.; Zeng, Z.; Zhang, A.; Ye, Q.; Su, S.; Xia, T. WGCNA Analysis Identifies Polycystic Ovary Syndrome-Associated Circular RNAs That Interact with RNA-Binding Proteins and Sponge miRNAs. Int. J. Gen. Med. 2021, 14, 8737–8751. [Google Scholar] [CrossRef]
- Krishnan, K.; Steptoe, A.L.; Martin, H.C.; Pattabiraman, D.R.; Nones, K.; Waddell, N.; Mariasegaram, M.; Simpson, P.T.; Lakhani, S.R.; Vlassov, A.; et al. miR-139-5p is a regulator of metastatic pathways in breast cancer. RNA 2013, 19, 1767–1780. [Google Scholar] [CrossRef]
- Schepeler, T.; Holm, A.; Halvey, P.; Nordentoft, I.; Lamy, P.; Riising, E.M.; Christensen, L.L.; Thorsen, K.; Liebler, D.C.; Helin, K.; et al. Attenuation of the beta-catenin/TCF4 complex in colorectal cancer cells induces several growth-suppressive microRNAs that target cancer promoting genes. Oncogene 2012, 31, 2750–2760. [Google Scholar] [CrossRef]
- Qiao, G.Y.; Dong, B.W.; Zhu, C.J.; Yan, C.Y.; Chen, B.L. Deregulation of WNT2/FZD3/beta-catenin pathway compromises the estrogen synthesis in cumulus cells from patients with polycystic ovary syndrome. Biochem. Biophys. Res. Commun. 2017, 493, 847–854. [Google Scholar] [CrossRef]
- Ruddenklau, A.; Campbell, R.E. Neuroendocrine Impairments of Polycystic Ovary Syndrome. Endocrinology 2019, 160, 2230–2242. [Google Scholar] [CrossRef]
- Wissing, M.L.; Sonne, S.B.; Westergaard, D.; Nguyen, K.; Belling, K.; Host, T.; Mikkelsen, A.L. The transcriptome of corona radiata cells from individual Mcapital I, Ukrainiancapital I, Ukrainian oocytes that after ICSI developed to embryos selected for transfer: PCOS women compared to healthy women. J. Ovarian. Res. 2014, 7, 110. [Google Scholar] [CrossRef] [PubMed]
- Hooten, N.N.; Fitzpatrick, M.; Wood, W.H., 3rd; De, S.; Ejiogu, N.; Zhang, Y.; Mattison, J.A.; Becker, K.G.; Zonderman, A.B.; Evans, M.K. Age-related changes in microRNA levels in serum. Aging 2013, 5, 725–740. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, H.; Zhang, C.; Jing, Y.; Wang, C.; Liu, C.; Zhang, R.; Wang, J.; Zhang, J.; Zen, K.; et al. Investigation of microRNA expression in human serum during the aging process. J. Gerontol. A Biol. Sci. Med. Sci. 2015, 70, 102–109. [Google Scholar] [CrossRef]
