
T12-L3 Nerve Transfer-Induced Locomotor Recovery in Rats with Thoracolumbar Contusion: Essential Roles of Sensory Input Rerouting and Central Neuroplasticity


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, SCI Modeling, and Perioperative Management

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group | SCI Model or Laminectomy | N | T12 Intraparenchymal Implantation of hMSCs | T12-L3 Nerve Transfer (NT) | FGF2 Injection (i.m.) | EEE Cage Housing |
|---|---|---|---|---|---|---|
| 1. LAM/NTsham | T13-L1 Laminectomy and sham NT (control) | 12 | None | Bilateral, n = 12 | None | No |
| 2. SCImi | Mild contusion (10 g × 12.5 mm) only (control) | 14 | None | None | None | Starting 1 week post SCI for 4 weeks |
| 3. NTsami | Mild contusion with subacute (7–9 days p.i.) T12-L3 NT | 14 | None | Unilateral, n = 7 Bilateral, n = 7 | None | No |
| 4. NTdmi | Mild contusion with delayed (8 weeks p.i.) T12-L3 NT | 8 | Injection of 4 µL hMSCs (5 × 104 cells/µL) per rat done 7 weeks p.i. | Unilateral, n = 3 Bilateral, n = 5 | Administration of 0.8 μg FGF2 per rat in the quadriceps | Starting 1 week post NT for 4 weeks |
| 5. SCImo | Moderate contusion (10 g × 25 mm) only (control) | 18 | None | None | None | Starting 1 week post SCI for 4 weeks |
| 6. NTsamo | Moderate contusion with subacute (7–9 days p.i.) T12-L3 NT | 14 | None | Unilateral, n = 7 Bilateral, n = 7 | None | No |
| 7. NTdmo | Moderate contusion with delayed (8 weeks p.i.) T12-L3 NT | 12 | Injection of 4 µL hMSCs (5 × 104 cells/µL) per rat done 7 weeks p.i. | Unilateral, n = 7 Bilateral, n = 7 | Administration of 0.8 μg FGF2 per rat in the quadriceps | Starting 1 week post NT for 4 weeks |
