
Ubiquitin Engineering for Interrogating the Ubiquitin–Proteasome System and Novel Therapeutic Strategies

Abstract
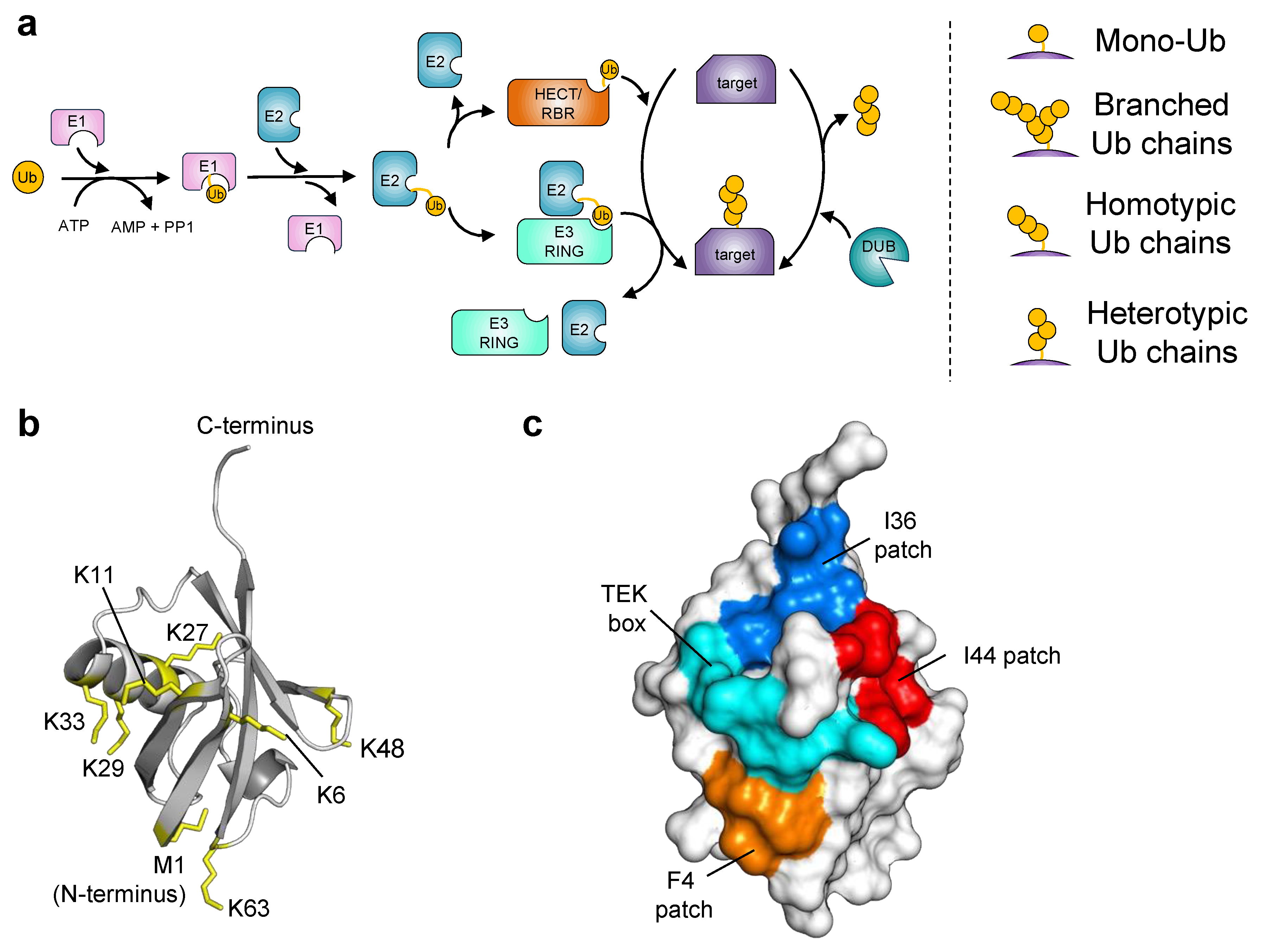
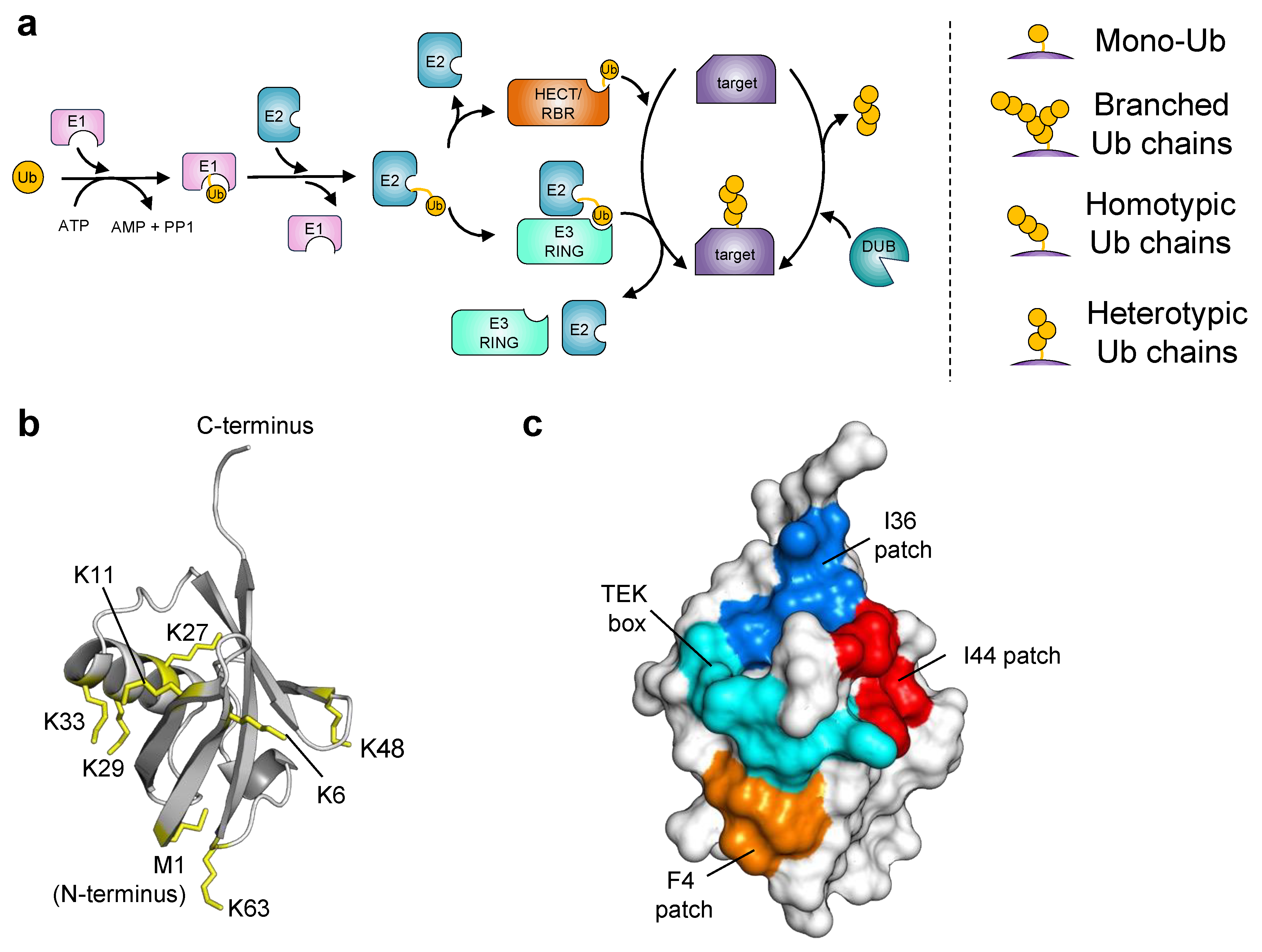
:1. Introduction
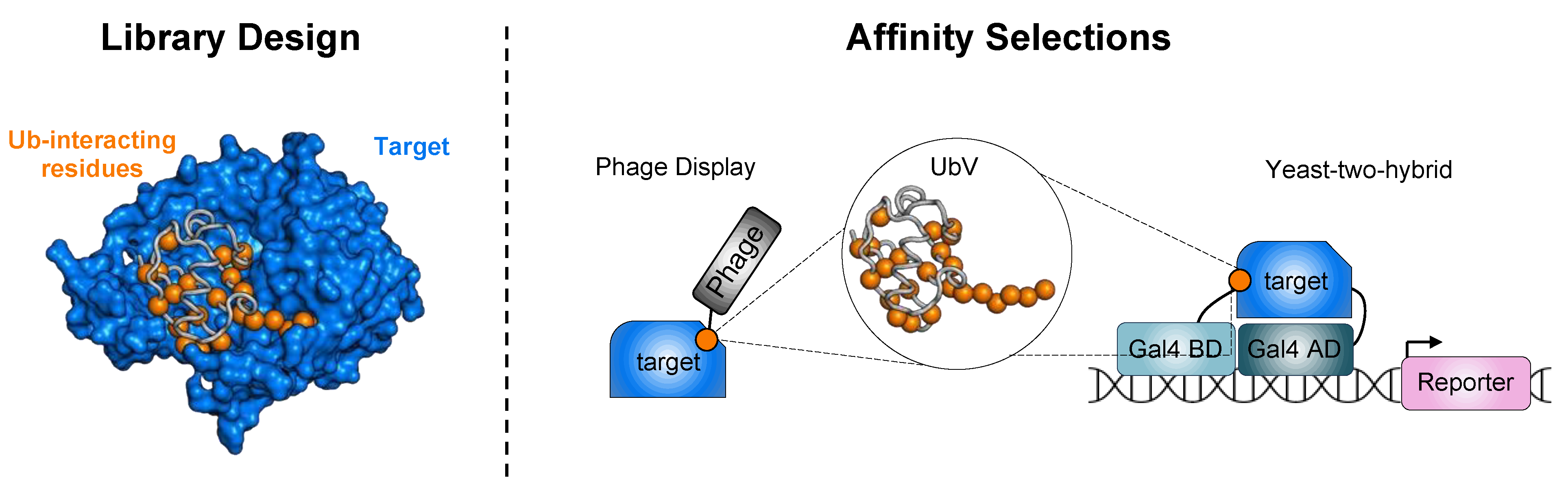
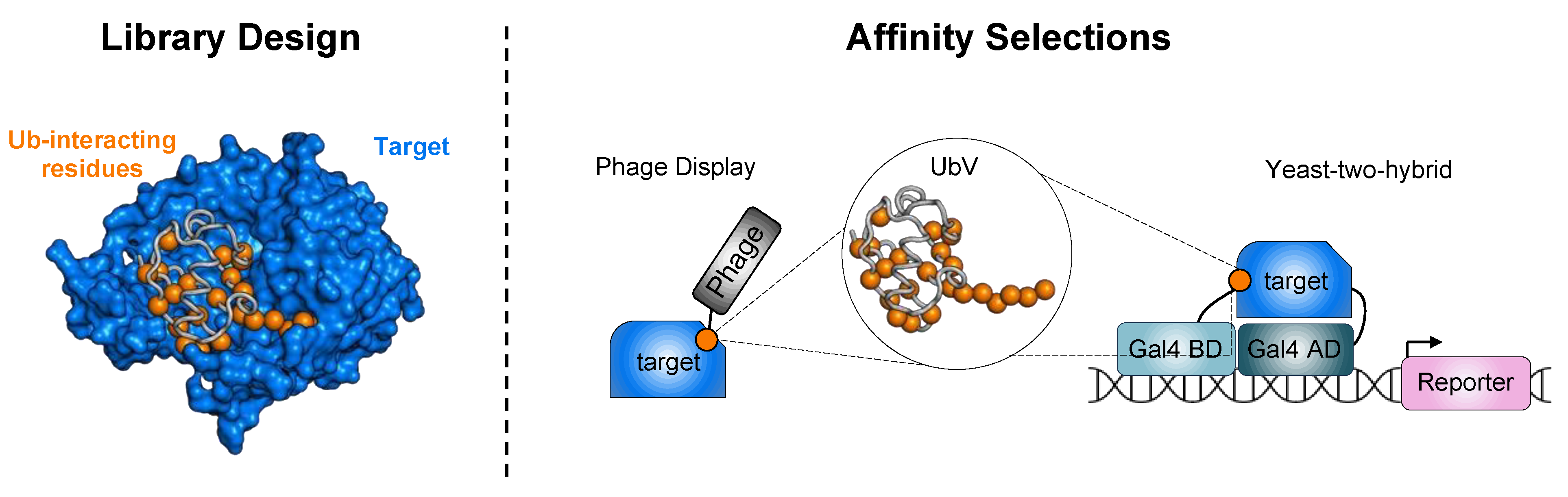
2. Ubiquitin Engineering
3. Targeting Enzymes of the Ubiquitin–Proteasome System
3.1. UbVs Targeting E2 Ubiquitin-Conjugating Enzymes
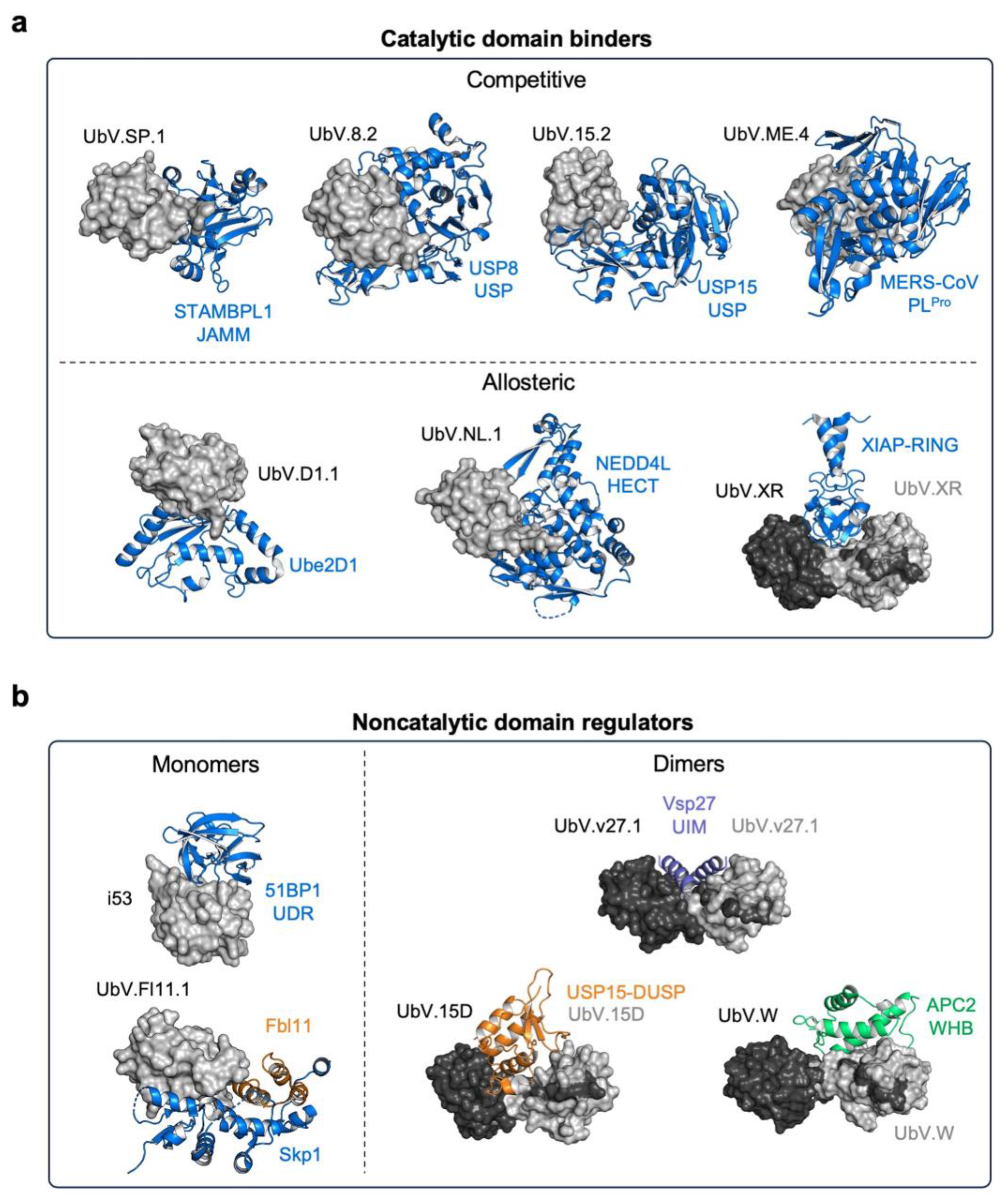
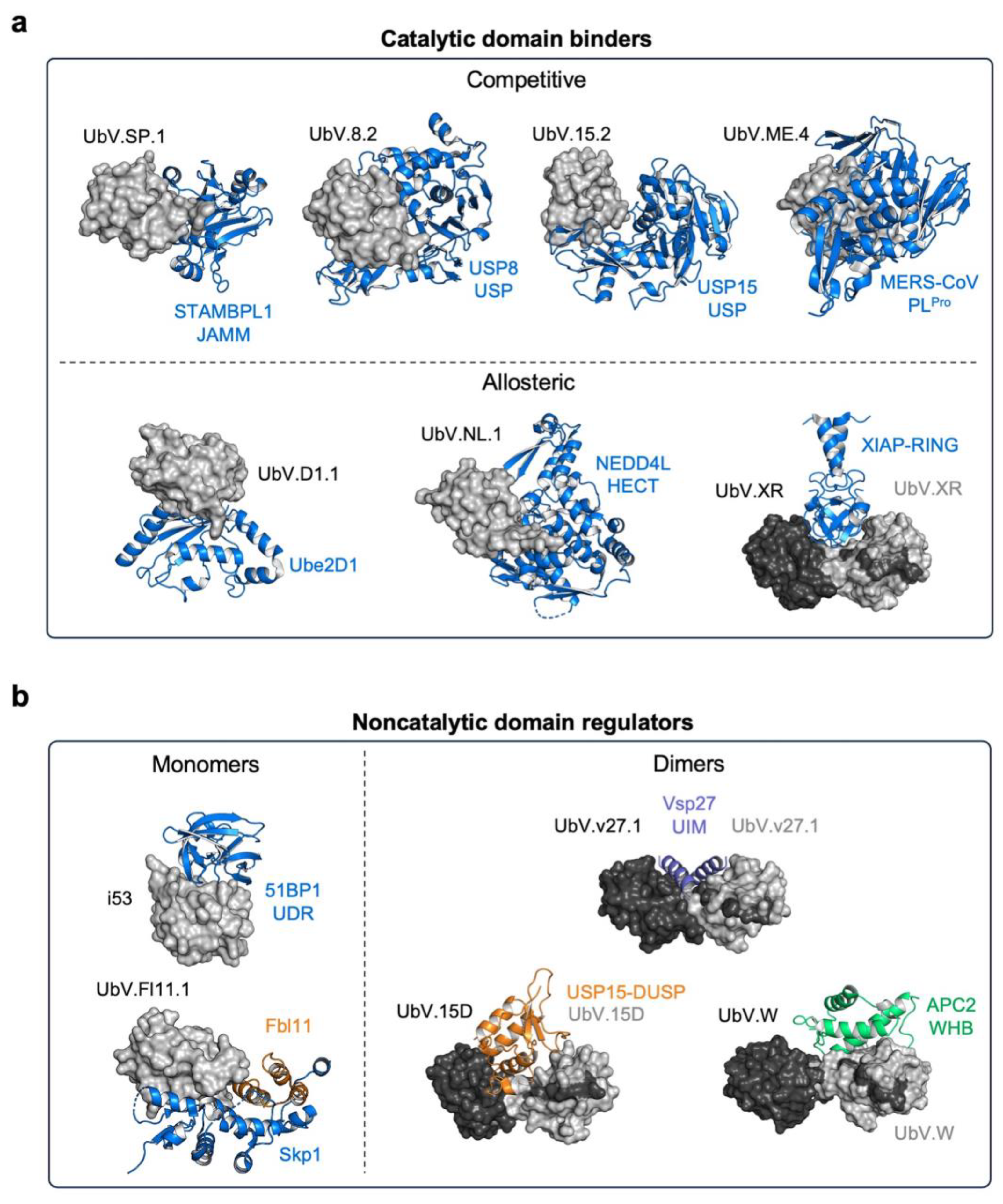
3.2. UbVs Targeting E3 Ligases
3.2.1. UbVs for HECT-E3 Ligases
3.2.2. UbVs for U-Box E3 Ligases
3.2.3. UbVs for RING-E3 Ligases
3.3. UbVs Targeting Deubiquitinating Enzymes
3.3.1. UbV Inhibitors of Ubiquitin-Specific Proteases
3.3.2. UbV Inhibitors of the JAMM Family
3.3.3. UbVs as Inhibitors of Viral DUBs
3.3.4. UbVs as Probes of DUB Activity
3.3.5. UbVs Targeting Noncatalytic Protein Domains in the UPS
4. Targeting Non-UPS Proteins with UbVs
5. UbV-Mediated Targeted Protein Degradation
6. Conclusions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Target Domain | Protein ID | UbV Isolation Method | UbV ID | Bound Site | Function | Reference |
|---|---|---|---|---|---|---|---|
| E2 | Catalytic | Ube2k | Phage display | UbV.k.1 | UBC | Inhibitor | [31] |
| Catalytic | Ube2k | Phage display | UbV.k.2 | UBC | Inhibitor | [31] | |
| Catalytic | Ube2D1 | Phage display | UbV.D1.1 | backside | Inhibitor | [33] | |
| Catalytic | Ube2V1 | Phage display | UbV.V1.1 | backside | Inhibitor | [33] | |
| Catalytic | Ube2G1 | Phage display | UbV.G1.1 | backside | Inhibitor | [33] | |
| E3 | HECT | WWP1 | Phage display | UbV.P1.1 | E2-site | Inhibitor | [40] |
| HECT | WWP2 | Phage display | UbV.P2.3 | ND | Activator | [40] | |
| HECT | ITCH | Phage display | UbV.IT.2 | E2-site | Inhibitor | [40] | |
| HECT | S. cerevisiae Rsp5 | Phage display | UbV.R5.4 | N-lobe exosite | Activator | [40] | |
| HECT | NEDD4 | Phage display | UbV.N4.4 | N-lobe exosite | Activator | [40] | |
| HECT | NEDD4 | Phage display | UbV.N.2 | ND | Activator | [41] | |
| HECT | NEDD4L | Phage display | UbV.NL.1 | ND | Activator | [40] | |
| HECT | NEDD4L | Phage display | UbV.NL.3 | N-lobe exosite | Inhibitor | [40] | |
| HECT | HACE1 | Phage display | UbV.HA.3 | ND | Inhibitor | [40] | |
| HECT | SMURF2 | Phage display | UbV.S2.5 | ND | Inhibitor | [40] | |
| RING | pCBL | Phage display | UbV.pCBL | E2-site | Inhibitor | [43] | |
| RING | XIAP | Phage display | UbV.XR | RING/donor Ub | Activator | [43] | |
| U-Box | UBE4B | Phage display | UbV.E4B | E2 site | Inhibitor | [43] | |
| RCL | SCF | Phage display | UbV.Fw7.5 | F-box interface | Inhibitor | [46] | |
| RCL | SCF | Phage display | UbV.Fw11.2 | F-box interface | Inhibitor | [46] | |
| RCL | SCF | Phage display | UbV.Fl11.1 | ND | Inhibitor | [47] | |
| RCL | SCF | Phage display | UbV.L1.1 | ND | Inhibitor | [47] | |
| RCL | SCF | Phage display | UbVO11.1 | ND | Inhibitor | [47] | |
| RING | APC11 | Phage display | UbV.R | E2-site | Inhibitor | [93] | |
| APC/C | APC2 | Phage display | UbV.W | Ub exosite | Inhibitor | [93] | |
| DUBs | Catalytic | USP2a | Computational/phage display/Y2H | UbV2.3 and UbV2.1 | ND | Inhibitor | [41] |
| OTU | OTUB1 | Phage display | UbV.B1.1 | Ub-distal site | Inhibitor | [41] | |
| JAMM | STAMBP | Phage display | UbV.SP.1 | Ins-1 | Inhibitor | [74] | |
| JAMM | STAMBP | Phage display | UbV.SP.3 | ND | Inhibitor | [74] | |
| UCH | UCHL1 | Computational/ rational design | UbVT9F/T66K | ND | Inhibitor/ABP | [79] | |
| UCH | UCHL3 | Computational/ rational design | UbVQ40V/T66K/V70F | ND | Inhibitor/ABP | [81] | |
| Catalytic | USP8 | Phage display | UbV.8.2 | Ub-binding site | Inhibitor | [41] | |
| Catalytic | USP21 | Phage display | UbV21.4 | ND | Inhibitor | [41] | |
| Catalytic | USP7 | Phage display | Ub7Ub25 | ND | Inhibitor | [59] | |
| Catalytic | USP7 | Phage display | Ub7Ub25.2540 | ND | Inhibitor | [59] | |
| Catalytic | USP7 | Computational/phage display | UbV.7.2 and M6 | Ub-binding site | Inhibitor/ABP | [60,82] | |
| Catalytic | USP10 | Phage display | UbV10.1 | ND | Inhibitor | [60] | |
| Catalytic | USP2 | Y2H | UbV2.6 | Ub-binding site | Inhibitor | [63] | |
| Catalytic | USP15 | Phage display | UbV.15.1 | Ub-binding site | Inhibitor | [69] | |
| Catalytic | USP15 | Phage display | UbV.15.1/D (dimer) | Ub-binding site and DUSP | Inhibitor | [69] | |
| Catalytic | USP16 | Computational | M20 | ND | Inhibitor/ABP | [83] | |
| Catalytic | USP14 | Phage display | U14Ub14 | Ub-binding site | Inhibitor | [110] | |
