

Gal-1 Expression Analysis in the GLIOCAT Multicenter Study: Role as a Prognostic Factor and an Immune-Suppressive Biomarker

 , ,
, ,  , , ,
, , ,  , and add
Show full author list
, and add
Show full author list
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Ethical Details
2.3. Tissue Microarray Preparation
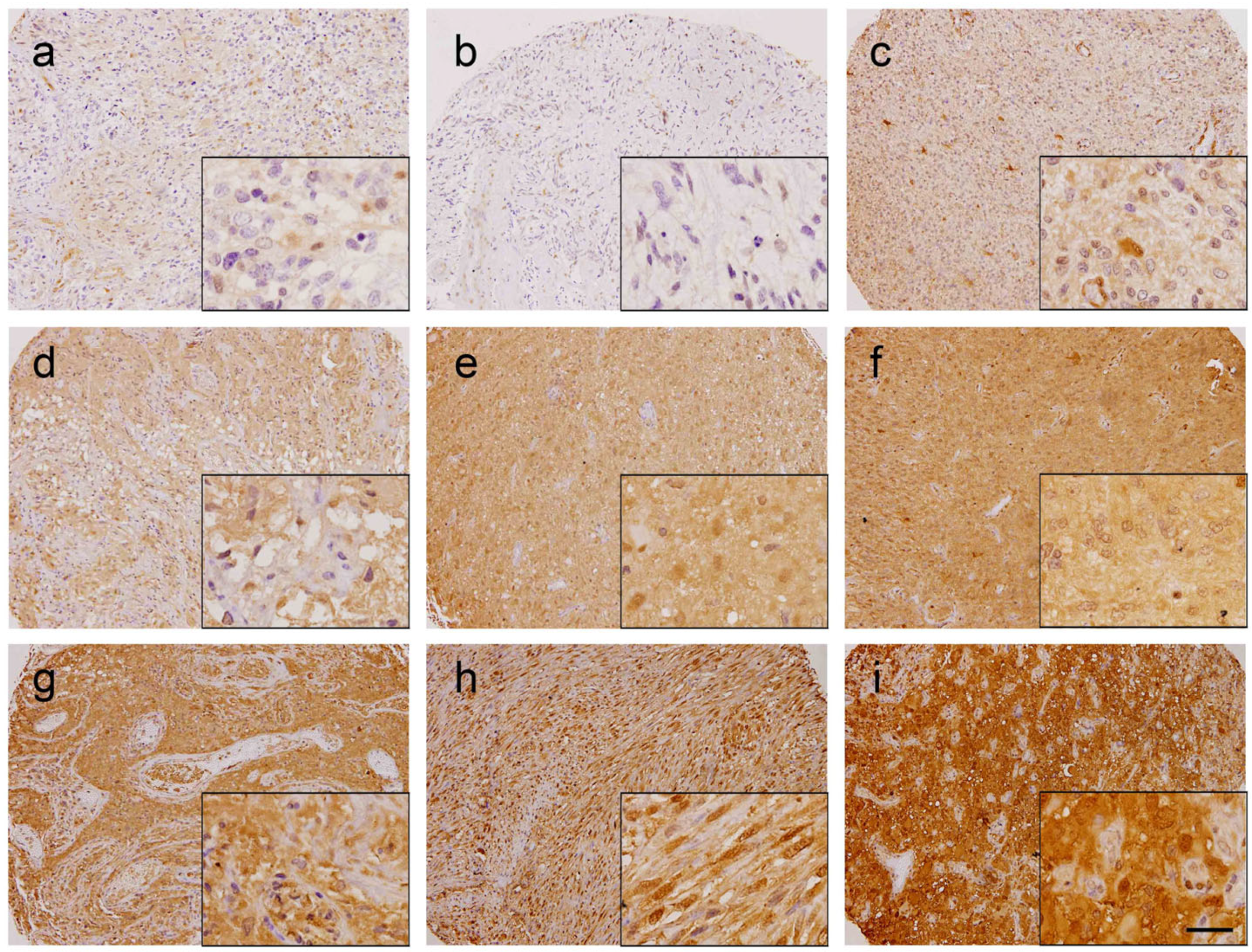
2.4. Immunohistochemistry Analysis
2.5. RNA Sequencing and GBM Molecular Subtypes
2.6. Cell-Type Deconvolution
2.7. Statistical Analysis
3. Results
3.1. Gal-1 Has Prognostic Value in GBM
3.2. Correlation between Gal-1 and Clinical or Molecular Data
3.3. Correlation between Gal-1 and GBM Molecular Subtypes
3.4. Correlation between Gal-1 Expression and Markers of Inflammation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cruz, J.V.R.; Batista, C.; Afonso, B.d.H.; Alexandre-Moreira, M.S.; Dubois, L.G.; Pontes, B.; Neto, V.M.; Mendes, F.d.A. Obstacles to Glioblastoma Treatment Two Decades after Temozolomide. Cancers 2022, 14, 3203. [Google Scholar] [CrossRef] [PubMed]
- Ostrom, Q.T.; Cote, D.J.; Ascha, M.; Kruchko, C.; Barnholtz-Sloan, J.S. Adult Glioma Incidence and Survival by Race or Ethnicity in the United States from 2000 to 2014. JAMA Oncol. 2018, 4, 1254–1262. [Google Scholar] [CrossRef]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.B.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus Concomitant and Adjuvant Temozolomide for Glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Taillibert, S.; Kanner, A.; Read, W.; Steinberg, D.M.; Lhermitte, B.; Toms, S.; Idbaih, A.; Ahluwalia, M.S.; Fink, K.; et al. Effect of Tumor-Treating Fields Plus Maintenance Temozolomide vs. Maintenance Temozolomide Alone on Survival in Patients With Glioblastoma: A Randomized Clinical Trial. JAMA 2017, 318, 2306–2316. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Waqas, M.; Shamim, M.S. Prognostic Significance of IDH 1 Mutation in Patients with Glioblastoma Multiforme. J. Pak. Med. Assoc. 2017, 67, 816–817. [Google Scholar] [PubMed]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A Summary. Neuro Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef]
- Audureau, E.; Chivet, A.; Ursu, R.; Corns, R.; Metellus, P.; Noel, G.; Zouaoui, S.; Guyotat, J.; le Reste, P.J.; Faillot, T.; et al. Prognostic Factors for Survival in Adult Patients with Recurrent Glioblastoma: A Decision-Tree-Based Model. J. Neurooncol. 2018, 136, 565–576. [Google Scholar] [CrossRef]
- Wang, Q.; Hu, B.; Hu, X.; Kim, H.; Squatrito, M.; Scarpace, L.; deCarvalho, A.C.; Lyu, S.; Li, P.; Li, Y.; et al. Tumor Evolution of Glioma-Intrinsic Gene Expression Subtypes Associates with Immunological Changes in the Microenvironment. Cancer Cell 2017, 32, 42–56.e6. [Google Scholar] [CrossRef]