| 8. Naïve | None | 7 | None | None | None | No |
2.2. Experimental Design
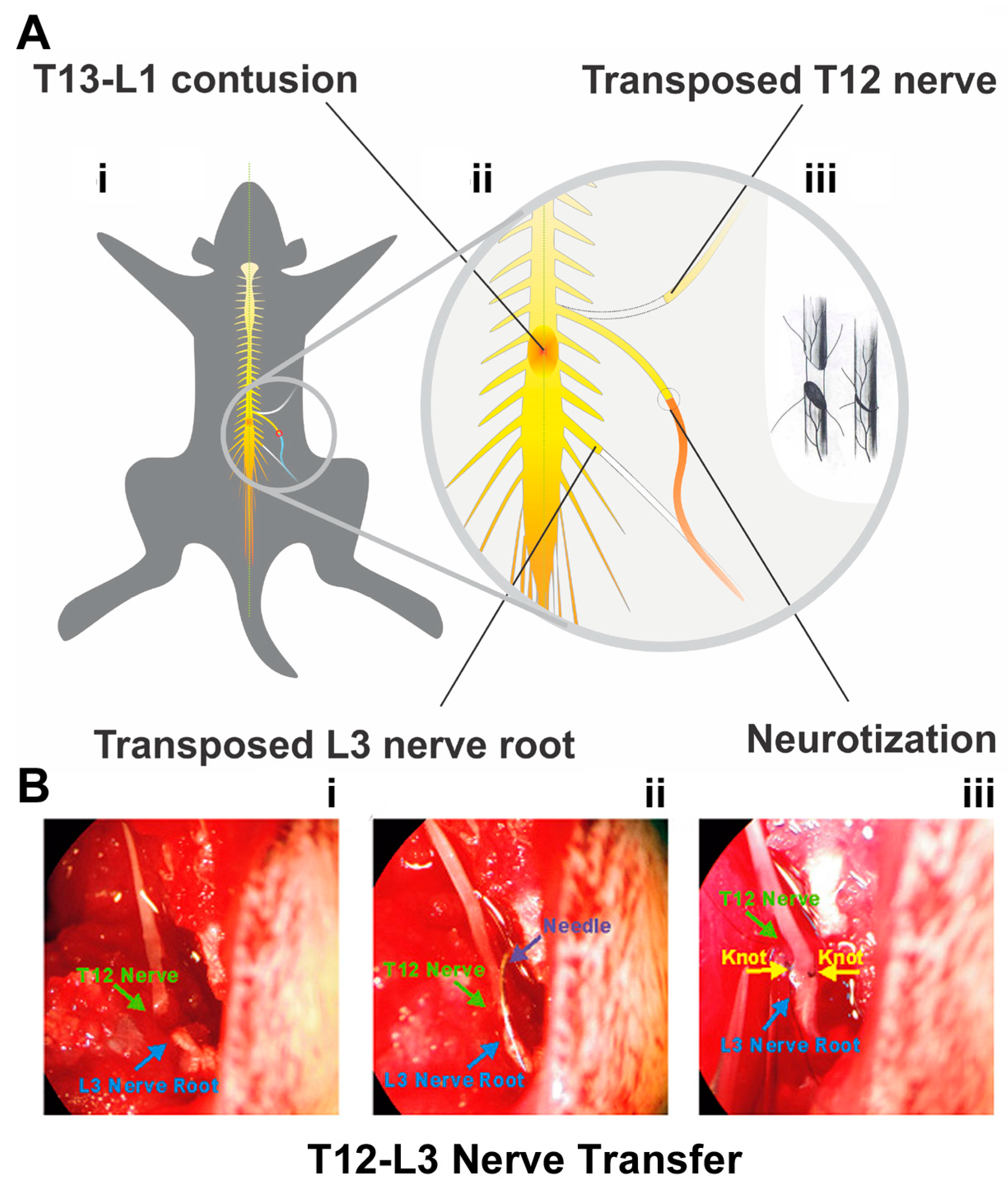
2.3. Thoracic 12 (T12) Intercostal Nerve and Lumbar 3 (L3) Nerve Root Transfer
2.4. Human MSC (hMSC) Implantation
2.5. Administration of FGF2
2.6. EEE Housing
2.7. Pharmacological Verification of Neuromodulation
2.8. Dorsal Rhizotomy and Ventral Root Transection
2.9. Behavioral Evaluations
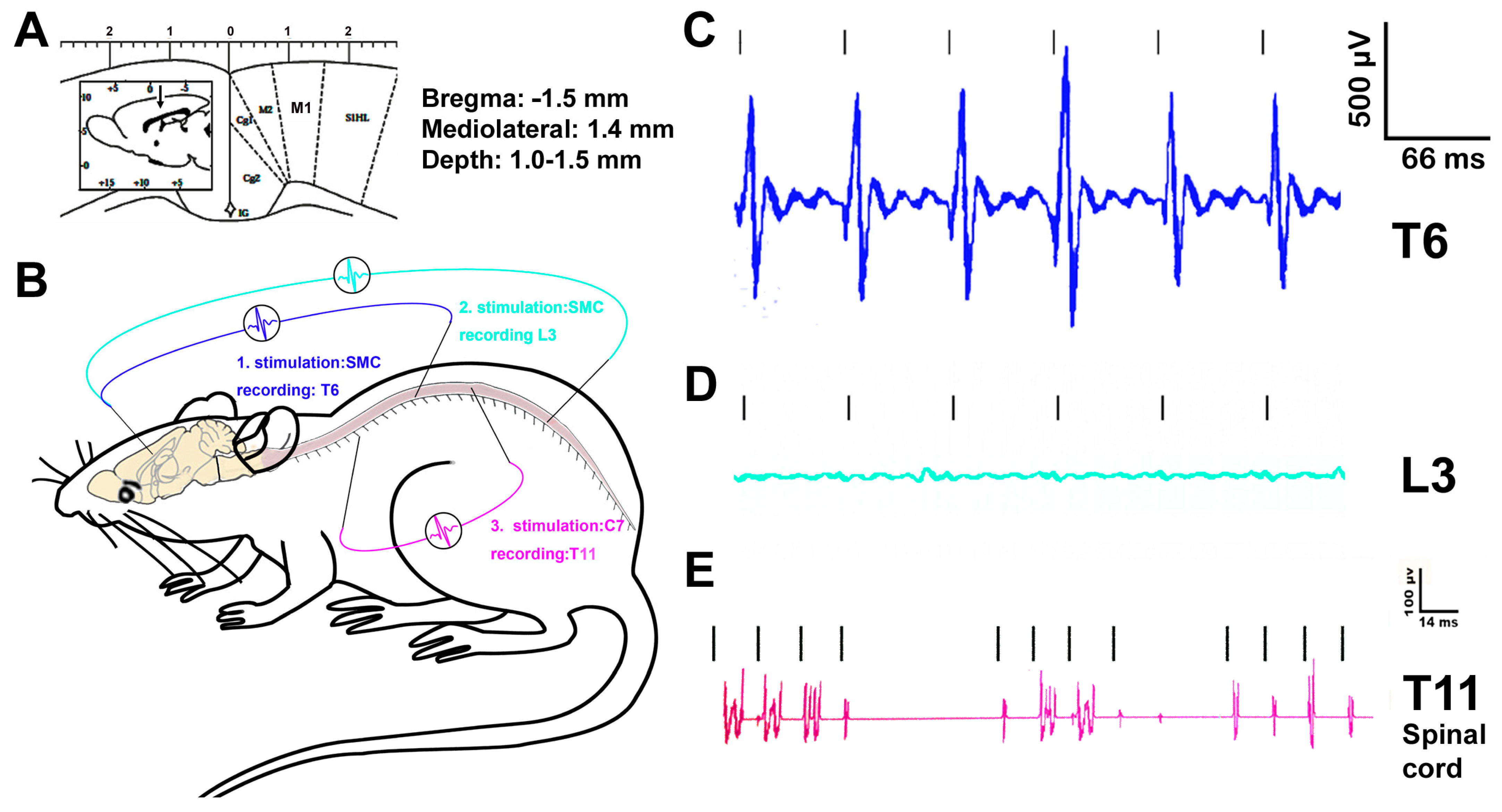
2.10. Electrophysiological Evaluations
2.11. Regular and Trans-Synaptic Neural Tracing
2.12. Neuromuscular Junction Labeling with α-Bungarotoxin
2.13. Tissue Preparation
2.14. Histopathology
2.15. IHC Assays
2.16. Statistical Analysis
3. Results
3.1. Hindlimb Motor Dysfunction following T13-L1 Contusion
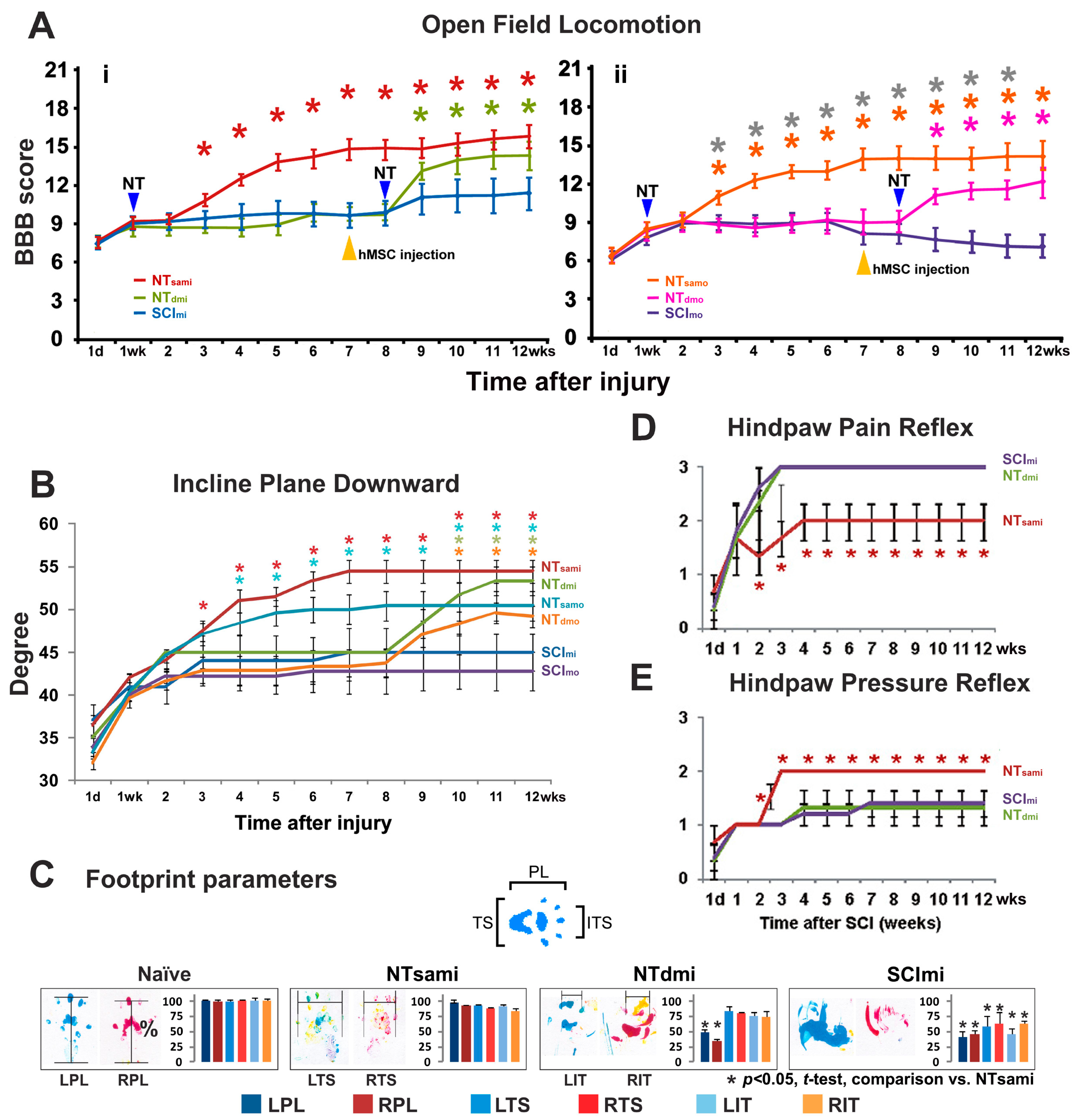
3.2. T12-L3 NT-Mediated Recovery of Coordinated Hindlimb Functions after T13-L1 Contusion
3.3. T12-L3 NT-Induced Sensory Reflex Improvement
3.4. The Impact of T12 Dorsal Root Input on the T12-L3 NT-Facilitated Locomotion
3.5. The Effect of 5-HT Neuromodulation on the T12-L3 NT-Mediated Locomotor Recovery
3.6. Electrophysiological Evaluation of the Effect of T12-L3 NT on the Neurocircuit Integrity
3.7. Histopathological Outcomes
3.8. Neural and Neuromuscular Tracing and IHC Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- National Spinal Cord Injury Statistical Center, University of Alabama. Traumatic Spinal Cord Injury Facts and Figures at a Glance [Fact Sheet]. 2022. Available online: https://www.nscisc.uab.edu/public/Facts%20and%20Figures%202022%20-%20English%20Final.pdf (accessed on 7 September 2022).
