
Inducing Angiogenesis in the Nucleus Pulposus
 , ,
, ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparing Treatments and Aminal Surgeries
2.2. Noninvasive Induced Angular Displacement Assessments
2.3. Bone Formation and Fusion Assessments
2.4. Micro-CT/Histology and Biomechanics Assessments
2.5. NP Cell and Disc Organ Preparation
2.6. Western Blotting Experiments
2.7. Statistical Testing and Analysis
3. Results
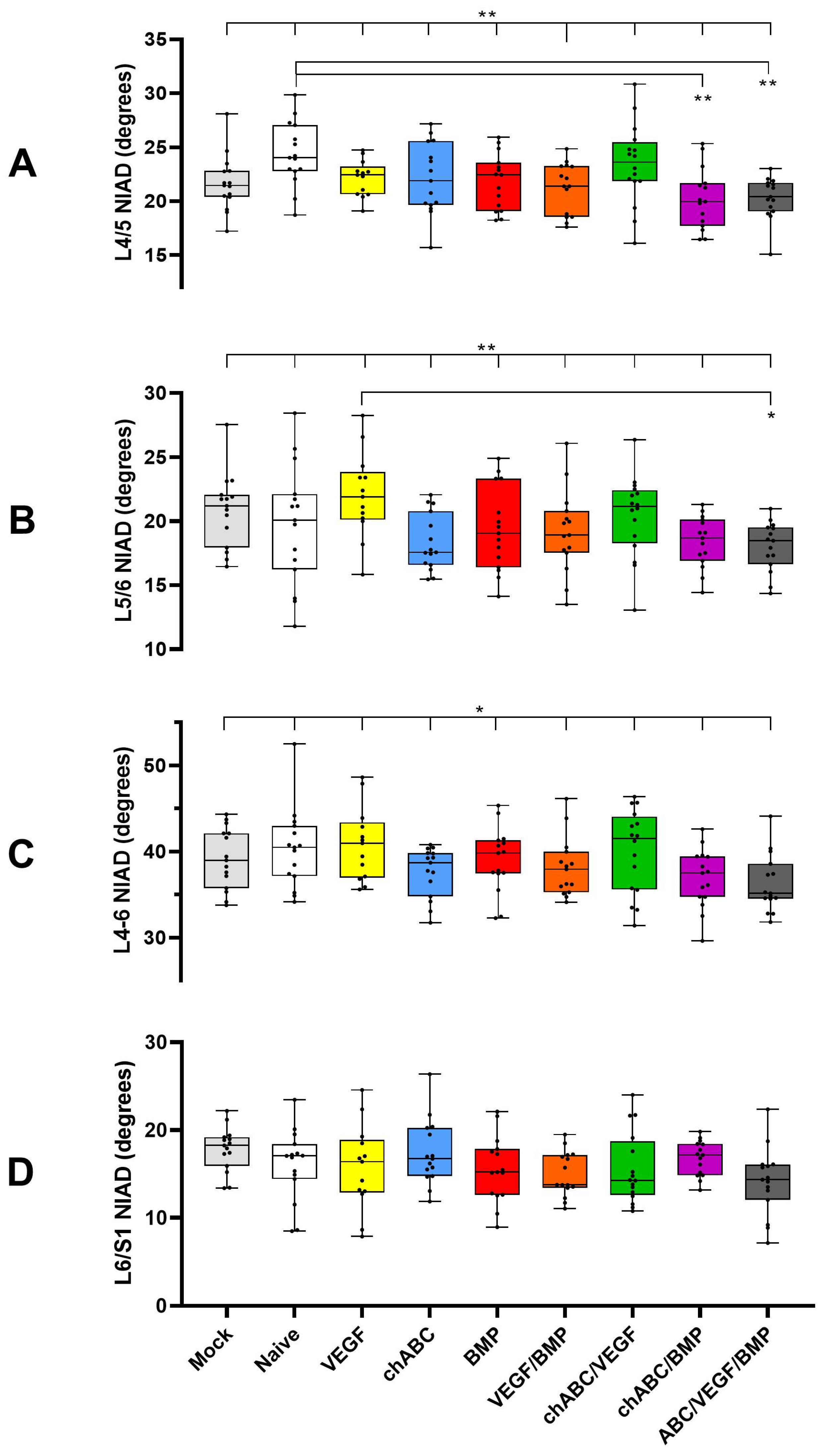
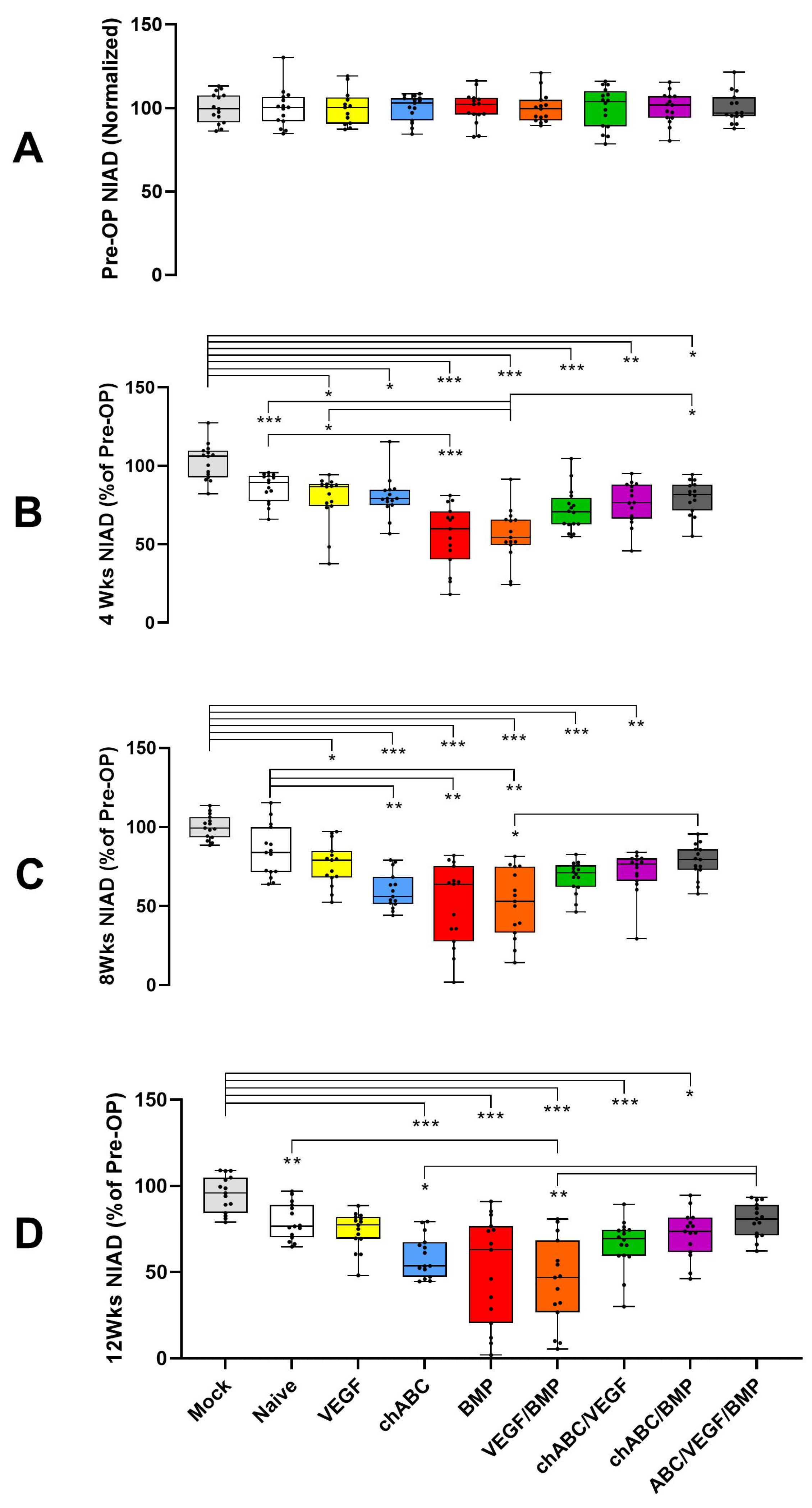
3.1. BMP and BMP/VEGF Decreased NIAD over a 12-Week Time Course
3.2. Mechanical Stiffness Testing Parallels NIAD Findings and Independently Confirms BMP- and BMP/VEGF-Dependent Increased Spinal Stiffness
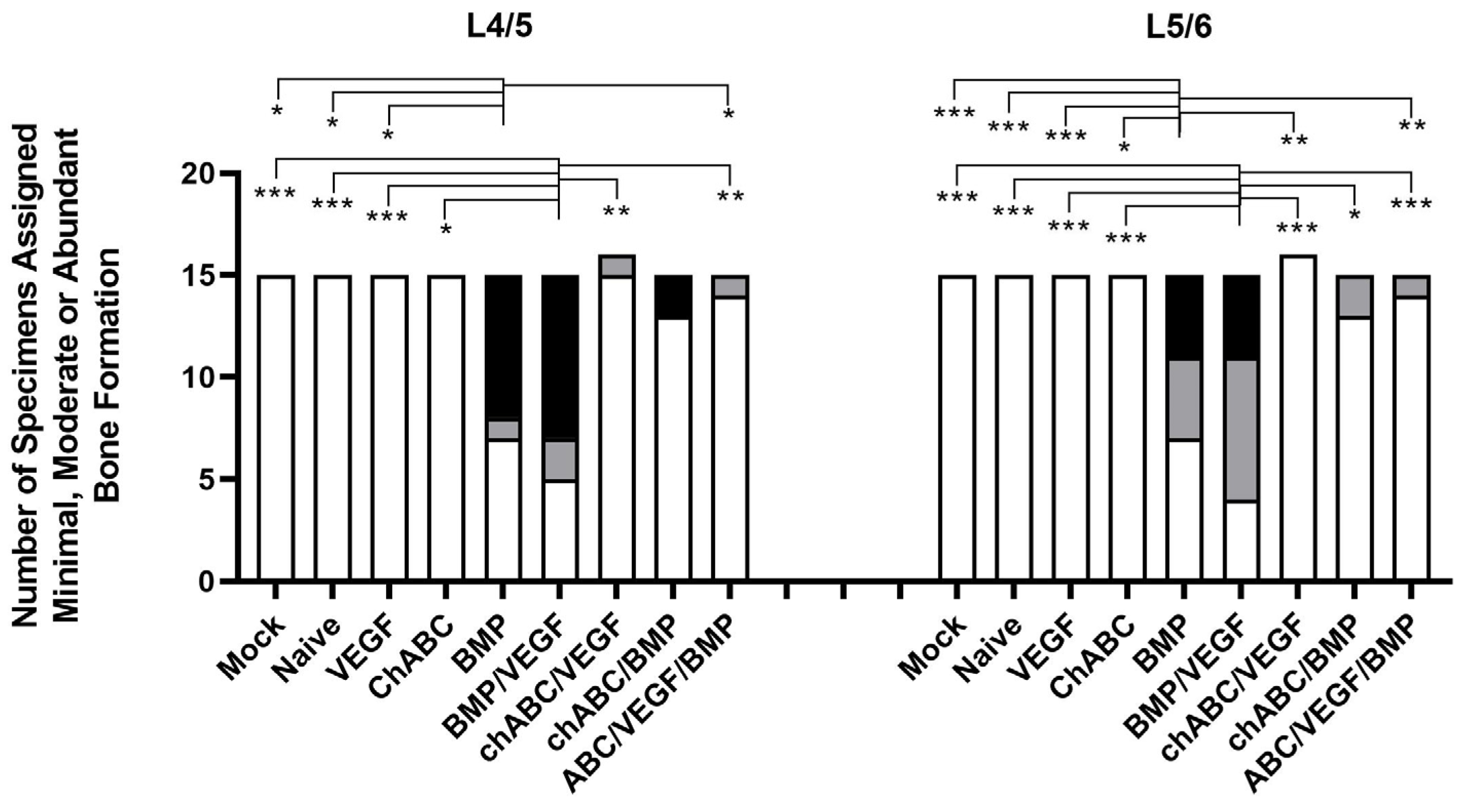
3.3. BMP and BMP/VEGF Treatment Resulted in Increased Frequency of Spinal Fusion Assessed at 12 Weeks
3.4. Decreased Mobility and Increased Spinal Fusion Was the Result of Bone Formation around the L4/5 and L5/6 IVDs, but Not Bone Production Inside the IVDs
3.5. Bone Induction Required BMP Treatment, VEGF Treatment Alone Did Not Induce Angiogenesis, and Chondroitinase ABC Did More Damage Than Expected
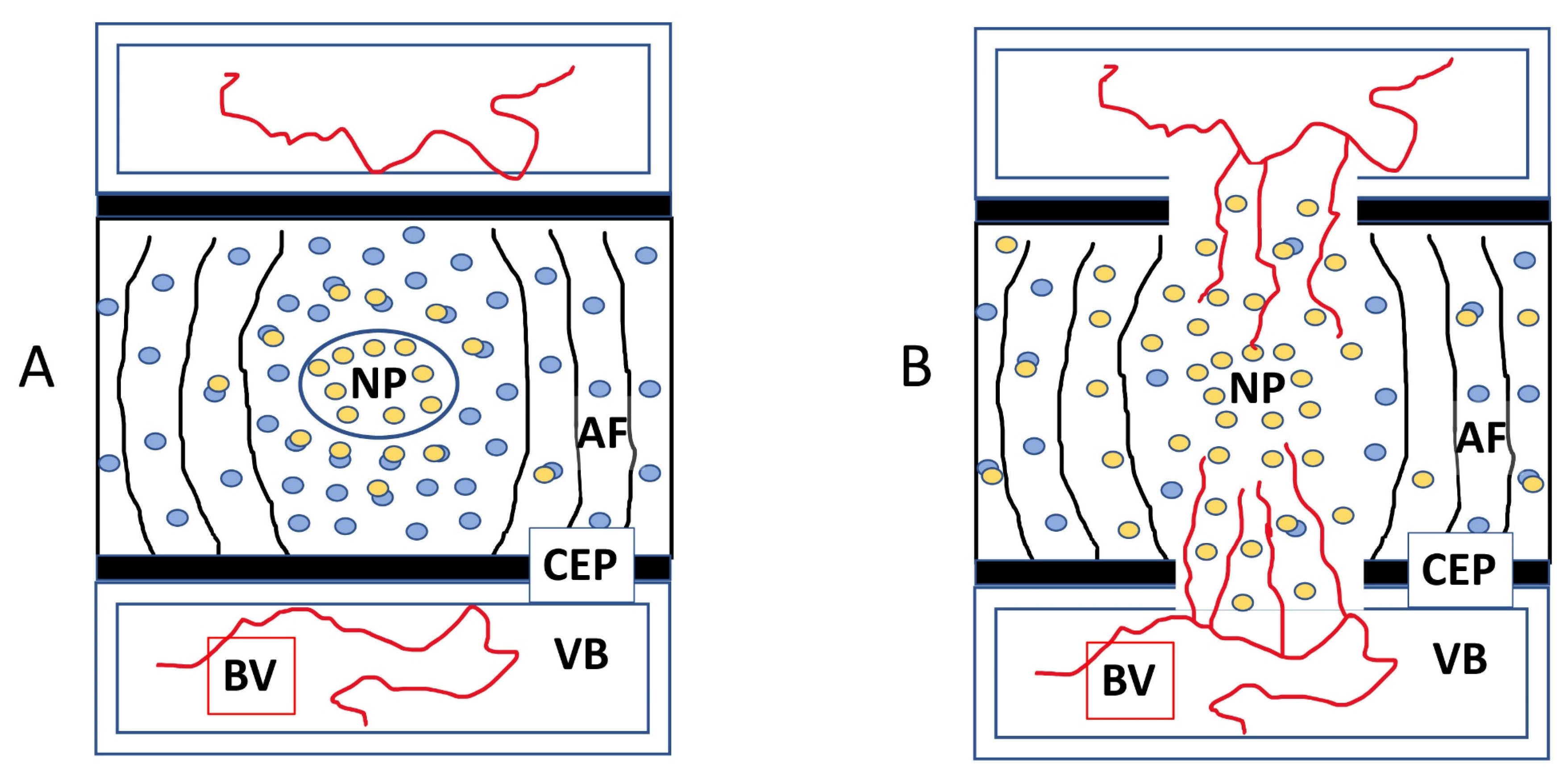
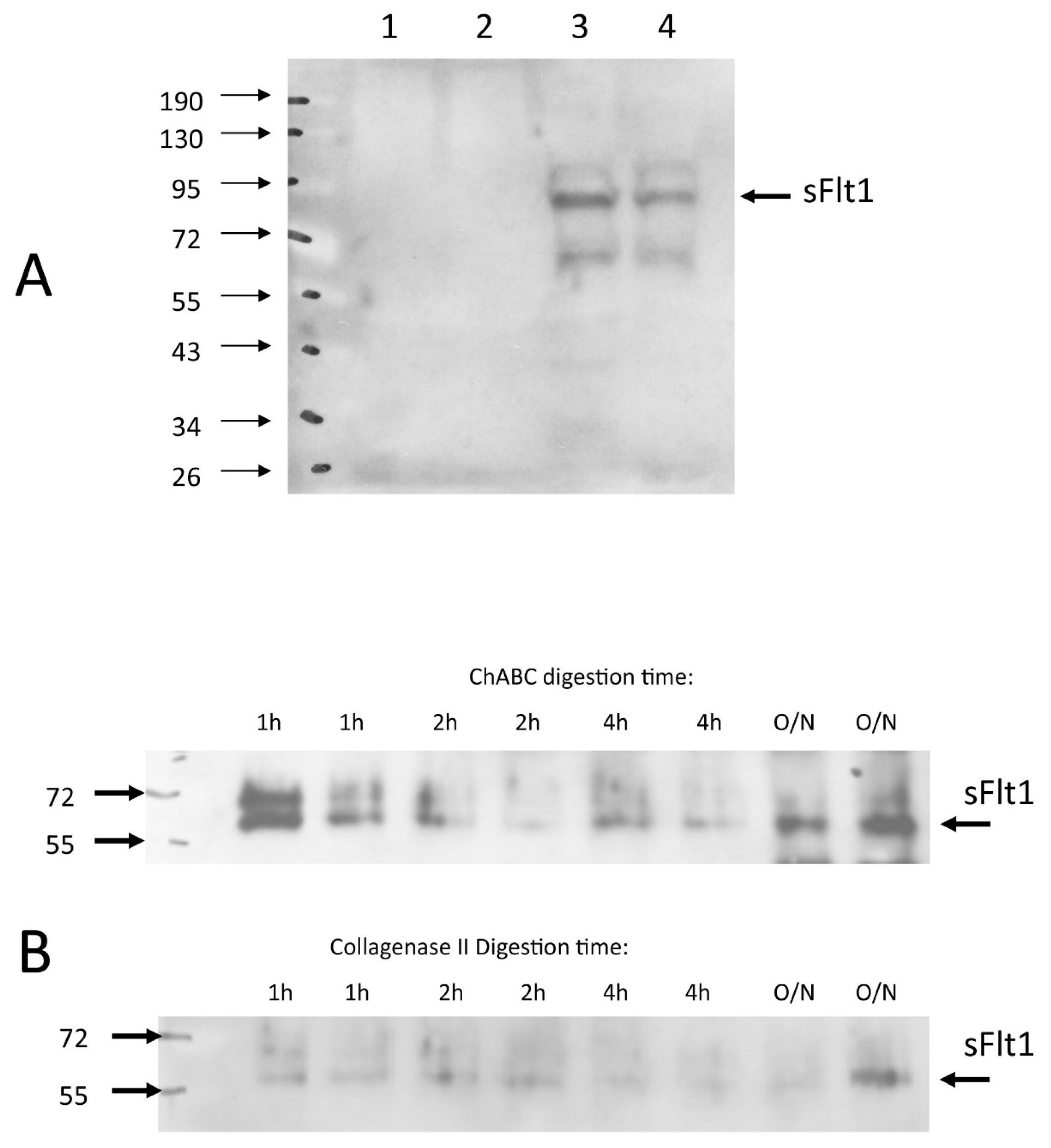
3.6. Chondroitinase ABC Disrupts Endogenously Expressed VEGF Sequestration, Making the IVD Permissive to Vascular Invasion
4. Discussion
5. Limitations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Score | Features | Category | Score | Features |
|---|---|---|---|---|---|
| NP Damage | 0 | Physaliferous cells(+), diffuse/nonfibrotic matrix | Induced Bone Extent | 0 | No Induced Bone (IB) |
| 1 | PF(-), NP now smaller rounded cells, matrix fibrous stranding (PSR(+) staining) not prominent | 1 | Sparse IB ventral to AF, ± connection to VB (EP-OP/ALL calcification/ossification) | ||
| 2 | PF(-), small rounded cells, prominent PSR(+) matrix | 2 | Thick IB with organized bone marrow compartment, ventral to AF, but not fused (partially spanning OP) | ||
| AF Damage | 0 | Organized lamellae, no clefts | |||
| 1 | Wavy/bulging lamellae, mild clefts in outer AF | 3 | IB spanning entire IVD and connecting VBs (IVD fusion) | ||
| 2 | Severe lamellae disorganization/bulging, severe clefts | NP Blood Vessel Structure or Staining Observed | 0 | Structures resembling blood vessels ± RBCs, or anti-VEC/anti-CD31+ immunostaining in only non-induced tissues: VBs, EPs, ALL, PLL, or perivertebral muscle. | |
| IVD Interface Damage | 0 | Normal NP/EP & AF/NP | |||
| 1 | Loss of NP/EP interface | ||||
| 2 | Loss of AF/NP interface | ||||
| IVD PG Damage | 0 | Normal Alcian Blue staining | 1 | Collagen-containing (PSR+) structures resembling blood vessels/RBCs, and or (+) vascular immunostaining cells seen in induced tissues (in VBs or EPs but not into the NP | |
| 1 | Reduced AB staining (>50% of control) | ||||
| 2 | Faint blue to no AB staining (<50% of control) | ||||
| 2 | Blood vessels/vascular immunostain observed crossing into the NP |




| Modality | p-Values for Comparisons | |||||||
|---|---|---|---|---|---|---|---|---|
| L4/5 Segment | L5/6 Segment | L4–6 Combined | ||||||
| Control | BMP | Control | BMP | Control | BMP | |||
| NIAD | 0.95 | 0.58 | 0.885 | 0.722 | 0.724 | 0.58 | ||
| Fusion Assessments | ||||||||
| Palpation | 1 | 0.285 | 1 | 0.458 | # | # | ||
| XR-Graded | 1 | 0.473 | 1 | 0.433 | # | # | ||
| XR-Categorical | 1 | 0.066 | 1 | 1 | # | # | ||
| Osteogenesis Quantifications | ||||||||
| XR-axitron | 1 | 0.019 * | 0.317 | 0.567 | 0.317 | 0.058 | ||
| Micro-CT | # | # | # | # | 0.806 | 0.327 | ||
| Stiffness Measurements were for the L4–6 Combined Segment | ||||||||
| Stiffness Assessments | IVAD | Four-Point Bending | ||||||
