
Unveiling Neuroprotection and Regeneration Mechanisms in Optic Nerve Injury: Insight from Neural Progenitor Cell Therapy with Focus on Vps35 and Syntaxin12

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Hypoxic Damage Conditioning
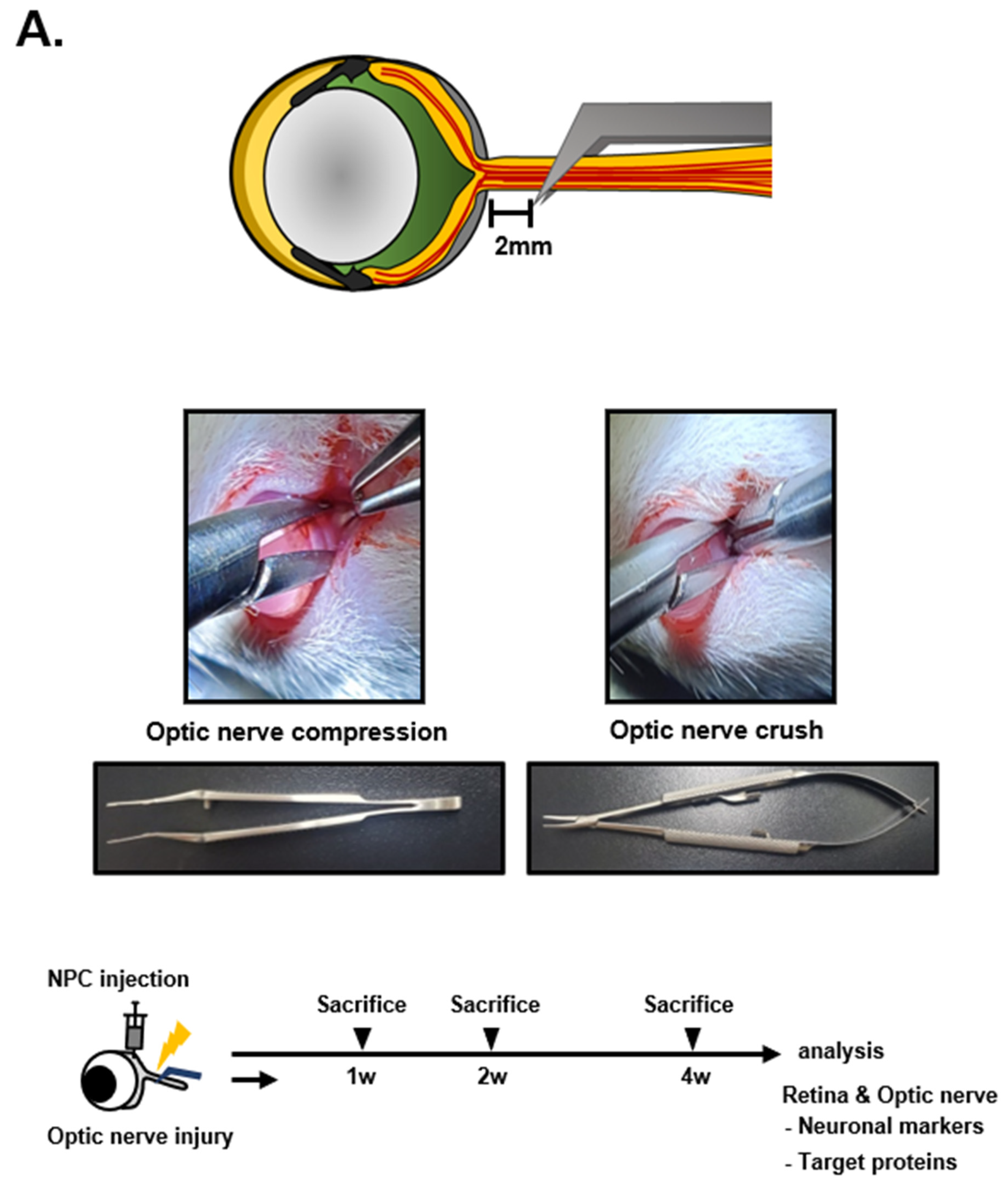
2.2. Construction of the Optic Nerve Compression (ONCo) and Crush (ONCr) Models
2.3. Immunoblot Analysis
2.4. Proteomics
2.5. Data Processing and Bioinformatics
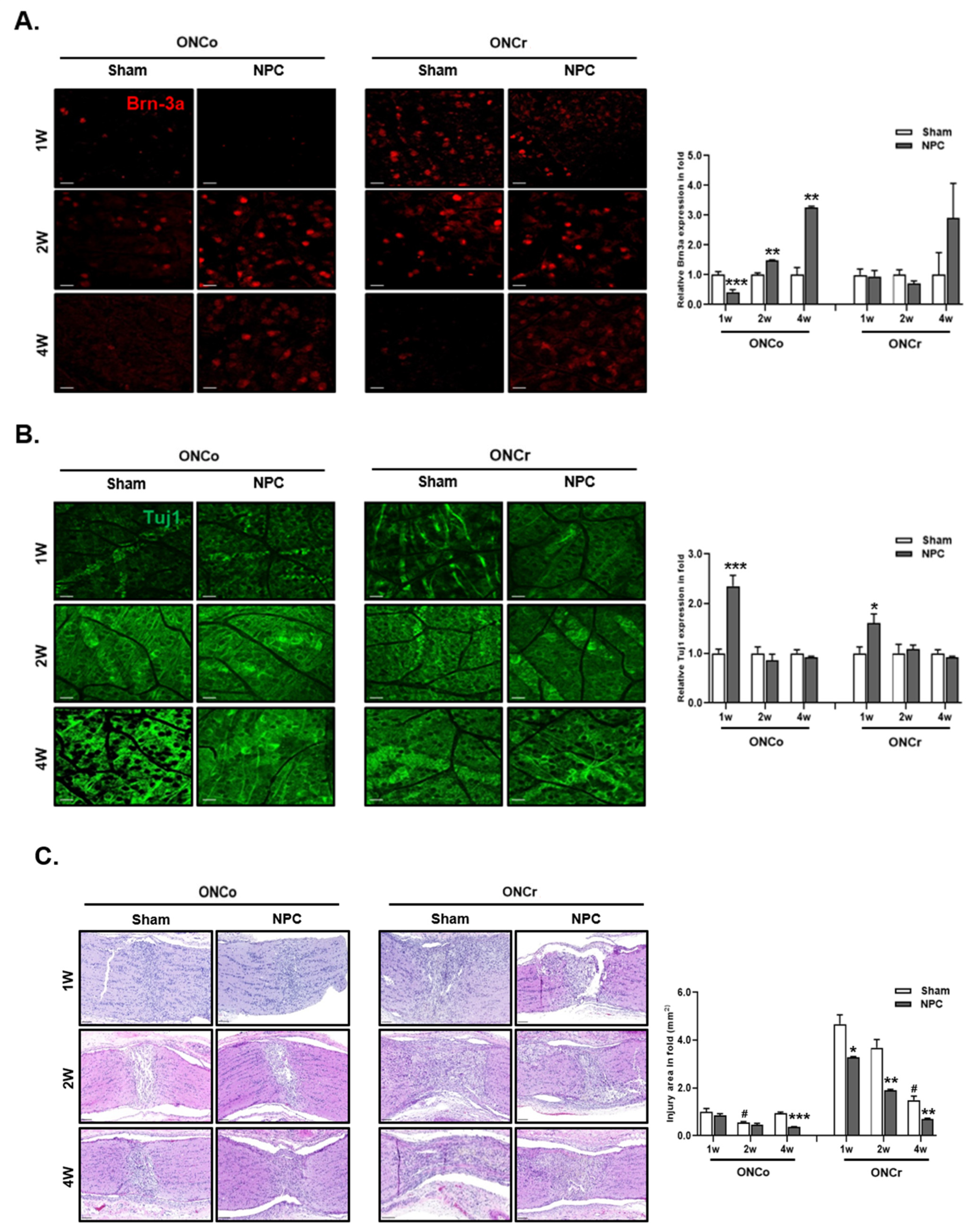
2.6. Immunofluorescence of Retinal Whole Mounts and RGC Survival Analysis
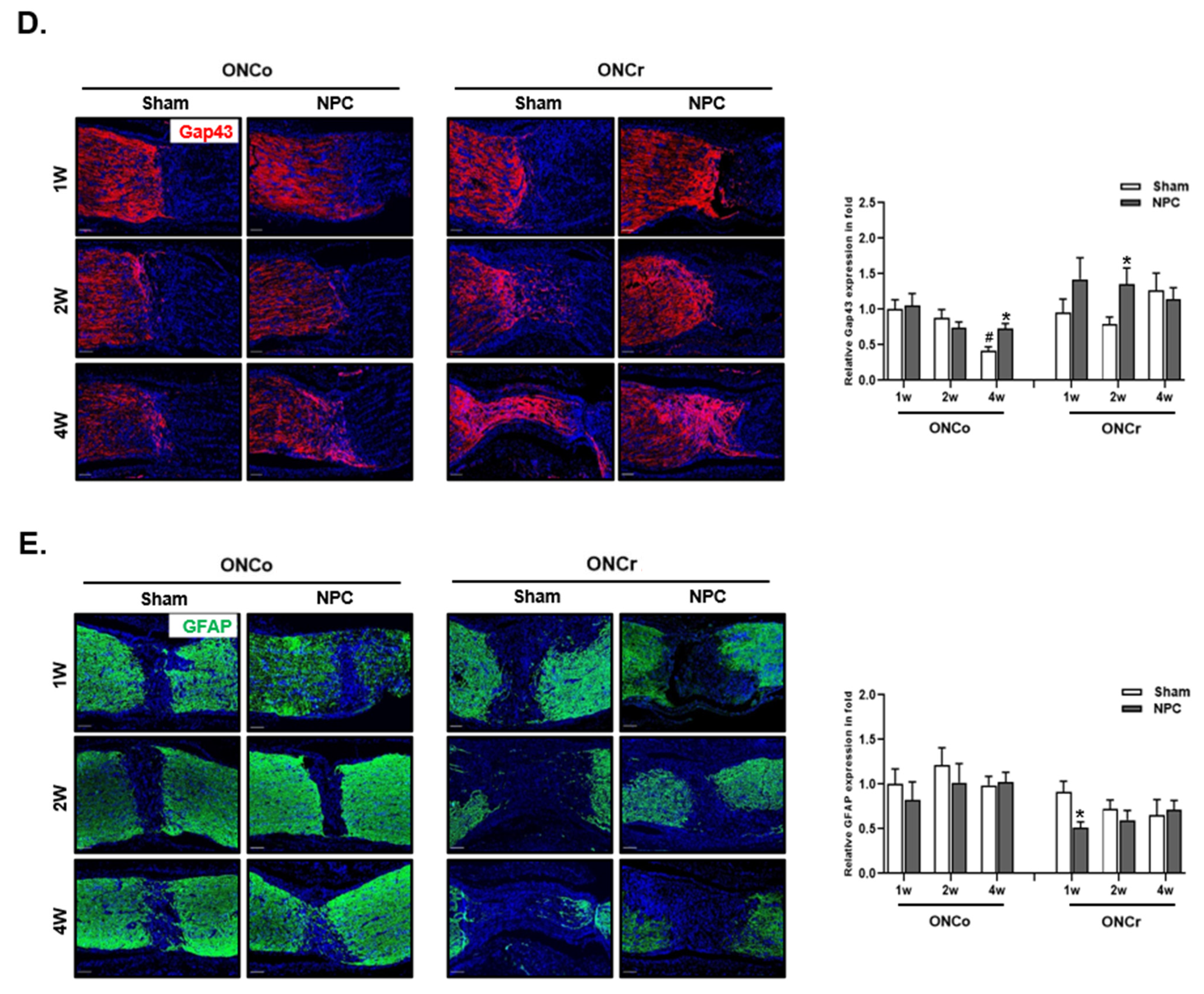
2.7. Optic Nerve Histological Analysis
2.8. Small-Interfering RNA for Vps35 Protein
2.9. Statistical Analyses
3. Results
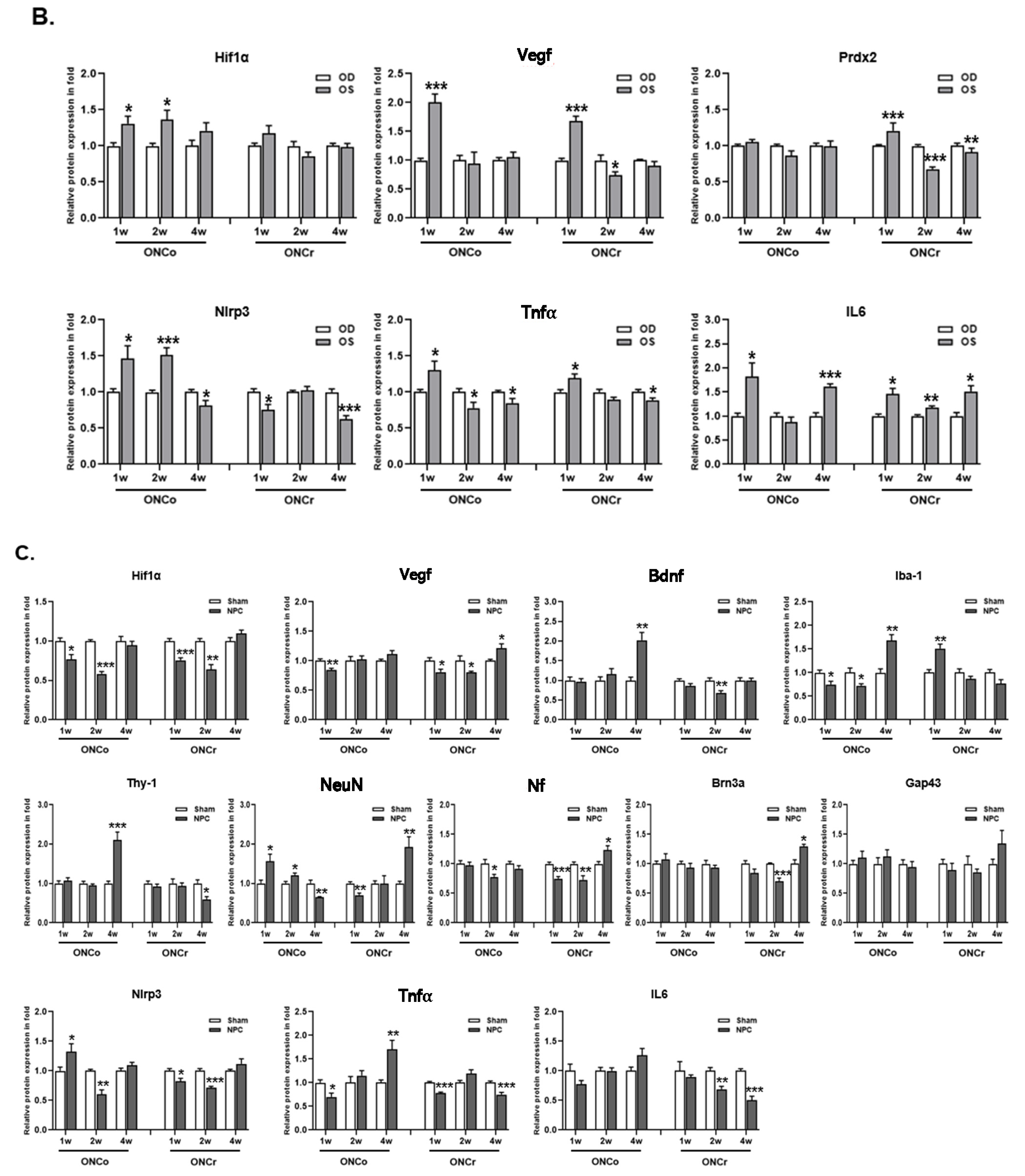
3.1. Comparison of Optic Nerve Injury Models
3.2. NPCs Protect Retinal RGCs in Optic Nerve Injury Models
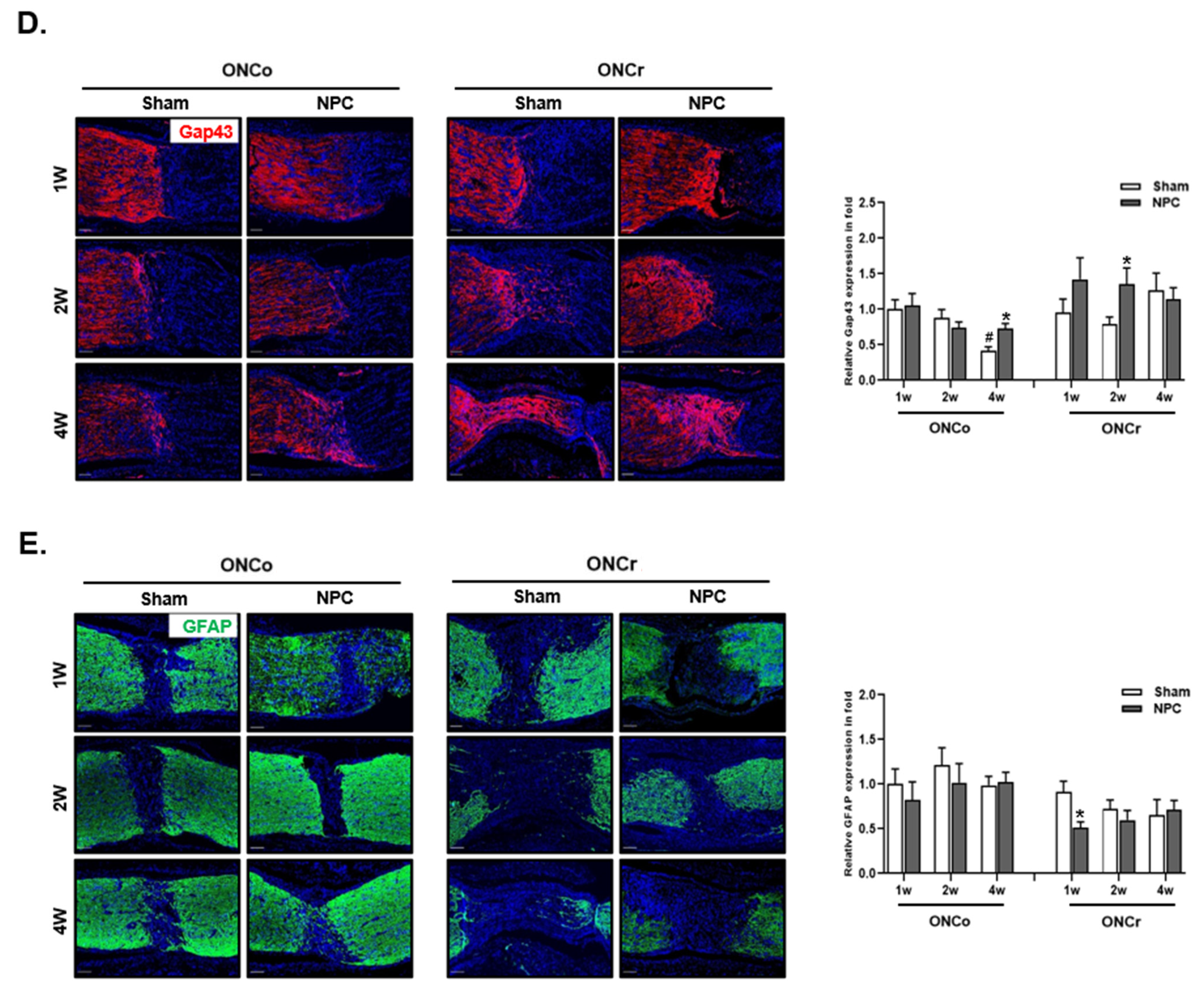
3.3. Effects of NPCs on Optic Nerve Axon Recovery in Each Model
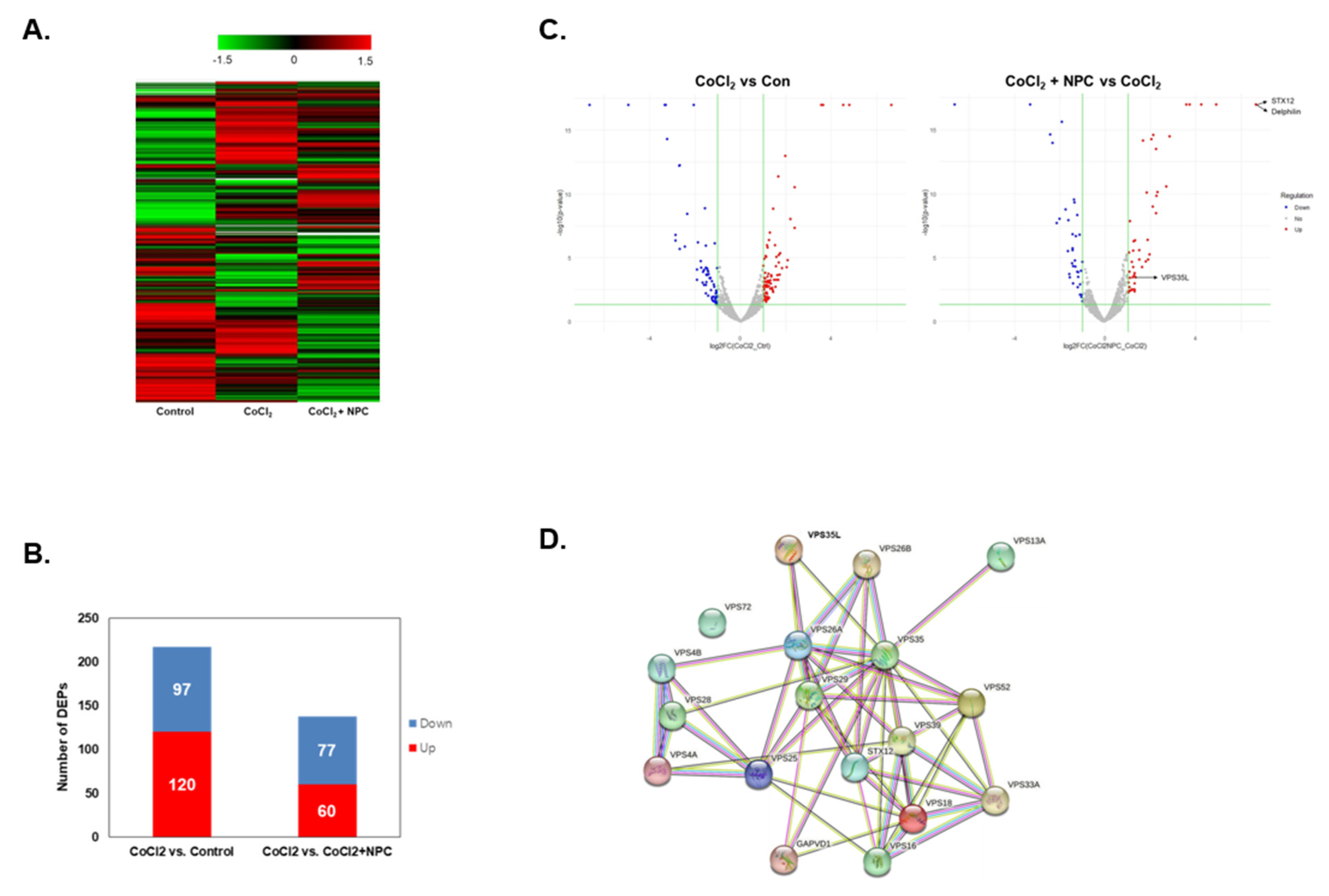
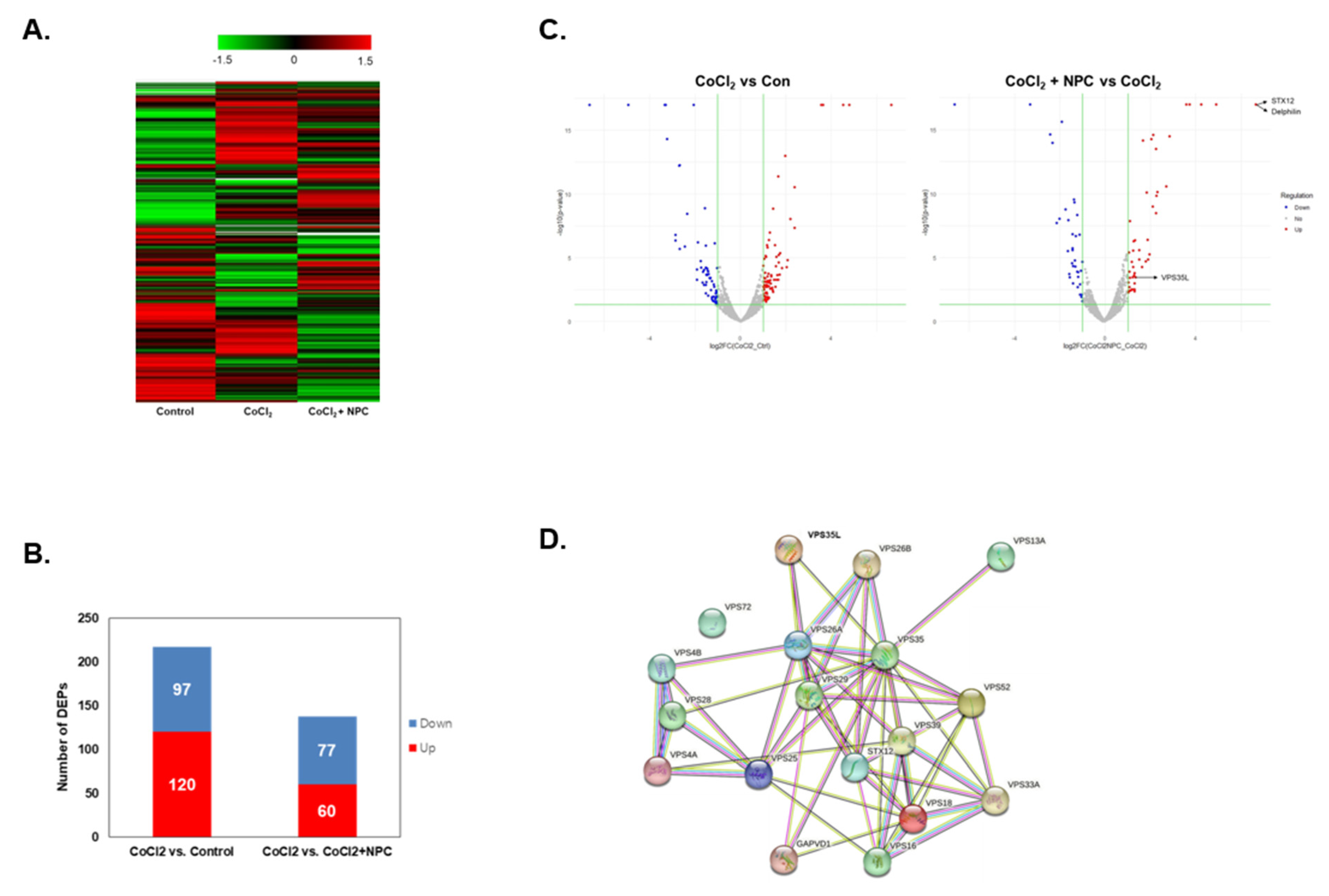
3.4. Hierarchical Clustering and Gene Ontology
3.5. Identification of Regulated Target Proteins during Recovery by NPC in Hypoxia-Damaged Retinal Precursor Cells
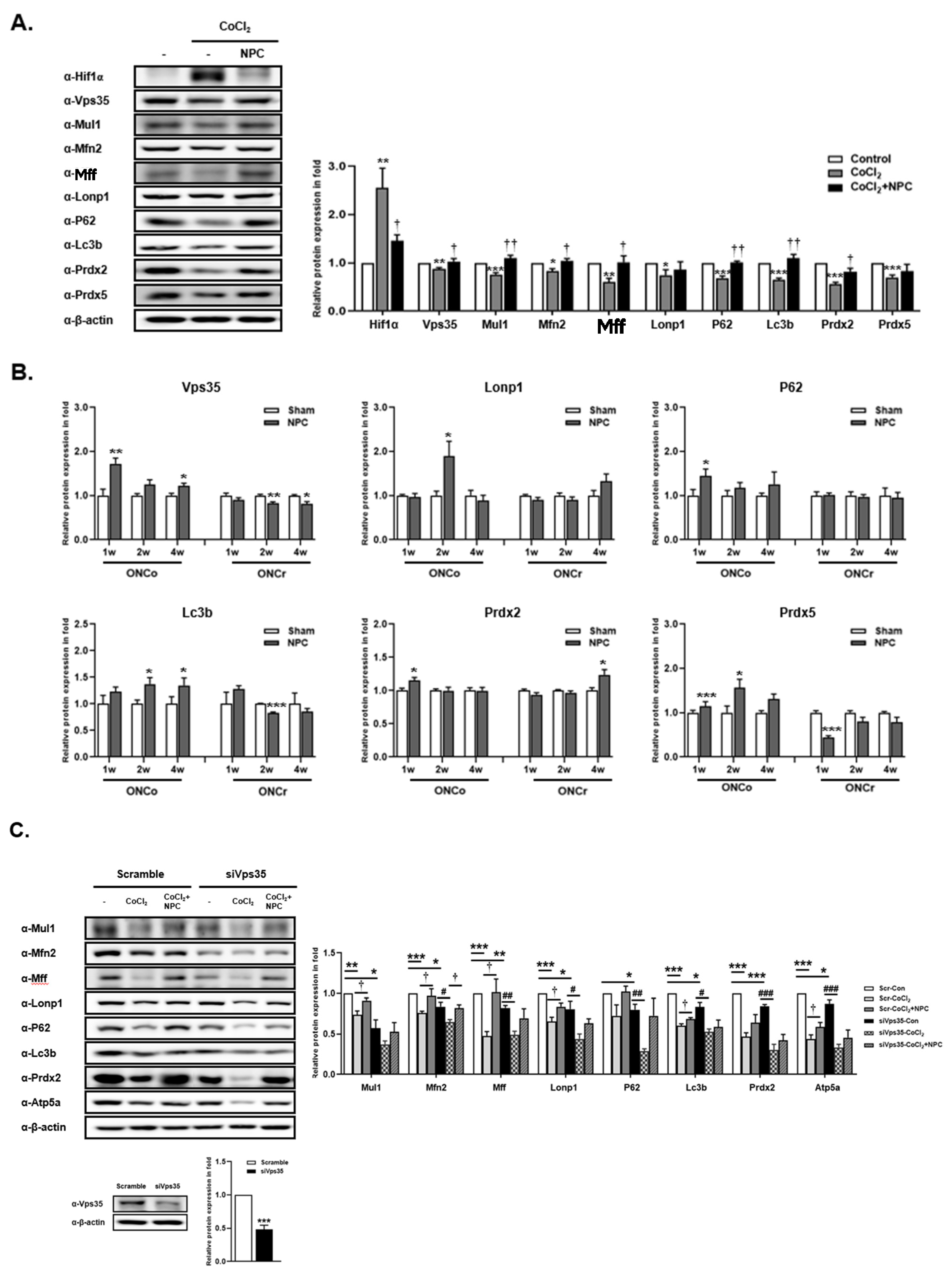
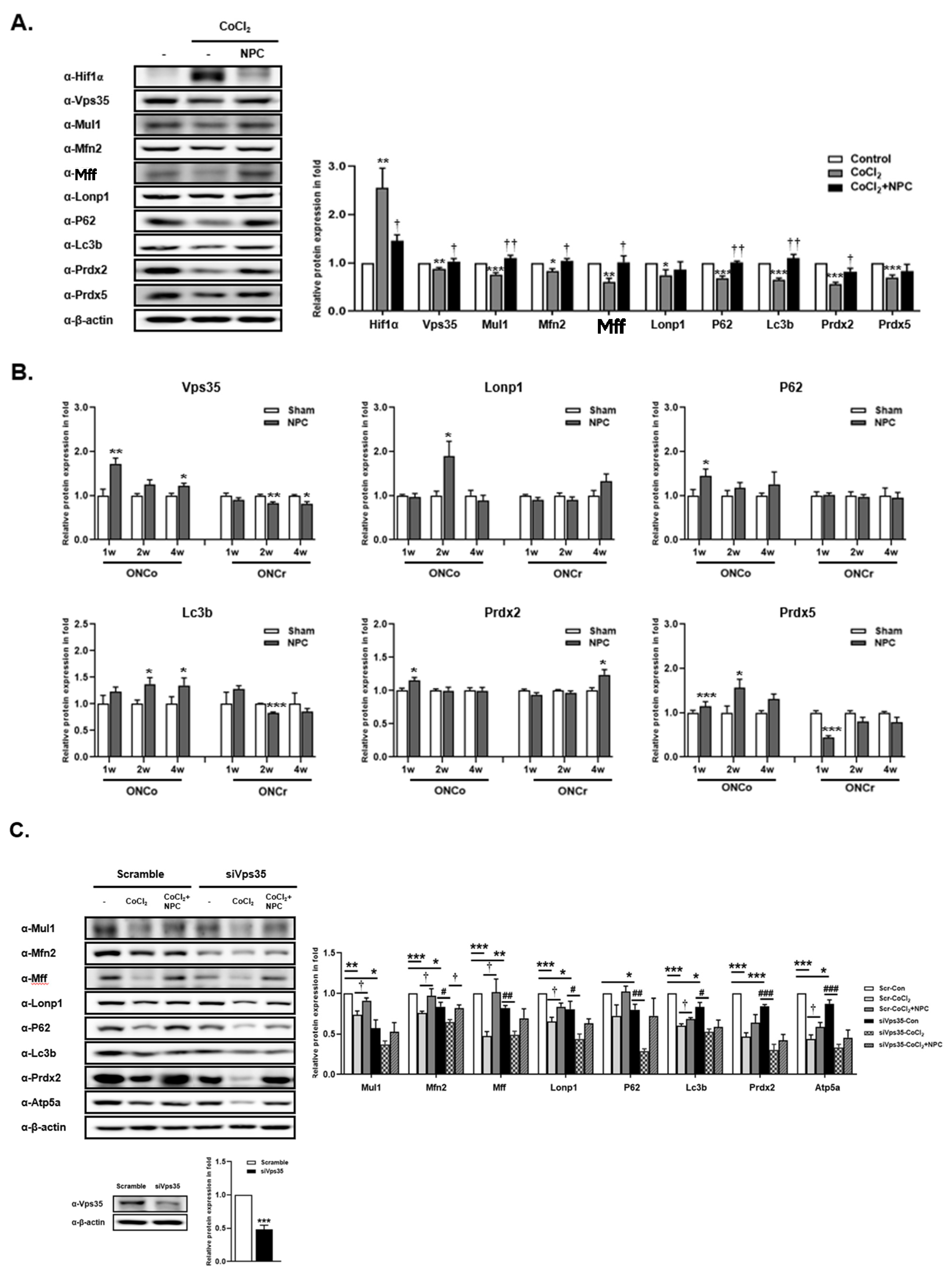
3.6. Role of VPS35 and Recovery of Mitochondrial Function Induced by NPCs
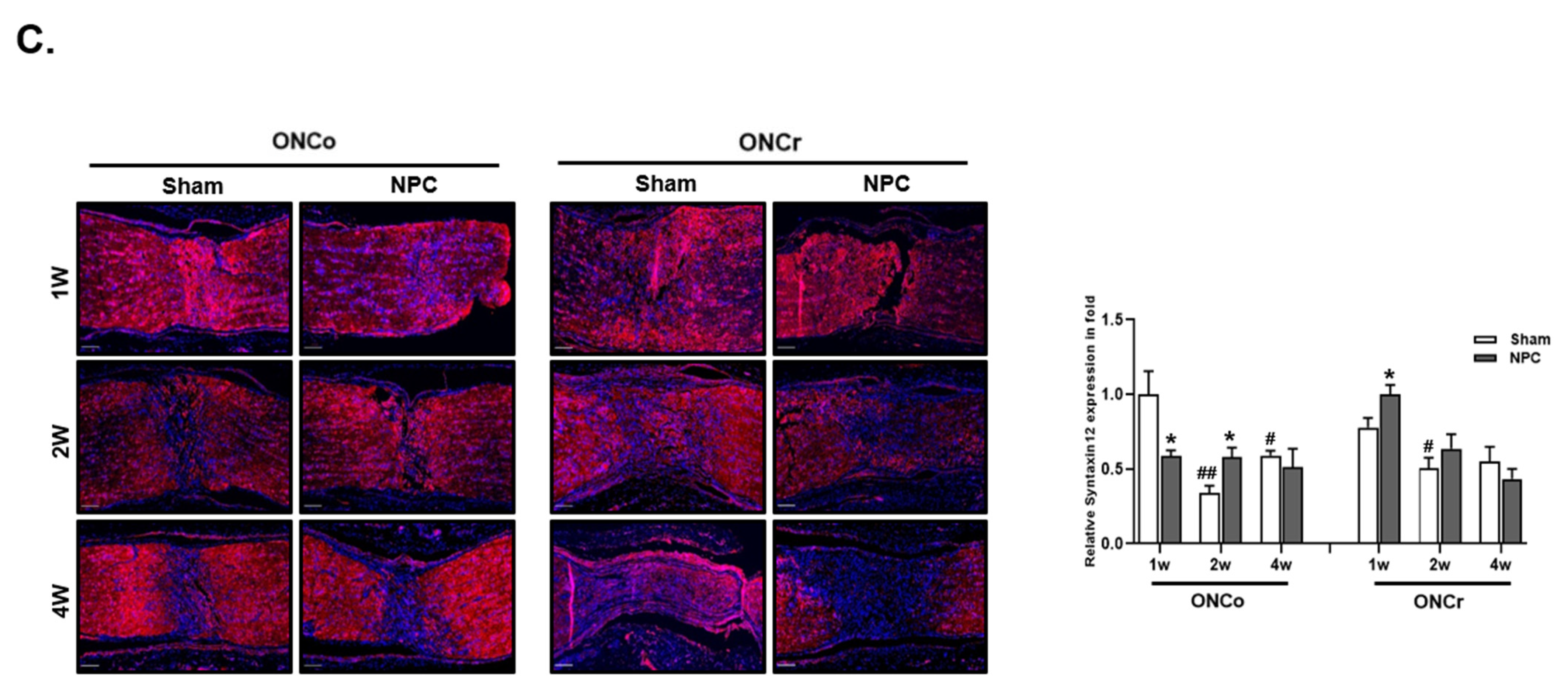
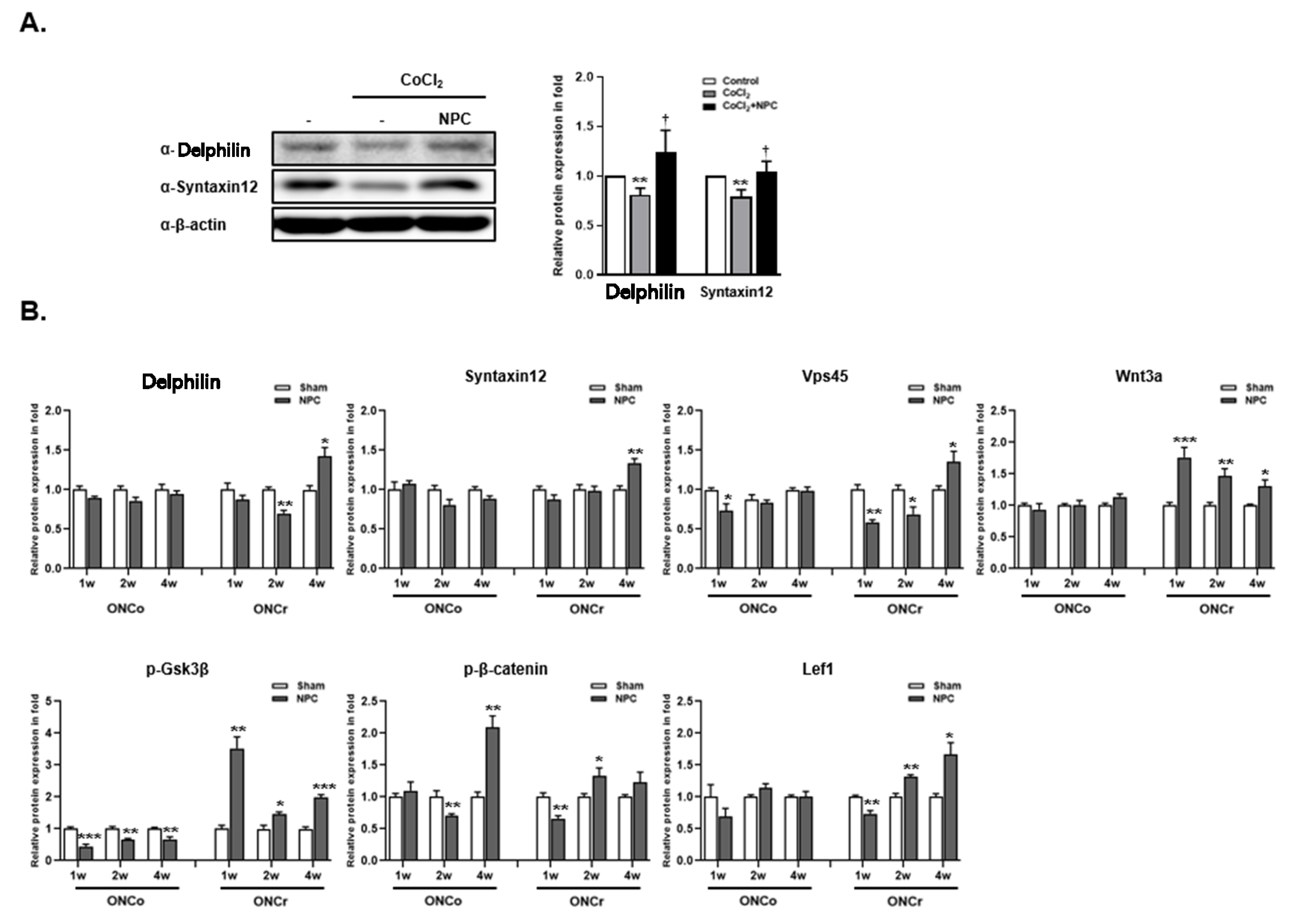
3.7. Identification of Syntaxin12, a New Target Protein Regulated by NPCs during RGCs Recovery
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
List of Abbreviations
| NPCs | Neural progenitor cells |
| ONCo | Optic nerve compression |
| ONCr | Optic nerve crush |
| RGCs | Retinal ganglion cells |
| OS | Oculus sinister |
| OD | Oculus dexter |
| PBS | Phosphate-buffered saline |
| BSS | Balanced salt solution |
| H&E | Hematoxylin and Eosin |
| DEPs | Differentially expressed proteins |
| Hif-1α | Hypoxia-inducible factor-1 α |
| Vegf | Vascular endothelial growth factor |
| Bdnf | Brain derived neurotrophic factor |
| Iba-1 | Ionized calcium-binding adapter molecule-1 |
| Thy-1 | Thy-1 cell surface antigen |
| NeuN | Neuronal nuclei |
| Nf | Neurofilament |