- Ahuja, C.S.; Wilson, J.R.; Nori, S.; Kotter, M.; Druschel, C.; Curt, A.; Fehlings, M.G. Traumatic spinal cord injury. Nat. Rev. Dis. Primers 2017, 3, 17018. [Google Scholar] [CrossRef] [PubMed]
- Pizzolato, C.; Gunduz, M.A.; Palipana, D.; Wu, J.; Grant, G.; Hall, S.; Dennison, R.; Zafonte, R.D.; Lloyd, D.G.; Teng, Y.D. Non-invasive approaches to functional recovery after spinal cord injury: Therapeutic targets and multimodal device interventions. Exp. Neurol. 2021, 339, 113612. [Google Scholar] [CrossRef]
- Vardjan, N.; Horvat, A.; Anderson, J.E.; Yu, D.; Croom, D.; Zeng, X.; Lužnik, Z.; Kreft, M.; Teng, Y.D.; Kirov, S.A.; et al. Adrenergic activation attenuates astrocyte swelling induced by hypotonicity and neurotrauma. Glia 2016, 64, 1034–1049. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, C.S.; Badhiwala, J.H.; Fehlings, M.G. “Time is spine”: The importance of early intervention for traumatic spinal cord injury. Spinal Cord 2020, 58, 1037–1039. [Google Scholar] [CrossRef] [PubMed]
- Baroncini, A.; Maffulli, N.; Eschweiler, J.; Tingart, M.; Migliorini, F. Pharmacological management of secondary spinal cord injury. Expert Opin. Pharmacother. 2021, 22, 1793–1800. [Google Scholar] [CrossRef]
- Teng, Y.D. Functional Multipotency of Stem Cells and Recovery Neurobiology of Injured Spinal Cords. Cell Transplant. 2019, 28, 451–459. [Google Scholar] [CrossRef]
- Kahn, L.C.; Evans, A.G.; Hill, E.J.R.; Fox, I.K. Donor activation focused rehabilitation approach to hand closing nerve transfer surgery in individuals with cervical level spinal cord injury. Spinal Cord Ser. Cases 2022, 8, 47. [Google Scholar] [CrossRef]
- Guth, L. Recovery following vagosympathetic anastomosis in the cat. Am. J. Physiol. 1956, 185, 205–208. [Google Scholar] [CrossRef]
- Gauthier, P.; Baussart, B.; Stamegna, J.C.; Tadie, M.; Vinit, S. Diaphragm recovery by laryngeal innervation after bilateral phrenicotomy or complete C2 spinal section in rats. Neurobiol. Dis. 2006, 24, 53–66. [Google Scholar] [CrossRef]
- Ray, W.Z.; Chang, J.; Hawasli, A.; Wilson, T.J.; Yang, L. Motor Nerve Transfers: A Comprehensive Review. Neurosurgery 2016, 78, 1–26. [Google Scholar] [CrossRef]
- Emamhadi, M.; Haghani Dogahe, M.; Gohritz, A. Nerve transfers in tetraplegia: A review and practical guide. J. Neurosurg. Sci. 2021, 65, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Javeed, S.; Dibble, C.F.; Greenberg, J.K.; Zhang, J.K.; Khalifeh, J.M.; Park, Y.; Wilson, T.J.; Zager, E.L.; Faraji, A.H.; Mahan, M.A.; et al. Upper Limb Nerve Transfer Surgery in Patients With Tetraplegia. JAMA Network Open 2022, 5, 2243890. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.J. Novel Uses of Nerve Transfers. Neurotherapeutics 2019, 16, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Sangalang, V.E.; Buhr, A.J.; Malik, H.G. Intercostal nerve transfer to lumbar nerve roots. Part II: Neuropathologic findings in the animal model. Spine 1979, 4, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Weber, R.A.; Warner, M.R.; Verheyden, C.N.; Proctor, W.H. Functional evaluation of gap vs. abutment repair of peripheral nerves in the rat. J. Reconstr. Microsurg. 1996, 12, 159–163. [Google Scholar] [CrossRef]
- Smith, B.W.; Sakamuri, S.; Spain, D.A.; Joseph, J.R.; Yang, L.J.; Wilson, T.J. An update on the management of adult traumatic nerve injuries-replacing old paradigms: A review. J. Trauma Acute Care Surg. 2019, 86, 299–306. [Google Scholar] [CrossRef]
- Qi, P.; Novak, C.B.; Dengler, J. Upper Extremity Reconstruction in Persons with Tetraplegia: Low Utilization and Postoperative Complications. Plast. Reconstr. Surg. 2021, 148, 860–862. [Google Scholar] [CrossRef]
- Konya, D.; Choi, H.; Liao, W.L.; Yu, D.; Woodard, M.C.; Newton, K.M.; King, A.M.; Pamir, N.M.; Black, P.M.; Frontera, W.R.; et al. Functional recovery in T13-L1 hemisected rats resulting from peripheral nerve rerouting: Role of central neuroplasticity. Regen. Med. 2008, 3, 309–327. [Google Scholar] [CrossRef]
- Ropper, A.E.; Thakor, D.K.; Han, I.; Yu, D.; Zeng, X.; Anderson, J.E.; Aljuboori, Z.; Kim, S.W.; Wang, H.; Sidman, R.L.; et al. Defining recovery neurobiology of injured spinal cord by synthetic matrix-assisted hMSC implantation. Proc. Natl. Acad. Sci. USA 2017, 114, 820–829. [Google Scholar] [CrossRef]
- Yuan, X.; Balog, B.M.; Lin, D.L.; Hanzlicek, B.; Kuang, M.; Yan, H.; Majerus, S.J.A.; Damaser, M.S. Brain-Derived Neurotrophic Factor Is an Important Therapeutic Factor in Mesenchymal Stem Cell Secretions for Treatment of Traumatic Peripheral Pelvic Injuries. Front. Cell. Neurosci. 2022, 16, 866094. [Google Scholar] [CrossRef]
- Gelderd, J.B.; Chopin, S.F. The vertebral level of origin of spinal nerves in the rat. Anat. Rec. 1977, 188, 45–47. [Google Scholar] [CrossRef]
- Teng, Y.D.; Choi, H.; Onario, R.C.; Zhu, S.; Desilets, F.C.; Lan, S.; Woodard, E.J.; Snyder, E.Y.; Eichler, M.E.; Friedlander, R.M. Minocycline inhibits contusion-triggered mitochondrial cytochrome c release and mitigates functional deficits after spinal cord injury. Proc. Natl. Acad. Sci. USA 2004, 101, 3071–3076. [Google Scholar] [CrossRef]