| Direction | Control | BMP | Control | BMP | ||||
| Extension | 0.001 * | 0.622 | 0.88 | 0.291 | ||||
| Flexion | 0.023 * | 0.622 | 0.174 | 0.159 | ||||
| Left | 0.001 * | 0.364 | 0.131 | 0.65 | ||||
| Right | 0.131 | 0.291 | 0.112 | 0.481 | ||||
| Coronal | 0.002 * | 0.496 | # | # | ||||
| Sagittal | 0.001 * | 0.573 | # | # | ||||


References
- Kuo, C.C.; Soliman, M.A.R.; Aguirre, A.O.; Youngs, D.; Kruk, M.; Hess, R.M.; Nyabuto, E.M.; Khan, A.; Jowdy, P.K.; Pollina, J.; et al. Risk factors of early complications after thoracic and lumbar spinal deformity surgery: A systematic review and meta-analysis. Eur. Spine J. 2023, 32, 899–913. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.S.; Klineberg, E.; Lafage, V.; Shaffrey, C.I.; Schwab, F.; Lafage, R.; Hostin, R.; Mundis, G.M., Jr.; Errico, T.J.; Kim, H.J.; et al. Prospective multicenter assessment of perioperative and minimum 2-year postoperative complication rates associated with adult spinal deformity surgery. J. Neurosurg. Spine 2016, 25, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Alhammoud, A.; Alborno, Y.; Baco, A.M.; Othman, Y.A.; Ogura, Y.; Steinhaus, M.; Sheha, E.D.; Qureshi, S.A. Minimally Invasive Scoliosis Surgery Is a Feasible Option for Management of Idiopathic Scoliosis and Has Equivalent Outcomes to Open Surgery: A Meta-Analysis. Glob. Spine J. 2022, 12, 483–492. [Google Scholar] [CrossRef]
- Skovrlj, B.; Belton, P.; Zarzour, H.; Qureshi, S.A. Perioperative outcomes in minimally invasive lumbar spine surgery: A systematic review. World J. Orthop. 2015, 6, 996–1005. [Google Scholar] [CrossRef] [PubMed]
- Alden, T.D.; Pittman, D.D.; Beres, E.J.; Hankins, G.R.; Kallmes, D.F.; Wisotsky, B.M.; Kerns, K.M.; Helm, G.A. Percutaneous spinal fusion using bone morphogenetic protein-2 gene therapy. J. Neurosurg. 1999, 90, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Laurent, J.J.; Webb, K.M.; Beres, E.J.; McGee, K.; Li, J.; van Rietbergen, B.; Helm, G.A. The use of bone morphogenetic protein-6 gene therapy for percutaneous spinal fusion in rabbits. J. Neurosurg. Spine 2004, 1, 90–94. [Google Scholar] [CrossRef]
- Cunningham, M.E.; Kelly, N.H.; Rawlins, B.A.; Boachie-Adjei, O.; van der Meulen, M.C.H.; Hidaka, C. Lumbar spine intervertebral disc gene delivery of BMPs induces anterior spine fusion in lewis rats. Sci. Rep. 2022, 12, 16847. [Google Scholar] [CrossRef]
- Kato, F.; Iwata, H.; Murakami, H.; Mimatsu, K.; Miura, T. Chemonucleolysis and response of postchemonucleolytic intervertebral disc to bone morphogenetic protein (BMP). Rinsho Seikeigeka 1987, 22, 965. [Google Scholar]
- Kato, F. Experimental study of chemical spinal fusion in the rabbit by means of bone morphogenetic protein. Nihon Seikeigeka Gakkai Zasshi 1990, 64, 442–452. [Google Scholar]
- Muschik, M.; Schlenzka, D.; Ritsila, V.; Tennstedt, C.; Lewandrowski, K.U. Experimental anterior spine fusion using bovine bone morphogenetic protein: A study in rabbits. J. Orthop. Sci. 2000, 5, 165–170. [Google Scholar] [CrossRef]
- Nordby, E.J.; Wright, P.H.; Schofield, S.R. Safety of chemonucleolysis. Adverse effects reported in the United States, 1982–1991. Clin. Orthop. Relat. Res. 1993, 293, 122–134. [Google Scholar] [CrossRef]
- Herkowitz, H.N. Current status of percutaneous discectomy and chemonucleolysis. Orthop. Clin. N. Am. 1991, 22, 327–332. [Google Scholar] [CrossRef]
- Kato, F.; Mimatsu, K.; Iwata, H.; Miura, T. Comparison of tissue reaction with chondroitinase ABC and chymopapain in rabbits as the basis of clinical application in chemonucleolysis. Clin. Orthop. Relat. Res. 1993, 294–302. [Google Scholar] [CrossRef]