| Brn3a | POU class4 homeobox1; POU4F1 |
| Gap43 | Growth associated protein 43 |
| Nlrp3 | NLR family pyrin domain containing 3 |
| Tnfα | Tumor necrosis factor α |
| IL6 | Interleukin 6 |
| Tuj1 | Neuron-specific class Ⅲ beta-tubulin |
| Gfap | Glial fibrillary acidic protein |
| Mul1 | Mitochondrial E3 ubiquitin protein ligase 1 |
| Mfn2 | Mitofusin 2 |
| Mff | Mitochondrial fission factor |
| Lonp1 | Lon peptidase 1, mitochondrial |
| P62 | Sequestosome 1; SQSTM1 |
| Prdx2 | Peroxiredoxin 2 |
| Prdx5 | Peroxiredoxin 5 |
| Atp5a | ATP synthase subunit α, mitochondrial |
| Gsk3β | Glycogen synthase kinase 3 β |
| Lef1 | Lymphoid enhancer binding factor 1 |
References
- Sanchez-Migallon, M.C.; Valiente-Soriano, F.J.; Nadal-Nicolas, F.M.; Vidal-Sanz, M.; Agudo-Barriuso, M. Apoptotic Retinal Ganglion Cell Death After Optic Nerve Transection or Crush in Mice: Delayed RGC Loss With BDNF or a Caspase 3 Inhibitor. Invest. Ophthalmol. Vis. Sci. 2016, 57, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Kang, E.Y.; Liu, P.K.; Wen, Y.T.; Quinn, P.M.J.; Levi, S.R.; Wang, N.K.; Tsai, R.K. Role of Oxidative Stress in Ocular Diseases Associated with Retinal Ganglion Cells Degeneration. Antioxidants 2021, 10, 1948. [Google Scholar] [CrossRef] [PubMed]
- Au, N.P.B.; Ma, C.H.E. Neuroinflammation, Microglia and Implications for Retinal Ganglion Cell Survival and Axon Regeneration in Traumatic Optic Neuropathy. Front. Immunol. 2022, 13, 860070. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.S.; Shanmugalingam, U.; Smith, P.D. Harnessing Astrocytes and Muller Glial Cells in the Retina for Survival and Regeneration of Retinal Ganglion Cells. Cells 2021, 10, 1339. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, N.; Chrenek, M.A.; Girardot, P.E.; Wang, J.; Sellers, J.T.; Geisert, E.E.; Brenner, C.; Nickerson, J.M.; Boatright, J.H.; et al. Systemic Treatment with Nicotinamide Riboside Is Protective in Two Mouse Models of Retinal Ganglion Cell Damage. Pharmaceutics 2021, 13, 893. [Google Scholar] [CrossRef]
- Tang, Z.; Zhang, S.; Lee, C.; Kumar, A.; Arjunan, P.; Li, Y.; Zhang, F.; Li, X. An optic nerve crush injury murine model to study retinal ganglion cell survival. J. Vis. Exp. 2011, 50, e2685. [Google Scholar]
