

Retinoid X Receptor Activation Prevents Diabetic Retinopathy in Murine Models

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Diabetes Induction
2.3. UAB126 Synthesis
2.4. Preparation of UAB126 Eye Drops
2.5. UAB126 Systemic Administration
2.6. UAB126 Topical Application
2.7. Retinal Explants
2.8. Quantitative Real-Time PCR Analysis (qRT-PCR)
2.9. Tissue Homogenization and Extraction for Mass Spectrometry
2.10. Mass Spectrometry
2.11. Mouse Bone Marrow-Derived Macrophages (BMDM) Generation
2.12. BMDM Activation and Pro-Inflammatory Assessment
2.13. Glucose Uptake Assay
2.14. Metabolic Stress Test (Seahorse Assay)
2.15. Electroretinography (ERG)
2.16. Immunoblotting
2.17. Immunohistochemistry
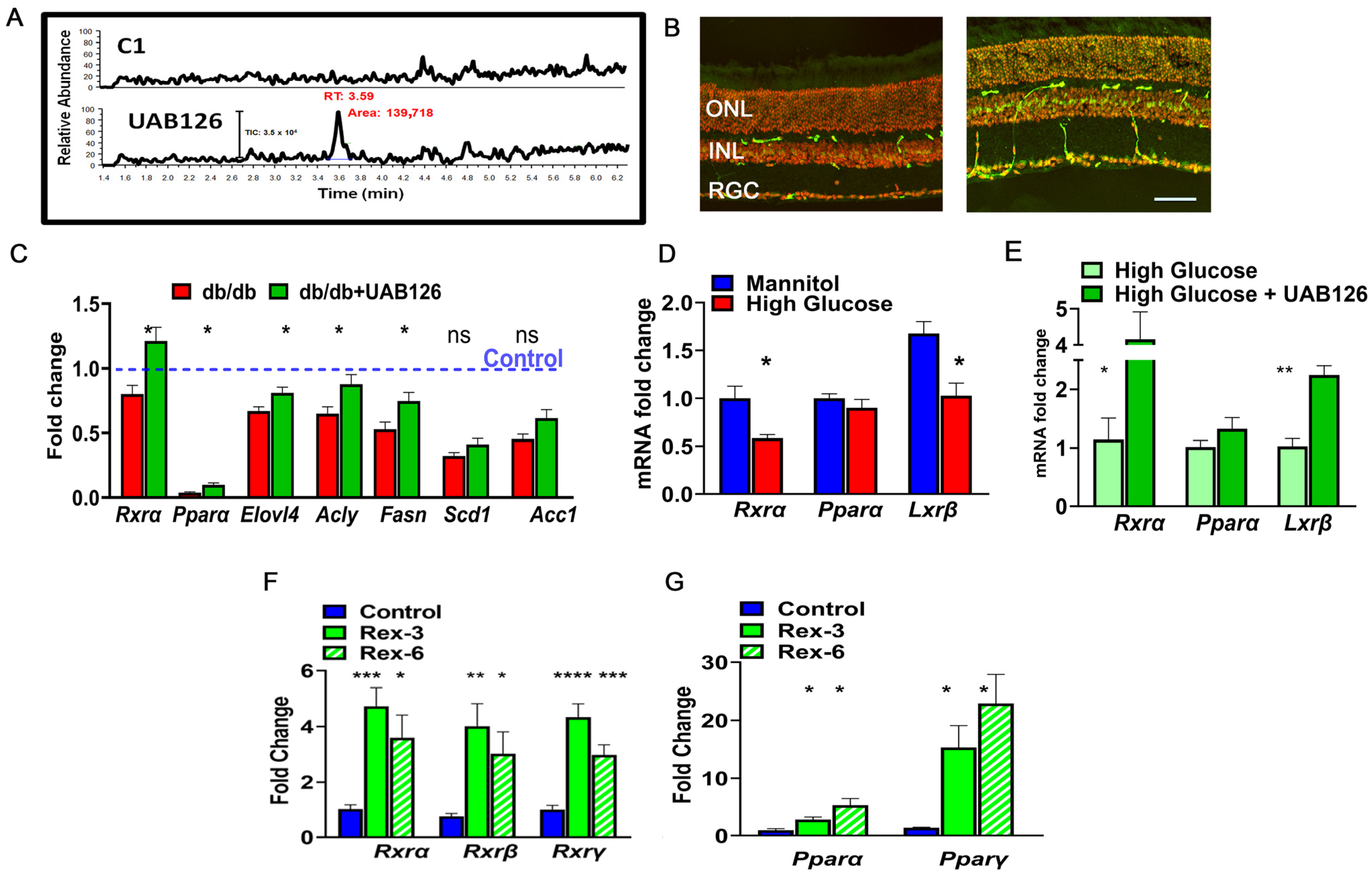
3. Results
3.1. RXR Is Reduced in the Diabetic Retinas of Humans and Mice
3.2. UAB126 Is Present in the Serum and Retina Following Oral Administration
3.3. UAB 126 Treatment Reduces Hyperglycemia in Diabetic Mice
3.4. UAB126 Treatment Improves the Diminished Retinal Function and Alters Cellular Signaling in Diabetic Retina
3.5. UAB126 Regulates Metabolism-Relative Biological Pathways in Diabetic Retinas
3.6. Topical Delivery of UAB 126 Activated RXR in Diabetic Retinas
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, X.K.; Lehmann, J.; Hoffmann, B.; Dawson, M.I.; Cameron, J.; Graupner, G.; Hermann, T.; Tran, P.; Pfahl, M. Homodimer formation of retinoid X receptor induced by 9-cis retinoic acid. Nature 1992, 358, 587–591. [Google Scholar] [CrossRef] [PubMed]
- Osz, J.; Brelivet, Y.; Peluso-Iltis, C.; Cura, V.; Eiler, S.; Ruff, M.; Bourguet, W.; Rochel, N.; Moras, D. Structural basis for a molecular allosteric control mechanism of cofactor binding to nuclear receptors. Proc. Natl. Acad. Sci. USA 2012, 109, E588–E594. [Google Scholar] [CrossRef]
- Chambon, P. A decade of molecular biology of retinoic acid receptors. FASEB J. 1996, 10, 940–954. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Chandra, V.; Rastinejad, F. Retinoic acid actions through mammalian nuclear receptors. Chem. Rev. 2014, 114, 233–254. [Google Scholar] [CrossRef] [PubMed]
- Gocek, E.; Baurska, H.; Marchwicka, A.; Marcinkowska, E. Regulation of Leukemic Cell Differentiation through the Vitamin D Receptor at the Levels of Intracellular Signal Transduction, Gene Transcription, and Protein Trafficking and Stability. Leuk. Res. Treat. 2012, 2012, 713243. [Google Scholar] [CrossRef][Green Version]
- Yasmin, R.; Williams, R.M.; Xu, M.; Noy, N. Nuclear import of the retinoid X receptor, the vitamin D receptor, and their mutual heterodimer. J. Biol. Chem. 2005, 280, 40152–40160. [Google Scholar] [CrossRef] [PubMed]
- Szeles, L.; Poliska, S.; Nagy, G.; Szatmari, I.; Szanto, A.; Pap, A.; Lindstedt, M.; Santegoets, S.J.; Ruhl, R.; Dezso, B.; et al. Research resource: Transcriptome profiling of genes regulated by RXR and its permissive and nonpermissive partners in differentiating monocyte-derived dendritic cells. Mol. Endocrinol. 2010, 24, 2218–2231. [Google Scholar] [CrossRef]
