
Targeting Neuroinflammation with Abscisic Acid Reduces Pain Sensitivity in Females and Hyperactivity in Males of an ADHD Mice Model


 ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
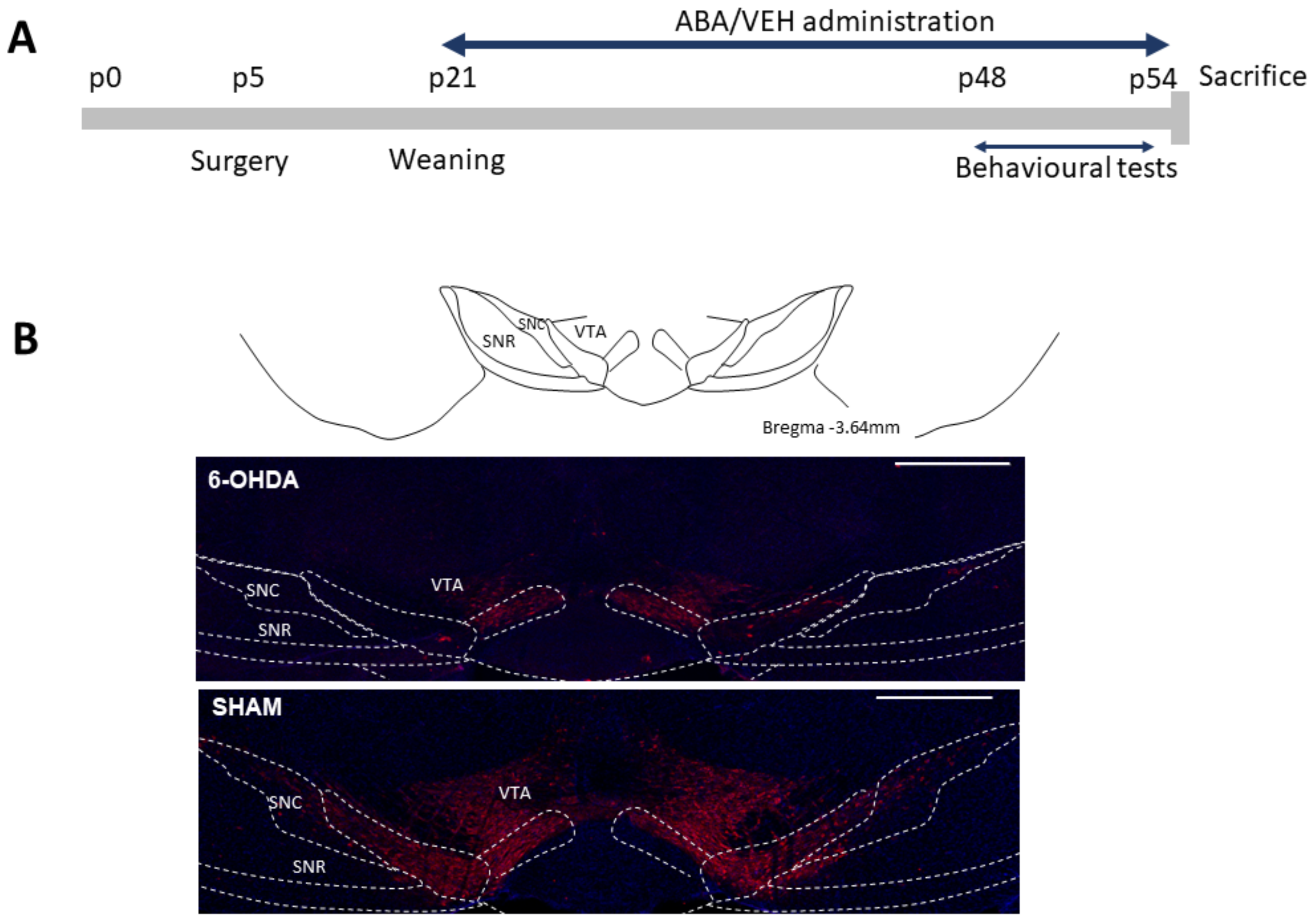
2.1. Animals and Surgical Procedures
2.2. Behavioral Procedures
2.3. Immunofluorescence Procedure
2.4. Statistical Analysis
3. Results
3.1. 6-OHDA Reduced TH Staining in the Ventral Tegmental Area
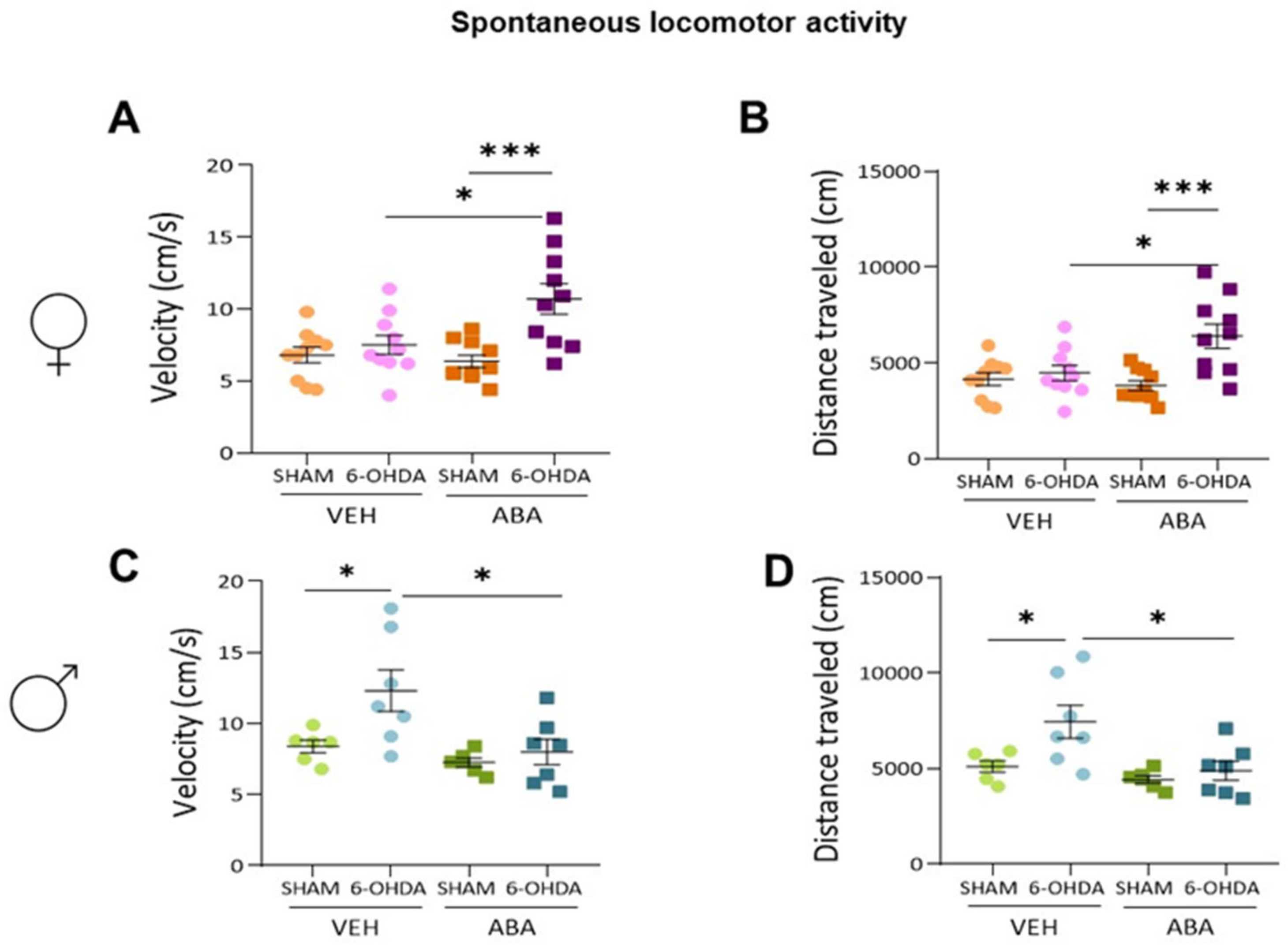
3.2. Effect of Perinatal i.c.v. 6-OHDA Injection and ABA Treatment on Spontaneous Locomotor Activity
3.3. 6-OHDA Injection Reduces Novel Object Recognition, and ABA Treatment Does Not Rescue the Lesion Effect
3.4. The 6-OHDA Lesion Did Not Alter Spatial Memory as Measured by Spontaneous Alternation (T-Maze) Nor Anxiety as Measured by Elevated plus Maze
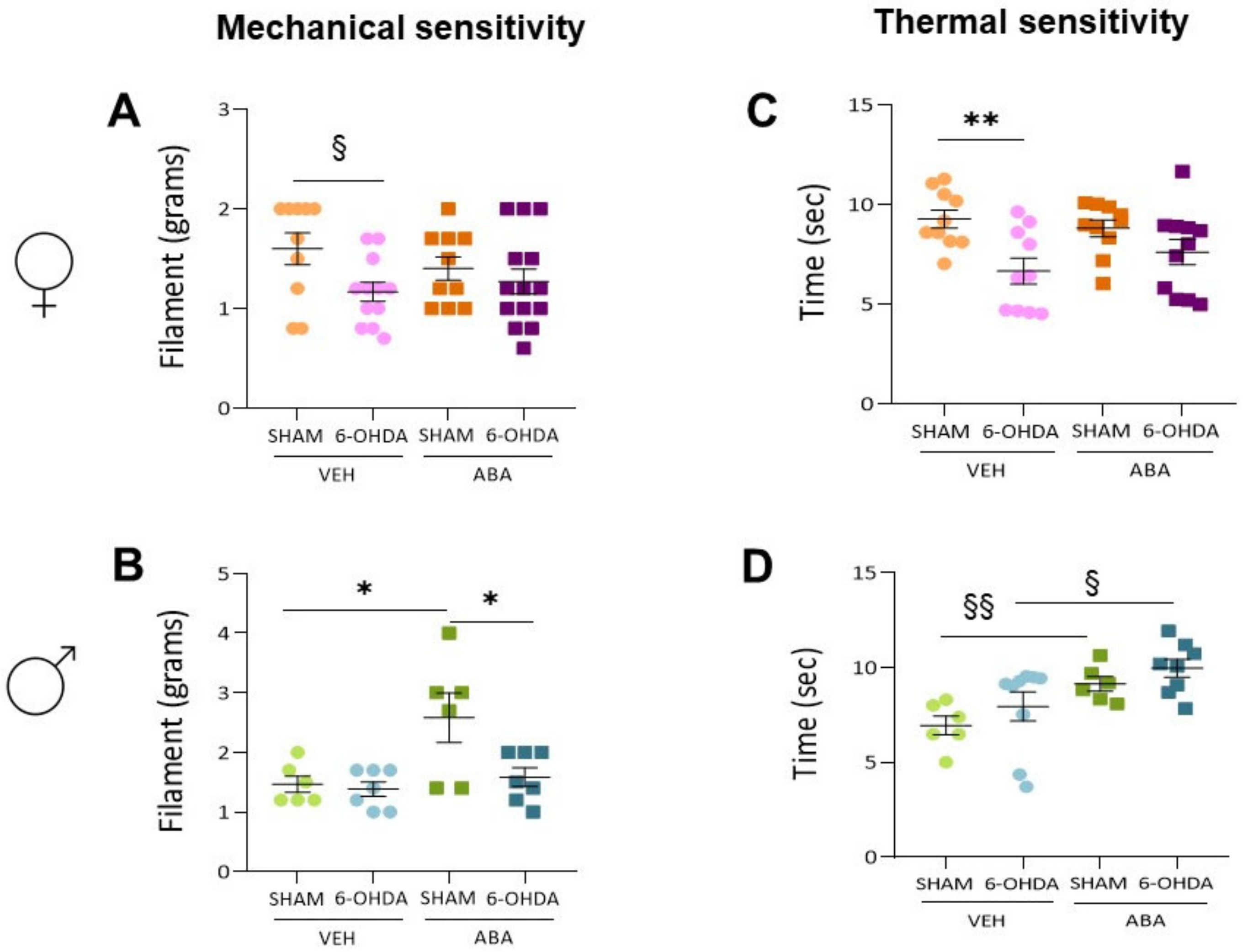
3.5. The 6-OHDA Lesion Had a Sex-Dependent Effect on Pain Sensitivity Tests—ABA Treatment Had a Potential Beneficial Effect Elevating Pain Sensitivity Threshold
3.6. Perinatal Injection of 6-OHDA Lesions Had a Sex-Dependent Effect on Sociability in Two-Month-Old Mice
3.7. Perinatal 6-OHDA Lesion Had a Sex-Dependent Effect on Pain Sensitivity by the Mechanical Stimulus, as Measured by the Von Frey Test
3.8. Perinatal 6-OHDA Lesions Had a Sex-Dependent Effect on Pain Sensitivity by Thermal Stimulus, and ABA Treatment Increased the Threshold in Both Females and Males
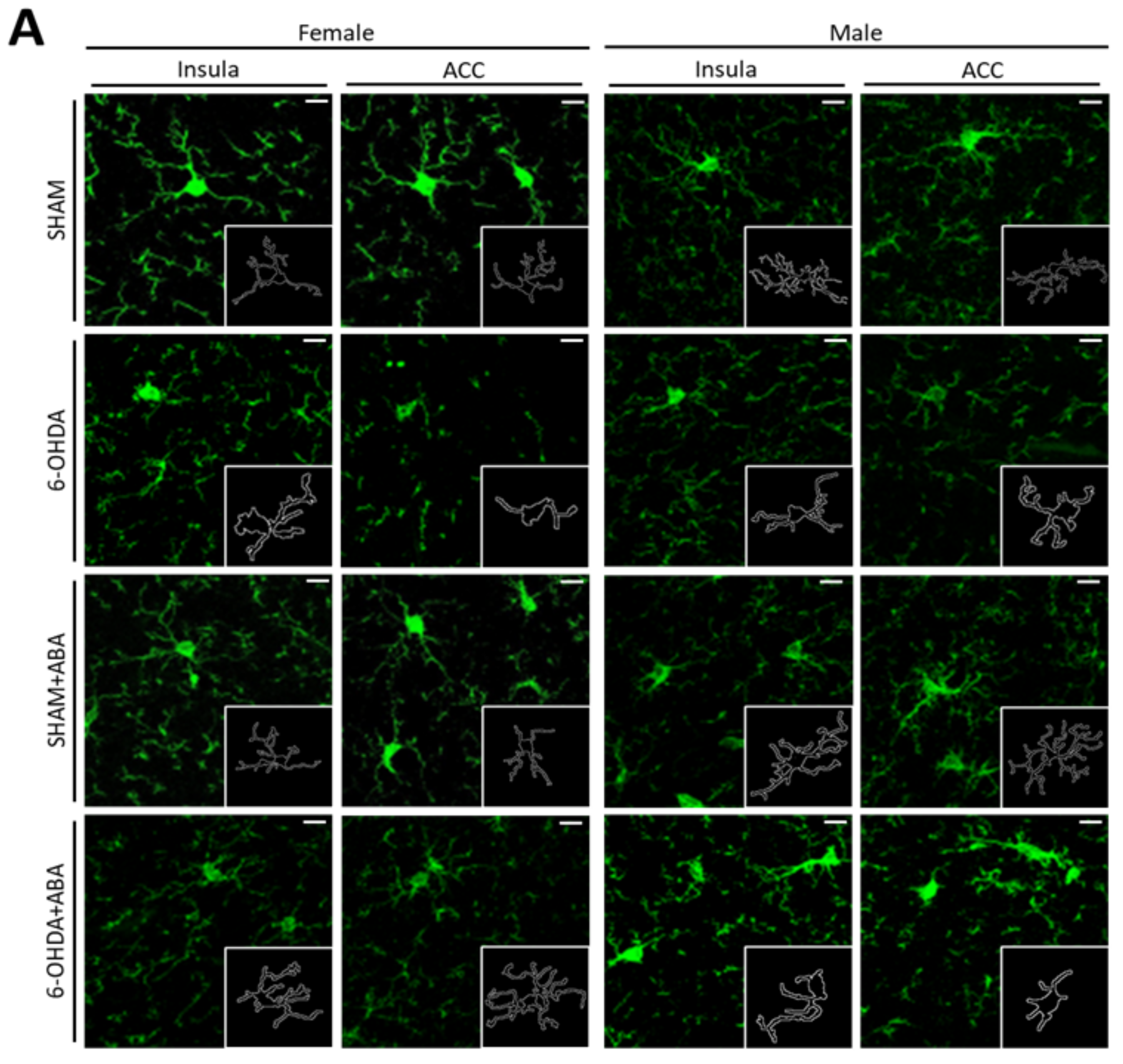
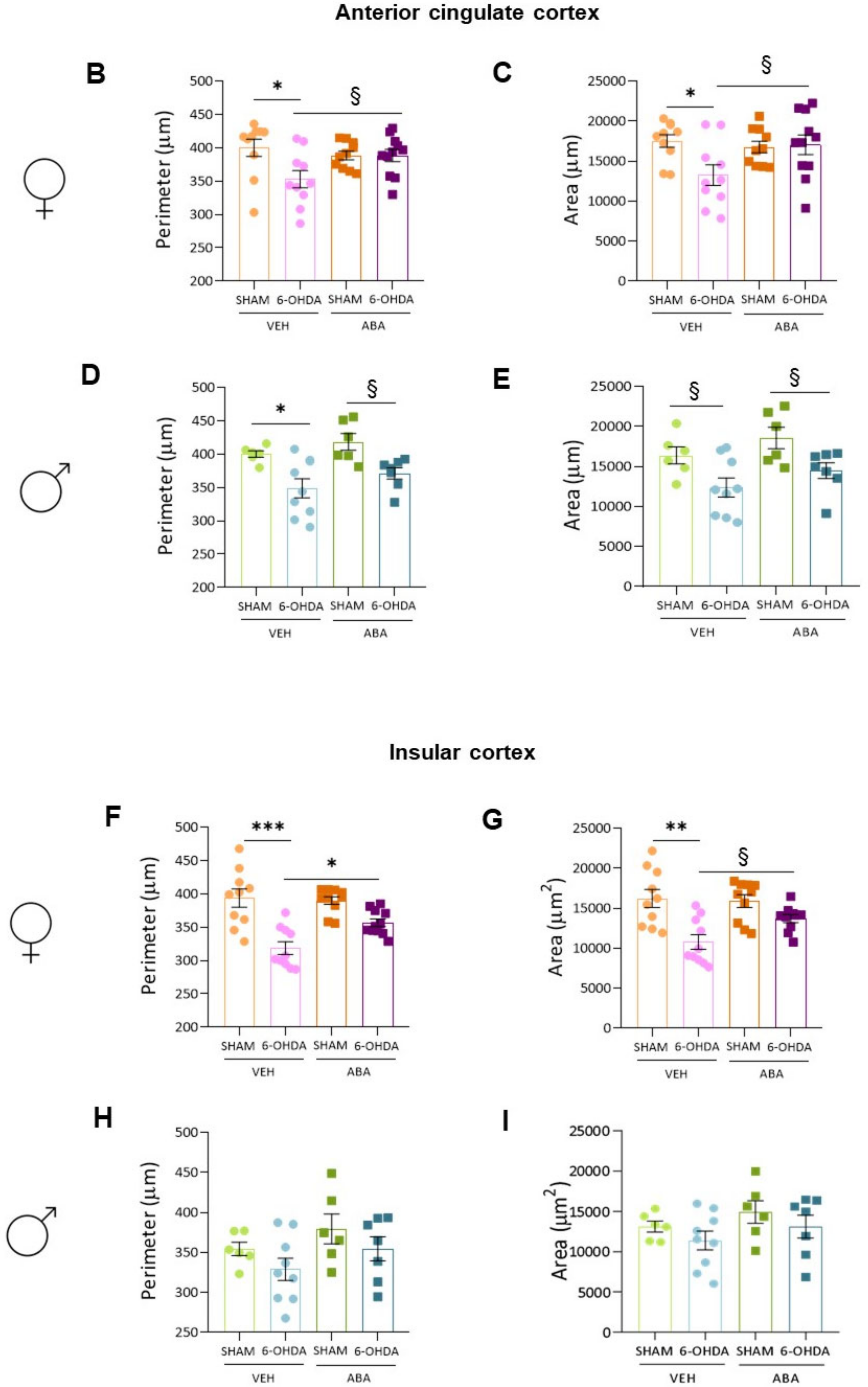
3.9. In the Anterior Cingulate Cortex (ACC), the 6-OHDA Lesion Induced Microglia Polarization to a Proinflammatory Status, and ABA Rescued the Effect in Females but Not in Males
3.10. In the Posterior Insular Cortex, 6-OHDA Lesions Induced Microglia Polarization to a Proinflammatory Status Only in Females, and ABA Rescued the Effect
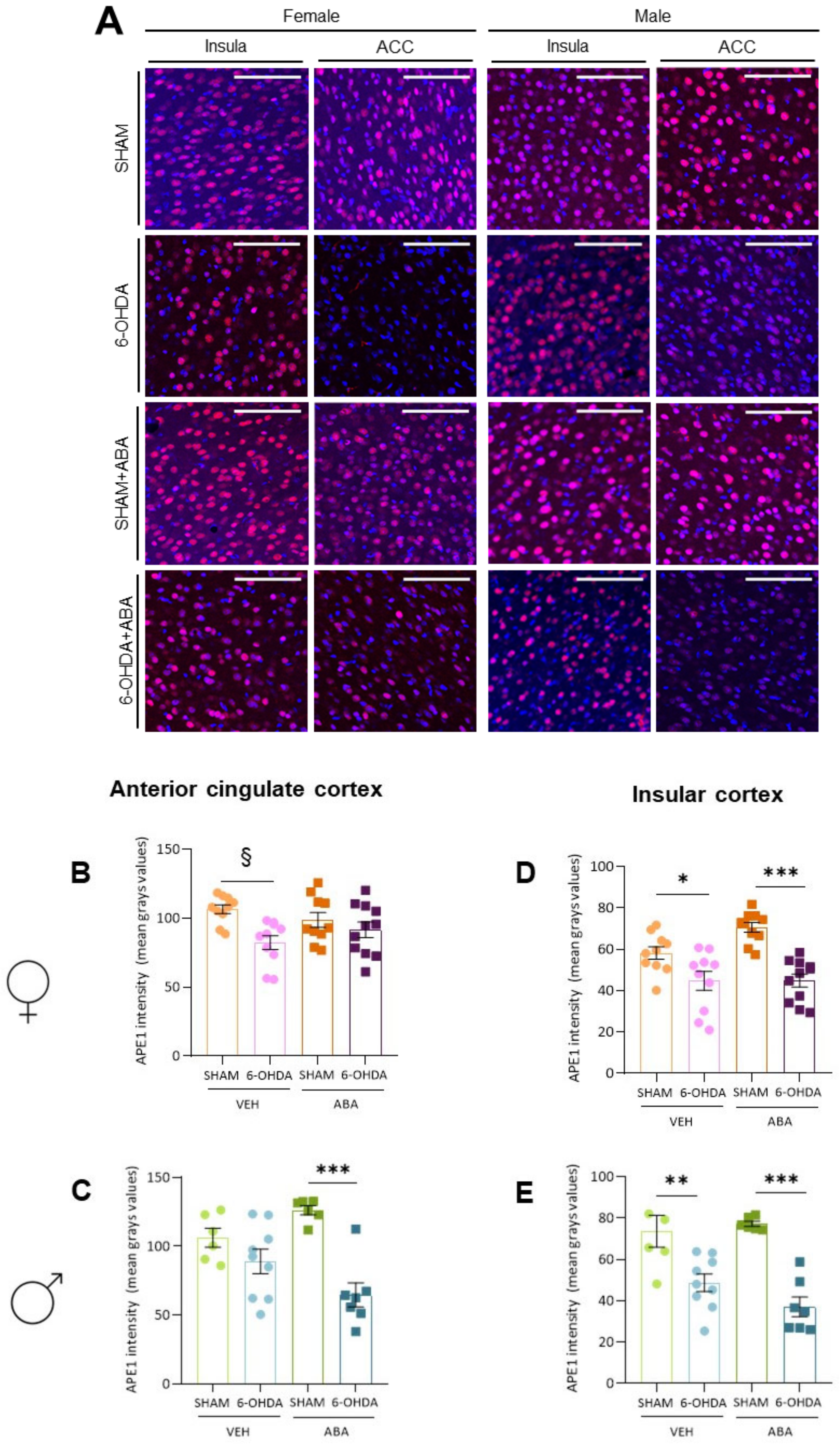
3.11. Perinatal 6-OHDA Lesion and ABA Treatment Affected APE1 Intensity Differently in the ACC of Females and Males
3.12. Perinatal 6-OHDA Lesion Reduces Ape1 Intensity in Both Females’ and Males’ Posterior Insula
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Faraone, S.V.; Asherson, P.; Banaschewski, T.; Biederman, J.; Buitelaar, J.K.; Ramos-Quiroga, J.A.; Rohde, L.A.; Sonuga-Barke, E.J.S.; Tannock, R.; Franke, B. Attention-Deficit/Hyperactivity Disorder. Nat. Rev. Dis. Prim. 2015, 1, 15020. [Google Scholar] [CrossRef] [PubMed]
- Avni, E.; Ben-Itzchak, E.; Zachor, D.A. The Presence of Comorbid ADHD and Anxiety Symptoms in Autism Spectrum Disorder: Clinical Presentation and Predictors. Front. Psychiatry 2018, 9, 717. [Google Scholar] [CrossRef] [PubMed]
- Borschuk, A.P.; Rodweller, C.; Salorio, C.F. The Influence of Comorbid Asthma on the Severity of Symptoms in Children with Attention-Deficit Hyperactivity Disorder. J. Asthma 2018, 55, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Slim, M.; Rico-Villademoros, F.; Calandre, E.P. Psychiatric Comorbidity in Children and Adults with Gluten-Related Disorders: A Narrative Review. Nutrients 2018, 10, 875. [Google Scholar] [CrossRef]
- Salem, H.; Vivas, D.; Cao, F.; Kazimi, I.F.; Teixeira, A.L.; Zeni, C.P. ADHD Is Associated with Migraine: A Systematic Review and Meta-Analysis. Eur. Child. Adolesc Psychiatry 2018, 27, 267–277. [Google Scholar] [CrossRef]