- Lindborg, J.A.; Tran, N.M.; Chenette, D.M.; DeLuca, K.; Foli, Y.; Kannan, R.; Sekine, Y.; Wang, X.; Wollan, M.; Kim, I.J.; et al. Optic nerve regeneration screen identifies multiple genes restricting adult neural repair. Cell Rep. 2021, 34, 108777. [Google Scholar] [CrossRef]
- Park, M.; Kim, H.M.; Shin, H.A.; Lee, S.H.; Hwang, D.Y.; Lew, H. Human Pluripotent Stem Cell-Derived Neural Progenitor Cells Promote Retinal Ganglion Cell Survival and Axon Recovery in an Optic Nerve Compression Animal Model. Int. J. Mol. Sci. 2021, 22, 12529. [Google Scholar] [CrossRef]
- Park, M.; Shin, H.A.; Duong, V.A.; Lee, H.; Lew, H. The Role of Extracellular Vesicles in Optic Nerve Injury: Neuroprotection and Mitochondrial Homeostasis. Cells 2022, 11, 3720. [Google Scholar] [CrossRef]
- Heuss, N.D.; Pierson, M.J.; Montaniel, K.R.; McPherson, S.W.; Lehmann, U.; Hussong, S.A.; Ferrington, D.A.; Low, W.C.; Gregerson, D.S. Retinal dendritic cell recruitment, but not function, was inhibited in MyD88 and TRIF deficient mice. J. Neuroinflammation. 2014, 11, 143. [Google Scholar] [CrossRef]
- Heuss, N.D.; Pierson, M.J.; Roehrich, H.; McPherson, S.W.; Gram, A.L.; Li, L.; Gregerson, D.S. Optic nerve as a source of activated retinal microglia post-injury. Acta Neuropathol. Commun. 2018, 6, 66. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods. 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Tomozumi, I.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 5, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.; Park, M.; Nepali, S.; Lew, H. Hypoxia-Preconditioned Placenta-Derived Mesenchymal Stem Cells Rescue Optic Nerve Axons Via Differential Roles of Vascular Endothelial Growth Factor in an Optic Nerve Compression Animal Model. Mol. Neurobiol. 2020, 57, 3362–3375. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Rho, S.; Kim, G.; Kim, S.R.; Baek, K.H.; Kang, M.; Lew, H. Human umbilical cord blood mononuclear cells and chorionic plate-derived mesenchymal stem cells promote axon survival in a rat model of optic nerve crush injury. Int. J. Mol. Med. 2016, 37, 1170–1180. [Google Scholar] [CrossRef]