| vDUBs | OTU | CCHFV | Phage display | UbV.CC.4 | Ub-binding site | Inhibitor | [76] |
| PLpro | MERS-CoV | Computational/phage display | UbV.ME.4 | Ub-binding site | Inhibitor | [76] | |
| PLpro | SARS-CoV-2 | Phage display | UbV.CV2.1a | Ub-binding site | Inhibitor | [78] | |
| UBDs | UIM | S. cerevisiae Vsp27 | Phage display | UbV.v27.1 | Ub-binding site | Inhibitor | [85,86] |
| UIM | ANKRD13D | Phage display | UbV.ANKRD13D.4 | Ub-binding site | Binding | [87] | |
| UIM | USP28 | Phage display | UbV.USP28 | ND | Inhibitor | [87] | |
| UIM | D. rerio USP37 | Phage display | UbV.UIM | ND | Inhibitor | [88] | |
| DUSP | USP11 | Phage display | UbV.11D.2 | ND | Inhibitor | [92] | |
| DUSP | USP15 | Phage display | UbV.15D.1 | DUSP | Inhibitor | [92] | |
| DUSP | USP20 | Phage display | UbV.20D2 | ND | Inhibitor | [92] | |
| Adaptor protein | SH2 | Grb2 | Phage display | UbV.G2.1 | ND | Binding | [23] |
| SH2 | Grb2 | Phage display | UbV.G2.2 | pTyr-binding site | Inhibitor | [23] | |
| SH2 | Grb2 | Phage display | UbV.G2.2/1 (dimer) | pTyr-binding site and SH2 | Inhibitor | [23] | |
| EGFR | Ecto-domain | Her3 | Phage display | UbV.H3.2 | Diverse from neuregulin site | Binding | [23] |
| Fibronectin | ED-B | Oncofetal fibronectin | Phage/ribosome display | Affilin-77405 | ND | Binding | [96] |
| Tumor suppressor | UDR | 53BP1 | Phage display | i53 | UDR domain | Gene editing/ targeted degradation | [94,95,101] |
7. Future Challenges in UbV Technology
- Specificity and selectivity: Ensuring high specificity and selectivity for target proteins is crucial for therapeutic applications of UbVs. As the number of identified UbVs increases, it becomes essential to address potential off-target effects and enhance the ability to discriminate between homologous proteins.
- UbV discovery strategies: While in vitro affinity selection has yielded successful protein-based inhibitors or activators of the UPS, some UbVs, like those targeting USP2, may lack functional activity within the cellular context. Exploring alternative strategies for isolating intracellularly functional UbVs can expedite discovery and enhance their therapeutic and research potential.
- Modulating proteasome activity: Small molecules like bortezomib and carfilzomib have demonstrated efficacy in inhibiting proteasome activity, leading to their approval for the treatment of certain cancers. Similarly, UbVs targeting specific proteasome subunits or regulatory proteins could offer a novel approach for modulating proteasome function with potentially greater selectivity and fewer off-target effects compared to traditional small molecules.
- Intracellular delivery: Efficient intracellular delivery of UbVs is essential for their therapeutic potential. Overcoming cellular barriers requires the development of novel delivery strategies such as novel CPPs, capable of escaping endosomes. Additionally, safer AAVs or lipid nanoparticles encapsulating the mRNA-encoding UbVs could unlock their translational potential.
- In vivo models: Establishing robust in vivo models that accurately represent the pathogenic activity of the UPS will be crucial for validating the therapeutic efficacy of UbVs.
- Addressing these challenges will unlock the full translational potential of UbVs and contribute to advancing our understanding of the UPS, leading to new opportunities for therapeutic interventions in various diseases.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lecker, S.H.; Goldberg, A.L.; Mitch, W.E. Protein Degradation by the Ubiquitin-Proteasome Pathway in Normal and Disease States. J. Am. Soc. Nephrol. 2006, 17, 1807–1819. [Google Scholar] [CrossRef] [PubMed]
- Clague, M.J.; Urbé, S. Ubiquitin: Same Molecule, Different Degradation Pathways. Cell 2010, 143, 682–685. [Google Scholar] [CrossRef]
- Swatek, K.N.; Komander, D. Ubiquitin Modifications. Cell Res. 2016, 26, 399–422. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Smits, A.H.; van Tilburg, G.B.A.; Jansen, P.W.T.C.; Makowski, M.M.; Ovaa, H.; Vermeulen, M. An Interaction Landscape of Ubiquitin Signaling. Mol. Cell 2017, 65, 941–955.e8. [Google Scholar] [CrossRef] [PubMed]
- Veggiani, G.; Sidhu, S.S. Peptides Meet Ubiquitin: Simple Interactions Regulating Complex Cell Signaling. Pept. Sci. 2019, 111, e24091. [Google Scholar] [CrossRef]
- Seet, B.T.; Dikic, I.; Zhou, M.-M.; Pawson, T. Reading Protein Modifications with Interaction Domains. Nat. Rev. Mol. Cell Biol. 2006, 7, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Sims, J.J.; Cohen, R.E. Linkage-Specific Avidity Defines the Lysine 63-Linked Polyubiquitin Binding Preference of Rap80. Mol. Cell 2009, 33, 775–783. [Google Scholar] [CrossRef]