- Brus, M.J.; Solanto, M.V.; Goldberg, J.F. Adult ADHD vs. Bipolar Disorder in the DSM-5 Era: A Challenging Differentiation for Clinicians. J. Psychiatr. Pr. 2014, 20, 428–437. [Google Scholar] [CrossRef]
- Velő, S.; Keresztény, Á.; Ferenczi-Dallos, G.; Pump, L.; Móra, K.; Balázs, J. The Association between Prosocial Behaviour and Peer Relationships with Comorbid Externalizing Disorders and Quality of Life in Treatment-Naïve Children and Adolescents with Attention Deficit Hyperactivity Disorder. Brain Sci. 2021, 11, 475. [Google Scholar] [CrossRef]
- Pawaskar, M.; Fridman, M.; Grebla, R.; Madhoo, M. Comparison of Quality of Life, Productivity, Functioning and Self-Esteem in Adults Diagnosed with ADHD and with Symptomatic ADHD. J. Atten. Disord. 2020, 24, 136–144. [Google Scholar] [CrossRef]
- Agarwal, R.; Goldenberg, M.; Perry, R.; William IsHak, W. The Quality of Life of Adults with Attention Deficit Hyperactivity Disorder: A Systematic Review. Innov. Clin. Neurosci. 2012, 9, 10–21. [Google Scholar]
- Luo, Y.; Weibman, D.; Halperin, J.M.; Li, X. A Review of Heterogeneity in Attention Deficit/Hyperactivity Disorder (ADHD). Front. Hum. Neurosci. 2019, 13, 42. [Google Scholar] [CrossRef]
- Rucklidge, J.J. Gender Differences in Attention-Deficit/Hyperactivity Disorder. Psychiatr. Clin. N. Am. 2010, 33, 357–373. [Google Scholar] [CrossRef]
- Loyer Carbonneau, M.; Demers, M.; Bigras, M.; Guay, M.C. Meta-Analysis of Sex Differences in ADHD Symptoms and Associated Cognitive Deficits. J. Atten. Disord. 2021, 25, 1640–1656. [Google Scholar] [CrossRef]
- Sudre, G.; Frederick, J.; Sharp, W.; Ishii-Takahashi, A.; Mangalmurti, A.; Choudhury, S.; Shaw, P. Mapping Associations between Polygenic Risks for Childhood Neuropsychiatric Disorders, Symptoms of Attention Deficit Hyperactivity Disorder, Cognition, and the Brain. Mol. Psychiatry 2020, 25, 2482–2492. [Google Scholar] [CrossRef]
- Nikolas, M.A.; Burt, S.A. Genetic and Environmental Influences on ADHD Symptom Dimensions of Inattention and Hyperactivity: A Meta-Analysis. J. Abnorm. Psychol. 2010, 119, 1–17. [Google Scholar] [CrossRef]
- Anand, D.; Colpo, G.D.; Zeni, G.; Zeni, C.P.; Teixeira, A.L. Attention-Deficit/Hyperactivity Disorder and Inflammation: What Does Current Knowledge Tell Us? A Systematic Review. Front. Psychiatry 2017, 8, 228. [Google Scholar] [CrossRef]
- Leffa, D.; Torres, I.; Rohde, L. A Review on the Role of Inflammation in Attention-Deficit/Hyperactivity Disorder. Neuroimmunomodulation 2018, 25, 328–333. [Google Scholar] [CrossRef]
- Gkougka, D.; Mitropoulos, K.; Tzanakaki, G.; Panagouli, E.; Psaltopoulou, T.; Thomaidis, L.; Tsolia, M.; Sergentanis, T.N.; Tsitsika, A. Gut Microbiome and Attention Deficit/Hyperactivity Disorder: A Systematic Review. Pediatr. Res. 2022, 92, 1507–1519. [Google Scholar] [CrossRef]
- Ellul, P.; Acquaviva, E.; Peyre, H.; Rosenzwajg, M.; Gressens, P.; Klatzmann, D.; Delorme, R. Parental Autoimmune and Autoinflammatory Disorders as Multiple Risk Factors for Common Neurodevelopmental Disorders in Offspring: A Systematic Review and Meta-Analysis. Transl. Psychiatry 2022, 12, 112. [Google Scholar] [CrossRef]
- Saccaro, L.F.; Schilliger, Z.; Perroud, N.; Piguet, C. Inflammation, Anxiety, and Stress in Attention-Deficit/Hyperactivity Disorder. Biomedicines 2021, 9, 1313. [Google Scholar] [CrossRef]