- Binda, C.S.; Nakamura, Y.; Henley, J.M.; Wilkinson, K.A. Sorting nexin 27 rescues neuroligin 2 from lysosomal degradation to control inhibitory synapse number. Biochem. J. 2019, 476, 293–306. [Google Scholar] [CrossRef]
- Li, X.; Fan, Z. The epidermal growth factor receptor antibody cetuximab induces autophagy in cancer cells by downregulating HIF-1alpha and Bcl-2 and activating the beclin 1/hVps34 complex. Cancer Res. 2010, 70, 5942–5952. [Google Scholar] [CrossRef]
- Ye, S.Y.; Apple, J.E.; Ren, X.; Tang, F.L.; Yao, L.L.; Wang, Y.G.; Mei, L.; Zhou, Y.G.; Xiong, W.C. Microglial VPS35 deficiency regulates microglial polarization and decreases ischemic stroke-induced damage in the cortex. J. Neuroinflammation. 2019, 16, 235. [Google Scholar] [CrossRef]
- Munsie, L.N.; Milnerwood, A.J.; Seibler, P.; Beccano-Kelly, D.A.; Tatarnikov, I.; Khinda, J.; Volta, M.; Kadgien, C.; Cao, L.P.; Tapia, L.; et al. Retromer-dependent neurotransmitter receptor trafficking to synapses is altered by the Parkinson’s disease VPS35 mutation p.D620N. Hum. Mol. Genet. 2015, 24, 1691–1703. [Google Scholar] [CrossRef]
- Tang, F.L.; Liu, W.; Hu, J.X.; Erion, J.R.; Ye, J.; Mei, L.; Xiong, W.C. VPS35 Deficiency or Mutation Causes Dopaminergic Neuronal Loss by Impairing Mitochondrial Fusion and Function. Cell Rep. 2015, 12, 1631–1643. [Google Scholar] [CrossRef]
- Vidyadhara, D.J.; Lee, J.E.; Chandra, S.S. Role of the endolysosomal system in Parkinson’s disease. J. Neurochem. 2019, 150, 487–506. [Google Scholar] [CrossRef] [PubMed]
- Ambrosio, A.L.; Febvre, H.P.; Di Pietro, S.M. Syntaxin 12 and COMMD3 are new factors that function with VPS33B in the biogenesis of platelet alpha-granules. Blood 2022, 139, 922–935. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.L.; Tan, A.E.; Lim, L.K.; Lee, S.S.; Low, D.Y.; Hong, W. Syntaxin 12, a member of the syntaxin family localized to the endosome. J. Biol. Chem. 1998, 273, 6944–6950. [Google Scholar] [CrossRef]
- Margiotta, A. Role of SNAREs in Neurodegenerative Diseases. Cells 2021, 10, 991. [Google Scholar] [CrossRef]
- Tang, B.L. Syntaxin 16′s Newly Deciphered Roles in Autophagy. Cells 2019, 8, 1655. [Google Scholar] [CrossRef] [PubMed]
