

New Treatment for Type 2 Diabetes Mellitus Using a Novel Bipyrazole Compound

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Lipinski’s Rule of Five for Drug-Likeness
2.2. Acute Oral Toxicity Test
2.3. Induction of T2D and Experimental Design
2.4. Biochemical Investigations
2.4.1. Measurement of Serum Glucose, Insulin, and Lipids in Rat Serum
2.4.2. Homeostasis Model Assessment of Insulin Resistance (HOMA-IR)
2.4.3. Measurement of Serum Glutathione (GSH), Malondialdehyde (MDA), and IL-6 Serum Concentrations
2.5. Western Blotting
2.6. Statistical Analysis
3. Results
3.1. Lipinski’s Rule and Drug-Likeness
3.2. Acute Toxicity Study
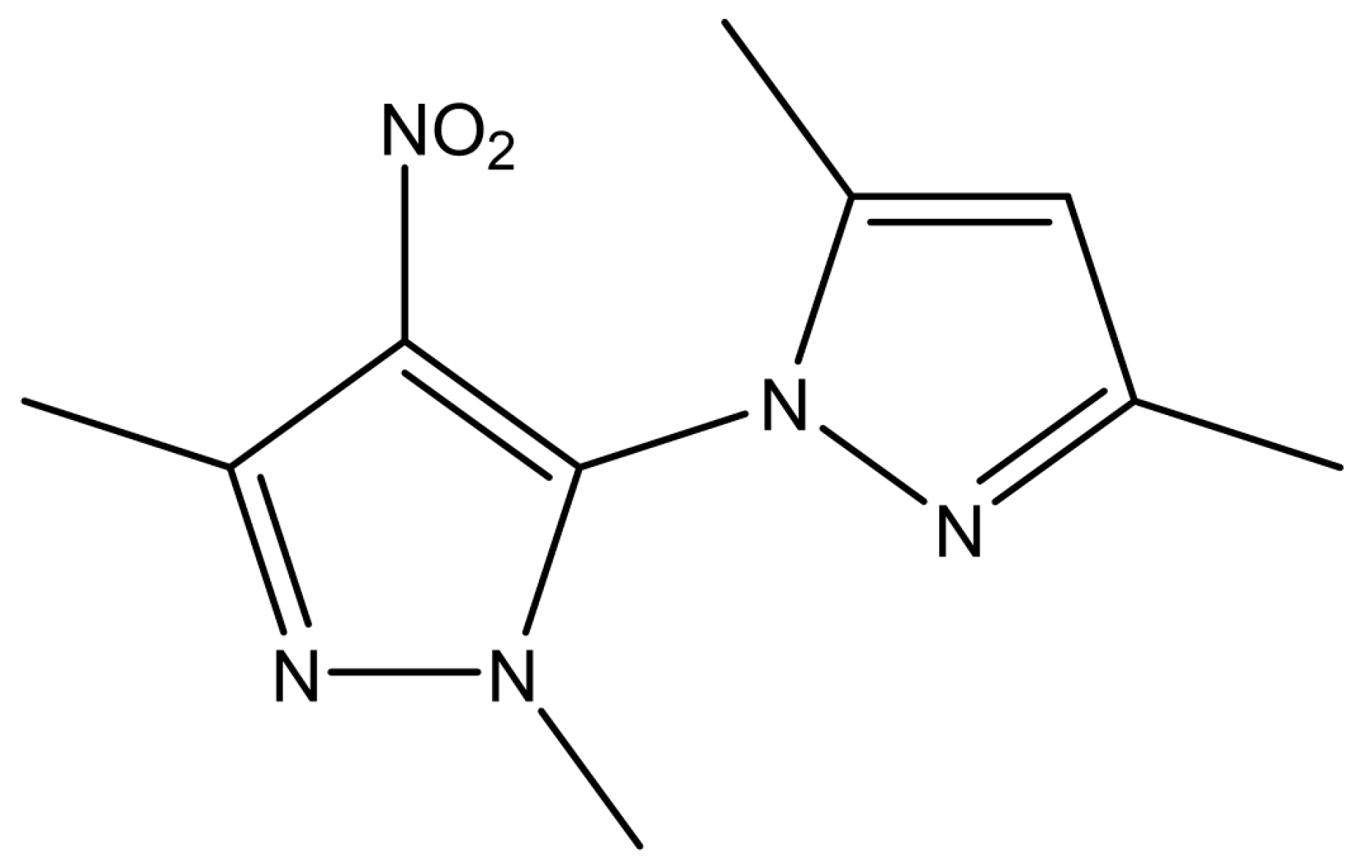
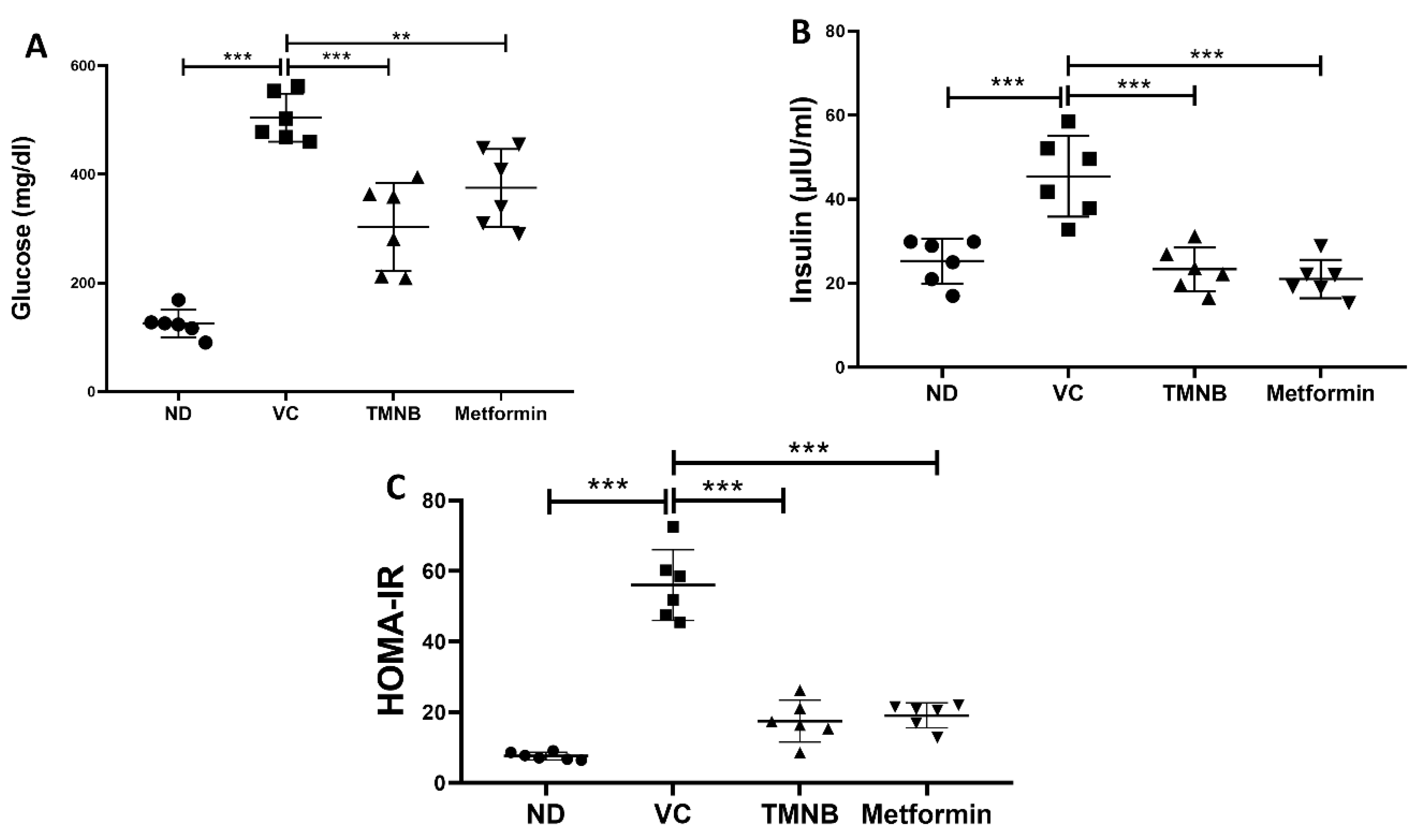
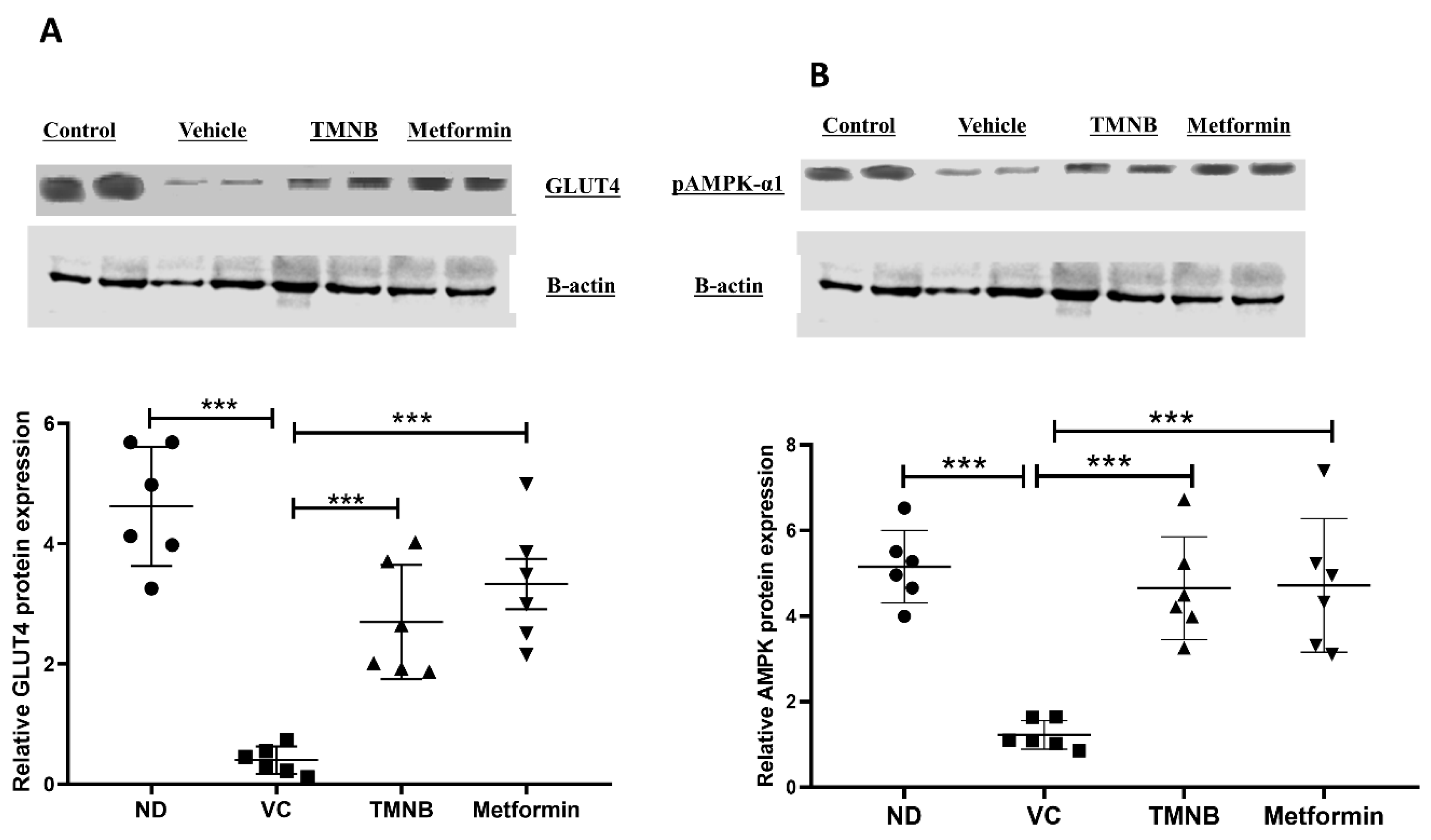
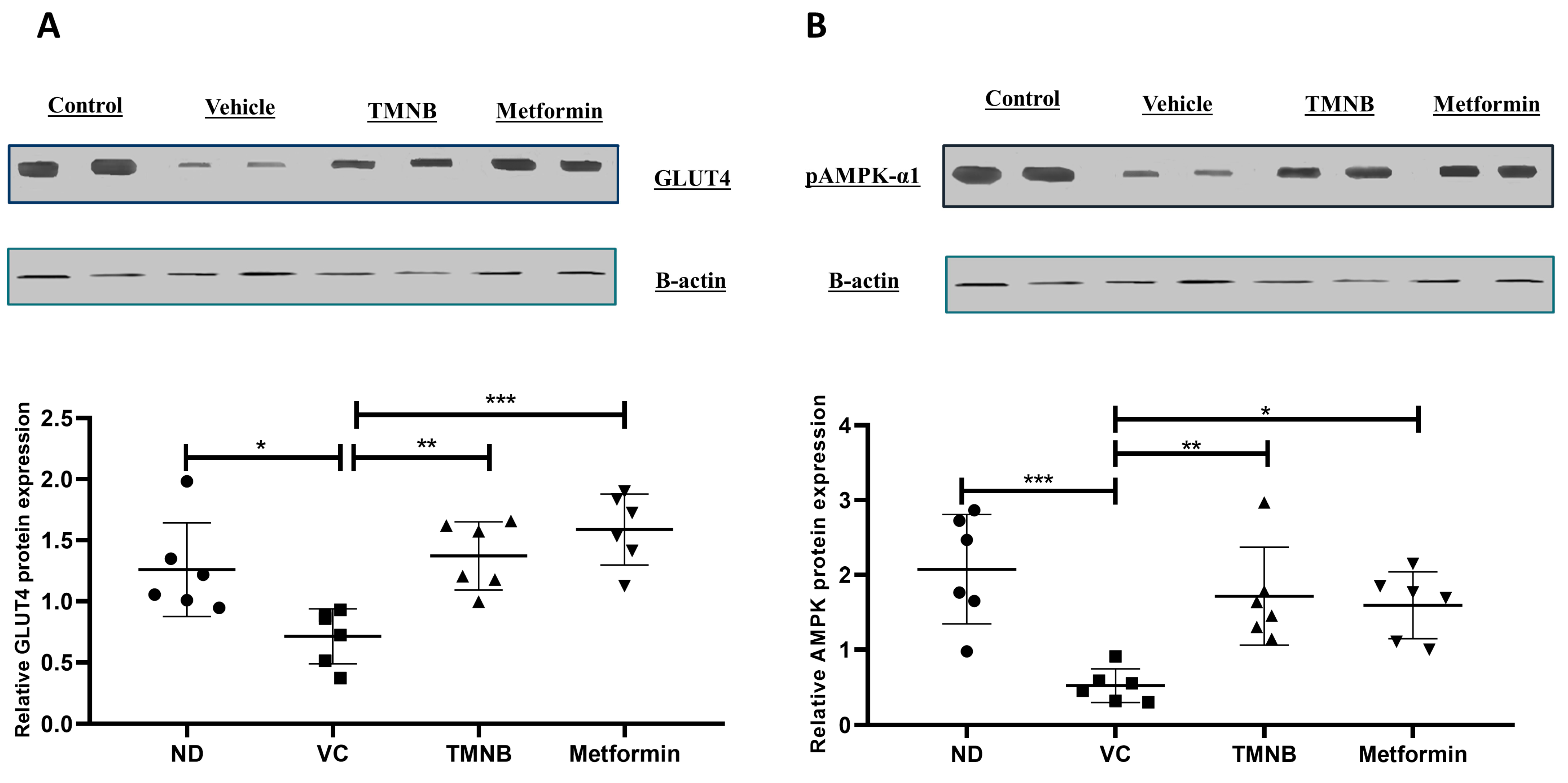
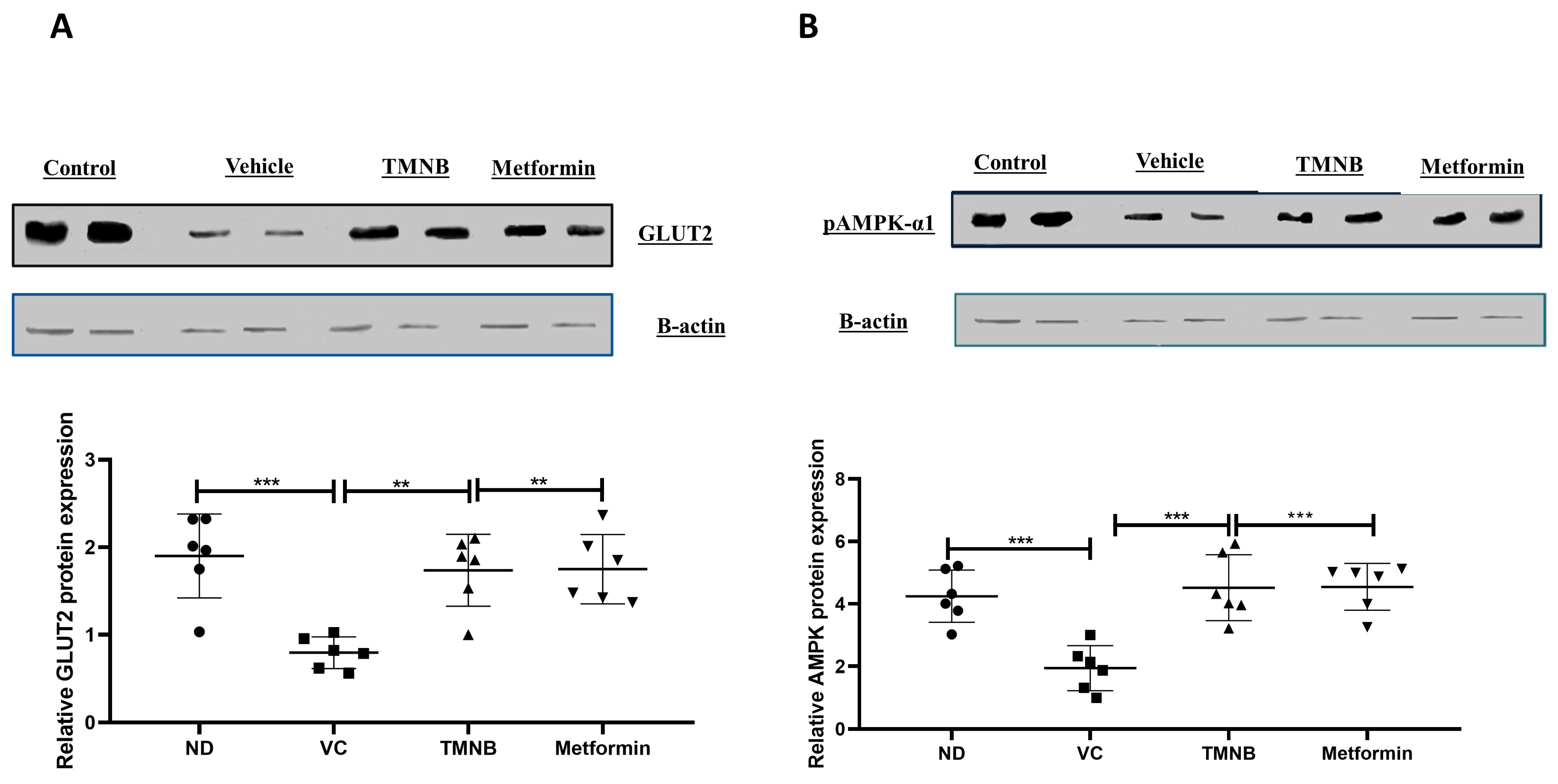
3.3. The Hypoglycemic Effect of 2′,3,3,5′-Tetramethyl-4′-Nitro-2′H-1,3′-Bipyrazole (TMNB)
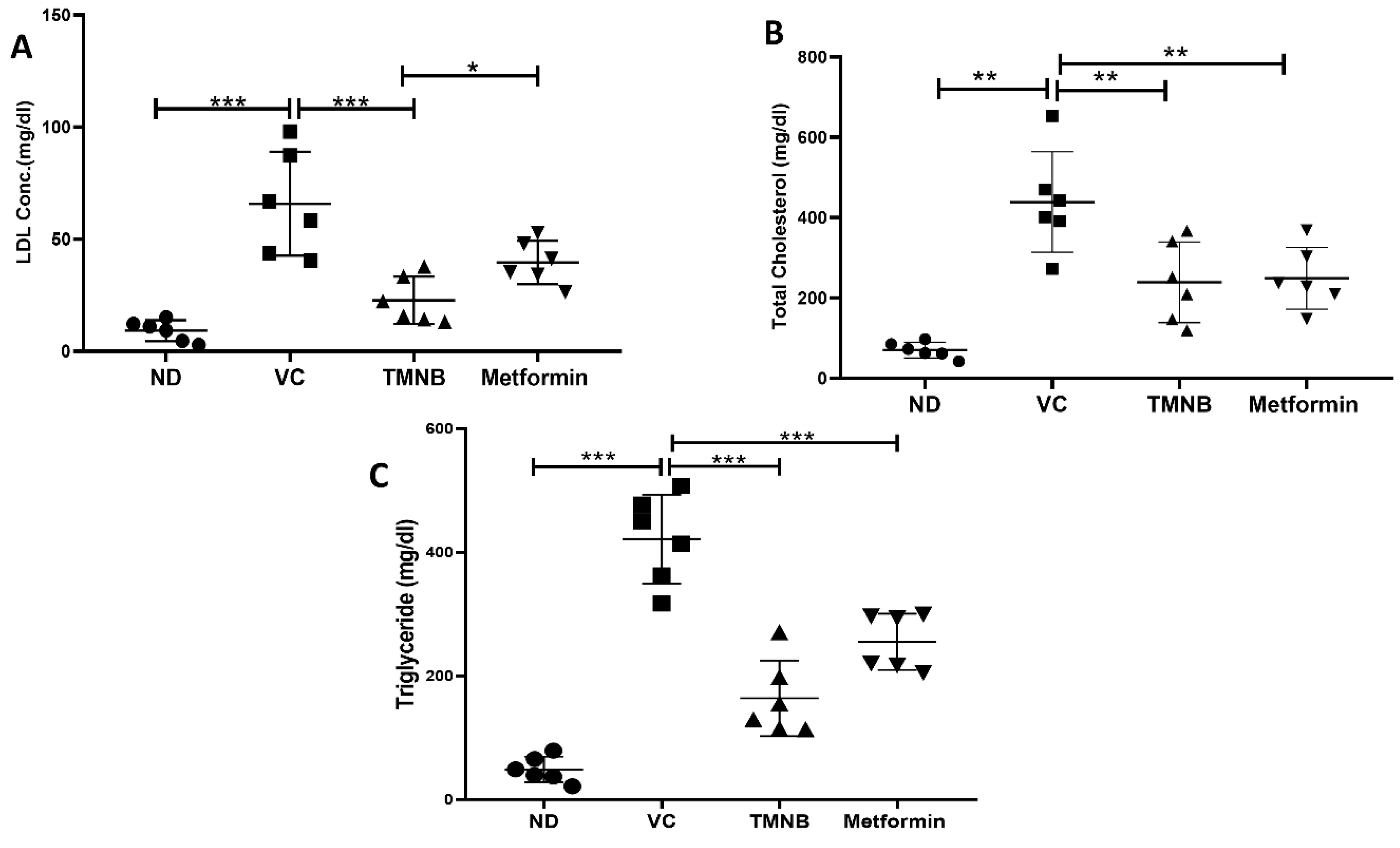
3.4. The Effect of 2′,3,3,5′-Tetramethyl-4′-Nitro-2′H-1,3′-Bipyrazole (TMNB) on Lipid Profile
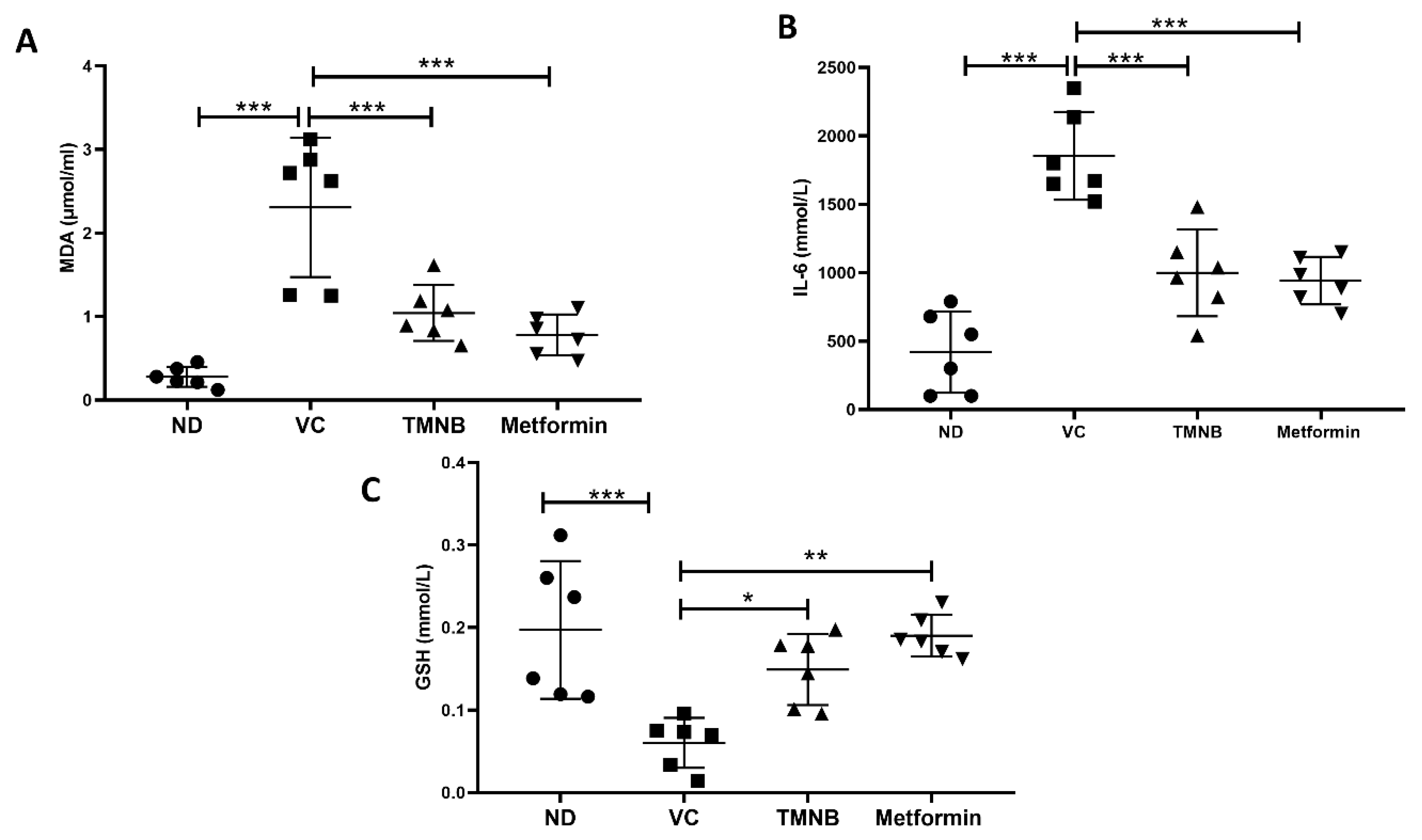
3.5. The Effect of 2′,3,3,5′-Tetramethyl-4′-Nitro-2′H-1,3′-Bipyrazole (TMNB) on MDA, GSH, and IL-6
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Of, D.; Mellitus, D. Diagnosis and classification of diabetes mellitus. Diabetes Care 2014, 37, 81–90. [Google Scholar] [CrossRef]
- Asmat, U.; Abad, K.; Ismail, K. Diabetes mellitus and oxidative stress—A concise review. Saudi Pharm. J. 2016, 24, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Czech, M.P. The GLUT4 Glucose Transporter. Cell Metab. 2007, 5, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.Y.; Shevchuk, M.; Opfermann, J.; Guo, T.; Santoro, M.; Fisher, J.P.; Kim, P.C.W. Trophoblast–endothelium signaling involves angiogenesis and apoptosis in a dynamic bioprinted placenta model. Biotechnol. Bioeng. 