- Kerekes, N.; Sanchéz-Pérez, A.M.; Landry, M. Neuroinflammation as a Possible Link between Attention-Deficit/Hyperactivity Disorder (ADHD) and Pain. Med. Hypotheses 2021, 157, 110717. [Google Scholar] [CrossRef]
- Chen, X.; Yao, T.; Cai, J.; Fu, X.; Li, H.; Wu, J. Systemic Inflammatory Regulators and 7 Major Psychiatric Disorders: A Two-Sample Mendelian Randomization Study. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2022, 116, 110534. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.E.; Teixeira, A.L. Inflammation in Psychiatric Disorders: What Comes First? Ann. N. Y. Acad. Sci. 2019, 1437, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.W.; Yoon, H.K.; Kim, Y.K. Role of Inflammation in Psychiatric Disorders. Adv. Exp. Med. Biol. 2019, 1192, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Dunn, G.A.; Nigg, J.T.; Sullivan, E.L. Neuroinflammation as a Risk Factor for Attention Deficit Hyperactivity Disorder. Pharmacol. Biochem. Behav. 2019, 182, 22–34. [Google Scholar] [CrossRef]
- Miyazaki, C.; Koyama, M.; Ota, E.; Swa, T.; Mlunde, L.B.; Amiya, R.M.; Tachibana, Y.; Yamamoto-Hanada, K.; Mori, R. Allergic Diseases in Children with Attention Deficit Hyperactivity Disorder: A Systematic Review and Meta-Analysis. BMC Psychiatry 2017, 17, 120. [Google Scholar] [CrossRef]
- Schans, J.v.d.; Çiçek, R.; de Vries, T.W.; Hak, E.; Hoekstra, P.J. Association of Atopic Diseases and Attention-Deficit/Hyperactivity Disorder: A Systematic Review and Meta-Analyses. Neurosci. Biobehav. Rev. 2017, 74, 139–148. [Google Scholar] [CrossRef]
- Bordeleau, M.; Fernández de Cossío, L.; Chakravarty, M.M.; Tremblay, M.È. From Maternal Diet to Neurodevelopmental Disorders: A Story of Neuroinflammation. Front. Cell. Neurosci. 2021, 14, 612705. [Google Scholar] [CrossRef]
- Han, V.X.; Patel, S.; Jones, H.F.; Dale, R.C. Maternal Immune Activation and Neuroinflammation in Human Neurodevelopmental Disorders. Nat. Rev. Neurol. 2021, 17, 564–579. [Google Scholar] [CrossRef]
- Thürmann, L.; Herberth, G.; Rolle-Kampczyk, U.; Röder, S.; Borte, M.; von Bergen, M.; Lehmann, I.; Trump, S. Elevated Gestational IL-13 During Fetal Development Is Associated with Hyperactivity and Inattention in Eight-Year-Old Children. Front. Immunol. 2019, 10, 1658. [Google Scholar] [CrossRef]
- Brewer, B.W.; Karoly, P. Effects of Attentional Focusing on Pain Perception. Motiv. Emot. 1989, 13, 193–203. [Google Scholar] [CrossRef]
- Longe, S.E.; Wise, R.; Bantick, S.; Lloyd, D.; Johansen-Berg, H.; McGlone, F.; Tracey, I. Counter-Stimulatory Effects on Pain Perception and Processing Are Significantly Altered by Attention: An FMRI Study. Neuroreport 2001, 12, 2021–2025. [Google Scholar] [CrossRef]
- Chan, S.C.C.; Chan, C.C.H.; Kwan, A.S.K.; Ting, K.H.; Chui, T. yi Orienting Attention Modulates Pain Perception: An ERP Study. PLoS ONE 2012, 7, e40215. [Google Scholar] [CrossRef]
- Asztély, K.; Kopp, S.; Gillberg, C.; Waern, M.; Bergman, S. Chronic Pain and Health-Related Quality of Life in Women with Autism And/or ADHD: A Prospective Longitudinal Study. J. Pain Res. 2019, 12, 2925–2932. [Google Scholar] [CrossRef]
- Kasahara, S.; Niwa, S.I.; Matsudaira, K.; Sato, N.; Oka, H.; Yamada, Y. Attention-Deficit/Hyperactivity Disorder and Chronic Pain. Psychosom. Med. 2020, 82, 346–347. [Google Scholar] [CrossRef]
- Stray, L.L.; Kristensen, Ø.; Lomeland, M.; Skorstad, M.; Stray, T.; Tønnessen, F.E. Motor Regulation Problems and Pain in Adults Diagnosed with ADHD. Behav. Brain Funct. 2013, 9, 18. [Google Scholar] [CrossRef]