- Kishida, S.; Hamao, K.; Inoue, M.; Hasegawa, M.; Matsuura, Y.; Mikoshiba, K.; Fukuda, M.; Kikuchi, A. Dvl regulates endo- and exocytotic processes through binding to synaptotagmin. Genes Cells 2007, 12, 49–61. [Google Scholar] [CrossRef]
- Merz, K.E.; Hwang, J.; Zhou, C.; Veluthakal, R.; McCown, E.M.; Hamilton, A.; Oh, E.; Dai, W.; Fueger, P.T.; Jiang, L.; et al. Enrichment of the exocytosis protein STX4 in skeletal muscle remediates peripheral insulin resistance and alters mitochondrial dynamics via Drp1. Nat. Commun. 2022, 13, 424. [Google Scholar] [CrossRef]
- An, H.K.; Chung, K.M.; Park, H.; Hong, J.; Gim, J.E.; Choi, H.; Lee, Y.W.; Choi, J.; Mun, J.Y.; Yu, S.W. CASP9 (caspase 9) is essential for autophagosome maturation through regulation of mitochondrial homeostasis. Autophagy 2020, 16, 1598–1617. [Google Scholar] [CrossRef]
- Li, N.; Gao, Y.; Zhang, Y.; Deng, Y. An integrated multi-level analysis reveals learning-memory deficits and synaptic dysfunction in the rat model exposure to austere environment. J. Proteom. 2023, 279, 104887. [Google Scholar] [CrossRef]
- Oda, H.; Kihara, K.; Morimoto, Y.; Takeuchi, S. Cell-Based Biohybrid Sensor Device for Chemical Source Direction Estimation. Cyborg Bionic Syst. 2021, 2021, 9. [Google Scholar] [CrossRef]
- Sargent, D.; Moore, D.J. Mechanisms of VPS35-Mediated Neurodegeneration in Parkinson’s Disease. Int. Rev. Mov. Disord. 2021, 2, 221–244. [Google Scholar] [PubMed]
- Chiu, C.C.; Weng, Y.H.; Huang, Y.Z.; Chen, R.S.; Liu, Y.C.; Yeh, T.H.; Lu, C.S.; Lin, Y.W.; Chen, Y.J.; Hsu, C.C.; et al. (D620N) VPS35 causes the impairment of Wnt/beta-catenin signaling cascade and mitochondrial dysfunction in a PARK17 knockin mouse model. Cell Death Dis. 2020, 11, 1018. [Google Scholar] [CrossRef] [PubMed]
- George, A.; Leahy, H.; Zhou, J.; Morin, P.J. The vacuolar-ATPase inhibitor bafilomycin and mutant VPS35 inhibit canonical Wnt signaling. Neurobiol. Dis. 2007, 26, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Gambardella, S.; Biagioni, F.; Ferese, R.; Busceti, C.L.; Frati, A.; Novelli, G.; Ruggieri, S.; Fornai, F. Vacuolar Protein Sorting Genes in Parkinson’s Disease: A Re-appraisal of Mutations Detection Rate and Neurobiology of Disease. Front. Neurosci. 2016, 10, 532. [Google Scholar] [CrossRef] [PubMed]