2019, 116, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.S.; Kim, T.H.; Kim, M.Y.; Park, J.M.; Ahn, Y.H. Transcriptional regulation of glucose sensors in pancreatic β cells and liver: An update. Sensors 2010, 10, 5031–5053. [Google Scholar] [CrossRef]
- Cool, B.; Zinker, B.; Chiou, W.; Kifle, L.; Cao, N.; Perham, M.; Dickinson, R.; Adler, A.; Gagne, G.; Iyengar, R.; et al. Identification and characterization of a small molecule AMPK activator that treats key components of type 2 diabetes and the metabolic syndrome. Cell Metab. 2006, 3, 403–416. [Google Scholar] [CrossRef]
- Vlavcheski, F.; Baron, D.; Vlachogiannis, I.A.; Macpherson, R.E.K.; Tsiani, E. Carnosol increases skeletal muscle cell glucose uptake via ampk-dependent glut4 glucose transporter translocation. Int. J. Mol. Sci. 2018, 19, 1321. [Google Scholar] [CrossRef]
- Esser, N.; Legrand-Poels, S.; Piette, J.; Scheen, A.J.; Paquot, N. Inflammation as a link between obesity, metabolic syndrome and type 2 diabetes. Diabetes Res. Clin. Pract. 2014, 105, 141–150. [Google Scholar] [CrossRef]
- Kubaszek, A.; Pihlajamäki, J.; Komarovski, V.; Lindi, V.; Lindström, J.; Eriksson, J.; Valle, T.T.; Hämäläinen, H.; Ilanne-Parikka, P.; Keinänen-Kiukaanniemi, S.; et al. Promoter Polymorphisms of the TNF-α (G-308A) and IL-6 (C-174G) Genes Predict the Conversion from Impaired Glucose Tolerance to Type 2 Diabetes the Finnish Diabetes Prevention Study. Diabetes 2003, 52, 1872–1876. [Google Scholar] [CrossRef]
- Tokuyama, Y.; Sturis, J.; DePaoli, A.M.; Takeda, J.; Stoffel, M.; Tang, J.; Sun, X.; Polonsky, K.S.; Bell, G.I. Evolution of beta-cell dysfunction in the male Zucker diabetic fatty rat. Diabetes 1995, 44, 1447–1457. [Google Scholar] [CrossRef]