- Kubickova, B.; Martinkova, S.; Bohaciakova, D.; Nezvedova, M.; Liu, R.; Brozman, O.; Spacil, Z.; Hilscherova, K. Effects of all-trans and 9-cis retinoic acid on differentiating human neural stem cells in vitro. Toxicology 2023, 487, 153461. [Google Scholar] [CrossRef]
- Sugawara, A.; Yen, P.M.; Qi, Y.; Lechan, R.M.; Chin, W.W. Isoform-specific retinoid-X receptor (RXR) antibodies detect differential expression of RXR proteins in the pituitary gland. Endocrinology 1995, 136, 1766–1774. [Google Scholar] [CrossRef]
- Volonte, Y.A.; Ayala-Pena, V.B.; Vallese-Maurizi, H.; Garelli, A.; Rotstein, N.P.; Politi, L.E.; German, O.L. Retinoid X receptor activation promotes photoreceptor survival and modulates the inflammatory response in a mouse model of retinitis pigmentosa. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 119098. [Google Scholar] [CrossRef]
- Roberts, M.R.; Hendrickson, A.; McGuire, C.R.; Reh, T.A. Retinoid X receptor (gamma) is necessary to establish the S-opsin gradient in cone photoreceptors of the developing mouse retina. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2897–2904. [Google Scholar] [CrossRef] [PubMed]
- UniProt, C. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Sharma, S.; Shen, T.; Chitranshi, N.; Gupta, V.; Basavarajappa, D.; Sarkar, S.; Mirzaei, M.; You, Y.; Krezel, W.; Graham, S.L.; et al. Correction to: Retinoid X Receptor: Cellular and Biochemical Roles of Nuclear Receptor with a Focus on Neuropathological Involvement. Mol. Neurobiol. 2022, 59, 2051. [Google Scholar] [CrossRef] [PubMed]
- Wietrzych-Schindler, M.; Szyszka-Niagolov, M.; Ohta, K.; Endo, Y.; Perez, E.; de Lera, A.R.; Chambon, P.; Krezel, W. Retinoid x receptor gamma is implicated in docosahexaenoic acid modulation of despair behaviors and working memory in mice. Biol. Psychiatry 2011, 69, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, R.; Takahashi, D.; Watanabe, M.; Nakatani, S.; Takamura, Y.; Kurosaki, Y.; Kakuta, H.; Hase, K. A Retinoid X Receptor Agonist Directed to the Large Intestine Ameliorates T-Cell-Mediated Colitis in Mice. Front. Pharmacol. 2021, 12, 715752. [Google Scholar] [CrossRef]
- Morichika, D.; Miyahara, N.; Fujii, U.; Taniguchi, A.; Oda, N.; Senoo, S.; Kataoka, M.; Tanimoto, M.; Kakuta, H.; Kiura, K.; et al. A retinoid X receptor partial agonist attenuates pulmonary emphysema and airway inflammation. Respir. Res. 2019, 20, 2. [Google Scholar] [CrossRef]
- Li, Y.; Xing, Q.; Wei, Y.; Zhao, L.; Zhang, P.; Han, X.; Wang, J. Activation of RXR by bexarotene inhibits inflammatory conditions in human rheumatoid arthritis fibroblast-like synoviocytes. Int. J. Mol. Med. 2019, 44, 1963–1970. [Google Scholar] [CrossRef]
- Nunez, V.; Alameda, D.; Rico, D.; Mota, R.; Gonzalo, P.; Cedenilla, M.; Fischer, T.; Bosca, L.; Glass, C.K.; Arroyo, A.G.; et al. Retinoid X receptor alpha controls innate inflammatory responses through the up-regulation of chemokine expression. Proc. Natl. Acad. Sci. USA 2010, 107, 10626–10631. [Google Scholar] [CrossRef]