- Hooten, N.N.; Martin-Montalvo, A.; Dluzen, D.F.; Zhang, Y.; Bernier, M.; Zonderman, A.B.; Becker, K.G.; Gorospe, M.; de Cabo, R.; Evans, M.K. Metformin-mediated increase in DICER1 regulates microRNA expression and cellular senescence. Aging Cell 2016, 15, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Udesen, P.B.; Glintborg, D.; Sorensen, A.E.; Svendsen, R.; Nielsen, N.L.S.; Wissing, M.L.M.; Andersen, M.S.; Englund, A.L.M.; Dalgaard, L.T. Metformin decreases miR-122, miR-223 and miR-29a in women with polycystic ovary syndrome. Endocr. Connect. 2020, 9, 1075–1084. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BL | FU | BL vs. FU | ||||||
|---|---|---|---|---|---|---|---|---|
| Control | PCOS | p | Control | PCOS | p | Control p | PCOS p | |
| Age (years) | 30.2 (4.9) | 28.5(4.2) | 36.0 (5.7) | 34.3 (4.3) | - | - | ||
| Weight (kg) | 74.6(13.5) | 77.2 (19.9) | 79.8 (12.1) | 78.2 (17.7) | - | - | ||
| Height (cm) | 169.8 (3.6) | 168.0 (6.8) | 170.1 (5.3) | 168.1 (6.7) | - | - | ||
| BMI (kg/m2) | 25.6 (4.7) | 26.8 (5.5) | 27.7 (4.4) | 27.7 (6.1) | * | * | ||
| Waist-Hip ratio | 0.78 (0.1) | 0.83 (0.1) | * | 0.85 (0.6) | 0.84 (0.1) | * | - | |
| Systolic Blood pressure (mmHg) | 124.1 (16.1) | 122.4 (10.9) | 116.2 (18.6) | 120.7 (13.5) | - | - | ||
| Diastolic Blood pressure (mmHg) | 78.9 (14.1) | 76.3 (10.3) | 72.7 (16.4) | 72.3 (11.2) | - | * | ||
| Metabolic Characteristics | ||||||||
| F-p-Glucose (mmol/L) | 5.3 (4.7–6.0) | 5.2 (5.0–5.4) | 4.7 (4.6–5.2) | 4.8 (4.6–5.1) | - | ** | ||
| F-s-Insulin (pmol/L) | 61.8 (44.9–75.9) | 45.8 (30.4–104.1) | 48.1 (37.0–67.0) | 70.0 (37.8–108.9) | - | ** | ||
| F-s-C-peptide (nmol/L) | 0.6 (0.6–0.7) | 0.6 (0.4–0.8) | 0.6 (0.6–0.7) | 0.7 (0.5–1.1) | - | *** | ||
| HOMA-IR (mU*mmol/L2) | 2.1 (1.5–2.6) | 1.5 (1.0–3.3) | 1.5 (1.1–1.9) | 1.9 (1.1–3.6) | - | * | ||
| HDL cholesterol (mmol/L) | 1.5 (1.3–1.6) | 1.5 (1.2–1.8) | 1.4 (1.3–1.4) | 1.4 (1.2–1.6) | - | - | ||
| LDL cholesterol (mmol/L) | 3.1 (2.6–3.5) | 2.6 (2.1–3.1) | 2.9 (2.1–3.1) | 2.4 (2.1–3.0) | - | - | ||
| Total cholesterol (mmol/L) | 4.9 (4.3–5.1) | 4.5 (4.1–5.0) | 4.6 (3.8–4.8) | 4.4 (4.0–5.0) | - | - | ||
| Triglyceride (mmol/L) | 0.7 (0.7–0.8) | 0.7 (0.6–1.2) | 0.9 (0.8–1.1) | 0.8 (0.7–1.5) | - | ** | ||
| Androgen Markers | ||||||||
| DHEAS (Umol/L) | 5.29 (1.86) | 6.27 (2.24) | 4.33 (2.41) | 4.81 (2.13) | - | ** | ||
| SHBG (nmol/L) | 71.0 (47.0–75.0) | 59.5 (36.0–89.0) | 53.0 (32.0–66.0) | 46.0 (35.0–82.0) | - | ** | ||
| Total testosterone (nmol/L) | 1.0 (0.8–1.5) | 1.9 (1.5–2.6) | *** | 0.9 (0.4–0.9) | 1.4 (0.9–2.0) | ** | * | *** |
| Free testosterone (nmol/L) | 0.017 (0.013–0.02) | 0.032 (0.019–0.05) | ** | 0.014 (0.010–0.015) | 0.023 (0.012–0.05) | - | ** | |
| Androstenedione (nmol/L) | 4.2 (3.4–5.2) | 6.9 (4.7–9.0) | ** | 2.6 (2.2–3.2) | 5.8 (4.2–8.9) | *** | *** | * |