- Gravendeel, L.A.M.; Kouwenhoven, M.C.M.; Gevaert, O.; de Rooi, J.J.; Stubbs, A.P.; Duijm, J.E.; Daemen, A.; Bleeker, F.E.; Bralten, L.B.C.; Kloosterhof, N.K.; et al. Intrinsic Gene Expression Profiles of Gliomas Are a Better Predictor of Survival than Histology. Cancer Res. 2009, 69, 9065–9072. [Google Scholar] [CrossRef]
- Nagy, Á.; Garzuly, F.; Padányi, G.; Szűcs, I.; Feldmann, Á.; Murnyák, B.; Hortobágyi, T.; Kálmán, B. Molecular Subgroups of Glioblastoma- an Assessment by Immunohistochemical Markers. Pathol. Oncol. Res. 2019, 25, 21–31. [Google Scholar] [CrossRef]
- Carrato, C.; Alameda, F.; Esteve-Codina, A.; Pineda, E.; Arpí, O.; Martinez-García, M.; Mallo, M.; Gut, M.; Lopez-Martos, R.; Barco, S.d.; et al. Glioblastoma TCGA Mesenchymal and IGS 23 Tumors Are Identifiable by IHC and Have an Immune-Phenotype Indicating a Potential Benefit from Immunotherapy. Clin. Cancer Res. 2020, 26, 6600–6609. [Google Scholar] [CrossRef]
- Camby, I.; le Mercier, M.; Lefranc, F.; Kiss, R. Galectin-1: A Small Protein with Major Functions. Glycobiology 2006, 16, 137R–157R. [Google Scholar] [CrossRef]
- Liu, F.T.; Rabinovich, G.A. Galectins as Modulators of Tumour Progression. Nat. Rev. Cancer 2005, 5, 29–41. [Google Scholar] [CrossRef]
- Thijssen, V.L.; Heusschen, R.; Caers, J.; Griffioen, A.W. Galectin Expression in Cancer Diagnosis and Prognosis: A Systematic Review. Biochim. Biophys. Acta 2015, 1855, 235–247. [Google Scholar] [CrossRef]
- Girotti, M.R.; Salatino, M.; Dalotto-Moreno, T.; Rabinovich, G.A. Sweetening the Hallmarks of Cancer: Galectins as Multifunctional Mediators of Tumor Progression. J. Exp. Med. 2020, 217, e20182041. [Google Scholar] [CrossRef]
- Chou, S.Y.; Yen, S.L.; Huang, C.C.; Huang, E.Y. Galectin-1 Is a Poor Prognostic Factor in Patients with Glioblastoma Multiforme after Radiotherapy. BMC Cancer 2018, 18, 105. [Google Scholar] [CrossRef]
- Camby, I.; Belot, N.; Lefranc, F.; Sadeghi, N.; de Launoit, Y.; Kaltner, H.; Musette, S.; Darro, F.; Danguy, A.; Salmon, I.; et al. Galectin-1 Modulates Human Glioblastoma Cell Migration into the Brain through Modifications to the Actin Cytoskeleton and Levels of Expression of Small GTPases. J. Neuropathol. Exp. Neurol. 2002, 61, 585–596. [Google Scholar] [CrossRef]
- D’Haene, N.; Maris, C.; Rorive, S.; Decaestecker, C.; le Mercier, M.; Salmon, I. Galectins and Neovascularization in Central Nervous System Tumors. Glycobiology 2014, 24, 892–898. [Google Scholar] [CrossRef]