- Ma, K.; Nunemaker, C.S.; Wu, R.; Chakrabarti, S.K.; Taylor-Fishwick, D.A.; Nadler, J.L. 12-Lipoxygenase Products Reduce Insulin Secretion and {beta}-Cell Viability in Human Islets. J. Clin. Endocrinol. Metab. 2010, 95, 887–893. [Google Scholar] [CrossRef] [PubMed]
- Kayama, Y.; Raaz, U.; Jagger, A.; Adam, M.; Schellinger, I.N.; Sakamoto, M.; Suzuki, H.; Toyama, K.; Spin, J.M.; Tsao, P.S. Diabetic Cardiovascular Disease Induced by Oxidative Stress. Int. J. Mol. Sci. 2015, 16, 25234–25263. [Google Scholar] [CrossRef] [PubMed]
- Kumar Bhaumik, P.; Ghosh, K.; Chattopadhyay, S. Synthetic strategies, crystal structures and biological activities of metal complexes with the members of azole family: A review. Polyhedron 2021, 200, 115093. [Google Scholar] [CrossRef]
- Ruffell, K.; Smith, F.R.; Green, M.T.; Nicolle, S.M.; Inman, M.; Lewis, W.; Hayes, C.J.; Moody, C.J. Diazophosphonates: Effective Surrogates for Diazoalkanes in Pyrazole Synthesis. Chem. Eur. J. 2021, 27, 13703–13708. [Google Scholar] [CrossRef] [PubMed]
- Dadiboyena, S.; Nefzi, A. Synthesis of functionalized tetrasubstituted pyrazolyl heterocycles—A review. Eur. J. Med. Chem. 2011, 46, 5258–5275. [Google Scholar] [CrossRef]
- El-Sayed, M.A.-A.; Abdel-Aziz, N.I.; Abdel-Aziz, A.A.-M.; El-Azab, A.S.; ElTahir, K.E. Synthesis, biological evaluation and molecular modeling study of pyrazole and pyrazoline derivatives as selective COX-2 inhibitors and anti-inflammatory agents. Part 2. Bioorg. Med. Chem. 2012, 20, 3306–3316. [Google Scholar] [CrossRef]
- Akhramez, S.; Oumessaoud, A.; Hibot, A.; Talbi, S.; Hamri, S.; Ketatni, E.M.; Ouchetto, H.; Hafid, A.; Ayad, H.; El Abbadi, N.; et al. Synthesis of pyrazolo-enaminones, bipyrazoles and pyrazolopyrimidines and evaluation of antioxidant and antimicrobial properties. Arab. J. Chem. 2022, 15, 103527. [Google Scholar] [CrossRef]