- Olmarker, K.; Danielsen, N.; Nannmark, U.; Sennerby, L.; Rydevik, B. Microvascular effects of chondroitinase ABC and chymopapain. An in vivo experimental study on hamsters and rabbits. Clin. Orthop. Relat. Res. 1990, 257, 274–279. [Google Scholar] [CrossRef]
- Park, J.S.; Ahn, J.I. The effect of chondroitinase ABC on rabbit intervertebral disc. Radiological, histological and electron microscopic findings. Int. Orthop. 1995, 19, 103–109. [Google Scholar] [CrossRef]
- Olmarker, K.; Stromberg, J.; Blomquist, J.; Zachrisson, P.; Nannmark, U.; Nordborg, C.; Rydevik, B. Chondroitinase ABC (pharmaceutical grade) for chemonucleolysis. Functional and structural evaluation after local application on intraspinal nerve structures and blood vessels. Spine 1996, 21, 1952–1956. [Google Scholar] [CrossRef]
- Boxberger, J.I.; Sen, S.; Yerramalli, C.S.; Elliott, D.M. Nucleus pulposus glycosaminoglycan content is correlated with axial mechanics in rat lumbar motion segments. J. Orthop. Res. 2006, 24, 1906–1915. [Google Scholar] [CrossRef]
- Boxberger, J.I.; Auerbach, J.D.; Sen, S.; Elliott, D.M. An in vivo model of reduced nucleus pulposus glycosaminoglycan content in the rat lumbar intervertebral disc. Spine 2008, 33, 146–154. [Google Scholar] [CrossRef]
- He, M.; Pang, J.; Sun, H.; Zheng, G.; Lin, Y.; Ge, W. Overexpression of TIMP3 inhibits discogenic pain by suppressing angiogenesis and the expression of substance P in nucleus pulposus. Mol. Med. Rep. 2020, 21, 1163–1171. [Google Scholar] [CrossRef]
- Tolofari, S.K.; Richardson, S.M.; Freemont, A.J.; Hoyland, J.A. Expression of semaphorin 3A and its receptors in the human intervertebral disc: Potential role in regulating neural ingrowth in the degenerate intervertebral disc. Arthritis Res. Ther. 2010, 12, R1. [Google Scholar] [CrossRef]
- Iruela-Arispe, M.L.; Liska, D.J.; Sage, E.H.; Bornstein, P. Differential expression of thrombospondin 1, 2, and 3 during murine development. Dev. Dyn. 1993, 197, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Ishii, Y.; Thomas, A.O.; Guo, X.E.; Hung, C.T.; Chen, F.H. Localization and distribution of cartilage oligomeric matrix protein in the rat intervertebral disc. Spine 2006, 31, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Kusafuka, K.; Hiraki, Y.; Shukunami, C.; Kayano, T.; Takemura, T. Cartilage-specific matrix protein, chondromodulin-I (ChM-I), is a strong angio-inhibitor in endochondral ossification of human neonatal vertebral tissues in vivo: Relationship with angiogenic factors in the cartilage. Acta Histochem. 2002, 104, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Minogue, B.M.; Richardson, S.M.; Zeef, L.A.; Freemont, A.J.; Hoyland, J.A. Transcriptional profiling of bovine intervertebral disc cells: Implications for identification of normal and degenerate human intervertebral disc cell phenotypes. Arthritis Res. Ther. 2010, 12, R22. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, T.; Tian, W.; Hu, H.; Xin, Z.; Ma, X.; Ye, C.; Hang, K.; Han, X.; Zhao, J.; et al. Loss of TIMP3 expression induces inflammation, matrix degradation, and vascular ingrowth in nucleus pulposus: A new mechanism of intervertebral disc degeneration. FASEB J. 2020, 34, 5483–5498. [Google Scholar] [CrossRef]
- Lin, D.; Alberton, P.; Delgado Caceres, M.; Prein, C.; Clausen-Schaumann, H.; Dong, J.; Aszodi, A.; Shukunami, C.; Iatridis, J.C.; Docheva, D. Loss of tenomodulin expression is a risk factor for age-related intervertebral disc degeneration. Aging Cell 2020, 19, e13091. [Google Scholar] [CrossRef]
- Fournier, D.E.; Kiser, P.K.; Shoemaker, J.K.; Battie, M.C.; Seguin, C.A. Vascularization of the human intervertebral disc: A scoping review. JOR Spine 2020, 3, e1123. [Google Scholar] [CrossRef]