- Kabatas, S.; Yu, D.; He, X.D.; Thatte, H.S.; Benedict, D.; Hepgul, K.T.; Black, P.M.; Sabharwal, S.; Teng, Y.D. Neural and anatomical abnormalities of the gastrointestinal system resulting from contusion spinal cord injury. Neuroscience 2008, 154, 1627–1638. [Google Scholar] [CrossRef]
- Teng, Y.D.; Bingaman, M.; Taveira-DaSilva, A.M.; Pace, P.P.; Gillis, R.A.; Wrathall, J.R. Serotonin 1A receptor agonists reverse respiratory abnormalities in spinal cord-injured rats. J. Neurosci. 2003, 23, 4182–41910. [Google Scholar] [CrossRef]
- Wang, L.; Gunduz, M.A.; Semeano, A.T.; Yılmaz, E.C.; Alanazi, F.A.H.; Imir, O.B.; Yener, U.; Arbelaez, C.A.; Usuga, E.; Teng, Y.D. Coexistence of chronic hyperalgesia and multilevel neuroinflammatory responses after experimental SCI: A systematic approach to profiling neuropathic pain. J. Neuroinflammation 2022, 19, 264. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.D.; Mocchetti, I.; Taveira-DaSilva, A.M.; Gillis, R.A.; Wrathall, J.R. Basic fibroblast growth factor increases long-term survival of spinal cord motor neurons and improves respiratory function after experimental SCI. J. Neurosci. 1999, 19, 7037–7047. [Google Scholar] [CrossRef]
- Teng, Y.D.; Mocchetti, I.; Wrathall, J.R. Basic and acidic fibroblast growth factors protect spinal motor neurones in vivo after experimental spinal cord injury. Eur. J. Neurosci. 1998, 10, 798–802. [Google Scholar] [CrossRef] [PubMed]
- Wrathall, J.R.; Choiniere, D.; Teng, Y.D. Dose-dependent reduction of tissue loss and functional impairment after spinal cord trauma with the AMPA/kainate antagonist NBQX. J. Neurosci. 1994, 14, 6598–6607. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.D.; Wrathall, J.R. Local blockade of sodium channels by tetrodotoxin ameliorates tissue loss and long-term functional deficits resulting from experimental spinal cord injury. J. Neurosci. 1997, 17, 4359–4366. [Google Scholar] [CrossRef]
- Yu, D.; Thakor, D.K.; Han, I.; Ropper, A.E.; Haragopal, H.; Sidman, R.L.; Zafonte, R.; Schachter, S.C.; Teng, Y.D. Alleviation of chronic pain following rat spinal cord compression injury with multimodal actions of huperzine A. Proc. Natl. Acad. Sci. USA 2013, 110, 746–755. [Google Scholar] [CrossRef]
- Jungnickel, J.; Haase, K.; Konitzer, J.; Timmer, M.; Grothe, C. Faster nerve regeneration after sciatic nerve injury in mice over-expressing basic fibroblast growth factor. J. Neurobiol. 2006, 66, 940–948. [Google Scholar] [CrossRef]
- Thakor, D.K.; Wang, L.; Benedict, D.; Kabatas, S.; Zafonte, R.D.; Teng, Y.D. Establishing an Organotypic System for Investigating Multimodal Neural Repair Effects of Human Mesenchymal Stromal Stem Cells. Curr. Protoc. Stem Cell Biol. 2018, 47, 58. [Google Scholar] [CrossRef]
- Zeng, X.; Han, I.; Abd-El-Barr, M.; Aljuboori, Z.; Anderson, J.E.; Chi, J.H.; Zafonte, R.D.; Teng, Y.D. The Effects of Thermal Preconditioning on Oncogenic and Intraspinal Cord Growth Features of Human Glioma Cells. Cell Transplant. 2016, 25, 2099–2109. [Google Scholar] [CrossRef] [PubMed]
- Vejsada, R.; Hník, P. Radicular innervation of hindlimb muscles of the rat. Physiol. Bohemoslov. 1980, 29, 385–392. [Google Scholar] [PubMed]
- Kohn, T.A.; Myburgh, K.H. Regional specialization of rat quadriceps myosin heavy chain isoforms occurring in distal to proximal parts of middle and deep regions is not mirrored by citrate synthase activity. J. Anat. 2007, 210, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.; Tsuji, T.; Kishimoto, Y.; Sugiyama, Y.; Nakamura, T.; Hirano, S. Regenerative Effects of Basic Fibroblast Growth Factor on Restoration of Thyroarytenoid Muscle Atrophy Caused by Recurrent Laryngeal Nerve Transection. J. Voice 2018, 32, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Grońska-Pęski, M.; Gonçalves, J.T.; Hébert, J.M. Enriched Environment Promotes Adult Hippocampal Neurogenesis through FGFRs. J. Neurosci. 2021, 41, 2899–2910. [Google Scholar] [CrossRef]
- Teng, Y.D. Method for improving respiratory function and inhibiting muscular degeneration. United States Pat. 2006, 7, 194. [Google Scholar]
- Okamoto, K.; Imbe, H.; Kimura, A.; Donishi, T.; Tamai, Y.; Senba, E. Activation of central 5HT2A receptors reduces the craniofacial nociception of rats. Neuroscience 2007, 147, 1090–1102. [Google Scholar] [CrossRef]
- Amato, D.; Pum, M.E.; Groos, D.; Lauber, A.C.; Huston, J.P.; Carey, R.J.; de Souza Silva, M.A.; Müller, C.P. Neuropharmacology of light-induced locomotor activation. Neuropharmacology 2015, 95, 243–251. [Google Scholar] [CrossRef]
- Goldberger, M.E. Locomotor recovery after unilateral hindlimb deafferentation in cats. Brain Res. 1977, 123, 59–74. [Google Scholar] [CrossRef] [PubMed]
- Basso, D.M.; Beattie, M.S.; Bresnahan, J.C. A sensitive and reliable locomotor rating scale for open field testing in rats. J. Neurotrauma 1995, 12, 1–21. [Google Scholar] [CrossRef] [PubMed]
