
Gbb Regulates Blood Cell Proliferation and Differentiation through JNK and EGFR Signaling Pathways in the Drosophila Lymph Gland


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Fly Stocks and Culture
2.2. Immunostaining
2.3. Image Analysis and Quantification
2.4. Quantitative Real-Time PCR
2.5. Statistical Analysis
3. Results
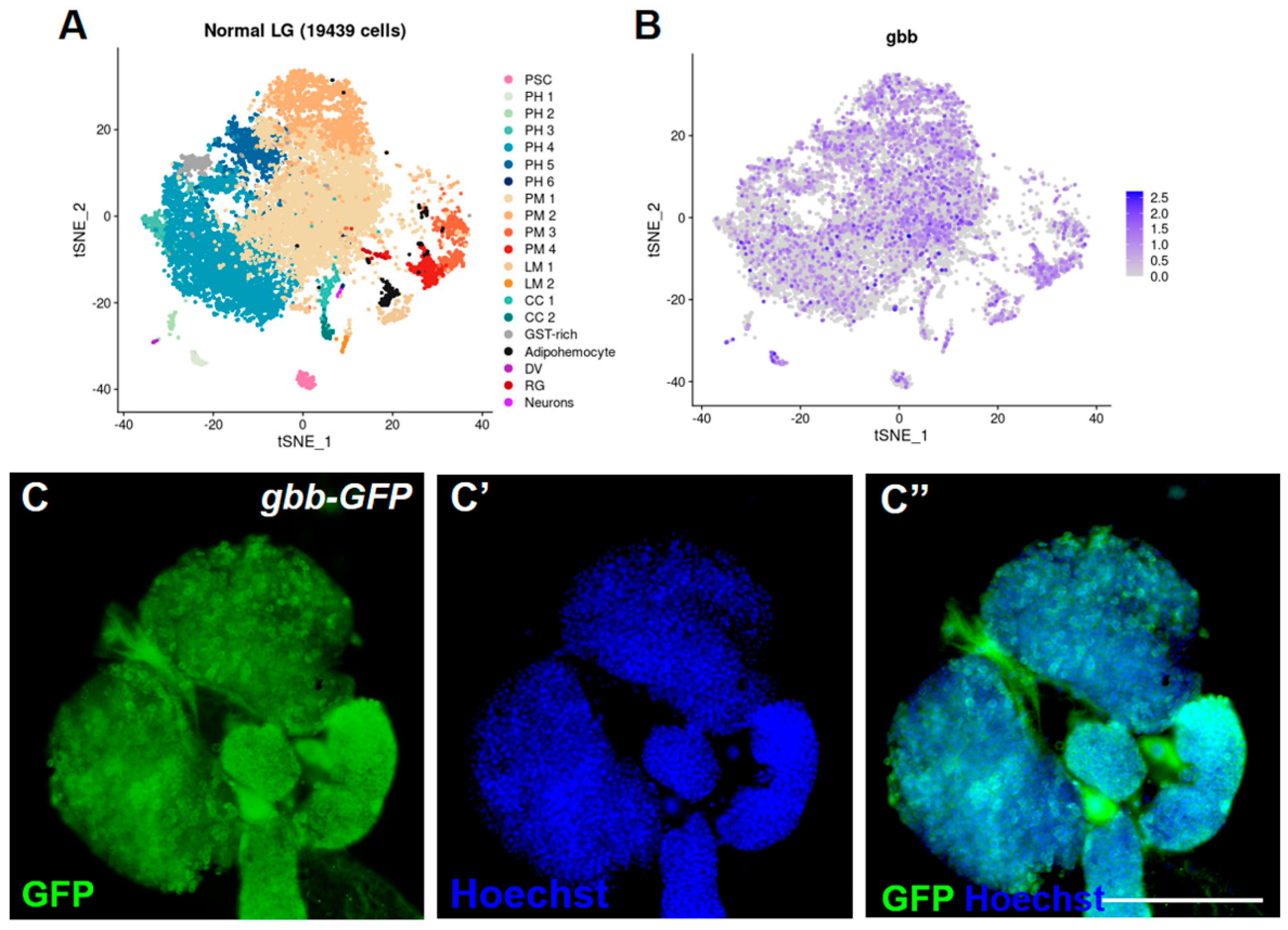
3.1. Gbb Is Widely Expressed in the Drosophila Lymph Gland
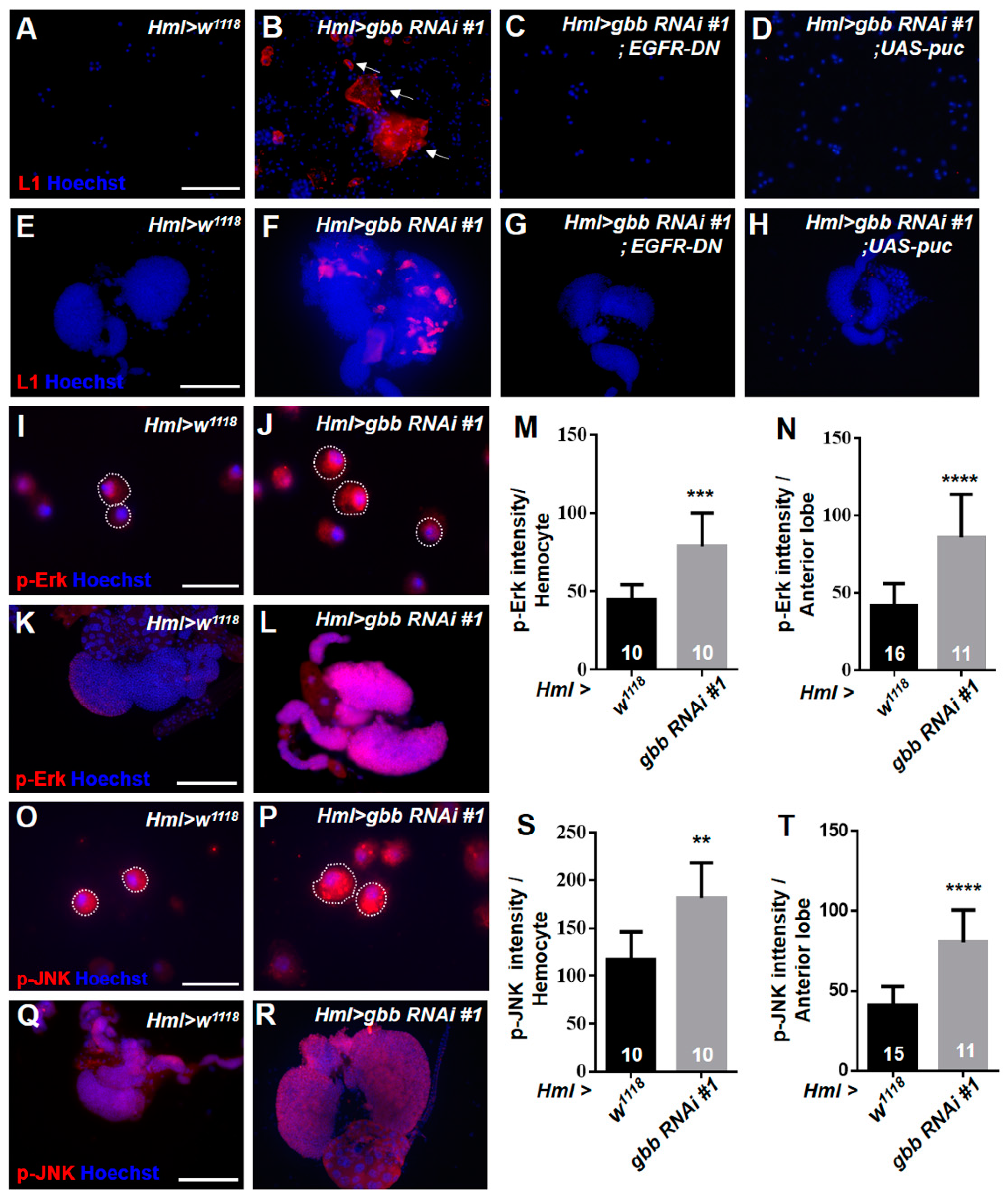
3.2. Knockdown of Gbb in the CZ Can Induce Lamellocyte Differentiation by Activating the EGFR and JNK Pathways
3.3. The Loss of Gbb in the CZ Can Induce Plasmatocyte Differentiation Primarily by Activating the EGFR Pathway but Not the JNK Pathway
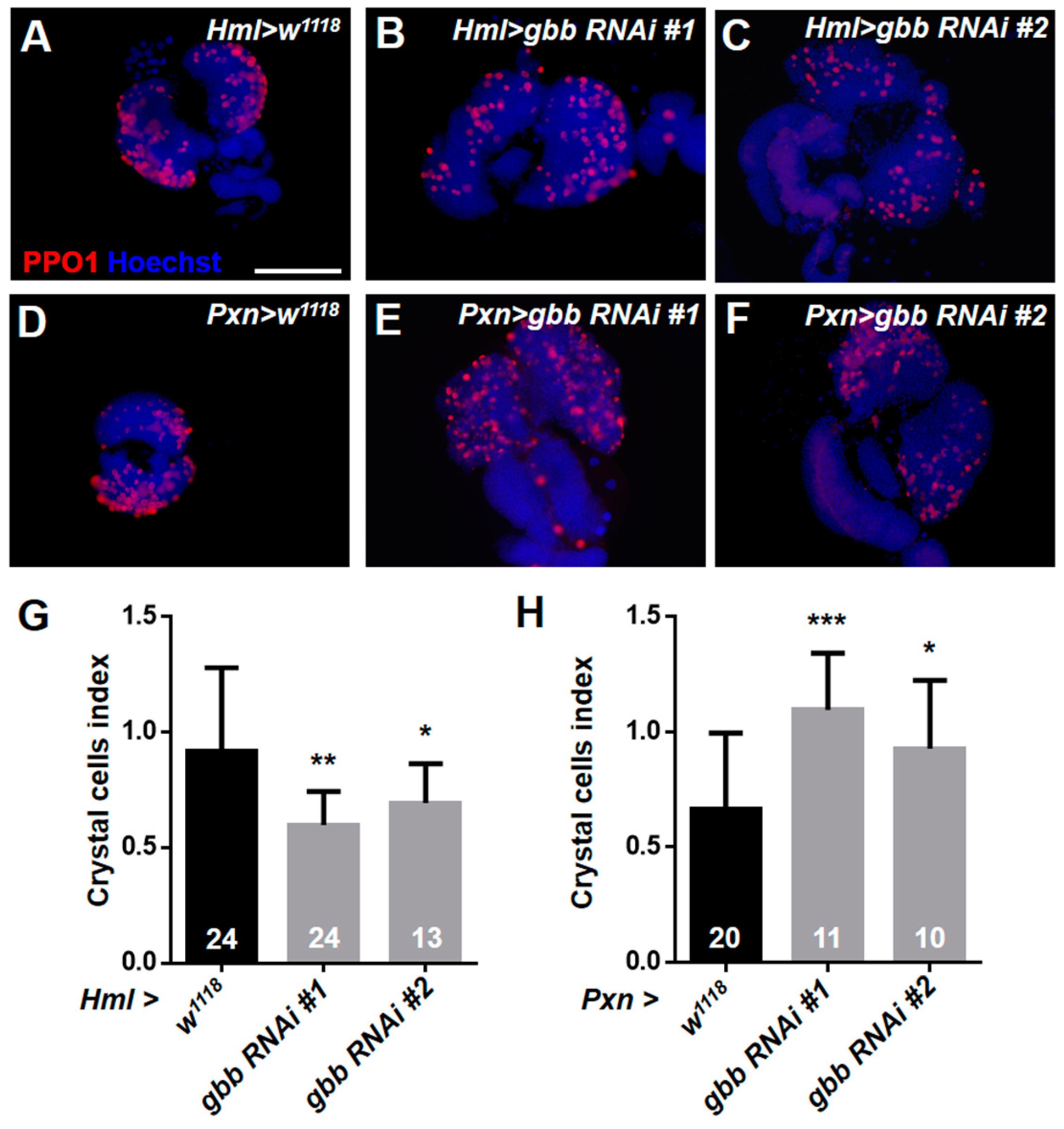
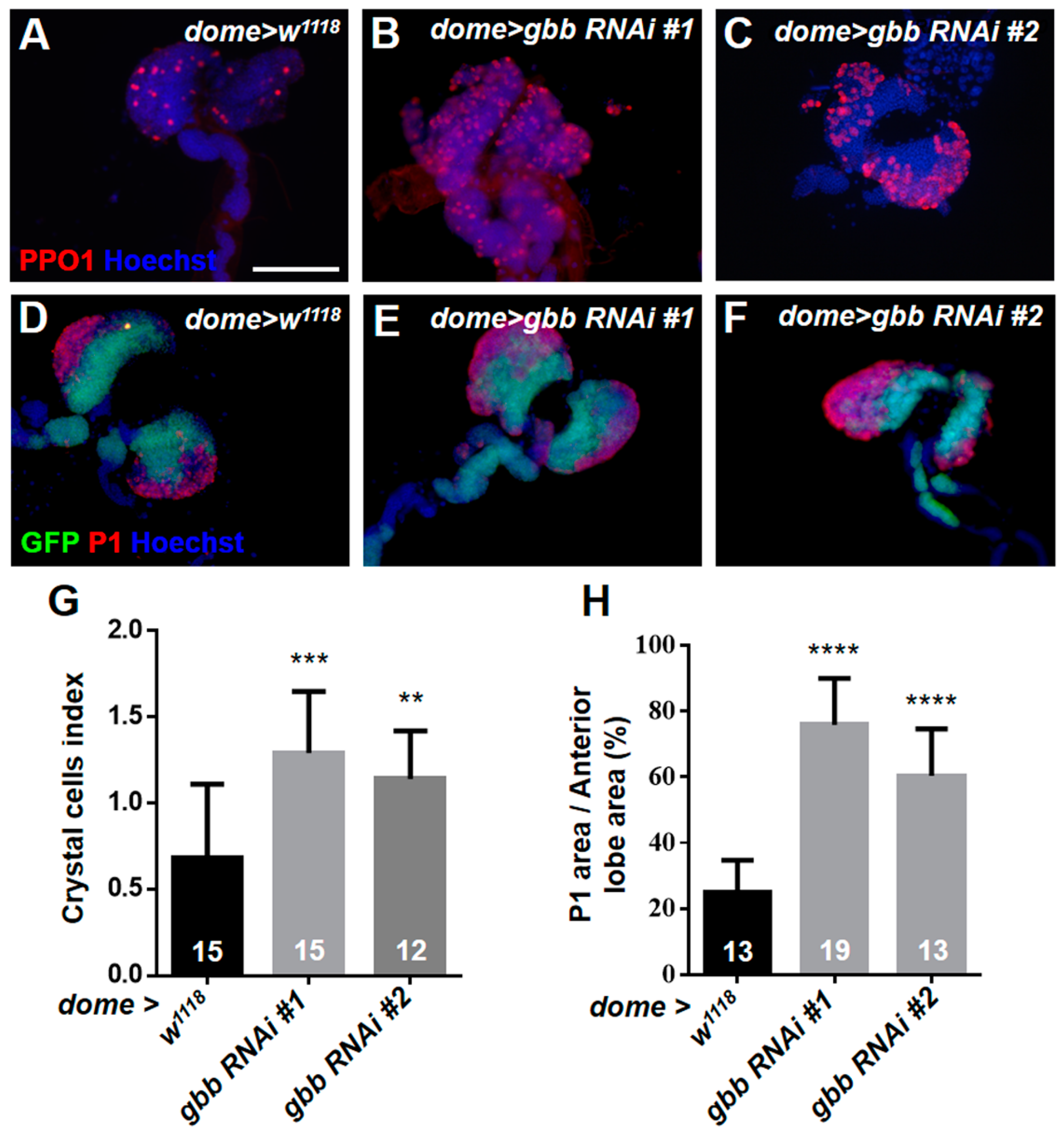
3.4. Knockdown of Gbb in Intermediate Progenitors and Progenitors Resulted in the Over-Differentiation of Crystal Cells and Plasmatocytes
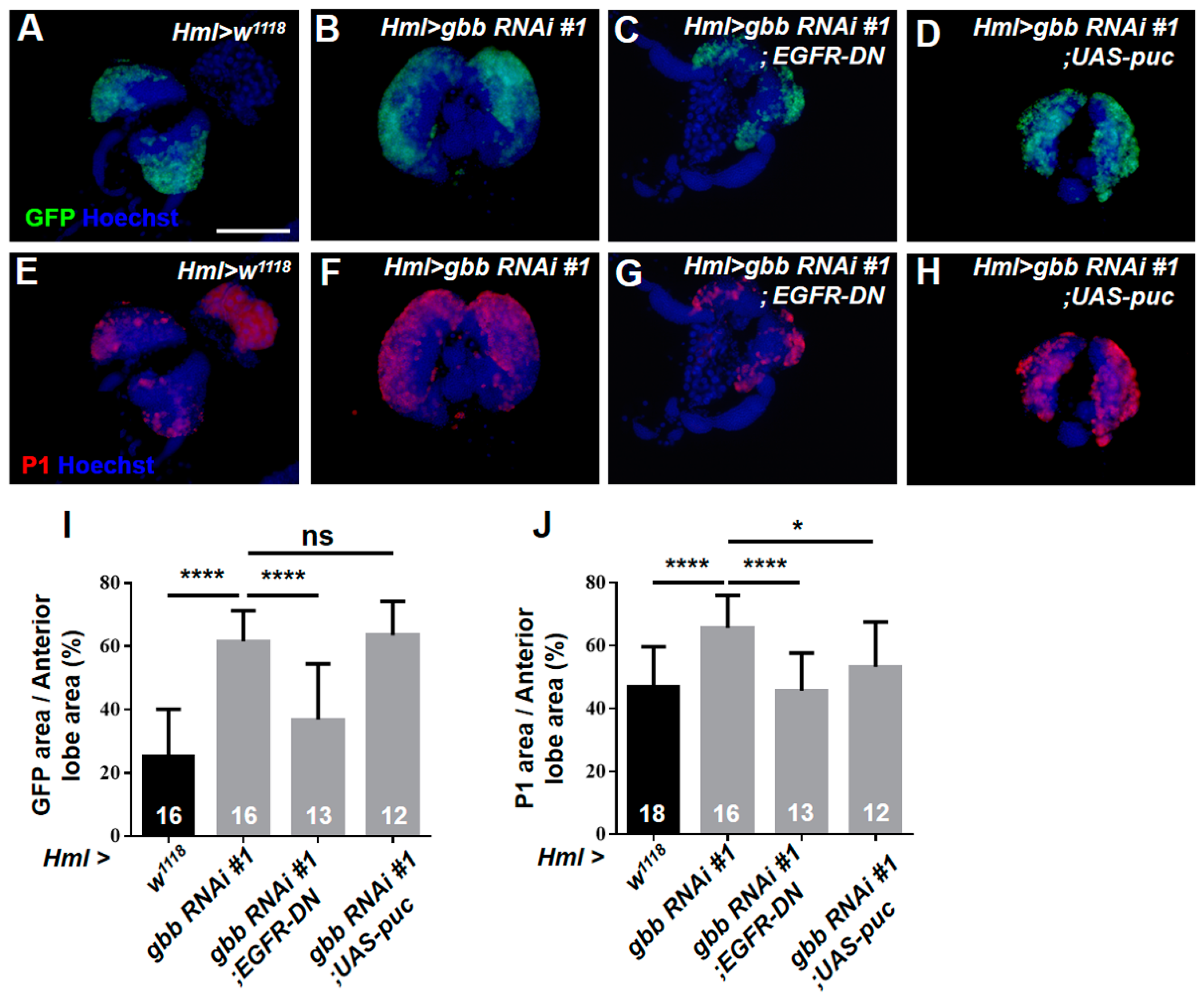
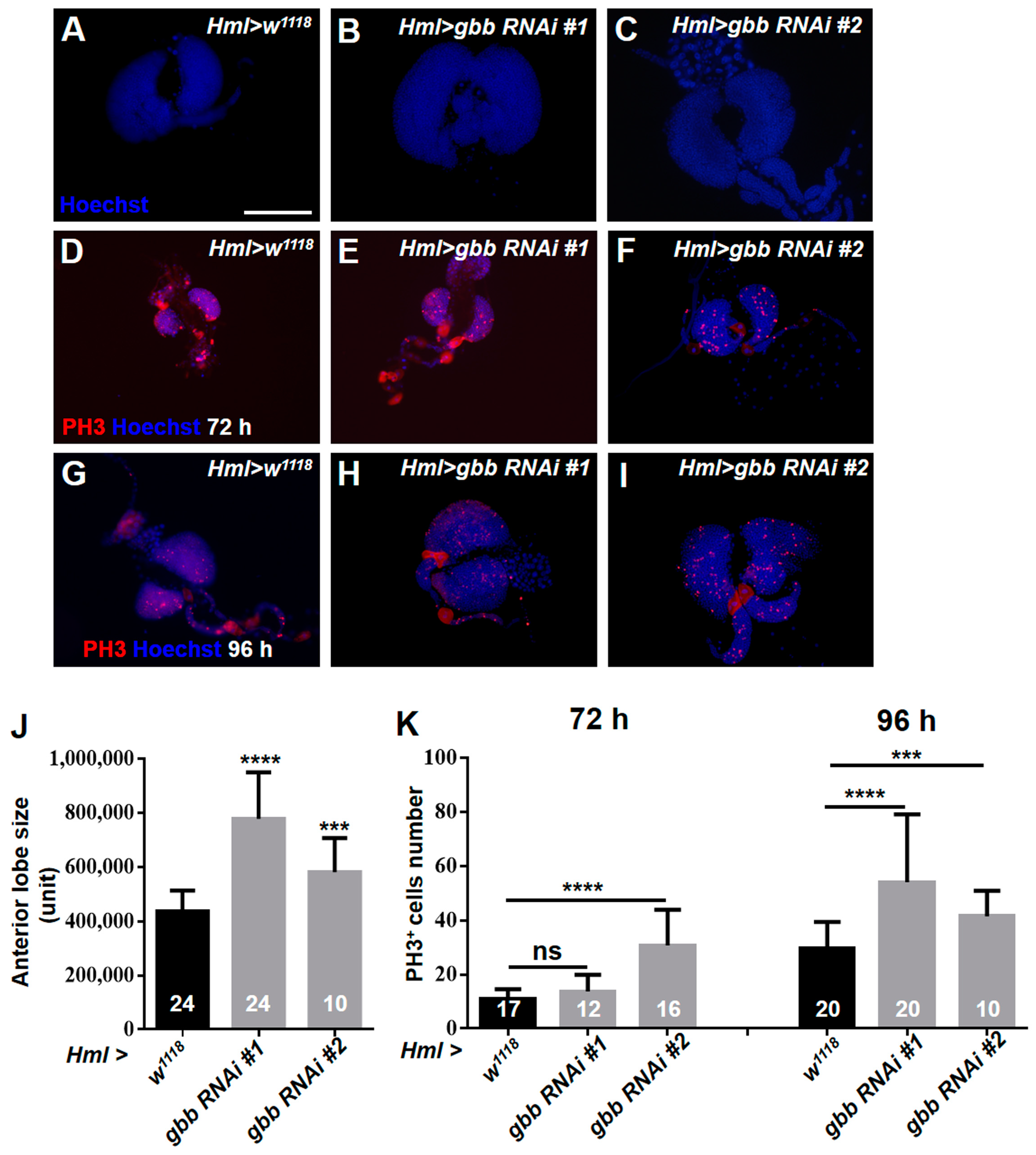
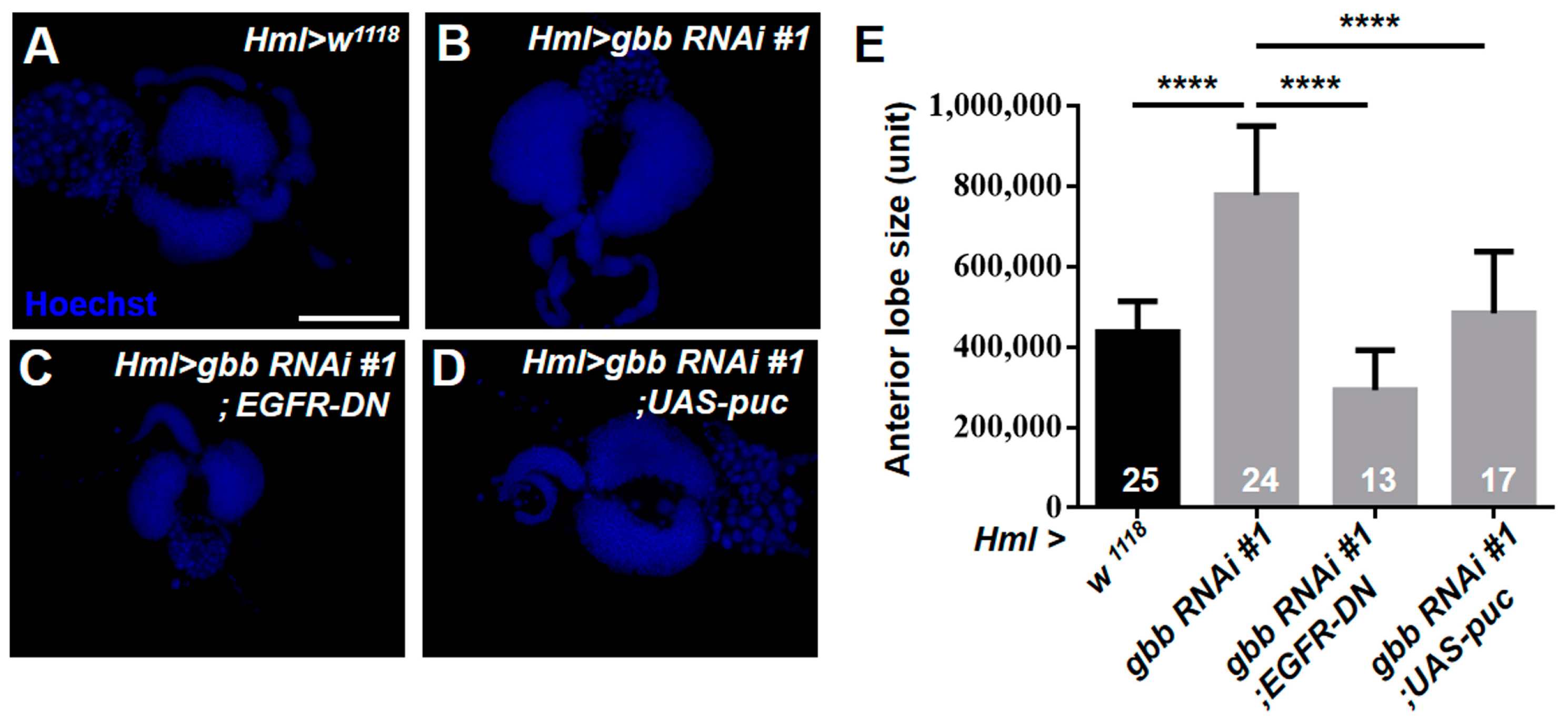
3.5. Gbb in the CZ Is Required for Preventing the Hyperproliferation of Lymph Glands by Inhibiting the Overactivation of EGFR and JNK Pathways
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boulet, M.; Miller, M.; Vandel, L.; Waltzer, L. From Drosophila Blood Cells to Human Leukemia. Adv. Exp. Med. Biol. 2018, 1076, 195–214. [Google Scholar]
- Harrison, D.A.; Binari, R.; Nahreini, T.S.; Gilman, M.; Perrimon, N. Activation of a Drosophila Janus kinase (JAK) causes hematopoietic neoplasia and developmental defects. EMBO J. 1995, 14, 2857–2865. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, R.P.; Melk, J.P.; Govind, S. Genetic analysis of contributions of dorsal group and JAK-Stat92E pathway genes to larval hemocyte concentration and the egg encapsulation response in Drosophila. Genetics 2004, 166, 1343–1356. [Google Scholar] [CrossRef] [PubMed]