- Kim, J.H.; Ham, C.H.; Kwon, W.K. Current Knowledge and Future Therapeutic Prospects in Symptomatic Intervertebral Disc Degeneration. Yonsei Med. J. 2022, 63, 199–210. [Google Scholar] [CrossRef]
- Binch, A.L.; Cole, A.A.; Breakwell, L.M.; Michael, A.L.; Chiverton, N.; Cross, A.K.; Le Maitre, C.L. Expression and regulation of neurotrophic and angiogenic factors during human intervertebral disc degeneration. Arthritis Res. Ther. 2014, 16, 416. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, J.; Xia, Q.; Wu, L.; Chen, F.; Li, L.; Zhu, C.; He, M.; Jiang, Y.; Huang, Y.; et al. Treatment outcomes of injectable thermosensitive hydrogel containing bevacizumab in intervertebral disc degeneration. Front. Bioeng. Biotechnol. 2022, 10, 976706. [Google Scholar] [CrossRef]
- Fujita, N.; Imai, J.; Suzuki, T.; Yamada, M.; Ninomiya, K.; Miyamoto, K.; Iwasaki, R.; Morioka, H.; Matsumoto, M.; Chiba, K.; et al. Vascular endothelial growth factor-A is a survival factor for nucleus pulposus cells in the intervertebral disc. Biochem. Biophys. Res. Commun. 2008, 372, 367–372. [Google Scholar] [CrossRef]
- Rodrigues-Pinto, R.; Ward, L.; Humphreys, M.; Zeef, L.A.H.; Berry, A.; Hanley, K.P.; Hanley, N.; Richardson, S.M.; Hoyland, J.A. Human notochordal cell transcriptome unveils potential regulators of cell function in the developing intervertebral disc. Sci. Rep. 2018, 8, 12866. [Google Scholar] [CrossRef]
- Sakai, D.; Nakai, T.; Mochida, J.; Alini, M.; Grad, S. Differential phenotype of intervertebral disc cells: Microarray and immunohistochemical analysis of canine nucleus pulposus and anulus fibrosus. Spine 2009, 34, 1448–1456. [Google Scholar] [CrossRef]
- Agrawal, A.; Gajghate, S.; Smith, H.; Anderson, D.G.; Albert, T.J.; Shapiro, I.M.; Risbud, M.V. Cited2 modulates hypoxia-inducible factor-dependent expression of vascular endothelial growth factor in nucleus pulposus cells of the rat intervertebral disc. Arthritis Rheum. 2008, 58, 3798–3808. [Google Scholar] [CrossRef] [PubMed]
- Kwon, W.K.; Moon, H.J.; Kwon, T.H.; Park, Y.K.; Kim, J.H. Influence of rabbit notochordal cells on symptomatic intervertebral disc degeneration: Anti-angiogenic capacity on human endothelial cell proliferation under hypoxia. Osteoarthr. Cartil. 2017, 25, 1738–1746. [Google Scholar] [CrossRef]
- Merceron, C.; Mangiavini, L.; Robling, A.; Wilson, T.L.; Giaccia, A.J.; Shapiro, I.M.; Schipani, E.; Risbud, M.V. Loss of HIF-1alpha in the notochord results in cell death and complete disappearance of the nucleus pulposus. PLoS ONE 2014, 9, e110768. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.J.; Zhang, X.K.; Zheng, X.F.; Yang, Y.H.; Jiang, S.D.; Jiang, L.S. SHH-dependent knockout of HIF-1 alpha accelerates the degenerative process in mouse intervertebral disc. Int. J. Immunopathol. Pharmacol. 2013, 26, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Zhuang, L.; Wang, J.; Liu, Z.; Wang, Y.; Xiao, D.; Zhang, X. Hypoxia-inducible factor (HIF)-1alpha knockout accelerates intervertebral disc degeneration in mice. Int. J. Clin. Exp. Pathol. 2018, 11, 548–557. [Google Scholar]
- Bouletreau, P.J.; Warren, S.M.; Spector, J.A.; Peled, Z.M.; Gerrets, R.P.; Greenwald, J.A.; Longaker, M.T. Hypoxia and VEGF up-regulate BMP-2 mRNA and protein expression in microvascular endothelial cells: Implications for fracture healing. Plast. Reconstr. Surg. 2002, 109, 2384–2397. [Google Scholar] [CrossRef]
- Dai, J.; Kitagawa, Y.; Zhang, J.; Yao, Z.; Mizokami, A.; Cheng, S.; Nor, J.; McCauley, L.K.; Taichman, R.S.; Keller, E.T. Vascular endothelial growth factor contributes to the prostate cancer-induced osteoblast differentiation mediated by bone morphogenetic protein. Cancer Res. 2004, 64, 994–999. [Google Scholar] [CrossRef]