- Sato, Y.; Yoshikawa, A.; Mimura, H.; Yamashita, M.; Yamagata, A.; Fukai, S. Structural Basis for Specific Recognition of Lys 63-Linked Polyubiquitin Chains by Tandem UIMs of RAP80. EMBO J. 2009, 28, 2461–2468. [Google Scholar] [CrossRef]
- Kliza, K.; Husnjak, K. Resolving the Complexity of Ubiquitin Networks. Front. Mol. Biosci. 2020, 7, 21. [Google Scholar] [CrossRef]
- Deng, L.; Meng, T.; Chen, L.; Wei, W.; Wang, P. The Role of Ubiquitination in Tumorigenesis and Targeted Drug Discovery. Signal Transduct. Target. Ther. 2020, 5, 11. [Google Scholar] [CrossRef]
- Shen, M.; Schmitt, S.; Buac, D.; Dou, Q.P. Targeting the Ubiquitin-Proteasome System for Cancer Therapy. Expert Opin. Ther. Targets 2013, 17, 1091–1108. [Google Scholar] [CrossRef]
- Edelmann, M.J.; Nicholson, B.; Kessler, B.M. Pharmacological Targets in the Ubiquitin System Offer New Ways of Treating Cancer, Neurodegenerative Disorders and Infectious Diseases. Expert Rev. Mol. Med. 2011, 13, e35. [Google Scholar] [CrossRef]
- Sasso, J.M.; Tenchov, R.; Wang, D.; Johnson, L.S.; Wang, X.; Zhou, Q.A. Molecular Glues: The Adhesive Connecting Targeted Protein Degradation to the Clinic. Biochemistry 2022, 62, 601–623. [Google Scholar] [CrossRef]
- Békés, M.; Langley, D.R.; Crews, C.M. PROTAC Targeted Protein Degraders: The Past Is Prologue. Nat. Rev. Drug Discov. 2022, 21, 181–200. [Google Scholar] [CrossRef] [PubMed]
- Manasanch, E.E.; Orlowski, R.Z. Proteasome Inhibitors in Cancer Therapy. Nat. Rev. Clin. Oncol. 2017, 14, 417–433. [Google Scholar] [CrossRef] [PubMed]
- Fricker, L.D. Proteasome Inhibitor Drugs. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 457–476. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, N.; Zhang, W.; Cheng, X.; Yan, Z.; Shao, G.; Wang, X.; Wang, R.; Fu, C. Therapeutic Peptides: Current Applications and Future Directions. Signal Transduct. Target. Ther. 2022, 7, 48. [Google Scholar] [CrossRef]
- La Manna, S.; Di Natale, C.; Florio, D.; Marasco, D. Peptides as Therapeutic Agents for Inflammatory-Related Diseases. Int. J. Mol. Sci. 2018, 19, 2714. [Google Scholar] [CrossRef]
- Owens, B. Faster, Deeper, Smaller—The Rise of Antibody-like Scaffolds. Nat. Biotechnol. 2017, 35, 602–603. [Google Scholar] [CrossRef]
- Zhang, W.; Sidhu, S.S. Generating Intracellular Modulators of E3 Ligases and Deubiquitinases from Phage-Displayed Ubiquitin Variant Libraries. Methods Mol. Biol. 2018, 1844, 101–119. [Google Scholar] [CrossRef]
- Ibarra-Molero, B.; Loladze, V.V.; Makhatadze, G.I.; Sanchez-Ruiz, J.M. Thermal versus Guanidine-Induced Unfolding of Ubiquitin. An Analysis in Terms of the Contributions from Charge-Charge Interactions to Protein Stability. Biochemistry 1999, 38, 8138–8149. [Google Scholar] [CrossRef] [PubMed]
- Winget, J.M.; Mayor, T. The Diversity of Ubiquitin Recognition: Hot Spots and Varied Specificity. Mol. Cell 2010, 38, 627–635. [Google Scholar] [CrossRef]
- Leung, I.; Jarvik, N.; Sidhu, S.S. A Highly Diverse and Functional Naïve Ubiquitin Variant Library for Generation of Intracellular Affinity Reagents. J. Mol. Biol. 2017, 429, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Roscoe, B.P.; Thayer, K.M.; Zeldovich, K.B.; Fushman, D.; Bolon, D.N.A. Analyses of the Effects of All Ubiquitin Point Mutants on Yeast Growth Rate. J. Mol. Biol. 2013, 425, 1363–1377. [Google Scholar] [CrossRef] [PubMed]
- Pruneda, J.N.; Smith, F.D.; Daurie, A.; Swaney, D.L.; Villén, J.; Scott, J.D.; Stadnyk, A.W.; Le Trong, I.; Stenkamp, R.E.; Klevit, R.E.; et al. E2~Ub Conjugates Regulate the Kinase Activity of Shigella Effector OspG during Pathogenesis. EMBO J. 2014, 33, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Wiener, R.; DiBello, A.T.; Lombardi, P.; Guzzo, C.M.; Zhang, X.; Matunis, M.J.; Wolberger, C. E2 Ubiquitin Conjugating Enzymes Regulate the Deubiquitinating Activity of OTUB1. Nat. Struct. Mol. Biol. 2013, 20, 1033–1039. [Google Scholar] [CrossRef] [PubMed]
- Alpi, A.F.; Chaugule, V.; Walden, H. Mechanism and Disease Association of E2-Conjugating Enzymes: Lessons from UBE2T and UBE2L3. Biochem. J. 2016, 473, 3401–3419. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Huang, P.; Qin, M.; Lu, Q.; Sang, X.; Cai, Y.; Wang, Y.; Liu, F.; Wu, R.; Wang, X.; et al. Reduction of HIP2 Expression Causes Motor Function Impairment and Increased Vulnerability to Dopaminergic Degeneration in Parkinson’s Disease Models. Cell Death Dis. 2018, 9, 1020. [Google Scholar] [CrossRef]