- Mughal, T.I.; Radich, J.P.; Deininger, M.W.; Apperley, J.F.; Hughes, T.P.; Harrison, C.J.; Gambacorti-Passerini, C.; Saglio, G.; Cortes, J.; Daley, G.Q. Chronic myeloid leukemia: Reminiscences and dreams. Haematologica 2016, 101, 541–558. [Google Scholar] [CrossRef] [PubMed]
- Shirinian, M.; Kambris, Z.; Hamadeh, L.; Grabbe, C.; Journo, C.; Mahieux, R.; Bazarbachi, A. A Transgenic Drosophila melanogaster Model To Study Human T-Lymphotropic Virus Oncoprotein Tax-1-Driven Transformation In Vivo. J. Virol. 2015, 89, 8092–8095. [Google Scholar] [CrossRef]
- Sinenko, S.A.; Hung, T.; Moroz, T.; Tran, Q.M.; Sidhu, S.; Cheney, M.D.; Speck, N.A.; Banerjee, U. Genetic manipulation of AML1-ETO-induced expansion of hematopoietic precursors in a Drosophila model. Blood 2010, 116, 4612–4620. [Google Scholar] [CrossRef] [PubMed]
- Baril, C.; Gavory, G.; Bidla, G.; Knævelsrud, H.; Sauvageau, G.; Therrien, M. Human NUP98-HOXA9 promotes hyperplastic growth of hematopoietic tissues in Drosophila. Dev. Biol. 2017, 421, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Holz, A.; Bossinger, B.; Strasser, T.; Janning, W.; Klapper, R. The two origins of hemocytes in Drosophila. Development 2003, 130, 4955–4962. [Google Scholar] [CrossRef] [PubMed]
- Lanot, R.; Zachary, D.; Holder, F.; Meister, M. Postembryonic hematopoiesis in Drosophila. Dev. Biol. 2001, 230, 243–257. [Google Scholar] [CrossRef]
- Yu, S.; Luo, F.; Jin, L.H. The Drosophila lymph gland is an ideal model for studying hematopoiesis. Dev. Comp. Immunol. 2018, 83, 60–69. [Google Scholar] [CrossRef]
- Hao, Y.; Jin, L.H. Dual role for Jumu in the control of hematopoietic progenitors in the Drosophila lymph gland. eLife 2017, 6, e25094. [Google Scholar] [CrossRef]
- Jung, S.H.; Evans, C.J.; Uemura, C.; Banerjee, U. The Drosophila lymph gland as a developmental model of hematopoiesis. Development 2005, 132, 2521–2533. [Google Scholar] [CrossRef] [PubMed]
- Crozatier, M.; Meister, M. Drosophila haematopoiesis. Cell. Microbiol. 2007, 9, 1117–1126. [Google Scholar] [CrossRef]
