

Olfactory Regeneration with Nasally Administered Murine Adipose-Derived Stem Cells in Olfactory Epithelium Damaged Mice

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Methimazole Administration
2.3. Nasal Administration of GFP-Positive Murine ADSCs
2.4. Assessment of Innate Odor Aversion Behavior to Butyric Acid in Mice
2.5. Immunohistochemical Analysis of Olfactory Epithelial Cells and ADSCs
2.6. ADSC Culture Supernatant Analysis by Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Statistical Analysis
3. Results
3.1. Odor Aversion Behavior Analysis in Mice with Olfactory Damage
3.2. Immunohistochemical Analysis of Olfactory Marker Protein Expression in the Nasal Epithelium of Mice on Day 14 after Left Side Nasal Administration of ADSCs
3.3. Immunohistochemical Analysis of the Olfactory Epithelium in Mice 24 h, 3 d, and 7 d after Left Side Nasal Administration of ADSCs
3.4. Neurotrophic Factors in ADSC Culture Medium Supernatant
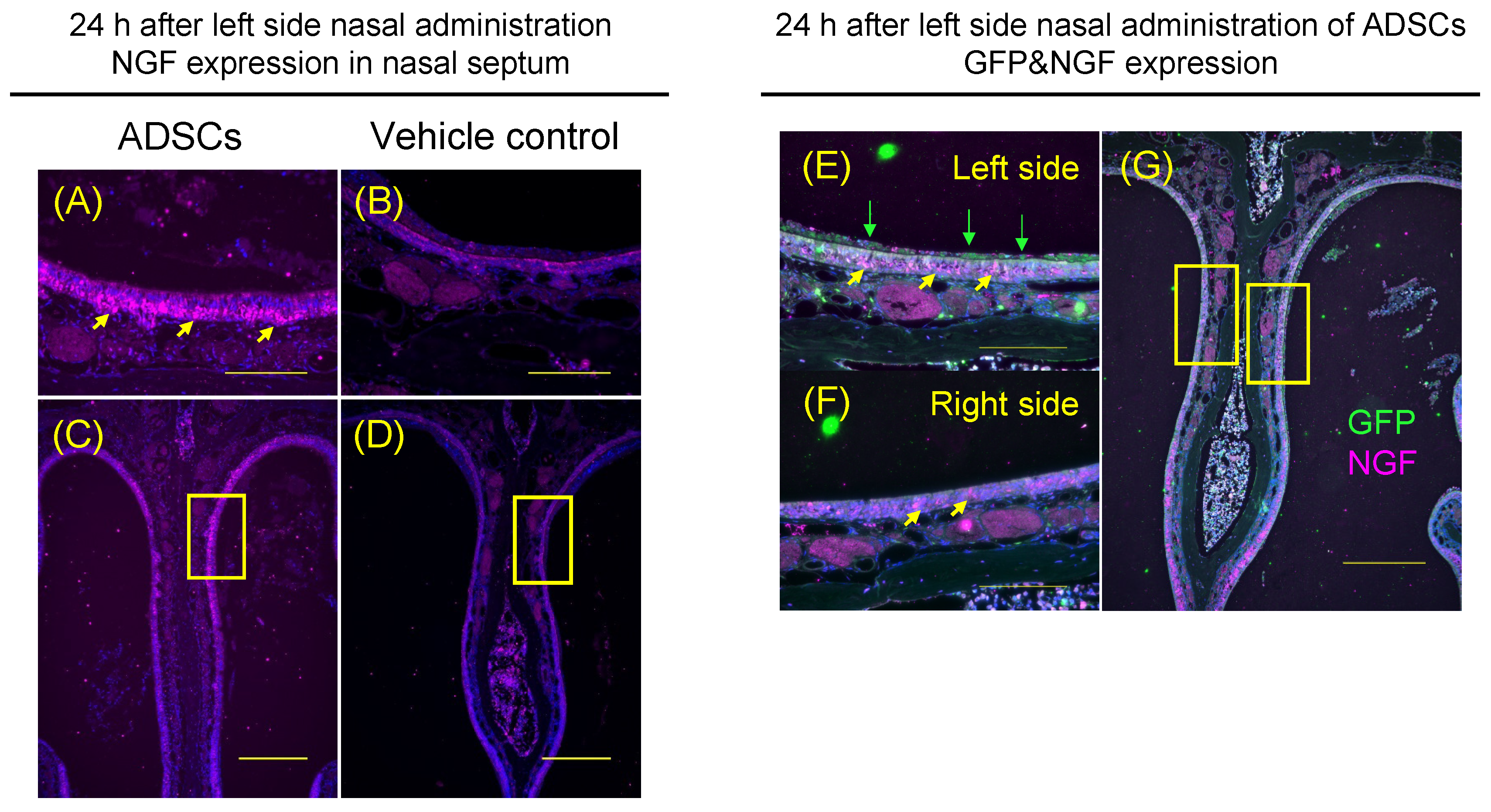
3.5. Immunohistochemical Analysis of Nerve Growth Factor Expression in Olfactory Epithelium with GFP-Positive ADSCs Adhering to the Surface of Nasal Epithelium in Mice after Left Side Nasal Administration of ADSCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goncalves, S.; Goldstein, B.J. Pathophysiology of olfactory disorders and potential treatment strategies. Curr. Otorhinolaryngol. Rep. 2016, 4, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Miwa, T.; Moriizumi, T.; Horikawa, I.; Uramoto, N.; Ishimaru, T.; Nishimura, T.; Furukawa, M. Role of nerve growth factor in the olfactory system. Microsc. Res. Tech. 2002, 58, 197–203. [Google Scholar] [CrossRef]
- Scolnick, J.A.; Cui, K.; Duggan, C.D.; Xuan, S.; Yuan, X.B.; Efstratiadis, A.; Ngai, J. Role of IGF signaling in olfactory sensory map formation and axon guidance. Neuron 2008, 57, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Umehara, T.; Udagawa, J.; Takamura, K.; Kimura, M.; Ishimitsu, R.; Kiyono, H.; Kawauchi, H.; Otani, H. Role of interleukin-15 in the development of mouse olfactory nerve. Congenit. Anom. 2009, 49, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Frontera, J.L.; Cervino, A.S.; Jungblut, L.D.; Paz, D.A. Brain-derived neurotrophic factor (BDNF) expression in normal and regenerating olfactory epithelium of Xenopus laevis. Ann. Anat. 2015, 198, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Tomita, K.; Madura, T.; Sakai, Y.; Yano, K.; Terenghi, G.; Hosokawa, K. Glial differentiation of human adipose-derived stem cells: Implications for cell-based transplantation therapy. Neuroscience 2013, 236, 55–65. [Google Scholar] [CrossRef]