- Chen, Q.; Han, B.; Meng, X.; Duan, C.; Yang, C.; Wu, Z.; Magafurov, D.; Zhao, S.; Safin, S.; Jiang, C.; et al. Immunogenomic Analysis Reveals LGALS1 Contributes to the Immune Heterogeneity and Immunosuppression in Glioma. Int. J. Cancer 2019, 145, 517–530. [Google Scholar] [CrossRef]
- Videla-Richardson, G.A.; Morris-Hanon, O.; Torres, N.I.; Esquivel, M.I.; Vera, M.B.; Ripari, L.B.; Croci, D.O.; Sevlever, G.E.; Rabinovich, G.A. Galectins as Emerging Glyco-Checkpoints and Therapeutic Targets in Glioblastoma. Int. J. Mol. Sci. 2021, 23, 316. [Google Scholar] [CrossRef]
- Le Mercier, M.; Fortin, S.; Mathieu, V.; Kiss, R.; Lefranc, F. Galectins and Gliomas. Brain Pathol. 2010, 20, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Esteve-Codina, A.; Alameda, F.; Carrato, C.; Pineda, E.; Arpí, O.; Martinez-García, M.; Mallo, M.; Gut, M.; Dabad, M.; Tortosa, A.; et al. RNA Sequencing and Immunohistochemistry Reveal ZFN7 as a Stronger Marker of Survival than Molecular Subtypes in G-CIMP-Negative Glioblastoma. Clin. Cancer Res. 2021, 27, 645–655. [Google Scholar] [CrossRef]
- Martínez-Bosch, N.; Manero-Rupérez, N.; Moreno, M.; Navarro, P. Exploring the Role of Galectins in Cancer: In Vitro and In Vivo Approaches. Methods Mol. Biol. 2022, 2442, 685–711. [Google Scholar] [CrossRef] [PubMed]
- Detre, S.; Saclani Jotti, G.; Dowsett, M. A “Quickscore” Method for Immunohistochemical Semiquantitation: Validation for Oestrogen Receptor in Breast Carcinomas. J. Clin. Pathol. 1995, 48, 876–878. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; Oshlack, A. A Scaling Normalization Method for Differential Expression Analysis of RNA-Seq Data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef]
- Chen, B.; Khodadoust, M.S.; Liu, C.L.; Newman, A.M.; Alizadeh, A.A. Profiling Tumor Infiltrating Immune Cells with CIBERSORT. Methods Mol. Biol. 2018, 1711, 243. [Google Scholar] [CrossRef]
- Hothorn, T.; Lausen, B. Maximally Selected Rank Statistics in R. R News, 1 February 2002; 3–5. [Google Scholar]
- Lausen, B.; Hothorn, T.; Bretz, F.; Schumacher, M. Assessment of Optimal Selected Prognostic Factors. Biom. J. 2004, 46, 364–374. [Google Scholar] [CrossRef]
- Budczies, J.; Klauschen, F.; Sinn, B.; Gyo Rffy, B.; Schmitt, W.D. Cutoff Finder: A Comprehensive and Straightforward Web Application Enabling Rapid Biomarker Cutoff Optimization. PLoS ONE 2012, 7, 51862. [Google Scholar] [CrossRef]
- Rabinovich, G.A. Galectin-1 as a Potential Cancer Target. Br. J. Cancer 2005, 92, 1188–1192. [Google Scholar] [CrossRef]
- Baker, G.J.; Chockley, P.; Yadav, V.N.; Doherty, R.; Ritt, M.; Sivaramakrishnan, S.; Castro, M.G.; Lowenstein, P.R. Natural Killer Cells Eradicate Galectin-1-Deficient Glioma in the Absence of Adaptive Immunity. Cancer Res. 2014, 74, 5079–5090. [Google Scholar] [CrossRef]