- Sorrentino, R.P.; Carton, Y.; Govind, S. Cellular immune response to parasite infection in the Drosophila lymph gland is developmentally regulated. Dev. Biol. 2002, 243, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Louradour, I.; Sharma, A.; Morin-Poulard, I.; Letourneau, M.; Vincent, A.; Crozatier, M.; Vanzo, N. Reactive oxygen species-dependent Toll/NF-κB activation in the Drosophila hematopoietic niche confers resistance to wasp parasitism. eLife 2017, 6, e25496. [Google Scholar] [CrossRef] [PubMed]
- Zettervall, C.J.; Anderl, I.; Williams, M.J.; Palmer, R.; Kurucz, E.; Ando, I.; Hultmark, D. A directed screen for genes involved in Drosophila blood cell activation. Proc. Natl. Acad. Sci. USA 2004, 101, 14192–14197. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Luo, F.; Jin, L.H. Rab5 and Rab11 maintain hematopoietic homeostasis by restricting multiple signaling pathways in Drosophila. eLife 2021, 10, e60870. [Google Scholar] [CrossRef]
- Stultz, B.G.; Hursh, D.A. Gene Regulation of BMP Ligands in Drosophila. Methods Mol. Biol. 2019, 1891, 75–89. [Google Scholar]
- Parker, L.; Stathakis, D.G.; Arora, K. Regulation of BMP and activin signaling in Drosophila. Prog. Mol. Subcell. Biol. 2004, 34, 73–101. [Google Scholar]
- Twombly, V.; Blackman, R.K.; Jin, H.; Graff, J.M.; Padgett, R.W.; Gelbart, W.M. The TGF-beta signaling pathway is essential for Drosophila oogenesis. Development 1996, 122, 1555–1565. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Spradling, A.C. Decapentaplegic is essential for the maintenance and division of germline stem cells in the Drosophila ovary. Cell 1998, 94, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Staehling-Hampton, K.; Hoffmann, F.M.; Baylies, M.K.; Rushton, E.; Bate, M. dpp induces mesodermal gene expression in Drosophila. Nature 1994, 372, 783–786. [Google Scholar] [CrossRef] [PubMed]