- Li, G.; Cui, Y.; McIlmurray, L.; Allen, W.E.; Wang, H. rhBMP-2, rhVEGF(165), rhPTN and thrombin-related peptide, TP508 induce chemotaxis of human osteoblasts and microvascular endothelial cells. J. Orthop. Res. 2005, 23, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Wright, V.; Usas, A.; Gearhart, B.; Shen, H.C.; Cummins, J.; Huard, J. Synergistic enhancement of bone formation and healing by stem cell-expressed VEGF and bone morphogenetic protein-4. J. Clin. Investig. 2002, 110, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Usas, A.; Olshanski, A.; Ho, A.M.; Gearhart, B.; Cooper, G.M.; Huard, J. VEGF improves, whereas sFlt1 inhibits, BMP2-induced bone formation and bone healing through modulation of angiogenesis. J. Bone Miner. Res. 2005, 20, 2017–2027. [Google Scholar] [CrossRef] [PubMed]
- Damle, S.R.; Rawlins, B.A.; Boachie-Adjei, O.; Crystal, R.G.; Hidaka, C.; Cunningham, M.E. Lumbar spine intervertebral disc gene delivery: A pilot study in lewis rats. HSS J. 2013, 9, 36–41. [Google Scholar] [CrossRef]
- Damle, S.R.; Krzyzanowska, A.; Frawley, R.J.; Cunningham, M.E. Surgical anatomy, transperitoneal approach, and early postoperative complications of a ventral lumbar spine surgical model in Lewis rats. Comp. Med. 2013, 63, 409–415. [Google Scholar]
- Cunningham, M.E.; Beach, J.M.; Bilgic, S.; Boachie-Adjei, O.; van der Meulen, M.C.; Hidaka, C. In vivo and in vitro analysis of rat lumbar spine mechanics. Clin. Orthop. Relat. Res. 2010, 468, 2695–2703. [Google Scholar] [CrossRef]
- Bae, H.W.; Zhao, L.; Kanim, L.E.; Wong, P.; Marshall, D.; Delamarter, R.B. Bone marrow enhances the performance of rhBMP-2 in spinal fusion: A rodent model. J. Bone Jt. Surg. Am. 2013, 95, 338–347. [Google Scholar] [CrossRef]
- Peterson, B.; Whang, P.G.; Iglesias, R.; Wang, J.C.; Lieberman, J.R. Osteoinductivity of commercially available demineralized bone matrix. Preparations in a spine fusion model. J. Bone Jt. Surg. Am. 2004, 86, 2243–2250. [Google Scholar] [CrossRef]
- Melgoza, I.P.; Chenna, S.S.; Tessier, S.; Zhang, Y.; Tang, S.Y.; Ohnishi, T.; Novais, E.J.; Kerr, G.J.; Mohanty, S.; Tam, V.; et al. Development of a standardized histopathology scoring system using machine learning algorithms for intervertebral disc degeneration in the mouse model-An ORS spine section initiative. JOR Spine 2021, 4, e1164. [Google Scholar] [CrossRef]
- Mohd Isa, I.L.; Abbah, S.A.; Kilcoyne, M.; Sakai, D.; Dockery, P.; Finn, D.P.; Pandit, A. Implantation of hyaluronic acid hydrogel prevents the pain phenotype in a rat model of intervertebral disc injury. Sci. Adv. 2018, 4, eaaq0597. [Google Scholar] [CrossRef]
- Yaniv, G.; Bader, S.; Lidar, M.; Herman, A.; Shazar, N.; Aharoni, D.; Eshed, I. The natural course of bridging osteophyte formation in diffuse idiopathic skeletal hyperostosis: Retrospective analysis of consecutive CT examinations over 10 years. Rheumatology 2014, 53, 1951–1957. [Google Scholar] [CrossRef] [PubMed]
- Krzyzanowska, A.K.; Frawley, R.J.; Damle, S.; Chen, T.; Otero, M.; Cunningham, M.E. Activation of nuclear factor-kappa B by TNF promotes nucleus pulposus mineralization through inhibition of ANKH and ENPP1. Sci. Rep. 2021, 11, 8271. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, M.E.; Greene, L.A. A function-structure model for NGF-activated TRK. EMBO J. 1998, 17, 7282–7293. [Google Scholar] [CrossRef]
- Cunningham, M.E.; Stephens, R.M.; Kaplan, D.R.; Greene, L.A. Autophosphorylation of activation loop tyrosines regulates signaling by the TRK nerve growth factor receptor. J. Biol. Chem. 1997, 272, 10957–10967. [Google Scholar] [CrossRef] [PubMed]