- Ingersoll, M.A.; Platt, A.M.; Potteaux, S.; Randolph, G.J. Monocyte trafficking in acute and chronic inflammation. Trends Immunol. 2011, 32, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Van Diest, S.A.; Stanisor, O.I.; Boeckxstaens, G.E.; de Jonge, W.J.; van den Wijngaard, R.M. Relevance of mast cell-nerve interactions in intestinal nociception. Biochim. Biophys. Acta. 2012, 1822, 74–84. [Google Scholar] [CrossRef]
- Borovikova, L.V.; Ivanova, S.; Zhang, M.; Yang, H.; Botchkina, G.I.; Watkins, L.R.; Wang, H.; Abumrad, N.; Eaton, J.W.; Tracey, K.J. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature 2000, 405, 458–462. [Google Scholar] [CrossRef]
- Feldman, E.L.; Bennettm, D.L.H.; Nave, K.A.; Jensen, T.S. New horizons in diabetic neuropathy: Mechanisms. Bioenerg. Pain Neuron 2017, 93, 1296–1313. [Google Scholar]
- Nakanishi, Y.; Kang, S.; Kumanogoh, A. Axon guidance molecules in immunometabolic diseases. Inflamm. Regen. 2022, 42, 5. [Google Scholar] [CrossRef]
- Cerani, A.; Tetreault, N.; Menard, C.; Lapalme, E.; Patel, C.; Sitaras, N.; Beaudoin, F.; Leboeuf, D.; De Guire, V.; Binet, F.; et al. Neuron-derived semaphorin 3A is an early inducer of vascular permeability in diabetic retinopathy vianeuropilin-1. Cell Metab. 2013, 18, 505–518. [Google Scholar] [CrossRef]
- Dejda, A.; Mawambo, G.; Cerani, A.; Miloudi, K.; Shao, Z.; Daudelin, J.F.; Boulet, S.; Oubaha, M.; Beaudoin, F.; Akla, N.; et al. Neuropilin-1 mediates myeloid cell chemoattraction and influences retinal neuroimmune crosstalk. J. Clin. Invest. 2014, 124, 4807–4822. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Liu, B.; Yu, Z.; Wang, T.; Song, Y.; Zhuang, R.; Wu, Y.; Su, Y.; Guo, S. Effects of CD100 promote wound healing in diabetic mice. J. Mol. Histol. 2018, 49, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.S.; Gao, N.; Dike, M.; Shkilnyy, O.; Me, R.; Zhang, Y. Opposing effects of neuropilin-1 and -2 on sensory nerve regeneration in wounded corneas: Role of Sema3C in ameliorating diabetic neurotrophic keratopathy. Diabetes 2019, 68, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.L.; Liu, Y.H.; Liu, C.; Qi, M.P.; Liu, R.N.; Zhu, X.F.; Zhou, Q.G.; Chen, Y.Y.; Guo, A.Z.; Hu, C.M. Indirubin Inhibits LPS-Induced Inflammation via TLR4 Abrogation Mediated by the NF-kB and MAPK Signaling Pathways. Inflammation 2017, 40, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sandall, C.F.; Ziehr, B.K.; MacDonald, J.A. ATP-Binding and Hydrolysis in Inflammasome Activation. Molecules 2020, 25, 4572. [Google Scholar] [CrossRef]