- Frasch, M. Induction of visceral and cardiac mesoderm by ectodermal Dpp in the early Drosophila embryo. Nature 1995, 374, 464–467. [Google Scholar] [CrossRef]
- Panganiban, G.E.; Reuter, R.; Scott, M.P.; Hoffmann, F.M. A Drosophila growth factor homolog, decapentaplegic, regulates homeotic gene expression within and across germ layers during midgut morphogenesis. Development 1990, 110, 1041–1050. [Google Scholar] [CrossRef] [PubMed]
- Spencer, F.A.; Hoffmann, F.M.; Gelbart, W.M. Decapentaplegic: A gene complex affecting morphogenesis in Drosophila melanogaster. Cell 1982, 28, 451–461. [Google Scholar] [CrossRef]
- Stultz, B.G.; Lee, H.; Ramon, K.; Hursh, D.A. Decapentaplegic head capsule mutations disrupt novel peripodial expression controlling the morphogenesis of the Drosophila ventral head. Dev. Biol. 2006, 296, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Kawase, E.; Wong, M.D.; Ding, B.C.; Xie, T. Gbb/Bmp signaling is essential for maintaining germline stem cells and for repressing bam transcription in the Drosophila testis. Development 2004, 131, 1365–1375. [Google Scholar] [CrossRef]
- Pennetier, D.; Oyallon, J.; Morin-Poulard, I.; Dejean, S.; Vincent, A.; Crozatier, M. Size control of the Drosophila hematopoietic niche by bone morphogenetic protein signaling reveals parallels with mammals. Proc. Natl. Acad. Sci. USA 2012, 109, 3389–3394. [Google Scholar] [CrossRef] [PubMed]
- McCabe, B.D.; Marqués, G.; Haghighi, A.P.; Fetter, R.D.; Crotty, M.L.; Haerry, T.E.; Goodman, C.S.; O’Connor, M.B. The BMP homolog Gbb provides a retrograde signal that regulates synaptic growth at the Drosophila neuromuscular junction. Neuron 2003, 39, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Khalsa, O.; Yoon, J.W.; Torres-Schumann, S.; Wharton, K.A. TGF-beta/BMP superfamily members, Gbb-60A and Dpp, cooperate to provide pattern information and establish cell identity in the Drosophila wing. Development 1998, 125, 2723–2734. [Google Scholar] [CrossRef]