- Treister, R.; Eisenberg, E.; Demeter, N.; Pud, D. Alterations in Pain Response Are Partially Reversed by Methylphenidate (Ritalin) in Adults with Attention Deficit Hyperactivity Disorder (ADHD). Pain Pr. 2015, 15, 4–11. [Google Scholar] [CrossRef]
- Kasanetz, F.; Acuña, M.A.; Nevian, T. Anterior Cingulate Cortex, Pain Perception, and Pathological Neuronal Plasticity during Chronic Pain. Book Neurobiol. Physiol. Psychol. Pain 2022, 193–202. [Google Scholar] [CrossRef]
- Newman, L.A.; McGaughy, J. Attentional Effects of Lesions to the Anterior Cingulate Cortex: How Prior Reinforcement Influences Distractibility. Behav. Neurosci. 2011, 125, 360–371. [Google Scholar] [CrossRef]
- Bai, Y.; Ma, L.T.; Chen, Y.B.; Ren, D.; Chen, Y.B.; Li, Y.Q.; Sun, H.K.; Qiu, X.T.; Zhang, T.; Zhang, M.M.; et al. Anterior Insular Cortex Mediates Hyperalgesia Induced by Chronic Pancreatitis in Rats. Mol. Brain 2019, 12, 76. [Google Scholar] [CrossRef]
- Isbrandt, D. A Mechanistic Link between Glia and Neuronal Excitability in Acute Neuroinflammation. J. Physiol. 2017, 595, 603–604. [Google Scholar] [CrossRef]
- Villemure, C.; Bushnell, M.C. Cognitive Modulation of Pain: How Do Attention and Emotion Influence Pain Processing? Pain 2002, 95, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.F.; Guan, S.Y.; Luo, L.; Li, Y.J.; Yang, L.; Zhou, X.X.; Guo, G.D.; Zhao, M.G.; Yang, Q.; Liu, G. Tetrahydroxystilbene Glucoside Relieves the Chronic Inflammatory Pain by Inhibiting Neuronal Apoptosis, Microglia Activation, and GluN2B Overexpression in Anterior Cingulate Cortex. Mol. Pain 2018, 14, 1744806918814367. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.R.; Nackley, A.; Huh, Y.; Terrando, N.; Maixner, W. Neuroinflammation and Central Sensitization in Chronic and Widespread Pain. Anesthesiology 2018, 129, 343–366. [Google Scholar] [CrossRef] [PubMed]
- Bouchatta, O.; Manouze, H.; Bouali-benazzouz, R.; Kerekes, N.; Ba-M’hamed, S.; Fossat, P.; Landry, M.; Bennis, M. Neonatal 6-OHDA Lesion Model in Mouse Induces Attention-Deficit/Hyperactivity Disorder (ADHD)-like Behaviour. Sci. Rep. 2018, 8, 15349. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Sarasúa, S.; Moustafa, S.; García-Avilés, Á.; López-Climent, M.F.; Gómez-Cadenas, A.; Olucha-Bordonau, F.E.; Sánchez-Pérez, A.M. The Effect of Abscisic Acid Chronic Treatment on Neuroinflammatory Markers and Memory in a Rat Model of High-Fat Diet Induced Neuroinflammation. Nutr. Metab. 2016, 13, 73. [Google Scholar] [CrossRef]
- Espinosa-Fernández, V.; Mañas-Ojeda, A.; Pacheco-Herrero, M.; Castro-Salazar, E.; Ros-Bernal, F.; Sánchez-Pérez, A.M. Early Intervention with ABA Prevents Neuroinflammation and Memory Impairment in a Triple Transgenic Mice Model of Alzheimer’s Disease. Behav. Brain Res. 2019, 374, 112106. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K.B.J. Paxinos and Franklin’s the Mouse Brain in Stereotaxic Coordinates; Elsevier: Cambridge, MA, USA, 2019. [Google Scholar]
- Davis, K.E.; Burnett, K.; Gigg, J. Water and T-Maze Protocols Are Equally Efficient Methods to Assess Spatial Memory in 3xTg Alzheimer’s Disease Mice. Behav. Brain Res. 2017, 331, 54–66. [Google Scholar] [CrossRef]
- Komada, M.; Takao, K.; Miyakawa, T. Elevated plus Maze for Mice. J. Vis. Exp. 2008, 22, e1088. [Google Scholar] [CrossRef]
