
New Phage-Derived Antibacterial Enzyme PolaR Targeting Rothia spp.

 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Sample Collection
2.2. DNA Extraction
2.3. Illumina Sequencing and Processing of Sequencing Data
2.4. Assembly of Metagenomic Contigs
2.5. Identification of Genes Coding New Antibacterial Enzymes
2.6. Analysis of the Genomic and Evolutionary Background of the Selected Lysin
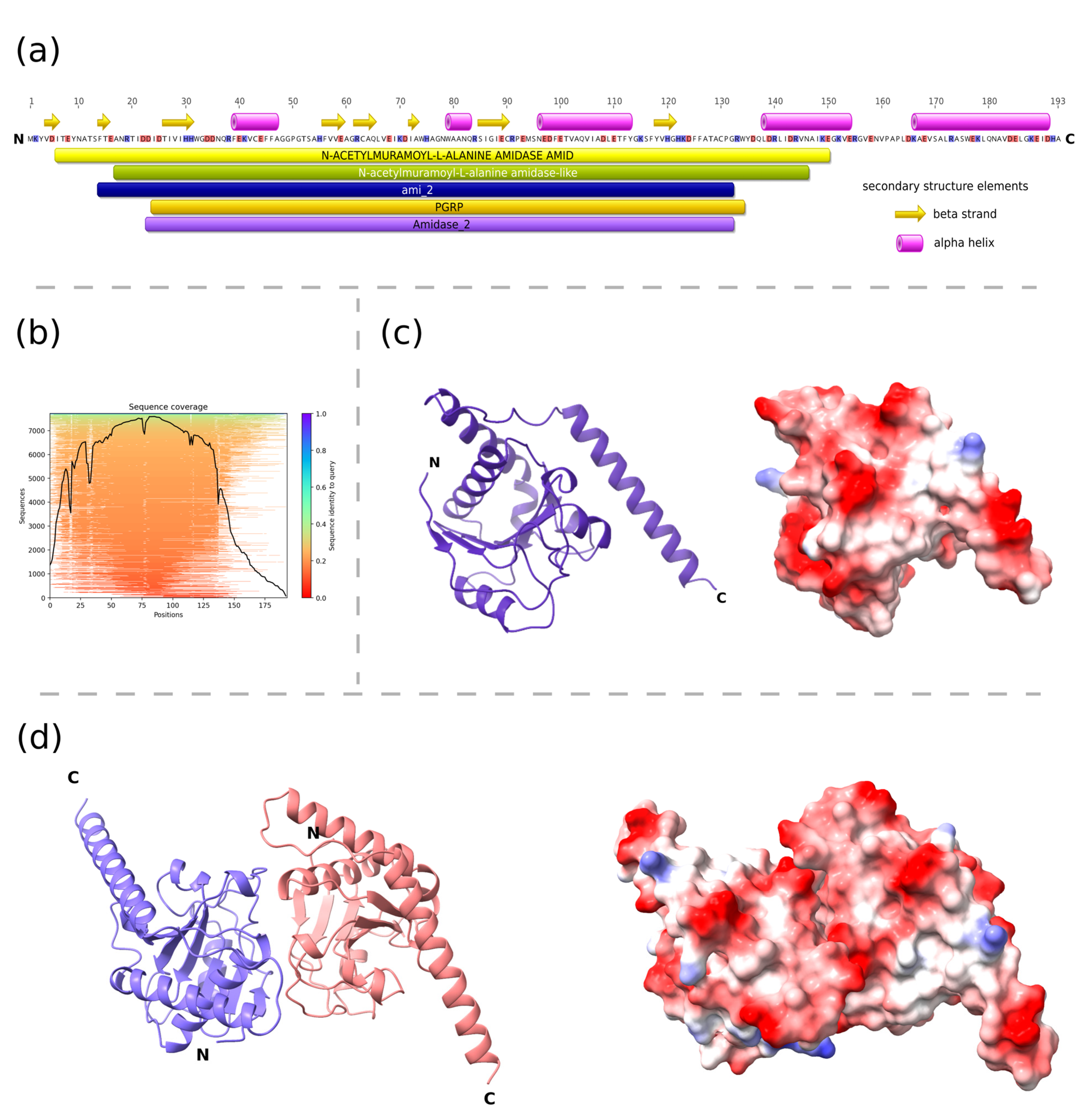
2.7. Prediction of Enzyme Structure
2.8. Protein Production
2.9. PolaR’s Lytic Activity—Fluorometric Reduction Assay