- Cooper, D.N.; Barondes, S.H. Evidence for Export of a Muscle Lectin from Cytosol to Extracellular Matrix and for a Novel Secretory Mechanism. J. Cell Biol. 1990, 110, 1681–1691. [Google Scholar] [CrossRef]
- Gao, Y.; Li, X.; Shu, Z.; Zhang, K.; Xue, X.; Li, W.; Hao, Q.; Wang, Z.; Zhang, W.; Wang, S.; et al. Nuclear Galectin-1-FOXP3 Interaction Dampens the Tumor-Suppressive Properties of FOXP3 in Breast Cancer. Cell Death Dis. 2018, 9, 416. [Google Scholar] [CrossRef]
- Patterson, R.J.; Wang, W.; Wang, J.L. Understanding the Biochemical Activities of Galectin-1 and Galectin-3 in the Nucleus. Glycoconj J. 2002, 19, 499–506. [Google Scholar] [CrossRef]
- Qi, C.; Lei, L.; Hu, J.; Wang, G.; Liu, J.; Ou, S. T Cell Immune Regulator 1 Is a Prognostic Marker Associated with Immune Infiltration in Glioblastoma Multiforme. Oncol. Lett. 2021, 21, 252. [Google Scholar] [CrossRef]
- Phillips, H.S.; Kharbanda, S.; Chen, R.; Forrest, W.F.; Soriano, R.H.; Wu, T.D.; Misra, A.; Nigro, J.M.; Colman, H.; Soroceanu, L.; et al. Molecular Subclasses of High-Grade Glioma Predict Prognosis, Delineate a Pattern of Disease Progression, and Resemble Stages in Neurogenesis. Cancer Cell 2006, 9, 157–173. [Google Scholar] [CrossRef]
- Verhaak, R.G.W.; Hoadley, K.A.; Purdom, E.; Wang, V.; Qi, Y.; Wilkerson, M.D.; Miller, C.R.; Ding, L.; Golub, T.; Mesirov, J.P.; et al. Integrated Genomic Analysis Identifies Clinically Relevant Subtypes of Glioblastoma Characterized by Abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 2010, 17, 98–110. [Google Scholar] [CrossRef]
- Van Woensel, M.; Mathivet, T.; Wauthoz, N.; Rosière, R.; Garg, A.D.; Agostinis, P.; Mathieu, V.; Kiss, R.; Lefranc, F.; Boon, L.; et al. Sensitization of Glioblastoma Tumor Micro-Environment to Chemo- and Immunotherapy by Galectin-1 Intranasal Knock-down Strategy. Sci. Rep. 2017, 7, 1217. [Google Scholar] [CrossRef]
- Correa, S.G.; Sotomayor, C.E.; Aoki, M.P.; Maldonado, C.A.; Rabinovich, G.A. Opposite Effects of Galectin-1 on Alternative Metabolic Pathways of L-Arginine in Resident, Inflammatory, and Activated Macrophages. Glycobiology 2003, 13, 119–128. [Google Scholar] [CrossRef]
- Barrionuevo, P.; Beigier-Bompadre, M.; Ilarregui, J.M.; Toscano, M.A.; Bianco, G.A.; Isturiz, M.A.; Rabinovich, G.A. A Novel Function for Galectin-1 at the Crossroad of Innate and Adaptive Immunity: Galectin-1 Regulates Monocyte/Macrophage Physiology through a Nonapoptotic ERK-Dependent Pathway. J. Immunol. 2007, 178, 436–445. [Google Scholar] [CrossRef]