- Salameh, B.A.; Abu-Safieh, K.A.; AL-Aqrabawi, I.S.; Alsoubani, F.; Tahtamouni, L.H. Synthesis and cytotoxic activity of some new bipyrazole derivatives. Heterocycles 2020, 100, 283–292. [Google Scholar] [CrossRef]
- Lipinski, C.A. Rule of five in 2015 and beyond: Target and ligand structural limitations, ligand chemistry structure and drug discovery project decisions. Adv. Drug Deliv. Rev. 2016, 101, 34–41. [Google Scholar] [CrossRef]
- Attique, S.A.; Hassan, M.; Usman, M.; Atif, R.M.; Mahboob, S.; Al-Ghanim, K.A.; Bilal, M.; Nawaz, M.Z. A Molecular Docking Approach to Evaluate the Pharmacological Properties of Natural and Synthetic Treatment Candidates for Use against Hypertension. Int. J. Environ. Res. Public Health 2019, 16, 923. [Google Scholar] [CrossRef]
- Bai, Q.; Ma, J.; Liu, S.; Xu, T.; Banegas-Luna, A.J.; Pérez-Sánchez, H.; Tian, Y.; Huang, J.; Liu, H.; Yao, X. WADDAICA: A webserver for aiding protein drug design by artificial intelligence and classical algorithm. Comput. Struct. Biotechnol. J. 2021, 19, 3573–3579. [Google Scholar] [CrossRef] [PubMed]
- OECD. Guideline for the Testing of Chemicals TG 471: Bacterial Reverse Mutation Test; OECD: Paris, France, 1997; pp. 1–13. [Google Scholar]
- Al-Ghaithi, F.; El-Ridi, M.R.; Adeghate, E.; Amiri, M.H. Biochemical effects of Citrullus colocynthis in normal and diabetic rats. Mol. Cell. Biochem. 2004, 261, 143–149. [Google Scholar] [CrossRef] [PubMed]
- De MagalhÃes, D.A.; Kume, W.T.; Correia, F.S.; Queiroz, T.S.; Allebrandt Neto, E.W.; Dos Santos, M.P.; Kawashita, N.H.; De França, S.A. High-fat diet and streptozotocin in the induction of type 2 diabetes mellitus: A new proposal. An. Acad. Bras. Cienc. 2019, 91. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Jeon, D.J.; Park, C.E.; You, H.S.; Moon, A.E. Relationship between homeostasis model assessment of insulin resistance and beta cell function and serum 25-hydroxyvitamin D in non-diabetic Korean adults. J. Clin. Biochem. Nutr. 2016, 59, 139–144. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association. 2. Classification and diagnosis of diabetes: Standards of medical care in diabetes 2019. Diabetes Care 2019, 42, S13–S28. [Google Scholar] [CrossRef]
- Babiker, A.; Dubayee, M. Anti-diabetic medications: How to make a choice? Sudan. J. Paediatr. 2017, 17, 11–20. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, Q.; Dang, H.; Liu, X.; Tian, F.; Zhao, J.; Chen, Y.; Zhang, H.; Chen, W. Oral administration of Lactobacillus rhamnosus CCFM0528 improves glucose tolerance and cytokine secretion in high-fat-fed, streptozotocin-induced type 2 diabetic mice. J. Funct. Foods 2014, 10, 318–326. [Google Scholar] [CrossRef]
- Priscilla, D.H.; Jayakumar, M.; Thirumurugan, K. Flavanone naringenin: An effective antihyperglycemic and antihyperlipidemic nutraceutical agent on high fat diet fed streptozotocin induced type 2 diabetic rats. J. Funct. Foods 2015, 14, 363–373. [Google Scholar] [CrossRef]
- Matboli, M.; Saad, M.; Hasanin, A.H.; Saleh, L.A.; Baher, W.; Bekhet, M.M.; Eissa, S. New insight into the role of isorhamnetin as a regulator of insulin signaling pathway in type 2 diabetes mellitus rat model: Molecular and computational approach. Biomed. Pharmacother. 2021, 135, 111176. [Google Scholar] [CrossRef]
- Akinlade, O.M.; Owoyele, B.V.; Soladoye, A.O. Streptozotocin-induced type 1 and 2 diabetes in rodents: A model for studying diabetic cardiac autonomic neuropathy. Afr. Health Sci. 2021, 21, 719–727. [Google Scholar] [CrossRef]