- Tak, Y.J.; Kang, S. The E2 Ubiquitin-Conjugating Enzyme HIP2 Is a Crucial Regulator of Quality Control against Mutant SOD1 Proteotoxicity. Biochim. Biophys. Acta Mol. Basis Dis. 2022, 1868, 166316. [Google Scholar] [CrossRef]
- Song, S.; Kim, S.-Y.; Hong, Y.-M.; Jo, D.-G.; Lee, J.-Y.; Shim, S.M.; Chung, C.-W.; Seo, S.J.; Yoo, Y.J.; Koh, J.-Y.; et al. Essential Role of E2-25K/Hip-2 in Mediating Amyloid-Beta Neurotoxicity. Mol. Cell 2003, 12, 553–563. [Google Scholar] [CrossRef]
- Middleton, A.J.; Teyra, J.; Zhu, J.; Sidhu, S.S.; Day, C.L. Identification of Ubiquitin Variants That Inhibit the E2 Ubiquitin Conjugating Enzyme, Ube2k. ACS Chem. Biol. 2021, 16, 1745–1756. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.D.; Ritterhoff, T.; Klevit, R.E.; Brzovic, P.S. E2 Enzymes: More than Just Middle Men. Cell Res. 2016, 26, 423–440. [Google Scholar] [CrossRef] [PubMed]
- Garg, P.; Ceccarelli, D.F.; Keszei, A.F.A.; Kurinov, I.; Sicheri, F.; Sidhu, S.S. Structural and Functional Analysis of Ubiquitin-Based Inhibitors That Target the Backsides of E2 Enzymes. J. Mol. Biol. 2020, 432, 952–966. [Google Scholar] [CrossRef] [PubMed]
- George, A.J.; Hoffiz, Y.C.; Charles, A.J.; Zhu, Y.; Mabb, A.M. A Comprehensive Atlas of E3 Ubiquitin Ligase Mutations in Neurological Disorders. Front. Genet. 2018, 9, 29. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Zhao, J.; Chen, D.; Wang, Y. E3 Ubiquitin Ligases: Styles, Structures and Functions. Mol. Biomed. 2021, 2, 23. [Google Scholar] [CrossRef] [PubMed]
- Potjewyd, F.M.; Axtman, A.D. Exploration of Aberrant E3 Ligases Implicated in Alzheimer’s Disease and Development of Chemical Tools to Modulate Their Function. Front. Cell. Neurosci. 2021, 15, 768655. [Google Scholar] [CrossRef] [PubMed]
- Jevtić, P.; Haakonsen, D.L.; Rapé, M. An E3 Ligase Guide to the Galaxy of Small-Molecule-Induced Protein Degradation. Cell Chem. Biol. 2021, 28, 1000–1013. [Google Scholar] [CrossRef]
- Wang, Z.; Hu, X.; Ye, M.; Lin, M.; Chu, M.; Shen, X. NEDD4 E3 Ligase: Functions and Mechanism in Human Cancer. Semin. Cancer Biol. 2020, 67, 92–101. [Google Scholar] [CrossRef]
- Rotin, D. Role of the UPS in Liddle Syndrome. BMC Biochem. 2008, 9, S5. [Google Scholar] [CrossRef]
- Zhang, W.; Wu, K.-P.; Sartori, M.A.; Kamadurai, H.B.; Ordureau, A.; Jiang, C.; Mercredi, P.Y.; Murchie, R.; Hu, J.; Persaud, A.; et al. System-Wide Modulation of HECT E3 Ligases with Selective Ubiquitin Variant Probes. Mol. Cell 2016, 62, 121–136. [Google Scholar] [CrossRef]
- Ernst, A.; Avvakumov, G.; Tong, J.; Fan, Y.; Zhao, Y.; Alberts, P.; Persaud, A.; Walker, J.R.; Neculai, A.-M.; Neculai, D.; et al. A Strategy for Modulation of Enzymes in the Ubiquitin System. Science 2013, 339, 590–595. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Leng, R.P. UBE4B, a Ubiquitin Chain Assembly Factor, Is Required for MDM2-Mediated P53 Polyubiquitination and Degradation. Cell Cycle 2011, 10, 1912–1915. [Google Scholar] [CrossRef] [PubMed]
- Gabrielsen, M.; Buetow, L.; Nakasone, M.A.; Ahmed, S.F.; Sibbet, G.J.; Smith, B.O.; Zhang, W.; Sidhu, S.S.; Huang, D.T. A General Strategy for Discovery of Inhibitors and Activators of RING and U-Box E3 Ligases with Ubiquitin Variants. Mol. Cell 2017, 68, 456–470.e10. [Google Scholar] [CrossRef] [PubMed]
- Visser Smit, G.D.; Place, T.L.; Cole, S.L.; Clausen, K.A.; Vemuganti, S.; Zhang, G.; Koland, J.G.; Lill, N.L. Cbl Controls EGFR Fate by Regulating Early Endosome Fusion. Sci. Signal. 2009, 2, ra86. [Google Scholar] [CrossRef] [PubMed]
- Pierce, N.W.; Lee, J.E.; Liu, X.; Sweredoski, M.J.; Graham, R.L.J.; Larimore, E.A.; Rome, M.; Zheng, N.; Clurman, B.E.; Hess, S.; et al. Cand1 Promotes Assembly of New SCF Complexes through Dynamic Exchange of F Box Proteins. Cell 2013, 153, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Gorelik, M.; Orlicky, S.; Sartori, M.A.; Tang, X.; Marcon, E.; Kurinov, I.; Greenblatt, J.F.; Tyers, M.; Moffat, J.; Sicheri, F.; et al. Inhibition of SCF Ubiquitin Ligases by Engineered Ubiquitin Variants That Target the Cul1 Binding Site on the Skp1–F-Box Interface. Proc. Natl. Acad. Sci. USA 2016, 113, 3527–3532. [Google Scholar] [CrossRef]
- Gorelik, M.; Manczyk, N.; Pavlenco, A.; Kurinov, I.; Sidhu, S.S.; Sicheri, F. A Structure-Based Strategy for Engineering Selective Ubiquitin Variant Inhibitors of Skp1-Cul1-F-Box Ubiquitin Ligases. Structure 2018, 26, 1226–1236.e3. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.; Carrasco, G.; Li, B.; Sophocleous, A.; Idris, A.I. TRAF6 as a Potential Target in Advanced Breast Cancer: A Systematic Review, Meta-Analysis, and Bioinformatics Validation. Sci. Rep. 2023, 13, 4646. [Google Scholar] [CrossRef]