| LH/FSH ratio | 0.9 (0.6–1.2) | 1.7 (1.2–2.4) | *** | 0.8 (0.8–1.1) | 1.5 (1.0–2.2) | * | - | * |
| Ferriman-Gallwey score | 2.0 (2.0–3.0) | 5.0 (3.0–9.5) | ** | 1.00 (0.0–5.0) | 6.0 (3.0–9.0) | ** | - | - |
| Lifestyle characteristics | ||||||||
| Metformin use (current/previous/no) | 9/22/14 | |||||||
| ART outcome successful (yes/no) | 7/2 | 38/8 | ||||||
| Smoking (current, previous, no) | 3/2/4 | 9/11/26 | ||||||
| Alcohol consumption (yes, no) | 3/6 | 10/36 | ||||||
| miRNA | Reported Association | Difference in dCt ± SD | Direction | n | p | padj |
|---|---|---|---|---|---|---|
| miR-451a | PCOS (circulating) | 6.3 ± 3.2 | Decreased | 34 | 5.7 × 10−13 | 5.5 × 10−11 |
| miR-24-3p | PCOS (FF) | 5.8 ± 5.2 | Decreased | 42 | 8.4 × 10−9 | 8.1 × 10−7 |
| miR-16-5p | PCOS | 5.3 ± 3.1 | Decreased | 39 | 4.8 × 10−13 | 4.6 × 10−11 |
| miR-21-5p | PCOS (circulating) | 4.6 ± 3 | Decreased | 27 | 2.2 × 10−8 | 2.1 × 10−6 |
| miR-142-3p | Dysregulated in PCOS granulosa cells | 4.5 ± 3.3 | Decreased | 24 | 8.4 × 10−7 | 8.1 × 10−5 |
| miR-223-3p | PCOS (circulating | 4.5 ± 3.2 | Decreased | 43 | 1.4 × 10−11 | 1.3 × 10−9 |
| miR-720 | PCOS (FF) | 4.1 ± 3.2 | Decreased | 44 | 1.1 × 10−10 | 1.1 × 10−8 |
| miR-19b-3p | PCOS (circulating) | 4 ± 3 | Decreased | 38 | 7.3 × 10−10 | 7.0 × 10−8 |
| miR-151-3p | PCOS (FF) | 3.8 ± 2 | Decreased | 36 | 2.2 × 10−13 | 2.1 × 10−11 |
| miR-342-3p | Circulating levels associated with insulin resistance | 3.7 ± 2.7 | Decreased | 35 | 2.3 × 10−9 | 2.2 × 10−7 |
| miR-146a-5p | PCOS (circulating) | 3.6 ± 2.5 | Decreased | 39 | 7.4 × 10−11 | 7.1 × 10−9 |
| miR-126-3p | PCOS (circulating) | 3.4 ± 2.6 | Decreased | 37 | 3.6 × 10−9 | 3.4 × 10−7 |
| miR-93-5p | PCOS (circulating) | 3.2 ± 2.5 | Decreased | 35 | 9.9 × 10−9 | 9.5 × 10−7 |
| miR-145-5p | PCOS (circulating) | 3.2 ± 2.1 | Decreased | 32 | 1.5 × 10−9 | 1.5 × 10−7 |
| miR-1305 | Circulating biomarkers of ovarian cancer | 3.1 ± 5.9 | Decreased | 24 | 1.6 × 10−2 | 1.0 |
| miR-320a-3p | PCOS (FF) | 3.1 ± 1.8 | Decreased | 42 | 3.2 × 10−14 | 3.1 × 10−12 |
| miR-151-5p | PCOS (FF) | 2.7 ± 4.2 | Decreased | 29 | 1.8 × 10−3 | 0.17 |
| miR-17-5p | Circulating biomarkers of metabolic syndrome | 2.7 ± 2.6 | Decreased | 41 | 9.8 × 10−8 | 9.4 × 10−6 |
| miR-423-5p | Dysregulated in PCOS granulosa cells | 2.5 ± 1.8 | Decreased | 33 | 4.0 × 10−9 | 3.8 × 10−7 |
| miR-139-5p | PCOS (circulating) | 2.5 ± 2.0 | Decreased | 30 | 2.4 × 10−7 | 2.3 × 10−5 |