- de Medinaceli, L.; Freed, W.J.; Wyatt, R.J. An index of the functional condition of rat sciatic nerve based on measurements made from walking tracks. Exp. Neurol. 1982, 77, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.D.; Lavik, E.B.; Qu, X.; Park, K.I.; Ourednik, J.; Zurakowski, D.; Langer, R.; Snyder, E.Y. Functional recovery following traumatic spinal cord injury mediated by a unique polymer scaffold seeded with neural stem cells. Proc. Natl. Acad. Sci. USA 2002, 99, 3024–3029. [Google Scholar] [CrossRef] [PubMed]
- Ueta, Y.; Otsuka, T.; Morishima, M.; Ushimaru, M.; Kawaguchi, Y. Multiple layer 5 pyramidal cell subtypes relay cortical feedback from secondary to primary motor areas in rats. Cereb. Cortex 2014, 24, 2362–2376. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 3rd ed.; Academic Press: San Diego, CA, USA, 1997; p. 49. [Google Scholar]
- Jang, D.K.; Park, S.I.; Han, Y.M.; Jang, K.S.; Park, M.S.; Chung, Y.A.; Kim, M.W.; Maeng, L.S.; Huh, P.W.; Yoo, D.S. Motor-evoked potential confirmation of functional improvement by transplanted bone marrow mesenchymal stem cell in the ischemic rat brain. J. Biomed. Biotechnol. 2011, 2011, 238409. [Google Scholar] [CrossRef]
- Molander, C.; Grant, G. Spinal cord cytoarchitecture. In The Rat Nervous System, 1st ed.; Academic Press: San Diego, CA, USA, 1995; pp. 39–45. [Google Scholar]
- Siebert, J.R.; Middleton, F.A.; Stelzner, D.J. Long descending cervical propriospinal neurons differ from thoracic propriospinal neurons in response to low thoracic spinal injury. BMC Neurosci. 2010, 11, 148. [Google Scholar] [CrossRef]
- Mitchell, E.J.; McCallum, S.; Dewar, D.; Maxwell, D.J. Corticospinal and Reticulospinal Contacts on Cervical Commissural and Long Descending Propriospinal Neurons in the Adult Rat Spinal Cord; Evidence for Powerful Reticulospinal Connections. PLoS ONE 2016, 11, 0152094. [Google Scholar]
- Goshgarian, H.G.; Buttry, J.L. The pattern and extent of retrograde transsynaptic transport of WGA-Alexa 488 in the phrenic motor system is dependent upon the site of application. J. Neurosci. Methods 2014, 222, 156–164. [Google Scholar] [CrossRef]
- Burns, A.S.; Jawaid, S.; Zhong, H.; Yoshihara, H.; Bhagat, S.; Murray, M.; Roy, R.R.; Tessler, A.; Son, Y.J. Paralysis by SCI evokes selective disassembly of NMJ synapses with and without terminal sprouting in ankle flexors of the adult rat. J. Comp. Neurol. 2007, 500, 116–133. [Google Scholar] [CrossRef]
- Delhaye, B.P.; Long, K.H.; Bensmaia, S.J. Neural Basis of Touch and Proprioception in Primate Cortex. Compr. Physiol. 2018, 8, 1575–1602. [Google Scholar]
- Perrin, F.E.; Noristani, H.N. Serotonergic mechanisms in spinal cord injury. Exp. Neurol. 2019, 318, 174–191. [Google Scholar] [CrossRef] [PubMed]
- Pehek, E.; Nocjar, C.; Roth, B.; Byrd, T.; Mabrouk, O. Evidence for the Preferential Involvement of 5-HT2A Serotonin Receptors in Stress-and Drug-Induced Dopamine Release in the Rat Medial Prefrontal Cortex. Neuropsychopharmacology 2006, 31, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Sławińska, U.; Jordan, L.M. Serotonergic influences on locomotor circuits. Curr. Opin. Physiol. 2019, 8, 63–69. [Google Scholar] [CrossRef]
- Bonhaus, D.W.; Bach, C.; DeSouza, A.; Salazar, F.H.; Matsuoka, B.D.; Zuppan, P.; Chan, H.W.; Eglen, R.M. The pharmacology and distribution of human 5-hydroxytryptamine2B (5-HT2B) receptor gene products: Comparison with 5-HT2A and 5-HT2C receptors. Br. J. Pharmacol. 1995, 115, 622–628. [Google Scholar] [CrossRef]
- Courtine, G.; Gerasimenko, Y.; van den Brand, R.; Yew, A.; Musienko, P.; Zhong, H.; Song, B.; Ao, Y.; Ichiyama, R.M.; Lavrov, I.; et al. Transformation of nonfunctional spinal circuits into functional states after the loss of brain input. Nat. Neurosci. 2009, 12, 1333–1342. [Google Scholar] [CrossRef]
- Zaniewska, M.; McCreary, A.C.; Wydra, K.; Filip, M. Differential effects of serotonin (5-HT)2 receptor-targeting ligands on locomotor responses to nicotine-repeated treatment. Synapse 2010, 64, 511–519. [Google Scholar] [CrossRef]
- Pędzich, B.D.; Rubens, S.; Sekssaoui, M.; Pierre, A.; Van Schuerbeek, A.; Marin, P.; Bockaert, J.; Valjent, E.; Bécamel, C.; De Bundel, D. Effects of a psychedelic 5-HT2A receptor agonist on anxiety-related behavior and fear processing in mice. Neuropsychopharmacology 2022, 47, 1304–1314. [Google Scholar] [CrossRef]
- Perrier, J.F.; Cotel, F. Serotonergic modulation of spinal motor control. Curr. Opin. Neurobiol. 2015, 33, 1–7. [Google Scholar] [CrossRef]
- Sienkiewicz, W.; Dudek, A. Sources of the motor and somatic sensory innervation of the trapezius muscle in the rat. Vet. Med. Czech 2010, 55, 242–252. [Google Scholar] [CrossRef]
- Alstermark, B.; Kümmel, H. Transneuronal transport of wheat germ agglutinin conjugated horseradish peroxidase into last order spinal interneurones projecting to acromio- and spinodeltoideus motoneurones in the cat. Exp. Brain Res. 1990, 80, 83–95. [Google Scholar] [CrossRef]