- Gudey, S.K.; Sundar, R.; Mu, Y.; Wallenius, A.; Zang, G.; Bergh, A.; Heldin, C.-H.; Landström, M. TRAF6 Stimulates the Tumor-Promoting Effects of TGFβ Type I Receptor through Polyubiquitination and Activation of Presenilin 1. Sci. Signal. 2014, 7, ra2. [Google Scholar] [CrossRef]
- Mu, Y.; Sundar, R.; Thakur, N.; Ekman, M.; Gudey, S.K.; Yakymovych, M.; Hermansson, A.; Dimitriou, H.; Bengoechea-Alonso, M.T.; Ericsson, J.; et al. TRAF6 Ubiquitinates TGFβ Type I Receptor to Promote Its Cleavage and Nuclear Translocation in Cancer. Nat. Commun. 2011, 2, 330. [Google Scholar] [CrossRef]
- Sacco, J.J.; Coulson, J.M.; Clague, M.J.; Urbé, S. Emerging Roles of Deubiquitinases in Cancer-associated Pathways. IUBMB Life 2010, 62, 140–157. [Google Scholar] [CrossRef] [PubMed]
- Harrigan, J.A.; Jacq, X.; Martin, N.M.; Jackson, S.P. Deubiquitylating Enzymes and Drug Discovery: Emerging Opportunities. Nat. Rev. Drug Discov. 2018, 17, 57–78. [Google Scholar] [CrossRef] [PubMed]
- Gennaro, V.J.; Stanek, T.J.; Peck, A.R.; Sun, Y.; Wang, F.; Qie, S.; Knudsen, K.E.; Rui, H.; Butt, T.; Diehl, J.A.; et al. Control of CCND1 Ubiquitylation by the Catalytic SAGA Subunit USP22 Is Essential for Cell Cycle Progression through G1 in Cancer Cells. Proc. Natl. Acad. Sci. USA 2018, 115, E9298–E9307. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Dent, S.Y.R. Functions of SAGA in Development and Disease. Epigenomics 2014, 6, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Popov, N.; Wanzel, M.; Madiredjo, M.; Zhang, D.; Beijersbergen, R.; Bernards, R.; Moll, R.; Elledge, S.J.; Eilers, M. The Ubiquitin-Specific Protease USP28 Is Required for MYC Stability. Nat. Cell Biol. 2007, 9, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-B.; Min, Y.; Hwang, S.; Baek, K.-H. Suppression of USP7 Negatively Regulates the Stability of ETS Proto-Oncogene 2 Protein. Biomed. Pharmacother. 2023, 162, 114700. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.; Young, M.A.; Donato, N.J. Emerging Potential of Therapeutic Targeting of Ubiquitin-Specific Proteases in the Treatment of Cancer. Cancer Res. 2014, 74, 4955–4966. [Google Scholar] [CrossRef]
- Ritorto, M.S.; Ewan, R.; Perez-Oliva, A.B.; Knebel, A.; Buhrlage, S.J.; Wightman, M.; Kelly, S.M.; Wood, N.T.; Virdee, S.; Gray, N.S.; et al. Screening of DUB Activity and Specificity by MALDI-TOF Mass Spectrometry. Nat. Commun. 2014, 5, 4763. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, L.; Rouge, L.; Phillips, A.H.; Lam, C.; Liu, P.; Sandoval, W.; Helgason, E.; Murray, J.M.; Wertz, I.E.; et al. Conformational Stabilization of Ubiquitin Yields Potent and Selective Inhibitors of USP7. Nat. Chem. Biol. 2013, 9, 51–58. [Google Scholar] [CrossRef]
- Zhang, W.; Sartori, M.A.; Makhnevych, T.; Federowicz, K.E.; Dong, X.; Liu, L.; Nim, S.; Dong, A.; Yang, J.; Li, Y.; et al. Generation and Validation of Intracellular Ubiquitin Variant Inhibitors for USP7 and USP10. J. Mol. Biol. 2017, 429, 3546–3560. [Google Scholar] [CrossRef]
- Yuan, J.; Luo, K.; Zhang, L.; Cheville, J.C.; Lou, Z. USP10 Regulates P53 Localization and Stability by Deubiquitinating P53. Cell 2010, 140, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, L.F.; Sparks, A.; Allende-Vega, N.; Xirodimas, D.P.; Lane, D.P.; Saville, M.K. The Deubiquitinating Enzyme USP2a Regulates the P53 Pathway by Targeting Mdm2. EMBO J. 2007, 26, 976–986. [Google Scholar] [CrossRef] [PubMed]
- Pascoe, N.; Seetharaman, A.; Teyra, J.; Manczyk, N.; Satori, M.A.; Tjandra, D.; Makhnevych, T.; Schwerdtfeger, C.; Brasher, B.B.; Moffat, J.; et al. Yeast Two-Hybrid Analysis for Ubiquitin Variant Inhibitors of Human Deubiquitinases. J. Mol. Biol. 2019, 431, 1160–1171. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.G.F.; Seo, M.-H.; Nim, S.; Corbi-Verge, C.; Kim, P.M. Protein Engineering by Highly Parallel Screening of Computationally Designed Variants. Sci. Adv. 2016, 2, e1600692. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Ni, D.; Zhang, H.; Li, X.; Jiang, Y.; Wu, J.; Gu, Y.; Gao, M.; Shi, W.; Song, J.; et al. Over-expression of USP15/MMP3 Predict Poor Prognosis and Promote Growth, Migration in Non-Small Cell Lung Cancer Cells. Cancer Genet. 2023, 272–273, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-J.; Min, Y.; Jeong, S.-K.; Son, J.; Kim, J.Y.; Lee, J.S.; Kim, D.-H.; Lee, J.S.; Chun, E.; Lee, K.-Y. USP15 Negatively Regulates Lung Cancer Progression through the TRAF6-BECN1 Signaling Axis for Autophagy Induction. Cell Death Dis. 2022, 13, 348. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Liao, Q.; Tan, W.; Peng, C.; Hu, Z.; Chen, Y.; Li, Z.; Li, J.; Zhen, B.; Zhu, W.; et al. The Deubiquitylating Enzyme USP15 Regulates Homologous Recombination Repair and Cancer Cell Response to PARP Inhibitors. Nat. Commun. 2019, 10, 1224. [Google Scholar] [CrossRef]