- Jia, S.; Wang, Z.; Han, R.; Zhang, Z.; Li, Y.; Qin, X.; Zhao, M.; Xiang, R.; Yang, J. Incretin mimetics and sodium-glucose co-transporter 2 inhibitors as monotherapy or add-on to metformin for treatment of type 2 diabetes: A systematic review and network meta-analysis. Acta Diabetol. 2021, 58, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Kemp, D.M.; Habener, J.F. Synergistic effect of dimethyl sulfoxide on glucagon-like peptide 1 (GLP-1)-stimulated insulin secretion and gene transcription in INS-1 cells: Characterization and implications. Biochem. Pharmacol. 2002, 64, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Carmean, C.M.; Zhao, L.; Landeche, M.; Chellan, B.; Sargis, R.M. Dimethyl sulfoxide acutely enhances regulated insulin secretion in the MIN6-K8 mouse insulinoma cell line. Histochem. Cell Biol. 2021, 156, 69–73. [Google Scholar] [CrossRef]
- Rivas, D.A.; Fielding, R.A. Skeletal Muscle. Encycl. Hum. Nutr. 2012, 4, 193–199. [Google Scholar] [CrossRef]
- Shan, W.F.; Chen, B.Q.; Zhu, S.J.; Jiang, L.; Zhou, Y.F. Effects of GLUT4 expression on insulin resistance in patients with advanced liver cirrhosis. J. Zhejiang Univ. Sci. B 2011, 12, 677–682. [Google Scholar] [CrossRef]
- Zheng, D.; Maclean, P.S.; Pohnert, S.C.; Knight, J.B.; Olson, A.L.; Winder, W.W.; Dohm, G.L. Regulation of muscle GLUT-4 transcription by AMP-activated protein kinase. J. Appl. Physiol. 2001, 91, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Viollet, B.; Lantier, L.; Devin-Leclerc, J.; Hebrard, S.; Amouyal, C.; Remi Mounier, M.; Foretz, F.A. Targeting the AMPK pathway for the treatment of Type 2 diabetes. Front. Biosci. 2009, 9, 3380–3400. [Google Scholar] [CrossRef]
- Asrafuzzaman, M.; Rahman, M.M.; Mandal, M.; Marjuque, M.; Bhowmik, A.; Rokeya, B.; Hassan, Z.; Faruque, M.O. Oyster mushroom functions as an anti-hyperglycaemic through phosphorylation of AMPK and increased expression of GLUT4 in type 2 diabetic model rats. J. Taibah Univ. Med. Sci. 2018, 13, 465–471. [Google Scholar] [CrossRef]