- Kim, Y.M.; Choi, Y.S.; Choi, J.W.; Park, Y.H.; Koo, B.S.; Roh, H.J.; Rha, K.S. Effects of systemic transplantation of adipose tissue-derived stem cells on olfactory epithelium regeneration. Laryngoscope 2009, 119, 993–999. [Google Scholar] [CrossRef] [PubMed]
- Franceschini, V.; Bettini, S.; Pifferi, S.; Menini, A.; Siciliano, G.; Ognio, E.; Brini, A.T.; Di Oto, E.; Revoltella, R.P. Transplanted human adipose tissue-derived stem cells engraft and induce regeneration in mice olfactory neuroepithelium in response to dichlobenil subministration. Chem. Sens. 2014, 39, 617–629. [Google Scholar] [CrossRef]
- Mizuno, H.; Tobita, M.; Uysal, A.C. Concise review: Adipose-derived stem cells as a novel tool for future regenerative medicine. Stem. Cells. 2012, 30, 804–810. [Google Scholar] [CrossRef]
- Kuroda, Y.; Kitada, M.; Wakao, S.; Nishikawa, K.; Tanimura, Y.; Makinoshima, H.; Goda, M.; Akashi, H.; Inutsuka, A.; Niwa, A.; et al. Unique multipotent cells in adult human mesenchymal cell populations. Proc. Natl. Acad. Sci. USA 2010, 107, 8639–8643. [Google Scholar] [CrossRef]
- Jung, J.W.; Kwon, M.; Choi, J.C.; Shin, J.W.; Park, I.W.; Choi, B.W.; Kim, J.Y. Familial occurrence of pulmonary embolism after intravenous, adipose tissue-derived stem cell therapy. Yonsei Med. J. 2013, 54, 1293–1296. [Google Scholar] [CrossRef]
- Beites, C.L.; Kawauchi, S.; Crocker, C.E.; Calof, A.L. Identification and molecular regulation of neural stem cells in the olfactory epithelium. Exp. Cell Res. 2005, 306, 309–316. [Google Scholar] [CrossRef]
- Ueda, T.; Sakamoto, T.; Kobayashi, M.; Kuwata, F.; Ishikawa, M.; Omori, K.; Nakagawa, T. Optical coherence tomography for observation of the olfactory epithelium in mice. Auris Nasus Larynx 2019, 46, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Kondo, K.; Kashio, A.; Suzukawa, K.; Yamasoba, T. Methimazole-induced cell death in rat olfactory receptor neurons occurs via apoptosis triggered through mitochondrial cytochrome c-mediated caspase-3 activation pathway. J. Neurosci. Res. 2007, 85, 548–557. [Google Scholar] [CrossRef]
- Sasajima, H.; Miyazono, S.; Noguchi, T.; Kashiwayanagi, M. Intranasal administration of rotenone in mice attenuated olfactory functions through the lesion of dopaminergic neurons in the olfactory bulb. Neurotoxicology 2015, 51, 106–115. [Google Scholar] [CrossRef]
- Buiakova, O.I.; Baker, H.; Scott, J.W.; Farbman, A.; Kream, R.; Grillo, M.; Franzen, L.; Richman, M.; Davis, L.M.; Abbondanzo, S.; et al. Olfactory marker protein (OMP) gene deletion causes altered physiological activity of olfactory sensory neurons. Proc. Natl. Acad. Sci. USA 1996, 93, 9858–9863. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Packard, A.; Krolewski, R.C.; Harris, M.T.; Manglapus, G.L.; Schwob, J.E. Expression of pax6 and sox2 in adult olfactory epithelium. J. Comp. Neurol. 2010, 518, 4395–4418. [Google Scholar] [CrossRef]