2.10. Antibacterial Activity Test Using Single Cell Microscopy
2.11. Antibacterial Aggregate Activity—Fluorometric and Turbidity Reduction Assay
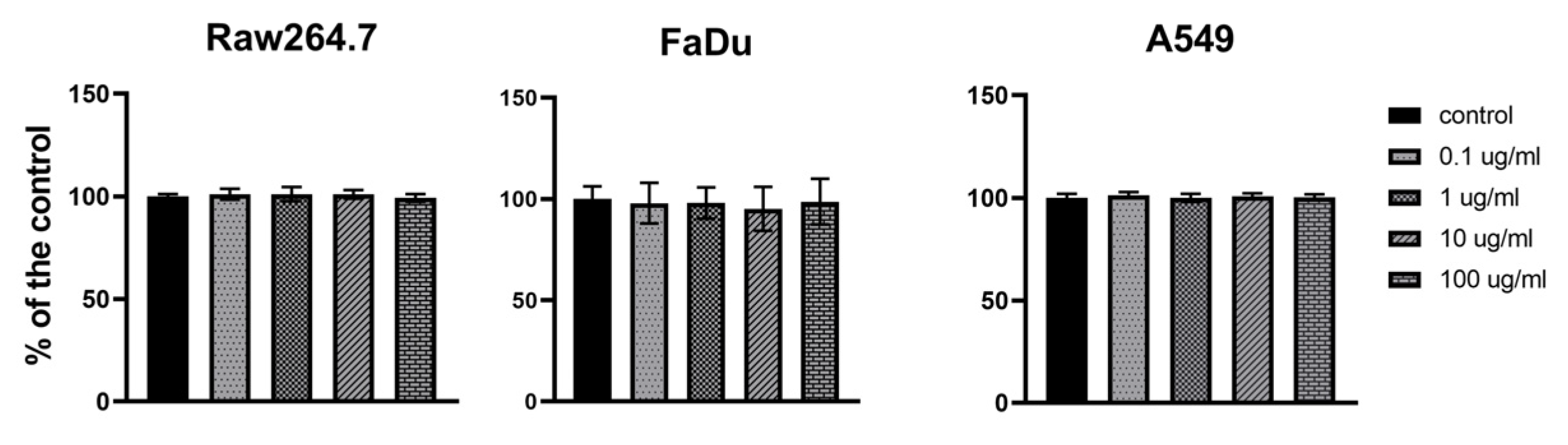
2.12. Safety Assay of PolaR
3. Results
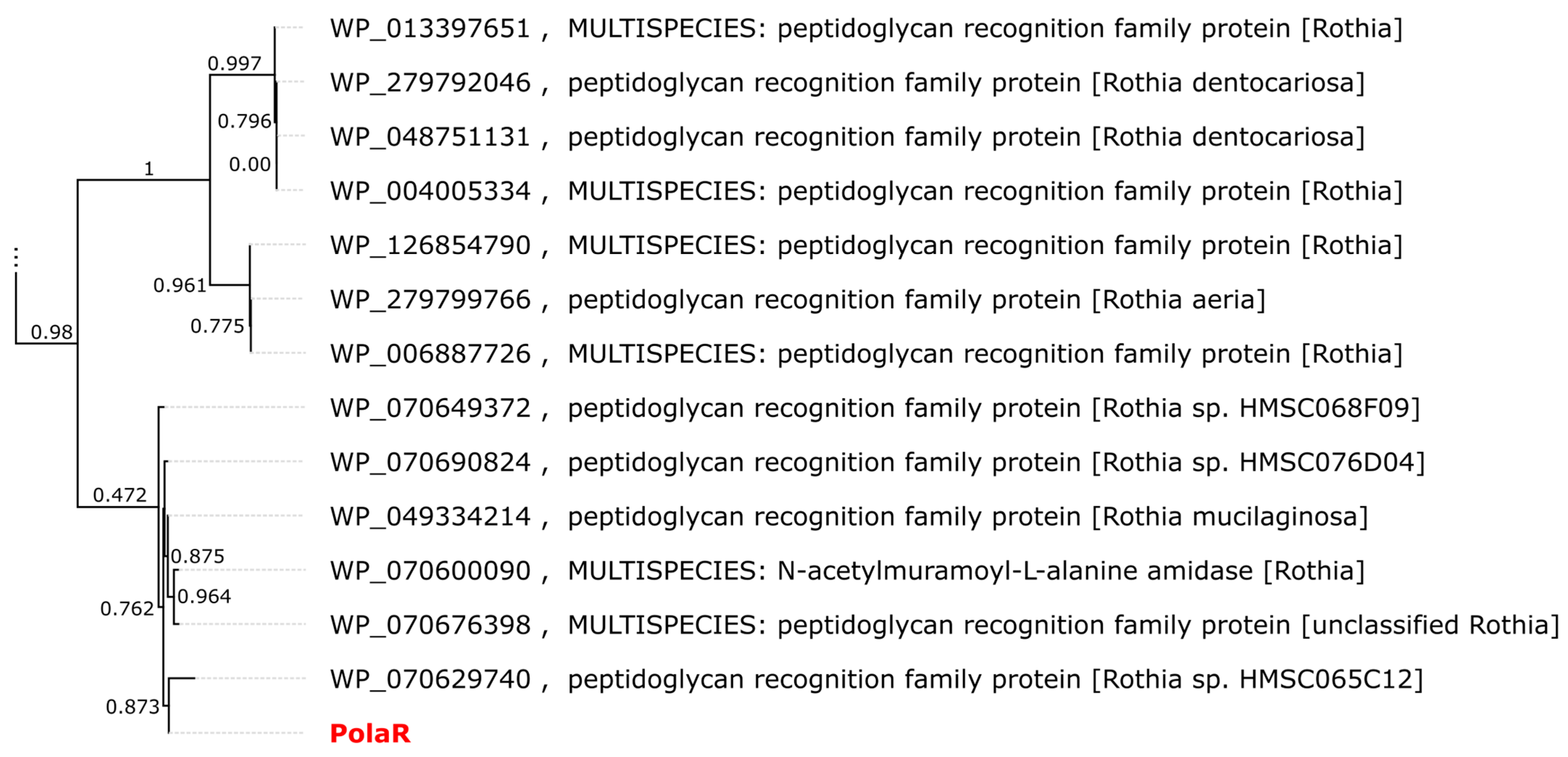
3.1. Investigating Human Stomach Phageomes for the Selection of Candidate Endolysins
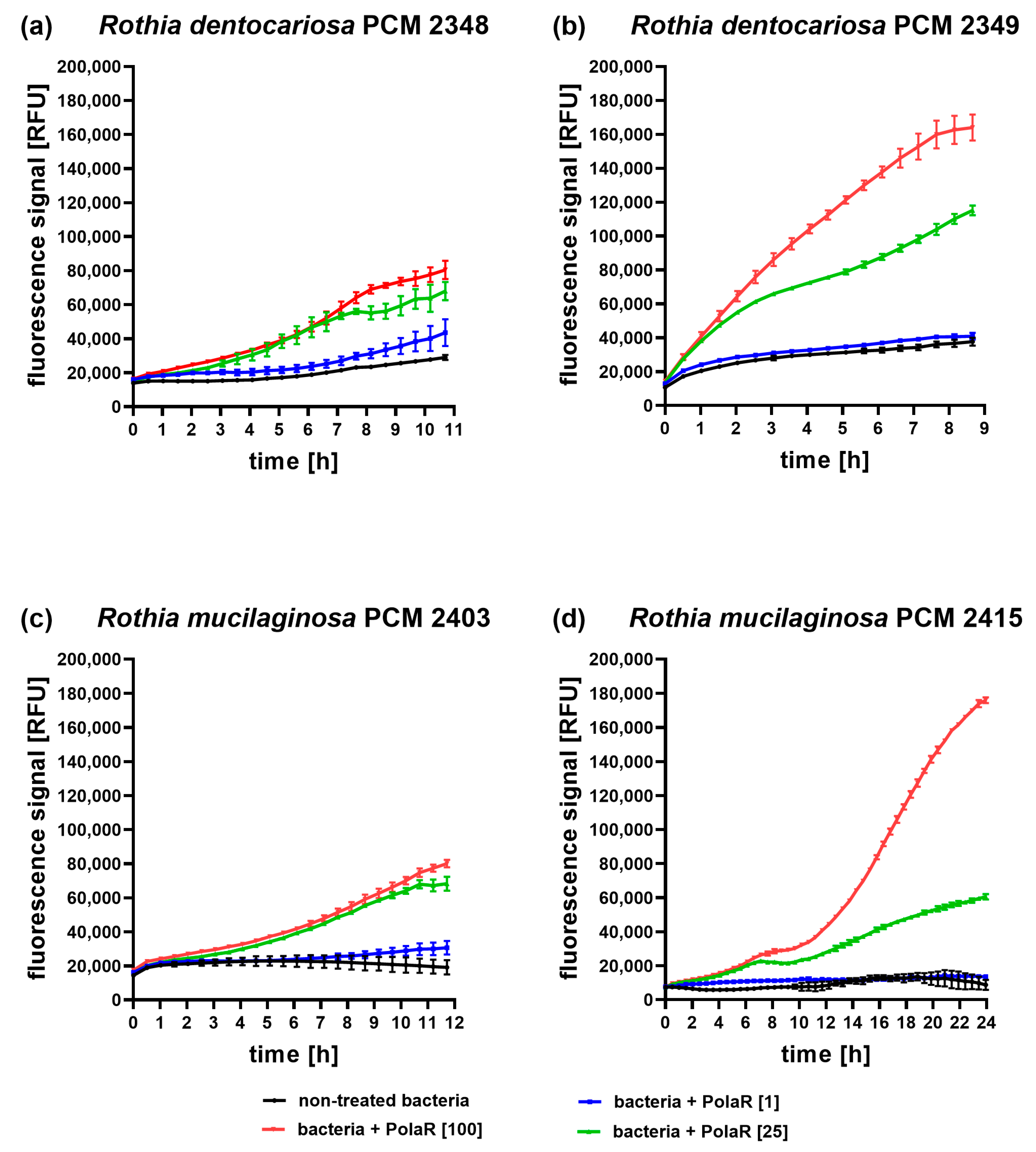
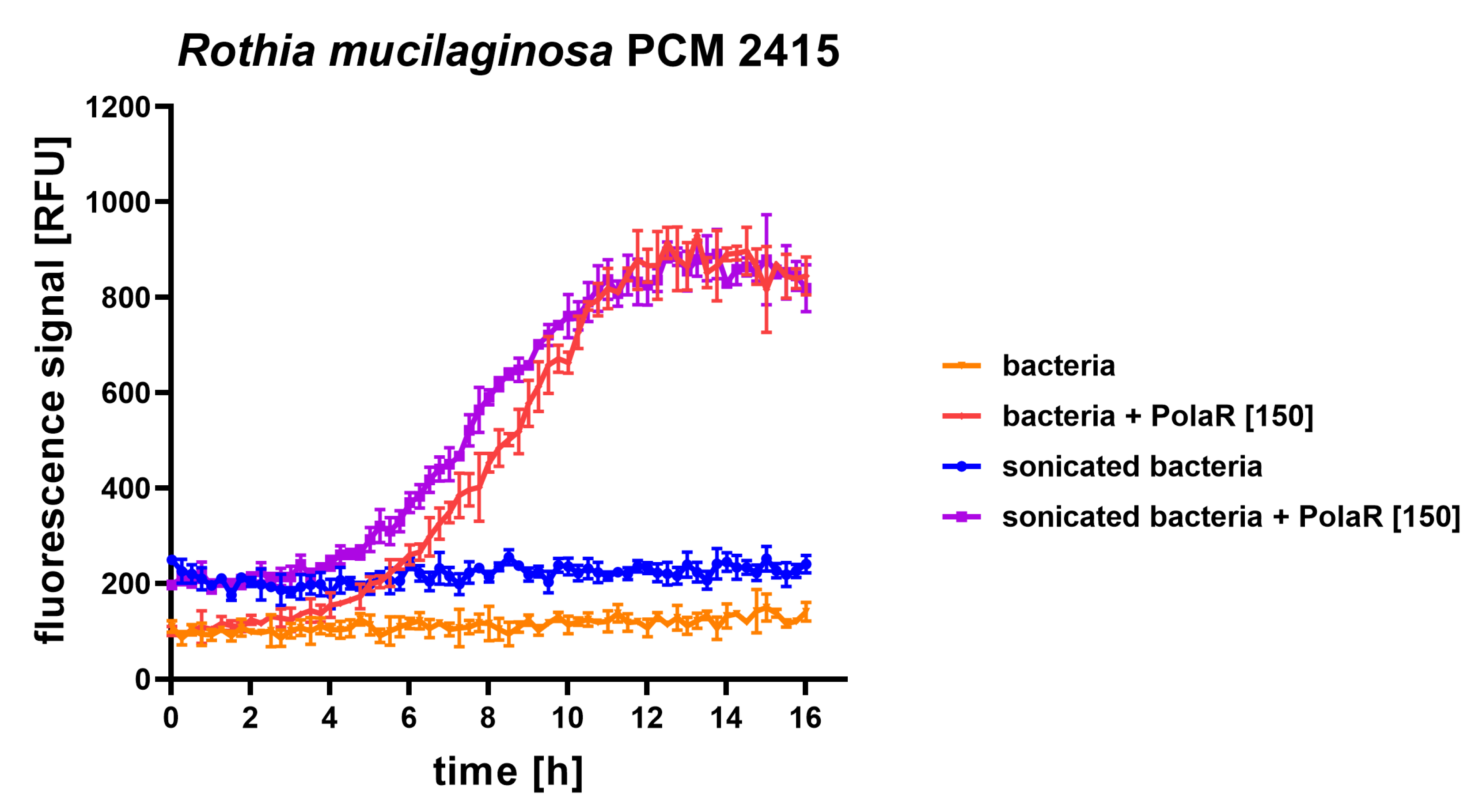
3.2. Antibacterial Activity of the New Candidate Endolysin
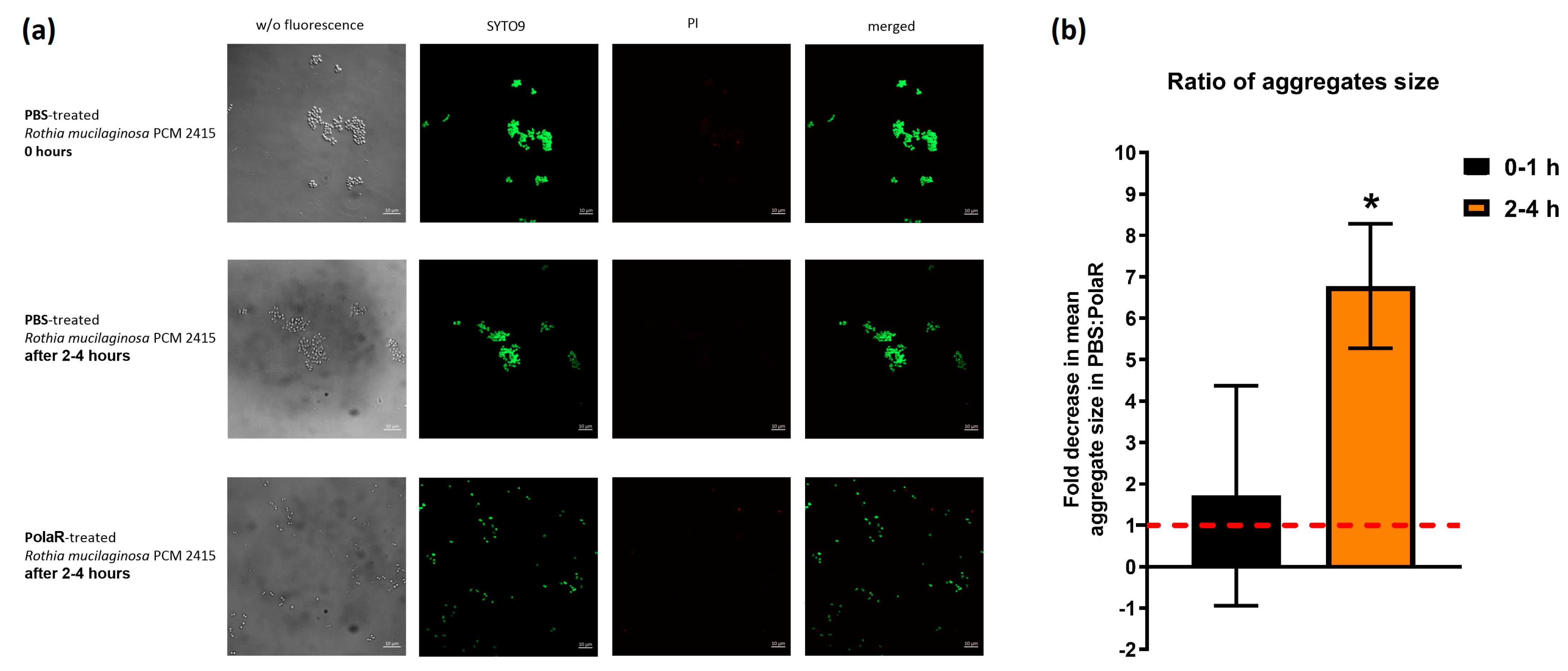
3.3. Antibacterial Aggregate Activity of the New Candidate Endolysin