| miR-28-3p | Granulosa cell proliferation | 2.3 ± 1.8 | Decreased | 25 | 7.9 × 10−7 | 7.6 × 10−5 |
| miR-30b-5p | Associated with NAFLD | 2.2 ± 2.4 | Decreased | 33 | 6.8 × 10−6 | 6.6 × 10−4 |
| miR-30c-5p | PCOS (circulating) | 2.2 ± 2.3 | Decreased | 37 | 1.2 × 10−6 | 1.2 × 10−4 |
| miR-20a-5p | PCOS (circulating) | 1.9 ± 2.9 | Decreased | 33 | 5.4 × 10−4 | 0.052 |
| miR-222-3p | PCOS (circulating) | 1.8 ± 2.5 | Decreased | 35 | 2.1 × 10−4 | 0.020 |
| miR-221-3p | Associated with inflammation | 1.7 ± 2.5 | Decreased | 26 | 1.4 × 10−3 | 0.130 |
| miR-1233-3p | Circulating biomarkers of ovarian cancer | 5.5 ± 7.1 | Increased | 28 | 3.2 × 10−4 | 0.031 |
| miR-518f-3p | PCOS (FF) | 6.6 ± 6.3 | Increased | 34 | 7.9 × 10−7 | 7.6 × 10−5 |
| miR-520c-3p | PCOS (FF) | 8.4 ± 8.1 | Increased | 30 | 3.7 × 10−6 | 3.6 × 10−4 |
| miR-618 | Not previously published | 10.3 ± 7.5 | Increased | 29 | 5.1 × 10−8 | 4.9 × 10−6 |
| Time Point | Clinical/Biochemical Marker | MiRNA | N | Spearman Rho (ρ) | p | padj. |
|---|---|---|---|---|---|---|
| BL | Total T | miR-139-5p | 34 | 0.50 | 0.002 | 0.013 |
| miR-28-3p | 31 | 0.37 | 0.043 | 0.090 | ||
| Free T | miR-139-5p | 34 | 0.47 | 0.005 | 0.051 | |
| FU | Waist-hip ratio | miR-376a-3p | 23 | 0.43 | 0.042 | 0.010 |
| Total T | miR-28-3p | 35 | 0.40 | 0.016 | 0.031 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Udesen, P.B.; Sørensen, A.E.; Svendsen, R.; Frisk, N.L.S.; Hess, A.L.; Aziz, M.; Wissing, M.L.M.; Englund, A.L.M.; Dalgaard, L.T. Circulating miRNAs in Women with Polycystic Ovary Syndrome: A Longitudinal Cohort Study. Cells 2023, 12, 983. https://doi.org/10.3390/cells12070983
Udesen PB, Sørensen AE, Svendsen R, Frisk NLS, Hess AL, Aziz M, Wissing MLM, Englund ALM, Dalgaard LT. Circulating miRNAs in Women with Polycystic Ovary Syndrome: A Longitudinal Cohort Study. Cells. 2023; 12(7):983. https://doi.org/10.3390/cells12070983
Chicago/Turabian StyleUdesen, Pernille B., Anja E. Sørensen, Rikke Svendsen, Nanna L. S. Frisk, Anne L. Hess, Mubeena Aziz, Marie Louise M. Wissing, Anne Lis M. Englund, and Louise T. Dalgaard. 2023. "Circulating miRNAs in Women with Polycystic Ovary Syndrome: A Longitudinal Cohort Study" Cells 12, no. 7: 983. https://doi.org/10.3390/cells12070983
APA StyleUdesen, P. B., Sørensen, A. E., Svendsen, R., Frisk, N. L. S., Hess, A. L., Aziz, M., Wissing, M. L. M., Englund, A. L. M., & Dalgaard, L. T. (2023). Circulating miRNAs in Women with Polycystic Ovary Syndrome: A Longitudinal Cohort Study. Cells, 12(7), 983. https://doi.org/10.3390/cells12070983