- Rostoker, R.; Yaseen, H.; Schif-Zuck, S.; Lichtenstein, R.G.; Rabinovich, G.A.; Ariel, A. Galectin-1 Induces 12/15-Lipoxygenase Expression in Murine Macrophages and Favors Their Conversion toward a pro-Resolving Phenotype. Prostaglandins Other Lipid Mediat. 2013, 107, 85–94. [Google Scholar] [CrossRef]
- Malik, R.K.J.; Ghurye, R.R.; Lawrence-Watt, D.J.; Stewart, H.J.S. Galectin-1 Stimulates Monocyte Chemotaxis via the P44/42 MAP Kinase Pathway and a Pertussis Toxin-Sensitive Pathway. Glycobiology 2009, 19, 1402–1407. [Google Scholar] [CrossRef]
- Shao, R.; Taylor, S.L.; Oh, D.S.; Schwartz, L.M. Vascular Heterogeneity and Targeting: The Role of YKL-40 in Glioblastoma Vascularization. Oncotarget 2015, 6, 40507–40518. [Google Scholar] [CrossRef] [PubMed]
- Faibish, M.; Francescone, R.; Bentley, B.; Yan, W.; Shao, R. A YKL-40-Neutralizing Antibody Blocks Tumor Angiogenesis and Progression: A Potential Therapeutic Agent in Cancers. Mol. Cancer Ther. 2011, 10, 742–751. [Google Scholar] [CrossRef] [PubMed]
- Francescone, R.A.; Scully, S.; Faibish, M.; Taylor, S.L.; Oh, D.; Moral, L.; Yan, W.; Bentley, B.; Shao, R. Role of YKL-40 in the Angiogenesis, Radioresistance, and Progression of Glioblastoma. J. Biol. Chem. 2011, 286, 15332–15343. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Jun, Y.J.; Kim, T.H.; Jung, J.Y.; Hwang, G.H.; Jung, K.J.; Lee, S.H.; Lee, H.M.; Lee, S.H. Increased Expression of YKL-40 in Mild and Moderate/Severe Persistent Allergic Rhinitis and Its Possible Contribution to Remodeling of Nasal Mucosa. Am. J. Rhinol. Allergy 2013, 27, 372–380. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HR | CI 95% | p Value | |
|---|---|---|---|
| Gal-1 mean H-score | |||
| <129.3 | 1 | ||
| ≥129.3 | 1.52 | 1.10–2.08 | 0.010 |
| Age | |||
| <70 | 1 | ||
| ≥70 | 1.45 | 1.06–1.98 | 0.019 |
| Gender | |||
| Male | 1 | ||
| Female | 0.90 | 0.69–1.17 | 0.428 |
| Clinical and Molecular Variables | n | Median Gal-1 | Percentiles (P25–P75) | p Value * |
|---|---|---|---|---|
| KPS | ||||
| <70% | 23 | 87.38 | (62.50–128.40) | 0.495 |
| ≥70% | 192 | 94.31 | (71.30–126.03) | |
| MGMT status | ||||
| Methylated | 123 | 93.63 | (66.88–127.92) | 0.503 |
| Non-methylated | 133 | 92.82 | (71.89–129.17) | |
| IDH-1 | ||||
| No mutated | 263 | 93.13 | (68.44–126.25) | <0.001 |
| Mutated | 10 | 7.28 | (3.68–29.57) | |
| Classical | Mesenchymal | Proneural | p Value * | |
|---|---|---|---|---|
| n | 36 | 13 | 22 | |
| Gal-1 median (P25-P75) | 98.60 (67.69–111.15) | 106.67 (93.28–151.46) | 66.58 (26.32–90.42) | <0.001 |
| Gal-1 Mean | Spearman Correlation Coefficient | p Value * |
|---|---|---|
| SHC1 | 0.230 | <0.001 |
| PD-L1 | 0.239 | <0.001 |
| PAX2 | 0.155 | 0.014 |
| MEOX2 | 0.159 | 0.012 |
| YKL-40 | 0.182 | 0.004 |