3.4. Safety of PolaR Testing on Mammalian Cell Cultures
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Franconieri, F.; Join-Lambert, O.; Creveuil, C.; Auzou, M.; Labombarda, F.; Aouba, A.; Verdon, R.; de La Blanchardière, A. Rothia spp. infective endocarditis: A systematic literature review. Infect. Dis. Now 2021, 51, 228–235. [Google Scholar] [CrossRef]
- Fatahi-Bafghi, M. Characterization of the Rothia spp. and their role in human clinical infections. Infect. Genet. Evol. 2021, 93, 104877. [Google Scholar] [CrossRef] [PubMed]
- Georg, L.K.; Brown, J.M. Rothia, gen. nov. an aerobic genus of the family Actinomycetaceae. Int. J. Syst. Bacteriol. 1967, 17, 79–88. [Google Scholar] [CrossRef]
- Onishi, M. Study on the Actinomyces isolated from the deeper layers of carious dentine. Shikagaku Zasshi 1949, 6, 273–318. [Google Scholar]
- Davis, G.H.G.; Freer, J.H. Studies Upon an Oral Aerobic Actinomycete. J. Gen. Microbiol. 1960, 23, 163–178. [Google Scholar] [CrossRef][Green Version]
- NCBI Taxonomy. Available online: https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?id=32207 (accessed on 27 June 2023).