- Koo, T.K.; Li, M.Y. A Guideline of Selecting and Reporting Intraclass Correlation Coefficients for Reliability Research. J. Chiropr. Med. 2016, 15, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Landis, J.R.; Koch, G.G. The measurement of observer agreement for categorical data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef]
- Kim, J.W.; Jeon, N.; Shin, D.E.; Lee, S.Y.; Kim, M.; Han, D.H.; Shin, J.Y.; Lee, S. Regeneration in Spinal Disease: Therapeutic Role of Hypoxia-Inducible Factor-1 Alpha in Regeneration of Degenerative Intervertebral Disc. Int. J. Mol. Sci. 2021, 22, 5281. [Google Scholar] [CrossRef]
- Risbud, M.V.; Schipani, E.; Shapiro, I.M. Hypoxic regulation of nucleus pulposus cell survival: From niche to notch. Am. J. Pathol. 2010, 176, 1577–1583. [Google Scholar] [CrossRef]
- Ambati, B.K.; Nozaki, M.; Singh, N.; Takeda, A.; Jani, P.D.; Suthar, T.; Albuquerque, R.J.; Richter, E.; Sakurai, E.; Newcomb, M.T.; et al. Corneal avascularity is due to soluble VEGF receptor-1. Nature 2006, 443, 993–997. [Google Scholar] [CrossRef]
- Capossela, S.; Bertolo, A.; Gunasekera, K.; Potzel, T.; Baur, M.; Stoyanov, J.V. VEGF vascularization pathway in human intervertebral disc does not change during the disc degeneration process. BMC Res. Notes 2018, 11, 333. [Google Scholar] [CrossRef]
- Carreon, L.Y.; Ito, T.; Yamada, M.; Uchiyama, S.; Takahashi, H.; Ikuta, F. Histologic changes in the disc after cervical spine trauma: Evidence of disc absorption. J. Spinal Disord. 1996, 9, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Foulsham, W.; Dohlman, T.H.; Mittal, S.K.; Taketani, Y.; Singh, R.B.; Masli, S.; Dana, R. Thrombospondin-1 in ocular surface health and disease. Ocul. Surf. 2019, 17, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Cursiefen, C.; Masli, S.; Ng, T.F.; Dana, M.R.; Bornstein, P.; Lawler, J.; Streilein, J.W. Roles of thrombospondin-1 and -2 in regulating corneal and iris angiogenesis. Investig. Ophthalmol. Vis. Sci. 2004, 45, 1117–1124. [Google Scholar] [CrossRef]
- Tucker, R.P.; Hagios, C.; Chiquet-Ehrismann, R.; Lawler, J. In situ localization of thrombospondin-1 and thrombospondin-3 transcripts in the avian embryo. Dev. Dyn. 1997, 208, 326–337. [Google Scholar] [CrossRef]
- Ebrahem, Q.; Qi, J.H.; Sugimoto, M.; Ali, M.; Sears, J.E.; Cutler, A.; Khokha, R.; Vasanji, A.; Anand-Apte, B. Increased neovascularization in mice lacking tissue inhibitor of metalloproteinases-3. Investig. Ophthalmol. Vis. Sci. 2011, 52, 6117–6123. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.H.; Zhang, F.; Shi, W.; Yao, J.Y.; Hsiao, C.H.; Wu, H.C.; Kim, W.S.; Hao, Y.X.; Hwang, D.G.; Chen, J.K.; et al. Expression of tissue inhibitor of metalloproteinase-4 in normal human corneal cells and experimental corneal neovascularization. Ophthalmic Res. 2003, 35, 199–207. [Google Scholar] [CrossRef]
- Azar, D.T. Corneal angiogenic privilege: Angiogenic and antiangiogenic factors in corneal avascularity, vasculogenesis, and wound healing (an American Ophthalmological Society thesis). Trans. Am. Ophthalmol. Soc. 2006, 104, 264–302. [Google Scholar]
- Barry, Z.; Park, B.; Corson, T.W. Pharmacological Potential of Small Molecules for Treating Corneal Neovascularization. Molecules 2020, 25, 3468. [Google Scholar] [CrossRef]
- Gutman, G.; Barak, V.; Maslovitz, S.; Amit, A.; Lessing, J.B.; Geva, E. Regulation of vascular endothelial growth factor-A and its soluble receptor sFlt-1 by luteinizing hormone in vivo: Implication for ovarian follicle angiogenesis. Fertil. Steril. 2008, 89, 922–926. [Google Scholar] [CrossRef]
- Ku, C.H.; White, K.E.; Dei Cas, A.; Hayward, A.; Webster, Z.; Bilous, R.; Marshall, S.; Viberti, G.; Gnudi, L. Inducible overexpression of sFlt-1 in podocytes ameliorates glomerulopathy in diabetic mice. Diabetes 2008, 57, 2824–2833. [Google Scholar] [CrossRef]