- Zaky, A.; Bouali-Benazzouz, R.; Favereaux, A.; Tell, G.; Landry, M. APE1/Ref-1 Redox Function Contributes to Inflammatory Pain Sensitization. Exp. Neurol. 2018, 307, 1–11. [Google Scholar] [CrossRef]
- Sánchez-Sarasúa, S.; Ribes-Navarro, A.; Beltrán-Bretones, M.T.; Sánchez-Pérez, A.M. AAV Delivery of ShRNA against IRS1 in GABAergic Neurons in Rat Hippocampus Impairs Spatial Memory in Females and Male Rats. Anat. Embryol. 2020, 226, 163–178. [Google Scholar] [CrossRef]
- Karperien, A. “FracLac for ImageJ”. Available online: https://imagej.nih.gov/ij/plugins/fraclac/FLHelp/Introduction.htm (accessed on 7 June 2022).
- Solanto, M.V. Dopamine Dysfunction in AD/HD: Integrating Clinical and Basic Neuroscience Research. Behav. Brain Res. 2002, 130, 65–71. [Google Scholar] [CrossRef]
- Avale, M.E.; Falzone, T.L.; Gelman, D.M.; Low, M.J.; Grandy, D.K.; Rubinstein, M. The Dopamine D4 Receptor Is Essential for Hyperactivity and Impaired Behavioral Inhibition in a Mouse Model of Attention Deficit/Hyperactivity Disorder. Mol. Psychiatry 2004, 9, 718–726. [Google Scholar] [CrossRef]
- Vidal, P.M.; Pacheco, R. Targeting the Dopaminergic System in Autoimmunity. J. Neuroimmune Pharmacol. 2020, 15, 57–73. [Google Scholar] [CrossRef]
- Broome, S.T.; Louangaphay, K.; Keay, K.; Leggio, G.; Musumeci, G.; Castorina, A. Dopamine: An Immune Transmitter. Neural Regen. Res. 2020, 15, 2173–2185. [Google Scholar] [CrossRef]
- Ribes-Navarro, A.; Atef, M.; Sánchez-Sarasúa, S.; Beltrán-Bretones, M.T.; Olucha-Bordonau, F.; Sánchez-Pérez, A.M. Abscisic Acid Supplementation Rescues High Fat Diet-Induced Alterations in Hippocampal Inflammation and IRSs Expression. Mol. Neurobiol. 2018, 56, 454–464. [Google Scholar] [CrossRef]
- Bouchatta, O.; Manouze, H.; Ba-M’Hamed, S.; Landry, M.; Bennis, M. Neonatal 6-OHDA Lesion Model in Mouse Induces Cognitive Dysfunctions of Attention-Deficit/Hyperactivity Disorder (ADHD) During Young Age. Front. Behav. Neurosci. 2020, 14, 27. [Google Scholar] [CrossRef] [PubMed]
- Kurzina, N.; Belskaya, A.; Gromova, A.; Ignashchenkova, A.; Gainetdinov, R.R.; Volnova, A. Modulation of Spatial Memory Deficit and Hyperactivity in Dopamine Transporter Knockout Rats via A2A-Adrenoceptors. Front. Psychiatry 2022, 13, 503. [Google Scholar] [CrossRef]
- Slobodin, O.; Davidovitch, M. Gender Differences in Objective and Subjective Measures of ADHD Among Clinic-Referred Children. Front. Hum. Neurosci. 2019, 13, 441. [Google Scholar] [CrossRef]
- De Sousa Macedo, L.L.B.; Antunes, F.T.T.; de Andrade Alvarenga, W.; Batista, M.C.C.; de Moura, M.S.B.; Farias, M.N.L.; Caminski, E.S.; Dallegrave, E.; Grivicich, I.; de Souza, A.H. Curcumin for Attention-Deficit-Hyperactivity Disorder: A Systematic Review and Preliminary Behavioral Investigation. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2022, 395, 803–813. [Google Scholar] [CrossRef]
- Denninger, J.K.; Smith, B.M.; Kirby, E.D. Novel Object Recognition and Object Location Behavioral Testing in Mice on a Budget. J. Vis. Exp. 2018, 2018, e58593. [Google Scholar] [CrossRef]
- Bishop, C.; Mulraney, M.; Rinehart, N.; Sciberras, E. An Examination of the Association between Anxiety and Social Functioning in Youth with ADHD: A Systematic Review. Psychiatry Res. 2019, 273, 402–421. [Google Scholar] [CrossRef] [PubMed]