- Entezari, M.; Hashemi, D.; Taheriazam, A.; Zabolian, A.; Mohammadi, S.; Fakhri, F.; Hashemi, M.; Hushmandi, K.; Ashrafizadeh, M.; Zarrabi, A.; et al. AMPK signaling in diabetes mellitus, insulin resistance and diabetic complications: A pre-clinical and clinical investigation. Biomed. Pharmacother. 2022, 146, 112563. [Google Scholar] [CrossRef]
- Thorens, B. GLUT2, glucose sensing and glucose homeostasis. Diabetologia 2015, 58, 221–232. [Google Scholar] [CrossRef]
- Chadt, A.; Al-Hasani, H. Glucose transporters in adipose tissue, liver, and skeletal muscle in metabolic health and disease. Pflugers Arch. Eur. J. Physiol. 2020, 472, 1273–1298. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.F.; Alam, M.M.; Verma, G.; Akhtar, W.; Akhter, M.; Shaquiquzzaman, M. The therapeutic voyage of pyrazole and its analogs: A review. Eur. J. Med. Chem. 2016, 120, 170–201. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Parhofer, K.G. Diabetic dyslipidemia. Metabolism 2014, 63, 1469–1479. [Google Scholar] [CrossRef] [PubMed]
- Eid, S.; Sas, K.M.; Abcouwer, S.F.; Feldman, E.L.; Gardner, T.W.; Pennathur, S.; Fort, P.E. New insights into the mechanisms of diabetic complications: Role of lipids and lipid metabolism. Diabetologia 2019, 62, 1539–1549. [Google Scholar] [CrossRef]
- Oguntibeju, O.O. Type 2 diabetes mellitus, oxidative stress and inflammation: Examining the links. Int. J. Physiol. Pathophysiol. Pharmacol. 2019, 11, 45–63. [Google Scholar]
- Chikezie, P.C.; Ojiako, O.A.; Ogbuji, A.C. Oxidative stress in diabetes mellitus. Int. J. Biol. Chem. 2015, 9, 92–109. [Google Scholar] [CrossRef]
- Su, L.J.; Zhang, J.H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.Y. Reactive Oxygen Species-Induced Lipid Peroxidation in Apoptosis, Autophagy, and Ferroptosis. Oxid. Med. Cell. Longev. 2019, 2019, 5080843. [Google Scholar] [CrossRef]