- Wang, X.; Kou, W.; Kong, W.; Ma, S.; Xue, Q.; Zou, Y.; Song, A. Magnetic Resonance Images, Pathological Features of Thrombus, and Expression of NLRP Inflammasome in Patients with Acute Ischemic Stroke. Contrast Media Mol. Imaging 2022, 2022, 3464042. [Google Scholar] [CrossRef]
- Cosin-Roger, J.; Simmen, S.; Melhem, H.; Atrott, K.; Frey-Wagner, I.; Hausmann, M.; de Valliere, C.; Spalinger, M.R.; Spielmann, P.; Wenger, R.H.; et al. Hypoxia ameliorates intestinal inflammation through NLRP3/mTOR downregulation and autophagy activation. Nat. Commun. 2017, 8, 98. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO Term | Definition | p-Value | No. up- or down Regulated | Total in Category |
|---|---|---|---|---|
| CoCl2 vs. Control | ||||
| Up | ||||
| GO:0007080 | mitotic metaphase plate congression | 9.36 × 10−5 | 5 | 111 |
| GO:0006915 | apoptotic process | 1.68 × 10−4 | 13 | 111 |
| GO:0005829 | cytosol | 9.57 × 10−12 | 66 | 115 |
| GO:0005654 | nucleoplasm | 1.64 × 10−8 | 49 | 115 |
| GO:0005634 | nucleus | 3.94 × 10−8 | 62 | 115 |
| GO:0005874 | microtubule | 2.48 × 10−7 | 13 | 115 |
| GO:0005737 | cytoplasm | 9.53 × 10−6 | 54 | 115 |
| GO:0072562 | blood microparticle | 1.57 × 10−4 | 7 | 115 |
| GO:0070062 | extracellular exosome | 4.06 × 10−4 | 26 | 115 |
| GO:0072686 | mitotic spindle | 0.001097823 | 6 | 115 |
| GO:0000776 | kinetochore | 0.00172263 | 6 | 115 |
| GO:0005871 | kinesin complex | 0.002838309 | 4 | 115 |
| GO:0005515 | protein binding | 6.20 × 10−9 | 102 | 113 |
| GO:0008017 | microtubule binding | 2.60 × 10−5 | 10 | 113 |
| GO:0005524 | ATP binding | 2.47 × 10−4 | 22 | 113 |
| GO:0042802 | identical protein binding | 9.06 × 10−4 | 22 | 113 |
| GO:0005198 | structural molecule activity | 0.001048323 | 7 | 113 |
| GO:0031267 | small GTPase binding | 0.001514117 | 8 | 113 |
| GO:0031625 | ubiquitin protein ligase binding | 0.00230867 | 8 | 113 |
| Down | ||||