- Mutter, W.P.; Karumanchi, S.A. Molecular mechanisms of preeclampsia. Microvasc. Res. 2008, 75, 1–8. [Google Scholar] [CrossRef]
- Purpura, K.A.; George, S.H.; Dang, S.M.; Choi, K.; Nagy, A.; Zandstra, P.W. Soluble Flt-1 regulates Flk-1 activation to control hematopoietic and endothelial development in an oxygen-responsive manner. Stem Cells 2008, 26, 2832–2842. [Google Scholar] [CrossRef]
- Shibuya, M. Vascular endothelial growth factor and its receptor system: Physiological functions in angiogenesis and pathological roles in various diseases. J. Biochem. 2013, 153, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.; Zhao, X.; Zhou, M.; Deng, Y.; Li, Y.; Peng, C. sFlt-1: A Double Regulator in Angiogenesis-related Diseases. Curr. Pharm. Des. 2021, 27, 4160–4170. [Google Scholar] [CrossRef] [PubMed]
- Russo, J.W.; Castellot, J.J. CCN5: Biology and pathophysiology. J. Cell Commun. Signal. 2010, 4, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Meadows, S.M.; Fletcher, P.J.; Moran, C.; Xu, K.; Neufeld, G.; Chauvet, S.; Mann, F.; Krieg, P.A.; Cleaver, O. Integration of repulsive guidance cues generates avascular zones that shape mammalian blood vessels. Circ. Res. 2012, 110, 34–46. [Google Scholar] [CrossRef]
- Masuda, T.; Shiga, T. Chemorepulsion and cell adhesion molecules in patterning initial trajectories of sensory axons. Neurosci. Res. 2005, 51, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Reese, D.E.; Hall, C.E.; Mikawa, T. Negative regulation of midline vascular development by the notochord. Dev. Cell 2004, 6, 699–708. [Google Scholar] [CrossRef]
- Bressan, M.; Davis, P.; Timmer, J.; Herzlinger, D.; Mikawa, T. Notochord-derived BMP antagonists inhibit endothelial cell generation and network formation. Dev. Biol. 2009, 326, 101–111. [Google Scholar] [CrossRef][Green Version]
- Rattner, A.; Yu, H.; Williams, J.; Smallwood, P.M.; Nathans, J. Endothelin-2 signaling in the neural retina promotes the endothelial tip cell state and inhibits angiogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, E3830–E3839. [Google Scholar] [CrossRef]
- Binch, A.L.; Cole, A.A.; Breakwell, L.M.; Michael, A.L.; Chiverton, N.; Creemers, L.B.; Cross, A.K.; Le Maitre, C.L. Class 3 semaphorins expression and association with innervation and angiogenesis within the degenerate human intervertebral disc. Oncotarget 2015, 6, 18338–18354. [Google Scholar] [CrossRef] [PubMed]
- Mima, Y.; Suzuki, S.; Fujii, T.; Morikawa, T.; Tamaki, S.; Takubo, K.; Shimoda, M.; Miyamoto, T.; Watanabe, K.; Matsumoto, M.; et al. Potential involvement of semaphorin 3A in maintaining intervertebral disc tissue homeostasis. J. Orthop. Res. 2019, 37, 972–980. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.S.; Brochmann Murray, E.J.; Wang, J.C.; Duarte, M.E. The history and histology of bone morphogenetic protein. Histol. Histopathol. 2016, 31, 721–732. [Google Scholar] [PubMed]
- Koosha, E.; Eames, B.F. Two Modulators of Skeletal Development: BMPs and Proteoglycans. J. Dev. Biol. 2022, 10, 15. [Google Scholar] [CrossRef] [PubMed]
- Yokobori, T.; Oohira, A.; Nogami, H. Proteoglycans synthesized in calluses at various stages of fracture healing in rats. Biochim. Biophys. Acta 1980, 628, 174–181. [Google Scholar] [CrossRef]
- Masuda, K.; Imai, Y.; Okuma, M.; Muehleman, C.; Nakagawa, K.; Akeda, K.; Thonar, E.; Andersson, G.; An, H.S. Osteogenic protein-1 injection into a degenerated disc induces the restoration of disc height and structural changes in the rabbit anular puncture model. Spine 2006, 31, 742–754. [Google Scholar] [CrossRef]