- Oliveira, I.M.F.; Ng, D.Y.K.; van Baarlen, P.; Stegger, M.; Andersen, P.S.; Wells, J.M. Comparative genomics of Rothia species reveals diversity in novel biosynthetic gene clusters and ecological adaptation to different eukaryotic hosts and host niches. Microb. Genom. 2022, 8, mgen000854. [Google Scholar] [CrossRef]
- Tsuzukibashi, O.; Uchibori, S.; Kobayashi, T.; Umezawa, K.; Mashimo, C.; Nambu, T.; Saito, M.; Hashizume-Takizawa, T.; Ochiai, T. Isolation and identification methods of Rothia species in oral cavities. J. Microbiol. Methods 2017, 134, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Zivkovic Zaric, R.S.; Pejcic, A.V.; Jankovic, S.M.; Kostic, M.J.; Milosavljevic, M.N.; Milosavljevic, M.J.; Opancina, V.D. Antimicrobial treatment of Kocuria kristinae invasive infections: Systematic review. J. Chemother. 2019, 31, 109–119. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Koch, C.; Gvozdiak, O.; Schumann, P. Taxonomic dissection of the genus Micrococcus: Kocuria gen. nov., Nesterenkonia gen. nov., Kytococcus gen. nov., Dermacoccus gen. nov., and Micrococcus Cohn 1872 gen. emend. Int. J. Syst. Bacteriol. 1995, 45, 682–692. [Google Scholar] [CrossRef]
- Khan, S.T.; Ahamed, M.; Musarrat, J.; Al-Khedhairy, A.A. Anti-biofilm and antibacterial activities of zinc oxide nanoparticles against the oral opportunistic pathogens Rothia dentocariosa and Rothia mucilaginosa. Eur. J. Oral. Sci. 2014, 122, 397–403. [Google Scholar] [CrossRef]
- Lim, Y.W.; Schmieder, R.; Haynes, M.; Furlan, M.; Matthews, T.D.; Whiteson, K.; Poole, S.J.; Hayes, C.S.; Low, D.A.; Maughan, H.; et al. Mechanistic model of Rothia mucilaginosa adaptation toward persistence in the CF lung, based on a genome reconstructed from metagenomic data. PLoS ONE 2013, 8, e64285. [Google Scholar] [CrossRef] [PubMed]
- Ramanan, P.; Barreto, J.N.; Osmon, D.R.; Tosh, P.K. Rothia Bacteremia: A 10-Year Experience at Mayo Clinic, Rochester, Minnesota. J. Clin. Microbiol. 2014, 52, 3184–3189. [Google Scholar] [CrossRef] [PubMed]
- Cunniffe, J.G.; Mallia, C.; Alcock, P.A. Stomatococcus mucilaginosus lower respiratory tract infection in a patient with AIDS. J. Infect. 1994, 29, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Tunney, M.M.; Field, T.R.; Moriarty, T.F.; Patrick, S.; Doering, G.; Muhlebach, M.S.; Wolfgang, M.C.; Boucher, R.; Gilpin, D.F.; McDowell, A.; et al. Detection of anaerobic bacteria in high numbers in sputum from patients with cystic fibrosis. Am. J. Respir. Crit. Care Med. 2008, 177, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Gallagher, T.; Zhang, Y.; Elbadawi-Sidhu, M.; Lai, Z.; Fiehn, O.; Whiteson, K.L. Tracking Polymicrobial Metabolism in Cystic Fibrosis Airways: Pseudomonas aeruginosa Metabolism and Physiology Are Influenced by Rothia mucilaginosa-Derived Metabolites. mSphere 2018, 3, e00151-18. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.W.; Evangelista, J.S., III; Schmieder, R.; Bailey, B.; Haynes, M.; Furlan, M.; Maughan, H.; Edwards, R.; Rohwer, F.; Conrad, D. Clinical insights from metagenomic analysis of sputum samples from patients with cystic fibrosis. J. Clin. Microbiol. 2014, 52, 425–437. [Google Scholar] [CrossRef]
- Silveira, C.B.; Cobián-Güemes, A.G.; Uranga, C.; Baker, J.L.; Edlund, A.; Rohwer, F.; Conrad, D. Multi-Omics Study of Keystone Species in a Cystic Fibrosis Microbiome. Int. J. Mol. Sci. 2021, 22, 12050. [Google Scholar] [CrossRef]
- Coburn, B.; Wang, P.W.; Diaz Caballero, J.; Clark, S.T.; Brahma, V.; Donaldson, S.; Zhang, Y.; Surendra, A.; Gong, Y.; Elizabeth Tullis, D.; et al. Lung microbiota across age and disease stage in cystic fibrosis. Sci. Rep. 2015, 5, 10241. [Google Scholar] [CrossRef]
- Rigauts, C.; Aizawa, J.; Taylor, S.L.; Rogers, G.B.; Govaerts, M.; Cos, P.; Ostyn, L.; Sims, S.; Vandeplassche, E.; Sze, M.; et al. Rothia mucilaginosa is an anti-inflammatory bacterium in the respiratory tract of patients with chronic lung disease. Eur. Respir. J. 2022, 59, 2101293. [Google Scholar] [CrossRef]