- Wharton, K.A.; Cook, J.M.; Torres-Schumann, S.; de Castro, K.; Borod, E.; Phillips, D.A. Genetic analysis of the bone morphogenetic protein-related gene, gbb, identifies multiple requirements during Drosophila development. Genetics 1999, 152, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Ballard, S.L.; Jarolimova, J.; Wharton, K.A. Gbb/BMP signaling is required to maintain energy homeostasis in Drosophila. Dev. Biol. 2010, 337, 375–385. [Google Scholar] [CrossRef]
- Tian, A.; Jiang, J. Intestinal epithelium-derived BMP controls stem cell self-renewal in Drosophila adult midgut. eLife 2014, 3, e01857. [Google Scholar] [CrossRef]
- Hao, Y.; Yu, S.; Luo, F.; Jin, L.H. Jumu is required for circulating hemocyte differentiation and phagocytosis in Drosophila. Cell Commun. Signal. 2018, 16, 95. [Google Scholar] [CrossRef]
- Cho, B.; Yoon, S.H.; Lee, D.; Koranteng, F.; Tattikota, S.G.; Cha, N.; Shin, M.; Do, H.; Hu, Y.; Oh, S.Y.; et al. Single-cell transcriptome maps of myeloid blood cell lineages in Drosophila. Nat. Commun. 2020, 11, 4483. [Google Scholar] [CrossRef]
- Sinenko, S.A.; Mandal, L.; Martinez-Agosto, J.A.; Banerjee, U. Dual role of wingless signaling in stem-like hematopoietic precursor maintenance in Drosophila. Dev. Cell 2009, 16, 756–763. [Google Scholar] [CrossRef] [PubMed]
- Krzemien, J.; Oyallon, J.; Crozatier, M.; Vincent, A. Hematopoietic progenitors and hemocyte lineages in the Drosophila lymph gland. Dev. Biol. 2010, 346, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Pinal, N.; Martín, M.; Medina, I.; Morata, G. Short-term activation of the Jun N-terminal kinase pathway in apoptosis-deficient cells of Drosophila induces tumorigenesis. Nat. Commun. 2018, 9, 1541. [Google Scholar] [CrossRef]
- Zhang, J.; Niu, C.; Ye, L.; Huang, H.; He, X.; Tong, W.G.; Ross, J.; Haug, J.; Johnson, T.; Feng, J.Q.; et al. Identification of the haematopoietic stem cell niche and control of the niche size. Nature 2003, 425, 836–841. [Google Scholar] [CrossRef]