- Miyamoto, K.; Masuda, K.; Kim, J.G.; Inoue, N.; Akeda, K.; Andersson, G.B.; An, H.S. Intradiscal injections of osteogenic protein-1 restore the viscoelastic properties of degenerated intervertebral discs. Spine J. 2006, 6, 692–703. [Google Scholar] [CrossRef]
- Patel, S.R.; Lee, L.Y.; Mack, C.A.; Polce, D.R.; El-Sawy, T.; Hackett, N.R.; Ilercil, A.; Jones, E.C.; Hahn, R.T.; Isom, O.W.; et al. Safety of direct myocardial administration of an adenovirus vector encoding vascular endothelial growth factor 121. Hum. Gene Ther. 1999, 10, 1331–1348. [Google Scholar] [CrossRef]
- Kremer, E.J.; Di Falco, N.D.; Opolon, P.; Perricaudet, M. Adenovirus vector-transduced hepatocytes implanted via a preformed collagen/PTFE support persist for at least 4 weeks in vivo. Gene Ther. 1996, 3, 932–936. [Google Scholar]
- Wu, J.C.; Sundaresan, G.; Iyer, M.; Gambhir, S.S. Noninvasive optical imaging of firefly luciferase reporter gene expression in skeletal muscles of living mice. Mol. Ther. 2001, 4, 297–306. [Google Scholar] [CrossRef]
- Hettiaratchi, M.H.; O’Meara, M.J.; O’Meara, T.R.; Pickering, A.J.; Letko-Khait, N.; Shoichet, M.S. Reengineering biocatalysts: Computational redesign of chondroitinase ABC improves efficacy and stability. Sci. Adv. 2020, 6, eabc6378. [Google Scholar] [CrossRef]
- Corvetti, L.; Rossi, F. Degradation of chondroitin sulfate proteoglycans induces sprouting of intact purkinje axons in the cerebellum of the adult rat. J. Neurosci. 2005, 25, 7150–7158. [Google Scholar] [CrossRef] [PubMed]
- Kaito, T.; Johnson, J.; Ellerman, J.; Tian, H.; Aydogan, M.; Chatsrinopkun, M.; Ngo, S.; Choi, C.; Wang, J.C. Synergistic effect of bone morphogenetic proteins 2 and 7 by ex vivo gene therapy in a rat spinal fusion model. J. Bone Jt. Surg. Am. 2013, 95, 1612–1619. [Google Scholar] [CrossRef]
- Zhu, W.; Rawlins, B.A.; Boachie-Adjei, O.; Myers, E.R.; Arimizu, J.; Choi, E.; Lieberman, J.R.; Crystal, R.G.; Hidaka, C. Combined bone morphogenetic protein-2 and -7 gene transfer enhances osteoblastic differentiation and spine fusion in a rodent model. J. Bone Miner. Res. 2004, 19, 2021–2032. [Google Scholar] [CrossRef]
- Ferrara, N. Vascular endothelial growth factor: Basic science and clinical progress. Endocr. Rev. 2004, 25, 581–611. [Google Scholar] [CrossRef] [PubMed]
- Owen-Woods, C.; Kusumbe, A. Fundamentals of bone vasculature: Specialization, interactions and functions. Semin. Cell Dev. Biol. 2022, 123, 36–47. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damle, S.R.; Krzyzanowska, A.K.; Korsun, M.K.; Morse, K.W.; Gilbert, S.; Kim, H.J.; Boachie-Adjei, O.; Rawlins, B.A.; van der Meulen, M.C.H.; Greenblatt, M.B.; et al. Inducing Angiogenesis in the Nucleus Pulposus. Cells 2023, 12, 2488. https://doi.org/10.3390/cells12202488
Damle SR, Krzyzanowska AK, Korsun MK, Morse KW, Gilbert S, Kim HJ, Boachie-Adjei O, Rawlins BA, van der Meulen MCH, Greenblatt MB, et al. Inducing Angiogenesis in the Nucleus Pulposus. Cells. 2023; 12(20):2488. https://doi.org/10.3390/cells12202488
Chicago/Turabian StyleDamle, Sheela R., Agata K. Krzyzanowska, Maximilian K. Korsun, Kyle W. Morse, Susannah Gilbert, Han Jo Kim, Oheneba Boachie-Adjei, Bernard A. Rawlins, Marjolein C. H. van der Meulen, Matthew B. Greenblatt, and et al. 2023. "Inducing Angiogenesis in the Nucleus Pulposus" Cells 12, no. 20: 2488. https://doi.org/10.3390/cells12202488
APA StyleDamle, S. R., Krzyzanowska, A. K., Korsun, M. K., Morse, K. W., Gilbert, S., Kim, H. J., Boachie-Adjei, O., Rawlins, B. A., van der Meulen, M. C. H., Greenblatt, M. B., Hidaka, C., & Cunningham, M. E. (2023). Inducing Angiogenesis in the Nucleus Pulposus. Cells, 12(20), 2488. https://doi.org/10.3390/cells12202488