- Willis, J.R.; Saus, E.; Iraola-Guzman, S.; Cabello-Yeves, E.; Ksiezopolska, E.; Cozzuto, L.; Bejarano, L.A.; Andreu-Somavilla, N.; Alloza-Trabado, M.; Blanco, A.; et al. Citizen-science based study of the oral microbiome in Cystic fibrosis and matched controls reveals major differences in diversity and abundance of bacterial and fungal species. J. Oral Microbiol. 2021, 13, 1897328. [Google Scholar] [CrossRef]
- Liu, H.; Hu, Z.; Li, M.; Yang, Y.; Lu, S.; Rao, X. Therapeutic potential of bacteriophage endolysins for infections caused by Gram-positive bacteria. J. Biomed. Sci. 2023, 30, 29. [Google Scholar] [CrossRef]
- Rashel, M.; Uchiyama, J.; Ujihara, T.; Uehara, Y.; Kuramoto, S.; Sugihara, S.; Yagyu, K.; Muraoka, A.; Sugai, M.; Hiramatsu, K.; et al. Efficient elimination of multidrug-resistant Staphylococcus aureus by cloned lysin derived from bacteriophage phi MR11. J. Infect. Dis. 2007, 196, 1237–1247. [Google Scholar] [CrossRef]
- Van Tassell, M.L.; Ibarra-Sanchez, L.A.; Hoepker, G.P.; Miller, M.J. Hot topic: Antilisterial activity by endolysin PlyP100 in fresh cheese. J. Dairy Sci. 2017, 100, 2482–2487. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cheng, M.; Zhang, H.; Dai, J.; Guo, Z.; Li, X.; Ji, Y.; Cai, R.; Xi, H.; Wang, X.; et al. Antibacterial Effects of Phage Lysin LysGH15 on Planktonic Cells and Biofilms of Diverse Staphylococci. Appl. Environ. Microbiol. 2018, 84, e00886-18. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Feng, C.; Ren, J.; Zhuang, X.; Zhang, Y.; Zhu, Y.; Dong, K.; He, P.; Guo, X.; Qin, J. A Novel Antimicrobial Endolysin, LysPA26, against Pseudomonas aeruginosa. Front. Microbiol. 2017, 8, 293. [Google Scholar] [CrossRef]
- Thurber, R.V.; Haynes, M.; Breitbart, M.; Wegley, L.; Rohwer, F. Laboratory procedures to generate viral metagenomes. Nat. Protoc. 2009, 4, 470–483. [Google Scholar] [CrossRef]
- Nurk, S.; Meleshko, D.; Korobeynikov, A.; Pevzner, P.A. metaSPAdes: A new versatile metagenomic assembler. Genome. Res. 2017, 27, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Kieft, K.; Zhou, Z.; Anantharaman, K. VIBRANT: Automated recovery, annotation and curation of microbial viruses, and evaluation of viral community function from genomic sequences. Microbiome 2020, 8, 90. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Zielezinski, A.; Deorowicz, S.; Gudys, A. PHIST: Fast and accurate prediction of prokaryotic hosts from metagenomic viral sequences. Bioinformatics 2022, 38, 1447–1449. [Google Scholar] [CrossRef] [PubMed]
- Sayers, E.W.; Beck, J.; Bolton, E.E.; Bourexis, D.; Brister, J.R.; Canese, K.; Comeau, D.C.; Funk, K.; Kim, S.; Klimke, W.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2021, 49, D10–D17. [Google Scholar] [CrossRef] [PubMed]
- Camargo, A.P.; Nayfach, S.; Chen, I.A.; Palaniappan, K.; Ratner, A.; Chu, K.; Ritter, S.J.; Reddy, T.B.K.; Mukherjee, S.; Schulz, F.; et al. IMG/VR v4: An expanded database of uncultivated virus genomes within a framework of extensive functional, taxonomic, and ecological metadata. Nucleic Acids Res. 2023, 51, D733–D743. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.Z.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed]
- Mirdita, M.; Schutze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making protein folding accessible to all. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef]
- Kabsch, W.; Sander, C. Dictionary of protein secondary structure: Pattern recognition of hydrogen-bonded and geometrical features. Biopolymers 1983, 22, 2577–2637. [Google Scholar] [CrossRef]
- Harhala, M.; Gembara, K.; Miernikiewicz, P.; Owczarek, B.; Kazmierczak, Z.; Majewska, J.; Nelson, D.C.; Dabrowska, K. DNA Dye Sytox Green in Detection of Bacteriolytic Activity: High Speed, Precision and Sensitivity Demonstrated With Endolysins. Front. Microbiol. 2021, 12, 752282. [Google Scholar] [CrossRef]
- van Meerloo, J.; Kaspers, G.J.; Cloos, J. Cell sensitivity assays: The MTT assay. Methods Mol. Biol. 2011, 731, 237–245. [Google Scholar]