- Ren, Y.; Song, Z.; Rieser, J.; Ackermann, J.; Koch, I.; Lv, X.; Ji, T.; Cai, X. USP15 Represses Hepatocellular Carcinoma Progression by Regulation of Pathways of Cell Proliferation and Cell Migration: A System Biology Analysis. Cancers 2023, 15, 1371. [Google Scholar] [CrossRef]
- Teyra, J.; Singer, A.U.; Schmitges, F.W.; Jaynes, P.; Kit Leng Lui, S.; Polyak, M.J.; Fodil, N.; Krieger, J.R.; Tong, J.; Schwerdtfeger, C.; et al. Structural and Functional Characterization of Ubiquitin Variant Inhibitors of USP15. Structure 2019, 27, 590–605.e5. [Google Scholar] [CrossRef]
- Berlin, I.; Schwartz, H.; Nash, P.D. Regulation of Epidermal Growth Factor Receptor Ubiquitination and Trafficking by the USP8·STAM Complex. J. Biol. Chem. 2010, 285, 34909–34921. [Google Scholar] [CrossRef]
- Pan, X.; Wu, S.; Wei, W.; Chen, Z.; Wu, Y.; Gong, K. Structural and Functional Basis of JAMM Deubiquitinating Enzymes in Disease. Biomolecules 2022, 12, 910. [Google Scholar] [CrossRef] [PubMed]
- McDonell, L.M.; Mirzaa, G.M.; Alcantara, D.; Schwartzentruber, J.; Carter, M.T.; Lee, L.J.; Clericuzio, C.L.; Graham, J.M.; Morris-Rosendahl, D.J.; Polster, T.; et al. Mutations in STAMBP, Encoding a Deubiquitinating Enzyme, Cause Microcephaly-Capillary Malformation Syndrome. Nat. Genet. 2013, 45, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Bednash, J.S.; Weathington, N.; Londino, J.; Rojas, M.; Gulick, D.L.; Fort, R.; Han, S.; McKelvey, A.C.; Chen, B.B.; Mallampalli, R.K. Targeting the Deubiquitinase STAMBP Inhibits NALP7 Inflammasome Activity. Nat. Commun. 2017, 8, 15203. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Liu, Q.; Mallette, E.; Caba, C.; Hou, F.; Fux, J.; LaPlante, G.; Dong, A.; Zhang, Q.; Zheng, H.; et al. Structural and Functional Characterization of Ubiquitin Variant Inhibitors for the JAMM-Family Deubiquitinases STAMBP and STAMBPL1. J. Biol. Chem. 2021, 297, 101107. [Google Scholar] [CrossRef]
- Luo, H. Interplay between the Virus and the Ubiquitin–Proteasome System: Molecular Mechanism of Viral Pathogenesis. Curr. Opin. Virol. 2016, 17, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Bailey-Elkin, B.A.; Knaap, R.C.M.; Khare, B.; Dalebout, T.J.; Johnson, G.G.; van Kasteren, P.B.; McLeish, N.J.; Gu, J.; He, W.; et al. Potent and Selective Inhibition of Pathogenic Viruses by Engineered Ubiquitin Variants. PLoS Pathog. 2017, 13, e1006372. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.I.; Hsieh, Y.-J.; Lu, W.-L.; Wu, K.-P.; Chang, C.A. Engineering Nanomolar Potent Protein-Based Inhibitors for Papain-like Protease Guided by Residue Correlation Network. bioRxiv 2023. bioRxiv:2023.03.15.532709. [Google Scholar] [CrossRef]
- van Vliet, V.J.E.; Huynh, N.; Palà, J.; Patel, A.; Singer, A.; Slater, C.; Chung, J.; van Huizen, M.; Teyra, J.; Miersch, S.; et al. Ubiquitin Variants Potently Inhibit SARS-CoV-2 PLpro and Viral Replication via a Novel Site Distal to the Protease Active Site. PLoS Pathog. 2022, 18, e1011065. [Google Scholar] [CrossRef]
- Hewitt, C.S.; Krabill, A.D.; Das, C.; Flaherty, D.P. Development of Ubiquitin Variants with Selectivity for Ubiquitin C-Terminal Hydrolase Deubiquitinases. Biochemistry 2020, 59, 3447–3462. [Google Scholar] [CrossRef]
- Tencer, A.H.; Liang, Q.; Zhuang, Z. Divergence in Ubiquitin Interaction and Catalysis among the Ubiquitin-Specific Protease Family Deubiquitinating Enzymes. Biochemistry 2016, 55, 4708–4719. [Google Scholar] [CrossRef]
- Hewitt, C.S.; Das, C.; Flaherty, D.P. Rational Development and Characterization of a Ubiquitin Variant with Selectivity for Ubiquitin C-Terminal Hydrolase L3. Biomolecules 2022, 12, 62. [Google Scholar] [CrossRef] [PubMed]
- Gjonaj, L.; Sapmaz, A.; González-Prieto, R.; Vertegaal, A.C.O.; Flierman, D.; Ovaa, H. USP7: Combining Tools towards Selectivity. Chem. Commun. 2019, 55, 5075–5078. [Google Scholar] [CrossRef] [PubMed]
- Gjonaj, L.; Sapmaz, A.; Flierman, D.; Janssen, G.M.C.; van Veelen, P.A.; Ovaa, H. Development of a DUB-Selective Fluorogenic Substrate. Chem. Sci. 2019, 10, 10290–10296. [Google Scholar] [CrossRef] [PubMed]