| GO:0005654 | nucleoplasm | 1.34 × 10−4 | 34 | 93 |
| GO:0005829 | cytosol | 1.82 × 10−4 | 42 | 93 |
| GO:0005515 | protein binding | 3.32 × 10−5 | 79 | 92 |
| CoCl2 vs. CoCl2+NPC | ||||
| up | ||||
| GO:0031508 | pericentric heterochromatin assembly | 1.43 × 10−4 | 3 | 75 |
| GO:0005829 | cytosol | 4.03 × 10−4 | 34 | 73 |
| GO:0005515 | protein binding | 1.12 × 10−4 | 61 | 70 |
| GO:0031492 | nucleosomal DNA binding | 4.49 × 10−4 | 4 | 70 |
| Down | ||||
| GO:0072562 | blood microparticle | 5.96 × 10−5 | 6 | 60 |
| GO:0070062 | extracellular exosome | 1.11 × 10−4 | 18 | 60 |
| GO:0005737 | cytoplasm | 2.97 × 10−4 | 30 | 60 |
| GO:0005829 | cytosol | 5.41 × 10−4 | 29 | 60 |
| GO:0005634 | nucleus | 9.02 × 10−4 | 30 | 60 |
| GO:0071682 | endocytic vesicle lumen | 0.001199461 | 3 | 60 |
| GO:0005615 | extracellular space | 0.002971796 | 14 | 60 |
| GO:0045095 | keratin filament | 0.003882598 | 4 | 60 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, H.-A.; Park, M.; Lee, H.J.; Duong, V.-A.; Kim, H.-M.; Hwang, D.-Y.; Lee, H.; Lew, H. Unveiling Neuroprotection and Regeneration Mechanisms in Optic Nerve Injury: Insight from Neural Progenitor Cell Therapy with Focus on Vps35 and Syntaxin12. Cells 2023, 12, 2412. https://doi.org/10.3390/cells12192412
Shin H-A, Park M, Lee HJ, Duong V-A, Kim H-M, Hwang D-Y, Lee H, Lew H. Unveiling Neuroprotection and Regeneration Mechanisms in Optic Nerve Injury: Insight from Neural Progenitor Cell Therapy with Focus on Vps35 and Syntaxin12. Cells. 2023; 12(19):2412. https://doi.org/10.3390/cells12192412
Chicago/Turabian StyleShin, Hyun-Ah, Mira Park, Hey Jin Lee, Van-An Duong, Hyun-Mun Kim, Dong-Youn Hwang, Hookeun Lee, and Helen Lew. 2023. "Unveiling Neuroprotection and Regeneration Mechanisms in Optic Nerve Injury: Insight from Neural Progenitor Cell Therapy with Focus on Vps35 and Syntaxin12" Cells 12, no. 19: 2412. https://doi.org/10.3390/cells12192412
APA StyleShin, H.-A., Park, M., Lee, H. J., Duong, V.-A., Kim, H.-M., Hwang, D.-Y., Lee, H., & Lew, H. (2023). Unveiling Neuroprotection and Regeneration Mechanisms in Optic Nerve Injury: Insight from Neural Progenitor Cell Therapy with Focus on Vps35 and Syntaxin12. Cells, 12(19), 2412. https://doi.org/10.3390/cells12192412