- Yamane, K.; Nambu, T.; Yamanaka, T.; Mashimo, C.; Sugimori, C.; Leung, K.-P.; Fukushima, H. Complete Genome Sequence of Rothia mucilaginosa DY-18: A Clinical Isolate with Dense Meshwork-Like Structures from a Persistent Apical Periodontitis Lesion. Sequencing 2010, 2010, 457236. [Google Scholar] [CrossRef][Green Version]
- Yuan, Z.; Panchal, D.; Syed, M.A.; Mehta, H.; Joo, M.; Hadid, W.; Sadikot, R.T. Induction of cyclooxygenase-2 signaling by Stomatococcus mucilaginosus highlights the pathogenic potential of an oral commensal. J. Immunol. 2013, 191, 3810–3817. [Google Scholar] [CrossRef]
- Cha, Y.; Son, B.; Ryu, S. Effective removal of staphylococcal biofilms on various food contact surfaces by Staphylococcus aureus phage endolysin LysCSA13. Food Microbiol. 2019, 84, 103245. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, X.; Liang, S.; Wang, J.; Zhu, Y.; Zhang, W.; Liu, S.; Schwarz, S.; Xie, F. Bactericidal synergism between phage endolysin Ply2660 and cathelicidin LL-37 against vancomycin-resistant Enterococcus faecalis biofilms. NPJ Biofilms Microbiomes 2023, 9, 16. [Google Scholar] [CrossRef]
- Arroyo-Moreno, S.; Cummings, M.; Corcoran, D.B.; Coffey, A.; McCarthy, R.R. Identification and characterization of novel endolysins targeting Gardnerella vaginalis biofilms to treat bacterial vaginosis. NPJ Biofilms Microbiomes 2022, 8, 29. [Google Scholar] [CrossRef]
- Shen, Y.; Koller, T.; Kreikemeyer, B.; Nelson, D.C. Rapid degradation of Streptococcus pyogenes biofilms by PlyC, a bacteriophage-encoded endolysin. J. Antimicrob. Chemother. 2013, 68, 1818–1824. [Google Scholar] [CrossRef] [PubMed]
- Schuch, R.; Khan, B.K.; Raz, A.; Rotolo, J.A.; Wittekind, M. Bacteriophage Lysin CF-301, a Potent Antistaphylococcal Biofilm Agent. Antimicrob. Agents Chemother. 2017, 61, e02666-16. [Google Scholar] [CrossRef]
- Henwick, S.; Koehler, M.; Patrick, C.C. Complications of bacteremia due to Stomatococcus mucilaginosus in neutropenic children. Clin. Infect. Dis. 1993, 17, 667–671. [Google Scholar] [CrossRef] [PubMed]
- Fanourgiakis, P.; Georgala, A.; Vekemans, M.; Daneau, D.; Heymans, C.; Aoun, M. Bacteremia due to Stomatococcus mucilaginosus in neutropenic patients in the setting of a cancer institute. Clin. Microbiol. Infect. 2003, 9, 1068–1072. [Google Scholar] [CrossRef] [PubMed]
- Treviño, M.; García-Zabarte, A.; Quintás, A.; Varela, E.; López-Paz, J.M.; Jato, A.; García-Riestra, C.; Regueiro, B.J. Stomatococcus mucilaginosus Septicemia in a Patient with Acute Lymphoblastic Leukaemia. Eur. J. Clin. Microbiol. Infect. Dis. 1998, 17, 505–507. [Google Scholar] [CrossRef]
- Granlund, M.; Linderholm, M.; Norgren, M.; Olofsson, C.; Wahlin, A.; Holm, S.E. Stomatococcus mucilaginosus septicemia in leukemic patients. Clin. Microbiol. Infect. 1996, 2, 179–185. [Google Scholar] [CrossRef][Green Version]
- McWhinney, P.H.; Kibbler, C.C.; Gillespie, S.H.; Patel, S.; Morrison, D.; Hoffbrand, A.V.; Prentice, H.G. Stomatococcus mucilaginosus: An emerging pathogen in neutropenic patients. Clin. Infect. Dis. 1992, 14, 641–646. [Google Scholar] [CrossRef]
- Crowe, A.; Ding, N.S.; Yong, E.; Sheorey, H.; Waters, M.J.; Daffy, J. Rothia aeria mitral valve endocarditis complicated by multiple mycotic aneurysms: Laboratory identification expedited using MALDI-TOF MS. Infection 2014, 42, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Getzenberg, R.B.; Hijano, D.R.; Hakim, H.; Dallas, R.H.; Ferrolino, J.A.; de Cardenas, J.B.; Garner, C.D.; Tang, L.; Su, Y.; Wolf, J.; et al. Rothia mucilaginosa Infections in Pediatric Cancer Patients. J. Pediatr. Infect. Dis. Soc. 2021, 10, 341–344. [Google Scholar] [CrossRef] [PubMed]
- Chavan, R.S.; Pannaraj, P.S.; Luna, R.A.; Szabo, S.; Adesina, A.; Versalovic, J.; Krance, R.A.; Kennedy-Nasser, A.A. Significant morbidity and mortality attributable to Rothia mucilaginosa infections in children with hematological malignancies or following hematopoietic stem cell transplantation. Pediatr. Hematol. Oncol. 2013, 30, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Brossard, J.; Cellot, S.; Dix, D.; Feusner, J.; Johnston, D.L.; Lewis, V.; Mitchell, D.; Portwine, C.; Price, V.; et al. Invasive Rothia infections in children with acute myeloid leukemia: A report from the Canadian infections in AML research group. Pediatr. Hematol. Oncol. 2016, 33, 277–281. [Google Scholar] [CrossRef]