- Leatherman, J.L.; Dinardo, S. Germline self-renewal requires cyst stem cells and stat regulates niche adhesion in Drosophila testes. Nat. Cell Biol. 2010, 12, 806–811. [Google Scholar] [CrossRef]
- Song, X.; Wong, M.D.; Kawase, E.; Xi, R.; Ding, B.C.; McCarthy, J.J.; Xie, T. Bmp signals from niche cells directly repress transcription of a differentiation-promoting gene, bag of marbles, in germline stem cells in the Drosophila ovary. Development 2004, 131, 1353–1364. [Google Scholar] [CrossRef]
- Harris, R.E.; Pargett, M.; Sutcliffe, C.; Umulis, D.; Ashe, H.L. Brat promotes stem cell differentiation via control of a bistable switch that restricts BMP signaling. Dev. Cell 2011, 20, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.I.; Woodcock, K.J.; Geissmann, F.; Trouillet, C.; Dionne, M.S. Multiple TGF-β superfamily signals modulate the adult Drosophila immune response. Curr. Biol. 2011, 21, 1672–1677. [Google Scholar] [CrossRef] [PubMed]
- Patrnogic, J.; Heryanto, C.; Eleftherianos, I. Wounding-induced upregulation of the Bone Morphogenic Protein signaling pathway in Drosophila promotes survival against parasitic nematode infection. Gene 2018, 673, 112–118. [Google Scholar] [CrossRef]
- Patrnogic, J.; Heryanto, C.; Eleftherianos, I. Transcriptional up-regulation of the TGF-β intracellular signaling transducer Mad of Drosophila larvae in response to parasitic nematode infection. Innate Immun. 2018, 24, 349–356. [Google Scholar] [CrossRef]
- Patrnogic, J.; Heryanto, C.; Ozakman, Y.; Eleftherianos, I. Transcript analysis reveals the involvement of NF-κB transcription factors for the activation of TGF-β signaling in nematode-infected Drosophila. Immunogenetics 2019, 71, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Gjelsvik, K.J.; Follansbee, T.L.; Ganter, G.K. Bone Morphogenetic Protein Glass Bottom Boat (BMP5/6/7/8) and its receptor Wishful Thinking (BMPRII) are required for injury-induced allodynia in Drosophila. Mol. Pain 2018, 14, 1744806918802703. [Google Scholar] [CrossRef] [PubMed]