- Roszer, T.; Menendez-Gutierrez, M.P.; Cedenilla, M.; Ricote, M. Retinoid X receptors in macrophage biology. Trends Endocrinol. Metab. 2013, 24, 460–468. [Google Scholar] [CrossRef]
- Lalloyer, F.; Fievet, C.; Lestavel, S.; Torpier, G.; van der Veen, J.; Touche, V.; Bultel, S.; Yous, S.; Kuipers, F.; Paumelle, R.; et al. The RXR agonist bexarotene improves cholesterol homeostasis and inhibits atherosclerosis progression in a mouse model of mixed dyslipidemia. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 2731–2737. [Google Scholar] [CrossRef]
- Grubbs, C.J.; Hill, D.L.; Bland, K.I.; Beenken, S.W.; Lin, T.H.; Eto, I.; Atigadda, V.R.; Vines, K.K.; Brouillette, W.J.; Muccio, D.D. 9cUAB30, an RXR specific retinoid, and/or tamoxifen in the prevention of methylnitrosourea-induced mammary cancers. Cancer Lett. 2003, 201, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Kim, T.; Kim, H.S.; Young, M.E.; Muccio, D.D.; Atigadda, V.R.; Blum, S.I.; Tse, H.M.; Habegger, K.M.; Bhatnagar, S.; et al. A Small Molecule, UAB126, Reverses Diet-Induced Obesity and its Associated Metabolic Disorders. Diabetes 2020, 69, 2003–2016. [Google Scholar] [CrossRef] [PubMed]
- Melo da Cunha, J.D.S.; Alfredo, T.M.; Dos Santos, J.M.; Alves Junior, V.V.; Rabelo, L.A.; Lima, E.S.; Boleti, A.P.A.; Carollo, C.A.; Dos Santos, E.L.; de Picoli Souza, K. Antioxidant, antihyperglycemic, and antidiabetic activity of Apis mellifera bee tea. PLoS ONE 2018, 13, e0197071. [Google Scholar] [CrossRef] [PubMed]
- Singh, Y.; Meher, J.G.; Raval, K.; Khan, F.A.; Chaurasia, M.; Jain, N.K.; Chourasia, M.K. Nanoemulsion: Concepts, development and applications in drug delivery. J. Control. Release 2017, 252, 28–49. [Google Scholar] [CrossRef]
- Fernandes, A.R.; Sanchez-Lopez, E.; Santos, T.D.; Garcia, M.L.; Silva, A.M.; Souto, E.B. Development and Characterization of Nanoemulsions for Ophthalmic Applications: Role of Cationic Surfactants. Materials 2021, 14, 7541. [Google Scholar] [CrossRef]
- Amend, S.R.; Valkenburg, K.C.; Pienta, K.J. Murine Hind Limb Long Bone Dissection and Bone Marrow Isolation. J. Vis. Exp. 2016, 110, e53936. [Google Scholar] [CrossRef]
- Saltykova, I.V.; Elahi, A.; Pitale, P.M.; Gorbatyuk, O.S.; Athar, M.; Gorbatyuk, M.S. Tribbles homolog 3-mediated targeting the AKT/mTOR axis in mice with retinal degeneration. Cell Death Dis. 2021, 12, 664. [Google Scholar] [CrossRef]
- Hernandez, C.; Bogdanov, P.; Corraliza, L.; Garcia-Ramirez, M.; Sola-Adell, C.; Arranz, J.A.; Arroba, A.I.; Valverde, A.M.; Simo, R. Topical Administration of GLP-1 Receptor Agonists Prevents Retinal Neurodegeneration in Experimental Diabetes. Diabetes 2016, 65, 172–187. [Google Scholar] [CrossRef]