- Robertson, R.D.; Panigrahi, A.; Cheema, R. Rothia mucilaginosa bacteremia, meningitis leading to diffuse cerebritis in an adolescent patient undergoing acute myeloid leukemia chemotherapy causing significant morbidity. SAGE Open Med. Case Rep. 2021, 9. [Google Scholar] [CrossRef]
- Pinsky, R.L.; Piscitelli, V.; Patterson, J.E. Endocarditis caused by relatively penicillin-resistant Stomatococcus mucilaginosus. J. Clin. Microbiol. 1989, 27, 215–216. [Google Scholar] [CrossRef]
- Ascher, D.P.; Zbick, C.; White, C.; Fischer, G.W. Infections due to Stomatococcus mucilaginosus: 10 cases and review. Rev. Infect. Dis. 1991, 13, 1048–1052. [Google Scholar] [CrossRef]
- Binder, D.; Zbinden, R.; Widmer, U.; Opravil, M.; Krause, M. Native and prosthetic valve endocarditis caused by Rothia dentocariosa: Diagnostic and therapeutic considerations. Infection 1997, 25, 22–26. [Google Scholar] [CrossRef]
- Korsholm, T.L.; Haahr, V.; Prag, J. Eight cases of lower respiratory tract infection caused by Stomatococcus mucilaginosus. Scand. J. Infect. Dis. 2007, 39, 913–917. [Google Scholar] [CrossRef]
- Maraki, S.; Papadakis, I.S. Rothia mucilaginosa pneumonia: A literature review. Infect. Dis. 2015, 47, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Bruminhent, J.; Tokarczyk, M.J.; Jungkind, D.; DeSimone, J.A., Jr. Rothia mucilaginosa prosthetic device infections: A case of prosthetic valve endocarditis. J. Clin. Microbiol. 2013, 51, 1629–1632. [Google Scholar] [CrossRef]
- Cho, E.J.; Sung, H.; Park, S.J.; Kim, M.N.; Lee, S.O. Rothia mucilaginosa pneumonia diagnosed by quantitative cultures and intracellular organisms of bronchoalveolar lavage in a lymphoma patient. Ann. Lab. Med. 2013, 33, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Bittar, F.; Richet, H.; Dubus, J.C.; Reynaud-Gaubert, M.; Stremler, N.; Sarles, J.; Raoult, D.; Rolain, J.M. Molecular detection of multiple emerging pathogens in sputa from cystic fibrosis patients. PLoS ONE 2008, 3, e2908. [Google Scholar] [CrossRef] [PubMed]
- Guss, A.M.; Roeselers, G.; Newton, I.L.; Young, C.R.; Klepac-Ceraj, V.; Lory, S.; Cavanaugh, C.M. Phylogenetic and metabolic diversity of bacteria associated with cystic fibrosis. ISME J. 2011, 5, 20–29. [Google Scholar] [CrossRef]
- Liepinsh, E.; Genereux, C.; Dehareng, D.; Joris, B.; Otting, G. NMR structure of Citrobacter freundii AmpD, comparison with bacteriophage T7 lysozyme and homology with PGRP domains. J. Mol. Biol. 2003, 327, 833–842. [Google Scholar] [CrossRef]
- National Library of Medicine, National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/bioproject/PRJNA934363 (accessed on 28 June 2023).






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miernikiewicz, P.; Barylski, J.; Wilczak, A.; Dragoš, A.; Rybicka, I.; Bałdysz, S.; Szymczak, A.; Dogsa, I.; Rokush, K.; Harhala, M.A.; et al. New Phage-Derived Antibacterial Enzyme PolaR Targeting Rothia spp. Cells 2023, 12, 1997. https://doi.org/10.3390/cells12151997
Miernikiewicz P, Barylski J, Wilczak A, Dragoš A, Rybicka I, Bałdysz S, Szymczak A, Dogsa I, Rokush K, Harhala MA, et al. New Phage-Derived Antibacterial Enzyme PolaR Targeting Rothia spp. Cells. 2023; 12(15):1997. https://doi.org/10.3390/cells12151997
Chicago/Turabian StyleMiernikiewicz, Paulina, Jakub Barylski, Aleksandra Wilczak, Anna Dragoš, Izabela Rybicka, Sophia Bałdysz, Aleksander Szymczak, Iztok Dogsa, Kostiantyn Rokush, Marek Adam Harhala, and et al. 2023. "New Phage-Derived Antibacterial Enzyme PolaR Targeting Rothia spp." Cells 12, no. 15: 1997. https://doi.org/10.3390/cells12151997
APA StyleMiernikiewicz, P., Barylski, J., Wilczak, A., Dragoš, A., Rybicka, I., Bałdysz, S., Szymczak, A., Dogsa, I., Rokush, K., Harhala, M. A., Ciekot, J., Ferenc, S., Gnus, J., Witkiewicz, W., & Dąbrowska, K. (2023). New Phage-Derived Antibacterial Enzyme PolaR Targeting Rothia spp. Cells, 12(15), 1997. https://doi.org/10.3390/cells12151997