- Ros, R.; Graziano, P.A. Social Functioning in Children with or At Risk for Attention Deficit/Hyperactivity Disorder: A Meta-Analytic Review. J. Clin. Child. Adolesc Psychol. 2018, 47, 213–235. [Google Scholar] [CrossRef] [PubMed]
- Greene, R.W.; Biederman, J.; Faraone, S.V.; Monuteaux, M.C.; Mick, E.; Dupre, E.P.; Fine, C.S.; Goring, J.C. Social Impairment in Girls with ADHD: Patterns, Gender Comparisons, and Correlates. J. Am. Acad. Child Adolesc. Psychiatry 2001, 40, 704–710. [Google Scholar] [CrossRef] [PubMed]
- Biele, G.; Overgaard, K.R.; Friis, S.; Zeiner, P.; Aase, H. Cognitive, Emotional, and Social Functioning of Preschoolers with Attention Deficit Hyperactivity Problems. BMC Psychiatry 2022, 22, 78. [Google Scholar] [CrossRef] [PubMed]
- Anastopoulos, A.D.; Smith, T.F.; Garrett, M.E.; Morrissey-Kane, E.; Schatz, N.K.; Sommer, J.L.; Kollins, S.H.; Ashley-Koch, A. Self-Regulation of Emotion, Functional Impairment, and Comorbidity Among ChildrenWith AD/HD. J. Atten. Disord. 2011, 15, 583–592. [Google Scholar] [CrossRef]
- Yue, X.; Liu, L.; Chen, W.; Preece, D.A.; Liu, Q.; Li, H.; Wang, Y.; Qian, Q. Affective-Cognitive-Behavioral Heterogeneity of Attention-Deficit/Hyperactivity Disorder (ADHD): Emotional Dysregulation as a Sentinel Symptom Differentiating “ADHD-Simplex” and “ADHD-Complex” Syndromes? J. Affec.t Disord. 2022, 307, 133–141. [Google Scholar] [CrossRef]
- McQuade, J.D. ADHD Symptoms, Peer Problems, and Emotion Dysregulation as Longitudinal and Concurrent Predictors of Adolescent Borderline Personality Features. J. Atten. Disord. 2022, 26, 1711–1724. [Google Scholar] [CrossRef]
- Bruton, A.M.; Senders, A.; Tost, G.; Ast, H.; Robinette, L.M.; Leung, B.; Hatsu, I.E.; Arnold, L.E.; Johnstone, J.M. Pain Sensitivity and Perceptual Sensitivity Are Associated with Severity of Emotional Dysregulation in Children with ADHD: A Cross-Sectional Analysis Using the Temperament in Middle Childhood Questionnaire. Disabil. Rehabil. 2022. ahead of print. [Google Scholar] [CrossRef]
- Northover, C.; Thapar, A.; Langley, K.; van Goozen, S. Pain Sensitivity in Adolescent Males with Attention-Deficit/Hyperactivity Disorder: Testing for Associations with Conduct Disorder and Callous and Unemotional Traits. PLoS ONE 2015, 10, e0134417. [Google Scholar] [CrossRef]
- Stickley, A.; Koyanagi, A.; Takahashi, H.; Kamio, Y. ADHD Symptoms and Pain among Adults in England. Psychiatry Res. 2016, 246, 326–331. [Google Scholar] [CrossRef]
- Wolff, N.; Rubia, K.; Knopf, H.; Hölling, H.; Martini, J.; Ehrlich, S.; Roessner, V. Reduced Pain Perception in Children and Adolescents with ADHD Is Normalized by Methylphenidate. Child Adolesc. Psychiatry Ment. Health 2016, 10, 24. [Google Scholar] [CrossRef]
- Ogata, M.; Noda, K.; Akita, H.; Ishibashi, H. Characterization of Nociceptive Response to Chemical, Mechanical, and Thermal Stimuli in Adolescent Rats with Neonatal Dopamine Depletion. Neuroscience 2015, 289, 43–55. [Google Scholar] [CrossRef]
- Bouchatta, O.; Aby, F.; Sifeddine, W.; Bouali-Benazzouz, R.; Brochoire, L.; Manouze, H.; Fossat, P.; Ba M’Hamed, S.; Bennis, M.; Landry, M. Pain Hypersensitivity in a Pharmacological Mouse Model of Attention-Deficit/Hyperactivity Disorder. Proc. Natl. Acad. Sci. USA 2022, 119, e2114094119. [Google Scholar] [CrossRef]