- Dikic, I.; Wakatsuki, S.; Walters, K.J. Ubiquitin Binding Domains—From Structures to Functions. Nat. Rev. Mol. Cell Biol. 2009, 10, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Manczyk, N.; Yates, B.P.; Veggiani, G.; Ernst, A.; Sicheri, F.; Sidhu, S.S. Structural and Functional Characterization of a Ubiquitin Variant Engineered for Tight and Specific Binding to an Alpha-Helical Ubiquitin Interacting Motif. Protein Sci. 2017, 26, 1060–1069. [Google Scholar] [CrossRef] [PubMed]
- Manczyk, N.; Veggiani, G.; Gish, G.D.; Yates, B.P.; Ernst, A.; Sidhu, S.S.; Sicheri, F. Dimerization of a Ubiquitin Variant Leads to High Affinity Interactions with a Ubiquitin Interacting Motif. Protein Sci. 2019, 28, 848–856. [Google Scholar] [CrossRef] [PubMed]
- Veggiani, G.; Yates, B.P.; Martyn, G.D.; Manczyk, N.; Singer, A.U.; Kurinov, I.; Sicheri, F.; Sidhu, S.S. Panel of Engineered Ubiquitin Variants Targeting the Family of Human Ubiquitin Interacting Motifs. ACS Chem. Biol. 2022, 17, 941–956. [Google Scholar] [CrossRef]
- Manczyk, N.; Veggiani, G.; Teyra, J.; Strilchuk, A.W.; Sidhu, S.S.; Sicheri, F. The Ubiquitin Interacting Motifs of USP37 Act on the Proximal Ub of a Di-Ub Chain to Enhance Catalytic Efficiency. Sci. Rep. 2019, 9, 4119. [Google Scholar] [CrossRef]
- Faesen, A.C.; Dirac, A.M.G.; Shanmugham, A.; Ovaa, H.; Perrakis, A.; Sixma, T.K. Mechanism of USP7/HAUSP Activation by Its C-Terminal Ubiquitin-like Domain and Allosteric Regulation by GMP-Synthetase. Mol. Cell 2011, 44, 147–159. [Google Scholar] [CrossRef]
- Mann, M.K.; Zepeda-Velázquez, C.A.; González-Álvarez, H.; Dong, A.; Kiyota, T.; Aman, A.M.; Loppnau, P.; Li, Y.; Wilson, B.; Arrowsmith, C.H.; et al. Structure–Activity Relationship of USP5 Inhibitors. J. Med. Chem. 2021, 64, 15017–15036. [Google Scholar] [CrossRef]
- Clerici, M.; Luna-Vargas, M.P.A.; Faesen, A.C.; Sixma, T.K. The DUSP-Ubl Domain of USP4 Enhances Its Catalytic Efficiency by Promoting Ubiquitin Exchange. Nat. Commun. 2014, 5, 5399. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.Q.; Veggiani, G.; Singer, A.; Teyra, J.; Chung, J.; Sidhu, S.S. A Panel of Engineered Ubiquitin Variants Targeting the Family of Domains Found in Ubiquitin Specific Proteases (DUSPs). J. Mol. Biol. 2021, 433, 167300. [Google Scholar] [CrossRef] [PubMed]
- Watson, E.R.; Grace, C.R.R.; Zhang, W.; Miller, D.J.; Davidson, I.F.; Prabu, J.R.; Yu, S.; Bolhuis, D.L.; Kulko, E.T.; Vollrath, R.; et al. Protein Engineering of a Ubiquitin-Variant Inhibitor of APC/C Identifies a Cryptic K48 Ubiquitin Chain Binding Site. Proc. Natl. Acad. Sci. USA 2019, 116, 17280–17289. [Google Scholar] [CrossRef] [PubMed]
- Canny, M.D.; Moatti, N.; Wan, L.C.K.; Fradet-Turcotte, A.; Krasner, D.; Mateos-Gomez, P.A.; Zimmermann, M.; Orthwein, A.; Juang, Y.-C.; Zhang, W.; et al. Inhibition of 53BP1 Favors Homology-Dependent DNA Repair and Increases CRISPR–Cas9 Genome-Editing Efficiency. Nat. Biotechnol. 2018, 36, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, G.; Huang, S.; Liu, Y.; Tang, J.; Zhong, M.; Wang, X.; Sun, W.; Yao, Y.; Ji, Q.; et al. Development of a Versatile Nuclease Prime Editor with Upgraded Precision. Nat. Commun. 2023, 14, 305. [Google Scholar] [CrossRef] [PubMed]
- Lorey, S.; Fiedler, E.; Kunert, A.; Nerkamp, J.; Lange, C.; Fiedler, M.; Bosse-Doenecke, E.; Meysing, M.; Gloser, M.; Rundfeldt, C.; et al. Novel Ubiquitin-Derived High Affinity Binding Proteins with Tumor Targeting Properties. J. Biol. Chem. 2014, 289, 8493–8507. [Google Scholar] [CrossRef] [PubMed]
- Lieverse, R.I.Y.; Marcus, D.; van der Wiel, A.M.A.; Van Limbergen, E.J.; Theys, J.; Yaromina, A.; Lambin, P.; Dubois, L.J. Human Fibronectin Extra Domain B as a Biomarker for Targeted Therapy in Cancer. Mol. Oncol. 2020, 14, 1555–1568. [Google Scholar] [CrossRef]
- Zhao, L.; Zhao, J.; Zhong, K.; Tong, A.; Jia, D. Targeted Protein Degradation: Mechanisms, Strategies and Application. Signal Transduct. Target. Ther. 2022, 7, 113. [Google Scholar] [CrossRef]
- Au, Y.Z.; Wang, T.; Sigua, L.H.; Qi, J. Peptide-Based PROTAC: The Predator of Pathological Proteins. Cell Chem. Biol. 2020, 27, 637–639. [Google Scholar] [CrossRef]
- Stephens, E.A.; Ludwicki, M.B.; Meksiriporn, B.; Li, M.; Ye, T.; Monticello, C.; Forsythe, K.J.; Kummer, L.; Zhou, P.; Plückthun, A.; et al. Engineering Single Pan-Specific Ubiquibodies for Targeted Degradation of All Forms of Endogenous ERK Protein Kinase. ACS Synth. Biol. 2021, 10, 2396–2408. [Google Scholar] [CrossRef]