| TCIRG1 | 0.151 | 0.017 |
| YWHAG | 0.232 | <0.001 |
| OLIG2 | −0.313 | <0.001 |
| SOX2 | −0.192 | 0.003 |
| Ki-67 | −0.283 | <0.001 |
| SOX11 | −0.278 | <0.001 |
| Immune cell Type | LGALS1 Correlation | p Value * |
|---|---|---|
| B cells, naive | 0.032 | 0.733 |
| B cells, memory | −0.124 | 0.182 |
| Plasma cells | 0.038 | 0.681 |
| T cells CD8 | −0.062 | 0.505 |
| T cells CD4, naive | −0.193 | 0.036 |
| T cells CD4 memory, resting | 0.454 | <0.001 |
| T cells CD4 memory, activated | 0.105 | 0.255 |
| T cells folicular, helper | −0.035 | 0.707 |
| T cells regulatory | 0.104 | 0.263 |
| T cells gamma delta | −0.019 | 0.835 |
| NK cells resting | 0.116 | 0.212 |
| NK cells activated | −0.150 | 0.105 |
| Monocytes | 0.243 | 0.008 |
| Macrophages M0 | 0.119 | 0.198 |
| Macrophages M1 | 0.018 | 0.843 |
| Macrophages M2 | 0.437 | <0.001 |
| Dendritic cells, resting | 0.055 | 0.550 |
| Dendritic cells, activated | 0.163 | 0.078 |
| Mast cells, resting | 0.005 | 0.956 |
| Mast cells, activated | 0.106 | 0.253 |
| Eosinophils | 0.111 | 0.232 |
| Neutrophils | 0.067 | 0.474 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Bosch, N.; Vilariño, N.; Alameda, F.; Mojal, S.; Arumí-Uria, M.; Carrato, C.; Aldecoa, I.; Ribalta, T.; Vidal, N.; Bellosillo, B.; et al. Gal-1 Expression Analysis in the GLIOCAT Multicenter Study: Role as a Prognostic Factor and an Immune-Suppressive Biomarker. Cells 2023, 12, 843. https://doi.org/10.3390/cells12060843
Martínez-Bosch N, Vilariño N, Alameda F, Mojal S, Arumí-Uria M, Carrato C, Aldecoa I, Ribalta T, Vidal N, Bellosillo B, et al. Gal-1 Expression Analysis in the GLIOCAT Multicenter Study: Role as a Prognostic Factor and an Immune-Suppressive Biomarker. Cells. 2023; 12(6):843. https://doi.org/10.3390/cells12060843
Chicago/Turabian StyleMartínez-Bosch, Neus, Noelia Vilariño, Francesc Alameda, Sergi Mojal, Montserrat Arumí-Uria, Cristina Carrato, Iban Aldecoa, Teresa Ribalta, Noemí Vidal, Beatriz Bellosillo, and et al. 2023. "Gal-1 Expression Analysis in the GLIOCAT Multicenter Study: Role as a Prognostic Factor and an Immune-Suppressive Biomarker" Cells 12, no. 6: 843. https://doi.org/10.3390/cells12060843
APA StyleMartínez-Bosch, N., Vilariño, N., Alameda, F., Mojal, S., Arumí-Uria, M., Carrato, C., Aldecoa, I., Ribalta, T., Vidal, N., Bellosillo, B., Menéndez, S., Del Barco, S., Gallego, O., Pineda, E., López-Martos, R., Hernández, A., Mesia, C., Esteve-Codina, A., de la Iglesia, N., ... Navarro, P. (2023). Gal-1 Expression Analysis in the GLIOCAT Multicenter Study: Role as a Prognostic Factor and an Immune-Suppressive Biomarker. Cells, 12(6), 843. https://doi.org/10.3390/cells12060843