- Morales, M.; Munné-Bosch, S. Malondialdehyde: Facts and artifacts. Plant Physiol. 2019, 180, 1246–1250. [Google Scholar] [CrossRef]
- Akbari, M.; Hassan-Zadeh, V. IL-6 signalling pathways and the development of type 2 diabetes. Inflammopharmacology 2018, 26, 685–698. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Sequence | Animal ID | Dose (mg/kg) | Short-Term Result (48 h) | Long-Term Result (12 Days) |
|---|---|---|---|---|
| 1 | 01 | 20 | Survival | Survival |
| 2 | 02 | 20 | Survival | Survival |
| 3 | 03 | 20 | Survival | Survival |
| 4 | 04 | 20 | Survival | Survival |
| 5 | 05 | 20 | Survival | Survival |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alqudah, A.; Qnais, E.Y.; Wedyan, M.A.; Altaber, S.; Abudalo, R.; Gammoh, O.; Alkhateeb, H.; Bataineh, S.; Athamneh, R.Y.; Oqal, M.; et al. New Treatment for Type 2 Diabetes Mellitus Using a Novel Bipyrazole Compound. Cells 2023, 12, 267. https://doi.org/10.3390/cells12020267
Alqudah A, Qnais EY, Wedyan MA, Altaber S, Abudalo R, Gammoh O, Alkhateeb H, Bataineh S, Athamneh RY, Oqal M, et al. New Treatment for Type 2 Diabetes Mellitus Using a Novel Bipyrazole Compound. Cells. 2023; 12(2):267. https://doi.org/10.3390/cells12020267
Chicago/Turabian StyleAlqudah, Abdelrahim, Esam Y. Qnais, Mohammed A. Wedyan, Sara Altaber, Rawan Abudalo, Omar Gammoh, Hakam Alkhateeb, Sajeda Bataineh, Rabaa Y. Athamneh, Muna Oqal, and et al. 2023. "New Treatment for Type 2 Diabetes Mellitus Using a Novel Bipyrazole Compound" Cells 12, no. 2: 267. https://doi.org/10.3390/cells12020267
APA StyleAlqudah, A., Qnais, E. Y., Wedyan, M. A., Altaber, S., Abudalo, R., Gammoh, O., Alkhateeb, H., Bataineh, S., Athamneh, R. Y., Oqal, M., Abu-Safieh, K., & McClements, L. (2023). New Treatment for Type 2 Diabetes Mellitus Using a Novel Bipyrazole Compound. Cells, 12(2), 267. https://doi.org/10.3390/cells12020267