- Miwa, T.; Horikawa, I.; Uramoto, N.; Ishimaru, T.; Yamamoto, K.; Furukawa, M.; Kato, T.; Moriizumi, T. TrkA expression in mouse olfactory tract following axotomy of olfactory nerves. Acta Otolaryngol. Suppl. 1998, 539, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Shiga, H.; Noda, T.; Harita, M.; Ishikura, T.; Nakamura, Y.; Hatta, T.; Sakata-Haga, H.; Shimada, H.; Miwa, T. The impact of ovariectomy on olfactory neuron regeneration in mice. Chem. Sens. 2020, 45, 203–209. [Google Scholar] [CrossRef]
- Miwa, T.; Uramoto, N.; Ishimaru, T.; Furukawa, M.; Shiba, K.; Morjizumi, T. Retrograde transport of nerve growth factor from olfactory bulb to olfactory epithelium. NeuroReport 1998, 9, 153–155. [Google Scholar] [CrossRef]
- Aloe, L.; Rocco, M.L.; Balzamino, B.O.; Micera, A. Nerve growth factor: A focus on neuroscience and therapy. Curr. Neuropharmacol. 2015, 13, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Chiaretti, A.; Conti, G.; Falsini, B.; Buonsenso, D.; Crasti, M.; Manni, L.; Soligo, M.; Fantacci, C.; Genovese, O.; Calcagni, M.L.; et al. Intranasal nerve growth factor administration improves cerebral functions in a child with severe traumatic brain injury: A case report. Brain Inj. 2017, 31, 1538–1547. [Google Scholar] [CrossRef] [PubMed]
- Petty, B.G.; Cornblath, D.R.; Adornato, B.T.; Chaudhry, V.; Flexner, C.; Wachsman, M.; Sinicropi, D.; Burton, L.E.; Peroutka, S.J. The effect of systemically administered recombinant human nerve growth factor in healthy human subjects. Ann. Neurol. 1994, 36, 244–246. [Google Scholar] [CrossRef] [PubMed]
- Tria, M.A.; Fusco, M.; Vantini, G.; Mariot, R. Pharmacokinetics of nerve growth factor (NGF) following different routes of administration to adult rats. Exp. Neurol. 1994, 127, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Poduslo, J.F.; Curran, G.L. Permeability at the blood-brain and blood-nerve barriers of the neurotrophic factors: NGF, CNTF, NT-3, BDNF. Brain Res. Mol. Brain Res. 1996, 36, 280–286. [Google Scholar] [CrossRef]
- Fukuda, Y.; Katsunuma, S.; Uranagase, A.; Nota, J.; Nibu, K.I. Effect of intranasal administration of neurotrophic factors on regeneration of chemically degenerated olfactory epithelium in aging mice. Neuroreport 2018, 29, 1400–1404. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scored Part | ADSCs in PBS (n = 6) | Control (PBS) (n = 6) | p-Value (Mann–Whitney Test) |
|---|---|---|---|
| Left side 1 | 81.0 (60.3–99.3) | 17.0 (5.8–25.5) | 0.002 |