- Nakajima, R.; Hattori, S.; Funasaka, T.; Huang, F.L.; Miyakawa, T. Decreased Nesting Behavior, Selective Increases in Locomotor Activity in a Novel Environment, and Paradoxically Increased Open Arm Exploration in Neurogranin Knockout Mice. Neuropsychopharmacol. Rep. 2021, 41, 111–116. [Google Scholar] [CrossRef]
- Chung, A.S.; Miller, S.M.; Sun, Y.; Xu, X.; Zweifel, L.S. Sexual Congruency in the Connectome and Translatome of VTA Dopamine Neurons. Sci. Rep. 2017, 7, 11120. [Google Scholar] [CrossRef]
- Monroe, T.B.; Gore, J.C.; Bruehl, S.P.; Benningfield, M.M.; Dietrich, M.S.; Chen, L.M.; Newhouse, P.; Fillingim, R.; Chodkowski, B.A.; Atalla, S.; et al. Sex Differences in Psychophysical and Neurophysiological Responses to Pain in Older Adults: A Cross-Sectional Study. Biol. Sex Differ. 2015, 6, 25. [Google Scholar] [CrossRef]
- Riley, J.L.; Robinson, M.E.; Wise, E.A.; Myers, C.D.; Fillingim, R.B. Sex Differences in the Perception of Noxious Experimental Stimuli: A Meta-Analysis. Pain 1998, 74, 181–187. [Google Scholar] [CrossRef]
- Unruh, A.M. Gender Variations in Clinical Pain Experience. Pain 1996, 65, 123–167. [Google Scholar] [CrossRef]
- Mogil, J.S.; Chesler, E.J.; Wilson, S.G.; Juraska, J.M.; Sternberg, W.F. Sex Differences in Thermal Nociception and Morphine Antinociception in Rodents Depend on Genotype. Neurosci. Biobehav. Rev. 2000, 24, 375–389. [Google Scholar] [CrossRef]
- Maixner, D.W.; Christy, D.; Kong, L.; Viatchenko-Karpinski, V.; Horner, K.A.; Hooks, S.B.; Weng, H.-R. Phytohormone Abscisic Acid Ameliorates Neuropathic Pain via Regulating LANCL2 Protein Abundance and Glial Activation at the Spinal Cord. Mol. Pain 2022, 18, 174480692211077. [Google Scholar] [CrossRef]
- Becker, S.; Gandhi, W.; Schweinhardt, P. Cerebral Interactions of Pain and Reward and Their Relevance for Chronic Pain. Neurosci. Lett. 2012, 520, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Leknes, S.; Brooks, J.C.W.; Wiech, K.; Tracey, I. Pain Relief as an Opponent Process: A Psychophysical Investigation. Eur. J. Neurosci. 2008, 28, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Arjona, M.d.M.; Grondona, J.M.; Granados-Durán, P.; Fernández-Llebrez, P.; López-Ávalos, M.D. Microglia Morphological Categorization in a Rat Model of Neuroinflammation by Hierarchical Cluster and Principal Components Analysis. Front. Cell. Neurosci. 2017, 11, 235. [Google Scholar] [CrossRef] [PubMed]
- Young, K.; Morrison, H. Quantifying Microglia Morphology from Photomicrographs of Immunohistochemistry Prepared Tissue Using ImageJ. J. Vis. Exp. 2018, 2018, e57648. [Google Scholar] [CrossRef]
- Hovens, I.; Nyakas, C.; Schoemaker, R. A Novel Method for Evaluating Microglial Activation Using Ionized Calcium-Binding Adaptor Protein-1 Staining: Cell Body to Cell Size Ratio. Neuroimmunol. Neuroinflamm. 2014, 1, 82–88. [Google Scholar] [CrossRef]
- Matisz, C.E.; Gruber, A.J. Neuroinflammatory Remodeling of the Anterior Cingulate Cortex as a Key Driver of Mood Disorders in Gastrointestinal Disease and Disorders. Neurosci. Biobehav. Rev. 2022, 133, 104497. [Google Scholar] [CrossRef]
- Choi, S.; Kim, K.; Kwon, M.; Bai, S.J.; Cha, M.; Lee, B.H. Modulation of Neuropathic Pain by Glial Regulation in the Insular Cortex of Rats. Front. Mol. Neurosci. 2022, 15, 815945. [Google Scholar] [CrossRef]
- Corona, J.C. Role of Oxidative Stress and Neuroinflammation in Attention-Deficit/Hyperactivity Disorder. Antioxidants 2020, 9, 1039. [Google Scholar] [CrossRef]