- Izumi, A.; Kida, M.Y. Segmental distribution of the motoneurons innervating trunk muscles in the spinal cord of the cat and rat. Neurosci. Res. 1998, 30, 247–255. [Google Scholar] [CrossRef]
- Alvarez, F.J.; Titus-Mitchell, H.E.; Bullinger, K.L.; Kraszpulski, M.; Nardelli, P.; Cope, T.C. Permanent central synaptic disconnection of proprioceptors after nerve injury and regeneration. I. Loss of VGLUT1/IA synapses on motoneurons. J. Neurophysiol. 2011, 106, 2450–2470. [Google Scholar] [CrossRef] [PubMed]
- Mende, M.; Fletcher, E.V.; Belluardo, J.L.; Pierce, J.P.; Bommareddy, P.K.; Weinrich, J.A.; Kabir, Z.D.; Schierberl, K.C.; Pagiazitis, J.G.; Mendelsohn, A.I.; et al. Sensory-Derived Glutamate Regulates Presynaptic Inhibitory Terminals in Mouse Spinal Cord. Neuron 2016, 90, 1189–1202. [Google Scholar] [CrossRef] [PubMed]
- Richardson, P.M.; Issa, V.M. Peripheral injury enhances central regeneration of primary sensory neurones. Nature 1984, 309, 791–793. [Google Scholar] [CrossRef] [PubMed]
- Neumann, S.; Woolf, C.J. Regeneration of dorsal column fibers into and beyond the lesion site following adult spinal cord injury. Neuron 1999, 23, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Damhieu, P.; Devanze, P.; Said, G.; Heard, J.M.; Tadie, M. Efficient reinnervation of hindlimb muscles by thoracic motor neurons after nerve cross-anastomosis in rats. J. Neurosurg. 2003, 99, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Johnston, L.; Zhang, Z.; Ma, Y.; Hu, Y.; Wang, J.; Huang, P.; Wang, S. Restoration of stepping-forward and ambulatory function in patients with paraplegia: Rerouting of vascularized intercostal nerves to lumbar nerve roots using selected interfascicular anastomosis. Surg. Technol. Int. 2003, 11, 244–248. [Google Scholar]
- Krieger, A.J.; Danetz, I.; Wu, S.Z.; Spatola, M.; Sapru, H.N. Electrophrenic respiration following anastomosis of phrenic with branchial nerve in the cat. J. Neurosurg. 1983, 59, 262–267. [Google Scholar] [CrossRef]
- de Divitiis, E.; Donzelli, R.; Caputi, F.; Crisci, C.; Gargiulo, G.; Francica, D. Experimental model of nervous anastomosis between intercostal and lumbar nerves in the rabbit. J. Neurosurg. Sci. 1984, 28, 153–156. [Google Scholar]
- Huie, J.R.; Morioka, K.; Haefeli, J.; Ferguson, A.R. What is being trained? How divergent forms of plasticity compete to shape locomotor recovery after spinal cord injury. J. Neurotrauma 2017, 34, 1831–1840. [Google Scholar] [CrossRef]
- Martin, J.H. Neuroplasticity of spinal cord injury and repair. Handb. Clin. Neurol. 2022, 184, 317–330. [Google Scholar]
- Musienko, P.E.; Bogacheva, I.N.; Gerasimenko, Y.P. Significance of peripheral feedback in the generation of stepping movements during epidural stimulation of the spinal cord. Neurosci. Behav. Physiol. 2007, 37, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Formento, E.; Minassian, K.; Wagner, F.; Mignardot, J.B.; Le Goff-Mignardot, C.G.; Rowald, A.; Bloch, J.; Micera, S.; Capogrosso, M.; Courtine, G. Electrical spinal cord stimulation must preserve proprioception to enable locomotion in humans with spinal cord injury. Nat. Neurosci. 2018, 21, 1728–1741. [Google Scholar] [CrossRef] [PubMed]
- Harnie, J.; Doelman, A.; de Vette, E.; Audet, J.; Desrochers, E.; Gaudreault, N.; Frigon, A. The recovery of standing and locomotion after spinal cord injury does not require task-specific training. Elife 2019, 8, 50134. [Google Scholar] [CrossRef]
- Harkema, S.; Gerasimenko, Y.; Hodes, J.; Burdick, J.; Angeli, C.; Chen, Y.; Ferreira, C.; Willhite, A.; Rejc, E.; Grossman, R.G.; et al. Effect of epidural stimulation of the lumbosacral spinal cord on voluntary movement, standing, and assisted stepping after motor complete paraplegia: A case study. Lancet 2011, 377, 1938–1947. [Google Scholar] [CrossRef]
- Angeli, C.A.; Boakye, M.; Morton, R.A.; Vogt, J.; Benton, K.; Chen, Y.; Ferreira, C.K.; Harkema, S.J. Recovery of over-ground walking after chronic motor complete spinal cord injury. N. Engl. J. Med. 2018, 379, 1244–1250. [Google Scholar] [CrossRef]
- McPherson, J.G.; Miller, R.R.; Perlmutter, S.I. Targeted, activity-dependent spinal stimulation produces long-lasting motor recovery in chronic cervical spinal cord injury. Proc. Natl. Acad. Sci. USA 2015, 112, 12193–12198. [Google Scholar] [CrossRef] [PubMed]
- Krucoff, M.O.; Rahimpour, S.; Slutzky, M.W.; Edgerton, V.R.; Turner, D.A. Enhancing nervous system recovery through neurobiologics, neural interface training, and neurorehabilitation. Front. Neurosci. 2016, 10, 584. [Google Scholar] [CrossRef] [PubMed]
- Sasada, S.; Kato, K.; Kadowaki, S.; Groiss, S.J.; Ugawa, Y.; Komiyama, T.; Nishimura, Y. Volitional walking via upper limb muscle-controlled stimulation of the lumbar locomotor center in man. J. Neurosci. 2014, 34, 11131–11142. [Google Scholar] [CrossRef]
- Marqueste, T.; Alliez, J.R.; Alluin, O.; Jammes, Y.; Decherchi, P. Neuromuscular rehabilitation by treadmill running or electrical stimulation after peripheral nerve injury and repair. J. Appl. Physiol. 2004, 96, 1988–1995. [Google Scholar] [CrossRef]