- Aminu, B.; Fux, J.; Mallette, E.; Petersen, N.; Zhang, W. Targeted Degradation of 53BP1 Using Ubiquitin Variant Induced Proximity. Biomolecules 2022, 12, 479. [Google Scholar] [CrossRef] [PubMed]
- Cyrus, K.; Wehenkel, M.; Choi, E.-Y.; Han, H.-J.; Lee, H.; Swanson, H.; Kim, K.-B. Impact of Linker Length on the Activity of PROTACs. Mol. Biosyst. 2011, 7, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Zephyr, J.; Kurt Yilmaz, N.; Schiffer, C.A. Viral Proteases: Structure, Mechanism and Inhibition. Enzymes 2021, 50, 301–333. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Bi, Y.; Zhang, H.; Dong, S.; Teng, L.; Lee, R.J.; Yang, Z. Cell-Penetrating Peptides in Diagnosis and Treatment of Human Diseases: From Preclinical Research to Clinical Application. Front. Pharmacol. 2020, 11, 697. [Google Scholar] [CrossRef] [PubMed]
- El-Andaloussi, S.; Järver, P.; Johansson, H.J.; Langel, Ü. Cargo-Dependent Cytotoxicity and Delivery Efficacy of Cell-Penetrating Peptides: A Comparative Study. Biochem. J. 2007, 407, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Bulcha, J.T.; Wang, Y.; Ma, H.; Tai, P.W.L.; Gao, G. Viral Vector Platforms within the Gene Therapy Landscape. Signal Transduct. Target. Ther. 2021, 6, 53. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.; Kim, S. Recent Advances in the Development of Gene Delivery Systems. Biomater. Res. 2019, 23, 8. [Google Scholar] [CrossRef]
- Chang, J.R.; Song, E.-H.; Nakatani-Webster, E.; Monkkonen, L.; Ratner, D.M.; Catalano, C.E. Phage Lambda Capsids as Tunable Display Nanoparticles. Biomacromolecules 2014, 15, 4410–4419. [Google Scholar] [CrossRef]
- McClary, W.D.; Catala, A.; Zhang, W.; Gamboni, F.; Dzieciatkowska, M.; Sidhu, S.S.; D’Alessandro, A.; Catalano, C.E. A Designer Nanoparticle Platform for Controlled Intracellular Delivery of Bioactive Macromolecules: Inhibition of Ubiquitin-Specific Protease 7 in Breast Cancer Cells. ACS Chem. Biol. 2022, 17, 1853–1865. [Google Scholar] [CrossRef]
- Phillips, A.H.; Zhang, Y.; Cunningham, C.N.; Zhou, L.; Forrest, W.F.; Liu, P.S.; Steffek, M.; Lee, J.; Tam, C.; Helgason, E.; et al. Conformational Dynamics Control Ubiquitin-Deubiquitinase Interactions and Influence in Vivo Signaling. Proc. Natl. Acad. Sci. USA 2013, 110, 11379–11384. [Google Scholar] [CrossRef]

| Features | Small Molecule Degraders | UbVIPs |
|---|---|---|
| Target Protein Range | Broad spectrum | Selective targeting of specific proteins within the UPS and beyond |
| Mechanism of Action | Recruit target protein to E3 ligase for ubiquitination | Facilitate ubiquitination and degradation of target proteins. Can enhance E3 ligase ubiquitination activity |
| Delivery and Cellular Uptake | Diffusion across cell membranes | Require specialized methods |
| Interaction with the UPS | Engage only a small subset of E3 ligases | Recruit diverse E3 ligases with distinct activity and cellular localization. Can engage E2 enzymes and DUBs |
| Target Protein Degradation Speed | Rapid and reversible | Can vary depending on UbV design and abundance |
| Cellular Response | Transient degradation of target protein | Transient modulation of target protein degradation |
| Binding Site Specificity | Depend on ligand and target protein interaction | Highly specific due to modular customization of UbVs |
| Protein Degradation Scope | Limited by the availability of specific ligands | Flexible and can be applied to a wide range of target proteins |
| Druggability | Require development of specific ligands for targets | Expand the repertoire of druggable targets even beyond the UPS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, J.Q.; Marchand, M.M.; Veggiani, G. Ubiquitin Engineering for Interrogating the Ubiquitin–Proteasome System and Novel Therapeutic Strategies. Cells 2023, 12, 2117. https://doi.org/10.3390/cells12162117
Tang JQ, Marchand MM, Veggiani G. Ubiquitin Engineering for Interrogating the Ubiquitin–Proteasome System and Novel Therapeutic Strategies. Cells. 2023; 12(16):2117. https://doi.org/10.3390/cells12162117
Chicago/Turabian StyleTang, Jason Q., Mary M. Marchand, and Gianluca Veggiani. 2023. "Ubiquitin Engineering for Interrogating the Ubiquitin–Proteasome System and Novel Therapeutic Strategies" Cells 12, no. 16: 2117. https://doi.org/10.3390/cells12162117
APA StyleTang, J. Q., Marchand, M. M., & Veggiani, G. (2023). Ubiquitin Engineering for Interrogating the Ubiquitin–Proteasome System and Novel Therapeutic Strategies. Cells, 12(16), 2117. https://doi.org/10.3390/cells12162117