- Hernandez, C.; Bogdanov, P.; Sola-Adell, C.; Sampedro, J.; Valeri, M.; Genis, X.; Simo-Servat, O.; Garcia-Ramirez, M.; Simo, R. Topical administration of DPP-IV inhibitors prevents retinal neurodegeneration in experimental diabetes. Diabetologia 2017, 60, 2285–2298. [Google Scholar] [CrossRef]
- Atigadda, V.R.; Kashyap, M.P.; Yang, Z.; Chattopadhyay, D.; Melo, N.; Sinha, R.; Belyaeva, O.V.; Chou, C.F.; Chang, P.L.; Kedishvili, N.Y.; et al. Conformationally Defined Rexinoids for the Prevention of Inflammation and Nonmelanoma Skin Cancers. J. Med. Chem. 2022, 65, 14409–14423. [Google Scholar] [CrossRef]
- Zeng, H.; Qi, X.; Xu, X.; Wu, Y. TAB1 regulates glycolysis and activation of macrophages in diabetic nephropathy. Inflamm. Res. 2020, 69, 1215–1234. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Boytard, L.; Hadi, T.; Koelwyn, G.; Simon, R.; Ouimet, M.; Seifert, L.; Spiro, W.; Yan, B.; Hutchison, S.; et al. Enhanced glycolysis and HIF-1alpha activation in adipose tissue macrophages sustains local and systemic interleukin-1beta production in obesity. Sci. Rep. 2020, 10, 5555. [Google Scholar] [CrossRef] [PubMed]
- Serbulea, V.; Upchurch, C.M.; Schappe, M.S.; Voigt, P.; DeWeese, D.E.; Desai, B.N.; Meher, A.K.; Leitinger, N. Macrophage phenotype and bioenergetics are controlled by oxidized phospholipids identified in lean and obese adipose tissue. Proc. Natl. Acad. Sci. USA 2018, 115, E6254–E6263. [Google Scholar] [CrossRef]
- Matsuura, Y.; Shimizu-Albergine, M.; Barnhart, S.; Kramer, F.; Hsu, C.C.; Kothari, V.; Tang, J.; Gharib, S.A.; Kanter, J.E.; Abel, E.D.; et al. Diabetes Suppresses Glucose Uptake and Glycolysis in Macrophages. Circ. Res. 2022, 130, 779–781. [Google Scholar] [CrossRef]
- Pavlou, S.; Lindsay, J.; Ingram, R.; Xu, H.; Chen, M. Sustained high glucose exposure sensitizes macrophage responses to cytokine stimuli but reduces their phagocytic activity. BMC Immunol. 2018, 19, 24. [Google Scholar] [CrossRef]
- Viola, A.; Munari, F.; Sanchez-Rodriguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef]
- Cantero-Navarro, E.; Rayego-Mateos, S.; Orejudo, M.; Tejedor-Santamaria, L.; Tejera-Munoz, A.; Sanz, A.B.; Marquez-Exposito, L.; Marchant, V.; Santos-Sanchez, L.; Egido, J.; et al. Role of Macrophages and Related Cytokines in Kidney Disease. Front. Med. 2021, 8, 688060. [Google Scholar] [CrossRef]
- Cesario, R.M.; Klausing, K.; Razzaghi, H.; Crombie, D.; Rungta, D.; Heyman, R.A.; Lala, D.S. The rexinoid LG100754 is a novel RXR:PPARgamma agonist and decreases glucose levels in vivo. Mol. Endocrinol. 2001, 15, 1360–1369. [Google Scholar] [CrossRef][Green Version]
- Rajala, A.; Teel, K.; Bhat, M.A.; Batushansky, A.; Griffin, T.M.; Purcell, L.; Rajala, R.V.S. Insulin-like growth factor 1 receptor mediates photoreceptor neuroprotection. Cell Death Dis. 2022, 13, 613. [Google Scholar] [CrossRef]
- Kubota, S.; Ozawa, Y.; Kurihara, T.; Sasaki, M.; Yuki, K.; Miyake, S.; Noda, K.; Ishida, S.; Tsubota, K. Roles of AMP-activated protein kinase in diabetes-induced retinal inflammation. Investig. Ophthalmol. Vis. Sci. 2011, 52, 9142–9148. [Google Scholar] [CrossRef]