- Helgren, M.E.; Goldberger, M.E. The recovery of postural reflexes and locomotion following low thoracic hemisection in adult cats involves compensation by undamaged primary afferent pathways. Exp. Neurol. 1993, 123, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, C.A.; Smith, J.L. Stepping behaviors in chronic spinal cats with one hindlimb deafferented. J. Neurosci. 1987, 7, 2537–2546. [Google Scholar] [PubMed]
- Lavrov, I.; Courtine, G.; Dy, C.J.; van den Brand, R.; Fong, A.J.; Gerasimenko, Y.; Zhong, H.; Roy, R.R.; Edgerton, V.R. Facilitation of stepping with epidural stimulation in spinal rats: Role of sensory input. J. Neurosci. 2008, 28, 7774–7780. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Pang, Z.; Lei, G.; Chen, Y.; Cai, Z.; Zhu, S.; Lin, W.; Qiu, Z.; Wang, Y.; Shen, Y.; et al. Crossing nerve transfer drives sensory input-dependent plasticity for motor recovery after brain injury. Sci. Adv. 2022, 8, 5899–5918. [Google Scholar] [CrossRef] [PubMed]
- Barbeau, H.; Ladouceur, M.; Mirbagheri, M.M.; Kearney, R.E. The effect of locomotor training combined with functional electrical stimulation in chronic spinal cord injured subjects: Walking and reflex studies. Brain Res. Rev. 2002, 40, 274–291. [Google Scholar] [CrossRef] [PubMed]
- Dougherty, B.J.; Lee, K.Z.; Gonzalez-Rothi, E.J.; Lane, M.A.; Reier, P.J.; Fuller, D.D. Recovery of inspiratory intercostal muscle activity following high cervical hemisection. Respir. Physiol. Neurobiol. 2012, 183, 186–192. [Google Scholar] [CrossRef]
- Plant, G.W.; Weinrich, J.A.; Kaltschmidt, J.A. Sensory and descending motor circuitry during development and injury. Curr. Opin. Neurobiol. 2018, 53, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Siebert, J.R.; Middelton, F.A.; Stelzner, D.J. Intrinsic response of thoracic propriospinal neurons to axotomy. BMC Neurosci. 2010, 11, 69. [Google Scholar] [CrossRef]
- Spencer, T.; Filbin, M.T. A role for cAMP in regeneration of the adult mammalian CNS. J. Anat. 2004, 204, 49–55. [Google Scholar] [CrossRef]
- Teixeira, F.G.; Carvalho, M.M.; Panchalingam, K.M.; Rodrigues, A.J.; Mendes-Pinheiro, B.; Anjo, S.; Manadas, B.; Behie, L.A.; Sousa, N.; Salgado, A.J. Impact of the Secretome of Human Mesenchymal Stem Cells on Brain Structure and Animal Behavior in a Rat Model of Parkinson’s Disease. Stem Cells Transl. Med. 2017, 6, 634–646. [Google Scholar] [CrossRef]
- Tashiro, S.; Shinozaki, M.; Mukaino, M.; Renault-Mihara, F.; Toyama, Y.; Liu, M.; Nakamura, M.; Okano, H. BDNF Induced by treadmill training contributes to suppression of spasticity and allodynia after SCI via KCC2 upregulation. Neurorehabilit. Neural Repair 2015, 29, 677–689. [Google Scholar] [CrossRef]
- Madison, R.D.; Archibald, S.J.; Brushart, T.M. Reinnervation accuracy of the rat femoral nerve by motor and sensory neurons. J. Neurosci. 1996, 16, 5698–5703. [Google Scholar] [CrossRef]
- Bolívar, S.; Udina, E. Preferential regeneration and collateral dynamics of motor and sensory neurons after nerve injury in mice. Exp. Neurol. 2022, 358, 114227. [Google Scholar] [CrossRef]
- Madison, R.D.; Robinson, G.A.; Chadaram, S.R. The specificity of motor neurone regeneration (preferential reinnervation). Acta Physiol. 2007, 189, 201–206. [Google Scholar] [CrossRef] [PubMed]
- da Silva, C.F.; Madison, R.; Dikkes, P.; Chiu, T.H.; Sidman, R.L. An in vivo model to quantify motor and sensory peripheral nerve regeneration using bioresorbable nerve guide tubes. Brain Res. 1985, 342, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Madison, R.D.; da Silva, C.F.; Dikkes, P. Entubulation repair with protein additives increases the maximum nerve gap distance successfully bridged with tubular prostheses. Brain Res. 1988, 447, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, H.L.; Donoso, J.A. Trophic Mechanisms Underlying Nerve-Muscle Interactions. In Nerve-Muscle Cell Trophic Communication, 1st ed.; CRC Press: Boca Raton, FL, USA, 2020; pp. 217–234. [Google Scholar]
- Park, J.S.; Höke, A. Treadmill exercise induced functional recovery after peripheral nerve repair is associated with increased levels of neurotrophic factors. PLoS ONE 2014, 9, 90245. [Google Scholar] [CrossRef]
- Nishimune, H.; Stanford, J.A.; Mori, Y. Role of exercise in maintaining the integrity of the neuromuscular junction. Muscle Nerve 2014, 49, 315–324. [Google Scholar] [CrossRef]
- Ernfors, P.; Lee, K.F.; Kucera, J.; Jaenisch, R. Lack of neurotrophin-3 leads to deficiencies in the peripheral nervous system and loss of limb proprioceptive afferents. Cell 1994, 77, 503–512. [Google Scholar] [CrossRef]
- Hornby, T.G.; Tysseling-Mattiace, V.M.; Benz, E.N.; Schmit, B.D. Contribution of muscle afferents to prolonged flexion withdrawal reflexes in human spinal cord injury. J. Neurophysiol. 2004, 92, 3375–3384. [Google Scholar] [CrossRef]
- Goshgarian, H.G. The crossed phrenic phenomenon and recovery of function following spinal cord injury. Respir. Physiol. Neurobiol. 2009, 169, 85–93. [Google Scholar] [CrossRef]