| Left side 2 | 97.0 (85.0–123.5) | 104.0 (58.0–161.0) | >0.999 |
| Left side 3 | 87.5 (77.8–105.0) | 77.0 (57.0–91.5) | 0.320 |
| Right side 1 | 80.0 (62.5–85.5) | 9.0 (4.8–23.5) | 0.002 |
| Right side 2 | 106.5 (82.0–118.0) | 118.0 (61.5–153.5) | 0.650 |
| Right side 3 | 108.5 (84.3–134.5) | 71.0 (49.5–108.5) | 0.120 |
| Scored Part 1 | ADSCs in PBS (n = 5) | Control (PBS) (n = 5) | p-Value (Mann–Whitney Test) | |
|---|---|---|---|---|
| 24 h | Left side | 0.0 (0.0–7.5) | 0.0 (0.0–2.5) | 0.723 |
| Right side | 1.0 (0.5–2.0) | 0.0 (0.0–1.5) | 0.370 | |
| 3 d | Left side | 4.0 (1.5–7.5) | 2.0 (0.5–12.0) | 0.952 |
| Right side | 2.0 (1.0–4.5) | 6.0 (1.5–10.5) | 0.238 | |
| 7 d | Left side | 33.0 (14.0–38.5) | 9.0 (5.5–14.5) | 0.032 |
| Right side | 27.0 (16.5–47.0) | 2.0 (1.5–13.5) | 0.016 | |
| Scored Part 1 | ADSCs in PBS (n = 5) | Control (PBS) (n = 5) | p-Value (Mann–Whitney Test) | |
|---|---|---|---|---|
| 24 h | Left side | 2.0 (0.5–5.5) | 53.0 (21.0–58.5) | 0.008 |
| Right side | 4.0 (0.5–10.0) | 70.0 (20.5–81.5) | 0.008 | |
| 3 d | Left side | 2.0 (0.5–22.5) | 8.0 (6.0–16.0) | 0.643 |
| Right side | 2.0 (1.5–17.5) | 5.0 (4.0–18.5) | 0.341 | |
| 7 d | Left side | 5.0 (3.0–22.5) | 9.0 (5.0–27.5) | 0.651 |
| Right side | 3.0 (0.5–13.5) | 17.0 (4.5–22.5) | 0.151 | |
| Marker (pg/mL) | ADSC Culture Medium (n = 3) | Control Culture Medium (n = 3) |
|---|---|---|
| Mature NGF | 136.1 (74.7–246.8) | N.D. |
| Mature BDNF | 17.9 (5.1–23.8) | N.D. |
| IGF-1 | N.D. | N.D. |
| IL-15 & IL-15R | N.D. | N.D. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishikura, T.; Shiga, H.; Nakamura, Y.; Kanitani, T.; Ishigaki, Y.; Miwa, T. Olfactory Regeneration with Nasally Administered Murine Adipose-Derived Stem Cells in Olfactory Epithelium Damaged Mice. Cells 2023, 12, 765. https://doi.org/10.3390/cells12050765
Ishikura T, Shiga H, Nakamura Y, Kanitani T, Ishigaki Y, Miwa T. Olfactory Regeneration with Nasally Administered Murine Adipose-Derived Stem Cells in Olfactory Epithelium Damaged Mice. Cells. 2023; 12(5):765. https://doi.org/10.3390/cells12050765
Chicago/Turabian StyleIshikura, Tomoko, Hideaki Shiga, Yuka Nakamura, Takako Kanitani, Yasuhito Ishigaki, and Takaki Miwa. 2023. "Olfactory Regeneration with Nasally Administered Murine Adipose-Derived Stem Cells in Olfactory Epithelium Damaged Mice" Cells 12, no. 5: 765. https://doi.org/10.3390/cells12050765
APA StyleIshikura, T., Shiga, H., Nakamura, Y., Kanitani, T., Ishigaki, Y., & Miwa, T. (2023). Olfactory Regeneration with Nasally Administered Murine Adipose-Derived Stem Cells in Olfactory Epithelium Damaged Mice. Cells, 12(5), 765. https://doi.org/10.3390/cells12050765