- Song, S.; Bao, S.; Zhang, C.; Zhang, J.; Lv, J.; Li, X.; Chudhary, M.; Ren, X.; Kong, L. Stimulation of AMPK Prevents Diabetes-Induced Photoreceptor Cell Degeneration. Oxid. Med. Cell. Longev. 2021, 2021, 5587340. [Google Scholar] [CrossRef] [PubMed]
- Freemark, M.; Bursey, D. The effects of metformin on body mass index and glucose tolerance in obese adolescents with fasting hyperinsulinemia and a family history of type 2 diabetes. Pediatrics 2001, 107, E55. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Tikhonenko, M.; Bozack, S.N.; Lydic, T.A.; Yan, L.; Panchy, N.L.; McSorley, K.M.; Faber, M.S.; Yan, Y.; Boulton, M.E.; et al. Changes in the daily rhythm of lipid metabolism in the diabetic retina. PLoS ONE 2014, 9, e95028. [Google Scholar] [CrossRef] [PubMed]
- Tikhonenko, M.; Lydic, T.A.; Wang, Y.; Chen, W.; Opreanu, M.; Sochacki, A.; McSorley, K.M.; Renis, R.L.; Kern, T.; Jump, D.B.; et al. Remodeling of retinal Fatty acids in an animal model of diabetes: A decrease in long-chain polyunsaturated fatty acids is associated with a decrease in fatty acid elongases Elovl2 and Elovl4. Diabetes 2010, 59, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Fort, P.E.; Rajendiran, T.M.; Soni, T.; Byun, J.; Shan, Y.; Looker, H.C.; Nelson, R.G.; Kretzler, M.; Michailidis, G.; Roger, J.E.; et al. Diminished retinal complex lipid synthesis and impaired fatty acid beta-oxidation associated with human diabetic retinopathy. JCI Insight 2021, 6, e152109. [Google Scholar] [CrossRef]
- Yuan, T.; Dong, L.; Pearsall, E.A.; Zhou, K.; Cheng, R.; Ma, J.X. The Protective Role of Microglial PPARalpha in Diabetic Retinal Neurodegeneration and Neurovascular Dysfunction. Cells 2022, 11, 3869. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dorofeeva, I.; Zhylkibayev, A.; Saltykova, I.V.; Atigadda, V.; Adhikari, B.; Gorbatyuk, O.S.; Grant, M.B.; Gorbatyuk, M.S. Retinoid X Receptor Activation Prevents Diabetic Retinopathy in Murine Models. Cells 2023, 12, 2361. https://doi.org/10.3390/cells12192361
Dorofeeva I, Zhylkibayev A, Saltykova IV, Atigadda V, Adhikari B, Gorbatyuk OS, Grant MB, Gorbatyuk MS. Retinoid X Receptor Activation Prevents Diabetic Retinopathy in Murine Models. Cells. 2023; 12(19):2361. https://doi.org/10.3390/cells12192361
Chicago/Turabian StyleDorofeeva, Iuliia, Assylbek Zhylkibayev, Irina V. Saltykova, Venkatram Atigadda, Bibek Adhikari, Oleg S. Gorbatyuk, Maria B. Grant, and Marina S. Gorbatyuk. 2023. "Retinoid X Receptor Activation Prevents Diabetic Retinopathy in Murine Models" Cells 12, no. 19: 2361. https://doi.org/10.3390/cells12192361
APA StyleDorofeeva, I., Zhylkibayev, A., Saltykova, I. V., Atigadda, V., Adhikari, B., Gorbatyuk, O. S., Grant, M. B., & Gorbatyuk, M. S. (2023). Retinoid X Receptor Activation Prevents Diabetic Retinopathy in Murine Models. Cells, 12(19), 2361. https://doi.org/10.3390/cells12192361