- Tian, A.; Wang, B.; Jiang, J. Injury-stimulated and self-restrained BMP signaling dynamically regulates stem cell pool size during Drosophila midgut regeneration. Proc. Natl. Acad. Sci. USA 2017, 114, E2699–E2708. [Google Scholar] [CrossRef]
- Wan, Y.Y.; Flavell, R.A. TGF-beta and regulatory T cell in immunity and autoimmunity. J. Clin. Immunol. 2008, 28, 647–659. [Google Scholar] [CrossRef]
- Johnston, C.J.; Smyth, D.J.; Dresser, D.W.; Maizels, R.M. TGF-β in tolerance, development and regulation of immunity. Cell Immunol. 2016, 299, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Goldman, D.C.; Bailey, A.S.; Pfaffle, D.L.; Al Masri, A.; Christian, J.L.; Fleming, W.H. BMP4 regulates the hematopoietic stem cell niche. Blood 2009, 114, 4393–4401. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Florescu, S.; Boettcher, A.L.; Luo, L.; Dutta, D.; Kerr, G.; Cai, Y.; Edgar, B.A.; Boutros, M. Dpp/Gbb signaling is required for normal intestinal regeneration during infection. Dev. Biol. 2015, 15, 189–203. [Google Scholar] [CrossRef]
- Khadilkar, R.J.; Ho, K.Y.L.; Venkatesh, B.; Tanentzapf, G. Integrins Modulate Extracellular Matrix Organization to Control Cell Signaling during Hematopoiesis. Curr. Biol. 2020, 30, 3316–3329. [Google Scholar] [CrossRef]
- Yang, S.; Wu, X.; Daoutidou, E.I.; Zhang, Y.; Shimell, M.; Chuang, K.H.; Peterson, A.J.; O’Connor, M.B.; Zheng, X. The NDNF-like factor Nord is a Hedgehog-induced extracellular BMP modulator that regulates Drosophila wing patterning and growth. eLife 2022, 11, e73357. [Google Scholar] [CrossRef]
- Tokusumi, T.; Sorrentino, R.P.; Russell, M.; Ferrarese, R.; Govind, S.; Schulz, R.A. Characterization of a lamellocyte transcriptional enhancer located within the misshapen gene of Drosophila melanogaster. PLoS ONE 2009, 4, e6429. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Jin, Y.; Marchetti, M.; Lewis, M.R.; Hammouda, O.T.; Edgar, B.A. EGFR signaling activates intestinal stem cells by promoting mitochondrial biogenesis and β-oxidation. Curr. Biol. 2022, 32, 3704–3719. [Google Scholar] [CrossRef] [PubMed]