- Caston, R.A.; Gampala, S.; Armstrong, L.; Messmann, R.A.; Fishel, M.L.; Kelley, M.R. The Multifunctional APE1 DNA Repair-Redox Signaling Protein as a Drug Target in Human Disease. Drug Discov. Today 2021, 26, 218–228. [Google Scholar] [CrossRef]
- Mijit, M.; Caston, R.; Gampala, S.; Fishel, M.L.; Fehrenbacher, J.; Kelley, M.R. APE1/Ref-1—One Target with Multiple Indications: Emerging Aspects and New Directions. J. Cell. Signal. 2021, 2, 151–161. [Google Scholar]
- Sahakian, L.; Filippone, R.T.; Stavely, R.; Robinson, A.M.; Yan, X.S.; Abalo, R.; Eri, R.; Bornstein, J.C.; Kelley, M.R.; Nurgali, K. Inhibition of APE1/Ref-1 Redox Signaling Alleviates Intestinal Dysfunction and Damage to Myenteric Neurons in a Mouse Model of Spontaneous Chronic Colitis. Inflamm. Bowel Dis. 2021, 27, 388–406. [Google Scholar] [CrossRef]
- Wu, L.; Jiang, C.; Kang, Y.; Dai, Y.; Fang, W.; Huang, P. Curcumin Exerts Protective Effects against Hypoxia-reoxygenation Injury via the Enhancement of Apurinic/Apyrimidinic Endonuclease 1 in SH-SY5Y Cells: Involvement of the PI3K/AKT Pathway. Int. J. Mol. Med. 2020, 45, 993–1004. [Google Scholar] [CrossRef]
- Zaky, A.; Bassiouny, A.; Farghaly, M.; El-Sabaa, B.M. A Combination of Resveratrol and Curcumin Is Effective Against Aluminum Chloride-Induced Neuroinflammation in Rats. J. Alzheimers. Dis. 2017, 60, S221–S222. [Google Scholar] [CrossRef]
- Torregrosa-Muñumer, R.; Gómez, A.; Vara, E.; Kireev, R.; Barja, G.; Tresguerres, J.A.F.; Gredilla, R. Reduced Apurinic/Apyrimidinic Endonuclease 1 Activity and Increased DNA Damage in Mitochondria Are Related to Enhanced Apoptosis and Inflammation in the Brain of Senescence- Accelerated P8 Mice (SAMP8). Biogerontology 2016, 17, 325–335. [Google Scholar] [CrossRef]
- Yang, J.L.; Lin, Y.T.; Chuang, P.C.; Bohr, V.A.; Mattson, M.P. BDNF and Exercise Enhance Neuronal DNA Repair by Stimulating CREB-Mediated Production of Apurinic/Apyrimidinic Endonuclease 1. Neuromolecular Med. 2014, 16, 161–174. [Google Scholar] [CrossRef]
- Zaky, A.; Mohammad, B.; Moftah, M.; Kandeel, K.M.; Bassiouny, A.R. Apurinic/Apyrimidinic Endonuclease 1 Is a Key Modulator of Aluminum-Induced Neuroinflammation. BMC Neurosci. 2013, 14, 26. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meseguer-Beltrán, M.; Sánchez-Sarasúa, S.; Landry, M.; Kerekes, N.; Sánchez-Pérez, A.M. Targeting Neuroinflammation with Abscisic Acid Reduces Pain Sensitivity in Females and Hyperactivity in Males of an ADHD Mice Model. Cells 2023, 12, 465. https://doi.org/10.3390/cells12030465
Meseguer-Beltrán M, Sánchez-Sarasúa S, Landry M, Kerekes N, Sánchez-Pérez AM. Targeting Neuroinflammation with Abscisic Acid Reduces Pain Sensitivity in Females and Hyperactivity in Males of an ADHD Mice Model. Cells. 2023; 12(3):465. https://doi.org/10.3390/cells12030465
Chicago/Turabian StyleMeseguer-Beltrán, María, Sandra Sánchez-Sarasúa, Marc Landry, Nora Kerekes, and Ana María Sánchez-Pérez. 2023. "Targeting Neuroinflammation with Abscisic Acid Reduces Pain Sensitivity in Females and Hyperactivity in Males of an ADHD Mice Model" Cells 12, no. 3: 465. https://doi.org/10.3390/cells12030465
APA StyleMeseguer-Beltrán, M., Sánchez-Sarasúa, S., Landry, M., Kerekes, N., & Sánchez-Pérez, A. M. (2023). Targeting Neuroinflammation with Abscisic Acid Reduces Pain Sensitivity in Females and Hyperactivity in Males of an ADHD Mice Model. Cells, 12(3), 465. https://doi.org/10.3390/cells12030465