- Dougherty, B.J.; Terada, J.; Springborn, S.R.; Vinit, S.; MacFarlane, P.M.; Mitchell, G.S. Daily acute intermittent hypoxia improves breathing function with acute and chronic spinal injury via distinct mechanisms. Respir. Physiol. Neurobiol. 2018, 256, 50–57. [Google Scholar] [CrossRef]
- Coggeshall, R.E.; Applebaum, M.L.; Fazen, M.; Stubbs, T.B., 3rd; Sykes, M.T. Unmyelinated axons in human ventral roots, a possible explanation for the failure of dorsal rhizotomy to relieve pain. Brain 1975, 98, 157–166. [Google Scholar] [CrossRef]
- Madden, J.F.; Davis, O.C.; Boyle, K.A.; Iredale, J.A.; Browne, T.J.; Callister, R.J.; Smith, D.W.; Jobling, P.; Hughes, D.I.; Graham, B.A. Functional and Molecular Analysis of Proprioceptive Sensory Neuron Excitability in Mice. Front. Mol. Neurosci. 2020, 13, 36. [Google Scholar] [CrossRef]
- Jung, R.; Belanger, A.; Kanchiku, T.; Fairchild, M.; Abbas, J.J. Neuromuscular stimulation therapy after incomplete spinal cord injury promotes recovery of interlimb coordination during locomotion. J. Neural Eng. 2009, 6, 055010. [Google Scholar] [CrossRef]
- Emamhadi, M.; Naseri, A.; Aghaei, I.; Ashrafi, M.; Emamhadi, R.; Andalib, S. Soleus nerve transfer to deep peroneal nerve for treatment of foot drop. J. Clin. Neurosci. 2020, 78, 159–163. [Google Scholar] [CrossRef]
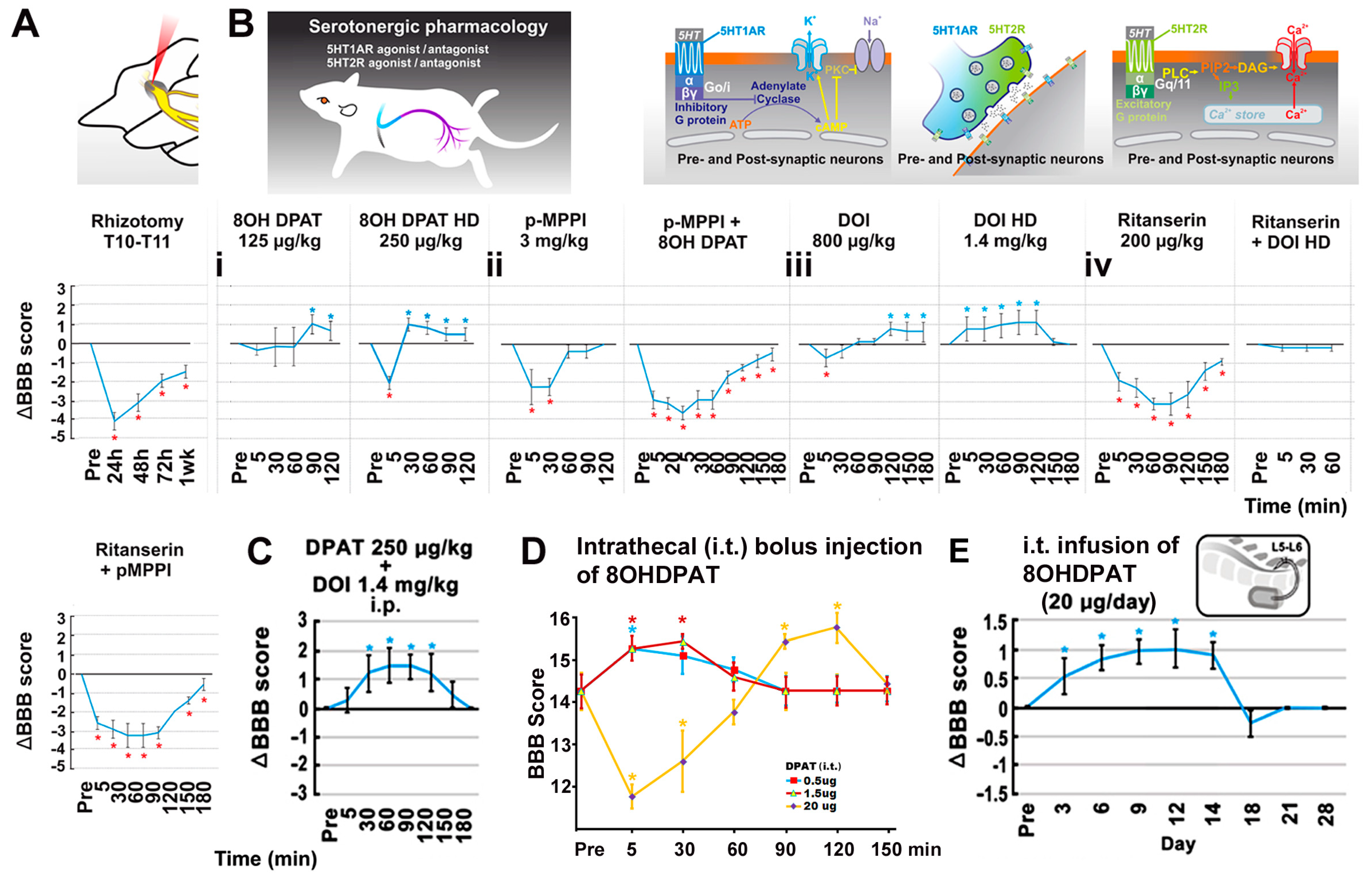
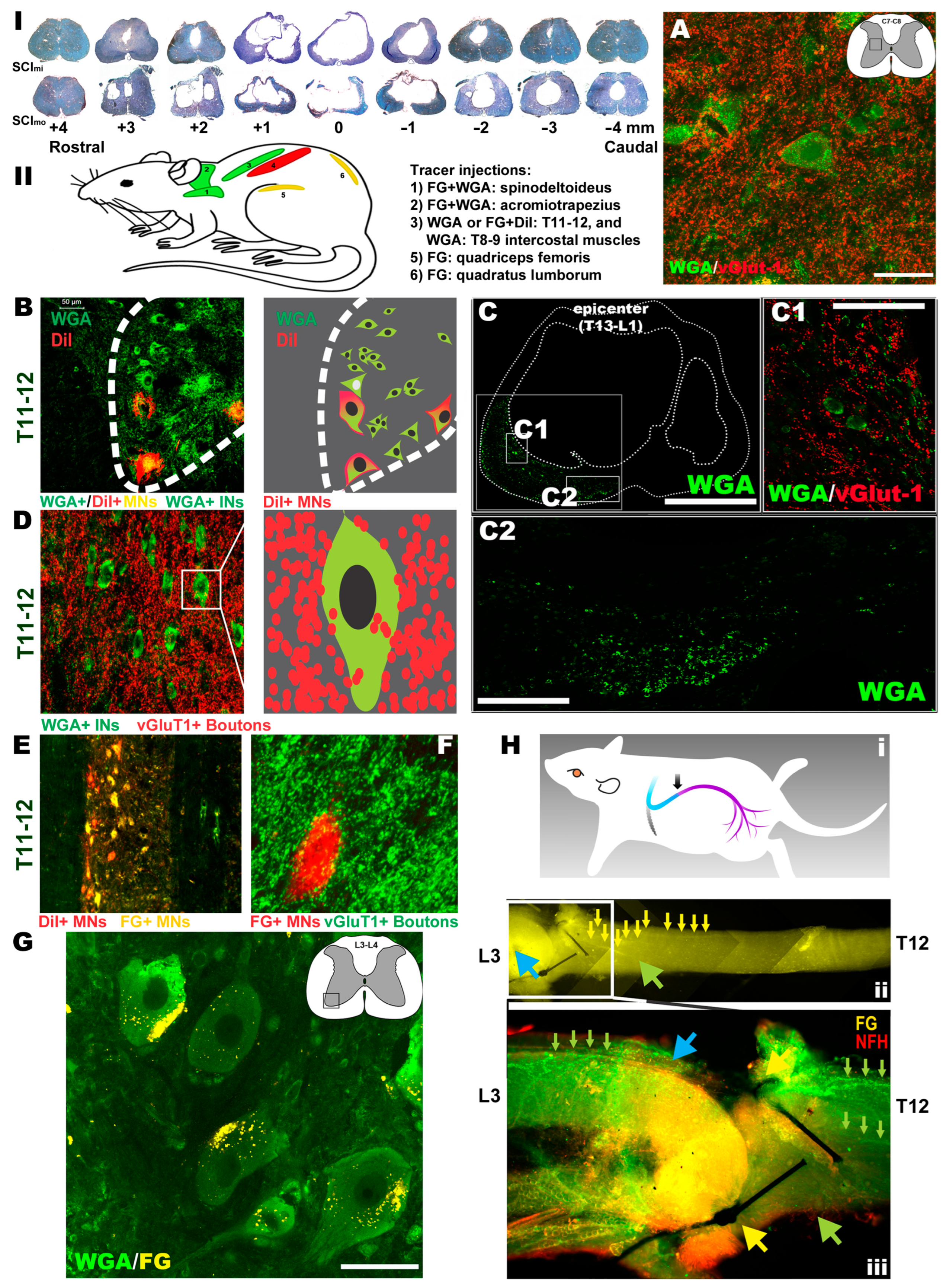
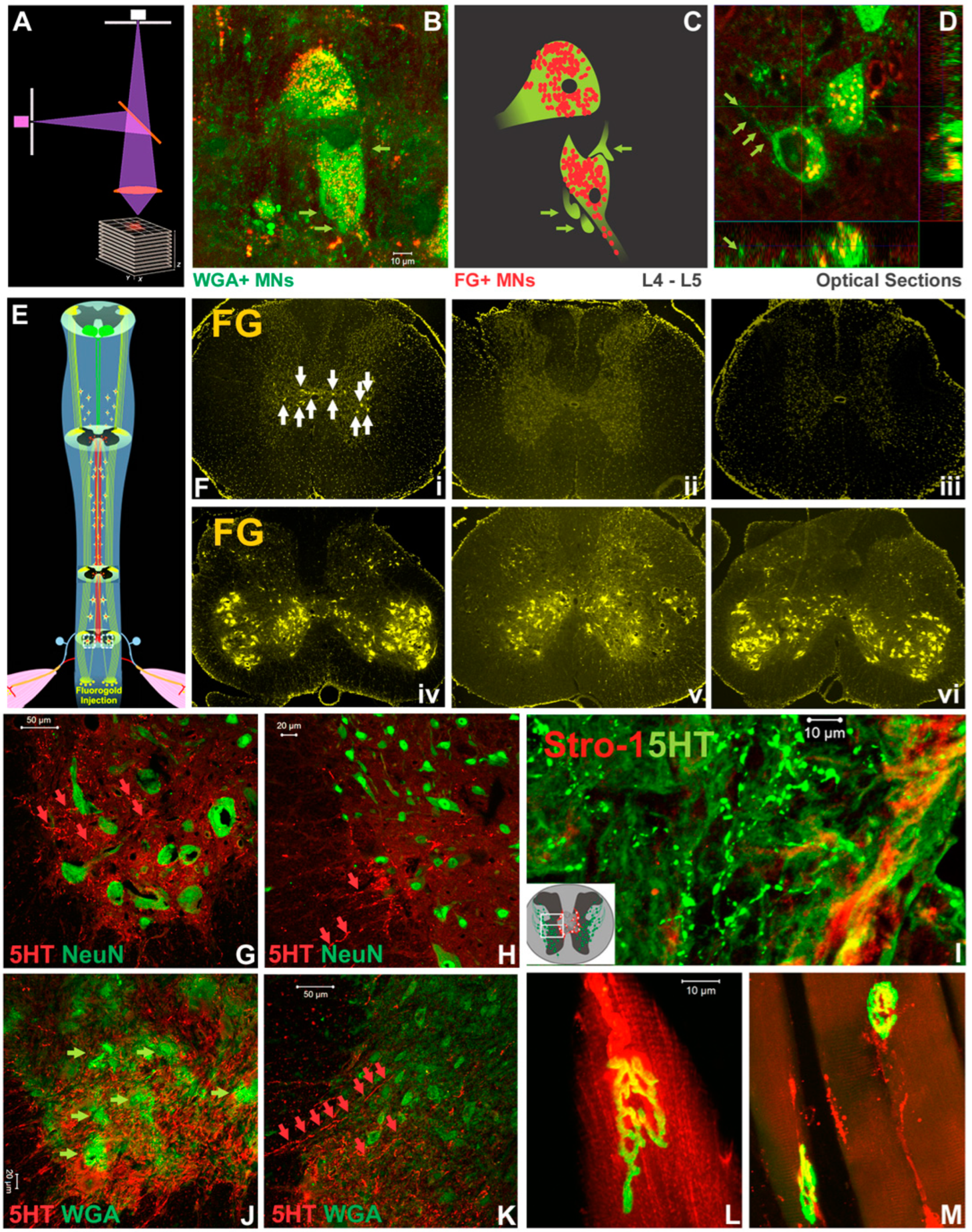
| Drug | Effect | Route | Dose | Vendor | References |
|---|---|---|---|---|---|
| 8-OH-DPAT: 8-hydroxy-2-(di-n-propylamino)tetralin | 5HT1A receptor agonist | i.p. i.t. | 0.125 or 0.25 µg/kg 20 µg/day × 14 | Tocris Bioscience | [25,39] |
| p-MPPI: 4-(2′-Methoxyphenyl)-1-[2′-[N-(2″-pyridinyl)-p-iodobenzamido]ethyl]piperazine | 5HT1A receptor antagonist | i.p. | 3 mg/kg | Tocris Bioscience | [25] |
| DOI: 1-(2,5-dimethoxy-4-iodopheny)-2-aminopropane | 5HT2A receptor agonist | i.p. | 0.8 or 1.4 mg/kg | Sigma-Aldrich | [40] |
| Ritanserin: 6-[2-[4-[Bis(4-fluorophenyl)methylene]-1-piperidinyl]ethyl]-7-methyl-5H-thiazolo[3,2-a]pyrimidin-5-one | 5HT2A receptor antagonist | i.p. | 1.5 mg/kg | Tocris Bioscience | [41] |
| Tracer | Location | Administration Side |
|---|---|---|
| NTsami | ||
| 2% FG | Quadriceps Femoris or Quadratus Lumborum | Unilateral (NT side; 1 mm3 Gelfoam × 4/muscle) |
| DiI crystal + 2% WGA (10 µL) | T11-T12 intercostal muscles | Unilateral (NT side) |
| NTsami/NTdmi/SCImi | ||
| 10% BDA (2 µL × 10) | Primary motor cortex | Contralateral to NT side |
| 2% WGA (10 µL) | T8-T9 intercostal muscles | Unilateral (NT side) |
| DiI crystal + 2% FG (10 µL) | T11-T12 intercostal muscles | Unilateral (NT side) |
| 2% FG + 2% WGA (10 µL) | Spinodeltoideus (SPD) and acromiotrapezius (ACZ) | Unilateral (NT side) |
| DiI crystal + 2% WGA (10 µL) | Quadriceps Femoris | Unilateral (NT side) |
| 2% FG (10 µL) | L3 lumbar cord | Bilateral (lateral white matter) |
| NTdmi/NTdmo/SCImi | ||
| DiI crystal + 2% WGA (10 µL) | Quadriceps Femoris | Unilateral (NT side) |
| 2% FG (10 µL) | L3 lumbar cord | Bilateral (lateral white matter) |
| Name | Host | Vender | Dilution |
|---|---|---|---|
| Primary antibody | |||
| Donor hMSC: | |||
| STRO-1 | Mouse | Santa Cruz Bio. | 1:400–500 |
| Neurotransmission: | |||
| vGluT1 | Mouse | MilliporeSigma | 1:200 |
| 5HT | Rabbit | Bio-Rad Laboratories | 1:10,000 |
| 5HT | Rabbit | Immunostar | 1:2000 |
| Neuron: | |||
| Neurofilament H | Rabbit | Chemicon | 1:1000 |
| Neuromuscular junction: | |||
| α-bungarotoxin | Molecular Probes | 1:200 | |
| Secondary antibody | |||
| Alexa Fluor® 488 Donkey | Jackson ImmunoRes. | 1:400–600 | |
| Alexa Fluor® 647 Donkey Anti-Rabbit IgG (H+L) | |||
| Anti-Mouse IgG (H+L) | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, D.; Zeng, X.; Aljuboori, Z.S.; Dennison, R.; Wu, L.; Anderson, J.A.; Teng, Y.D. T12-L3 Nerve Transfer-Induced Locomotor Recovery in Rats with Thoracolumbar Contusion: Essential Roles of Sensory Input Rerouting and Central Neuroplasticity. Cells 2023, 12, 2804. https://doi.org/10.3390/cells12242804
Yu D, Zeng X, Aljuboori ZS, Dennison R, Wu L, Anderson JA, Teng YD. T12-L3 Nerve Transfer-Induced Locomotor Recovery in Rats with Thoracolumbar Contusion: Essential Roles of Sensory Input Rerouting and Central Neuroplasticity. Cells. 2023; 12(24):2804. https://doi.org/10.3390/cells12242804
Chicago/Turabian StyleYu, Dou, Xiang Zeng, Zaid S. Aljuboori, Rachel Dennison, Liquan Wu, Jamie A. Anderson, and Yang D. Teng. 2023. "T12-L3 Nerve Transfer-Induced Locomotor Recovery in Rats with Thoracolumbar Contusion: Essential Roles of Sensory Input Rerouting and Central Neuroplasticity" Cells 12, no. 24: 2804. https://doi.org/10.3390/cells12242804
APA StyleYu, D., Zeng, X., Aljuboori, Z. S., Dennison, R., Wu, L., Anderson, J. A., & Teng, Y. D. (2023). T12-L3 Nerve Transfer-Induced Locomotor Recovery in Rats with Thoracolumbar Contusion: Essential Roles of Sensory Input Rerouting and Central Neuroplasticity. Cells, 12(24), 2804. https://doi.org/10.3390/cells12242804