- Richardson, H.E.; Portela, M. Modelling Cooperative Tumorigenesis in Drosophila. BioMed Res. Int. 2018, 2018, 4258387. [Google Scholar] [CrossRef] [PubMed]
- La Marca, J.E.; Richardson, H.E. Two-Faced: Roles of JNK Signalling During Tumourigenesis in the Drosophila Model. Front. Cell Dev. Biol. 2020, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Pinal, N.; Calleja, M.; Morata, G. Pro-apoptotic and pro-proliferation functions of the JNK pathway of Drosophila: Roles in cell competition, tumorigenesis and regeneration. Open Biol. 2019, 9, 180256. [Google Scholar] [CrossRef]
- Zhou, J.; Boutros, M. JNK-dependent intestinal barrier failure disrupts host-microbe homeostasis during tumorigenesis. Proc. Natl. Acad. Sci. USA 2020, 117, 9401–9412. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Wang, D.; Si, J.; Jin, L.H.; Hao, Y. Gbb Regulates Blood Cell Proliferation and Differentiation through JNK and EGFR Signaling Pathways in the Drosophila Lymph Gland. Cells 2023, 12, 661. https://doi.org/10.3390/cells12040661
Zhang W, Wang D, Si J, Jin LH, Hao Y. Gbb Regulates Blood Cell Proliferation and Differentiation through JNK and EGFR Signaling Pathways in the Drosophila Lymph Gland. Cells. 2023; 12(4):661. https://doi.org/10.3390/cells12040661
Chicago/Turabian StyleZhang, Wenhao, Dongmei Wang, Jingjing Si, Li Hua Jin, and Yangguang Hao. 2023. "Gbb Regulates Blood Cell Proliferation and Differentiation through JNK and EGFR Signaling Pathways in the Drosophila Lymph Gland" Cells 12, no. 4: 661. https://doi.org/10.3390/cells12040661
APA StyleZhang, W., Wang, D., Si, J., Jin, L. H., & Hao, Y. (2023). Gbb Regulates Blood Cell Proliferation and Differentiation through JNK and EGFR Signaling Pathways in the Drosophila Lymph Gland. Cells, 12(4), 661. https://doi.org/10.3390/cells12040661