
Co-Aggregation and Parallel Aggregation of Specific Proteins in Major Mental Illness

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
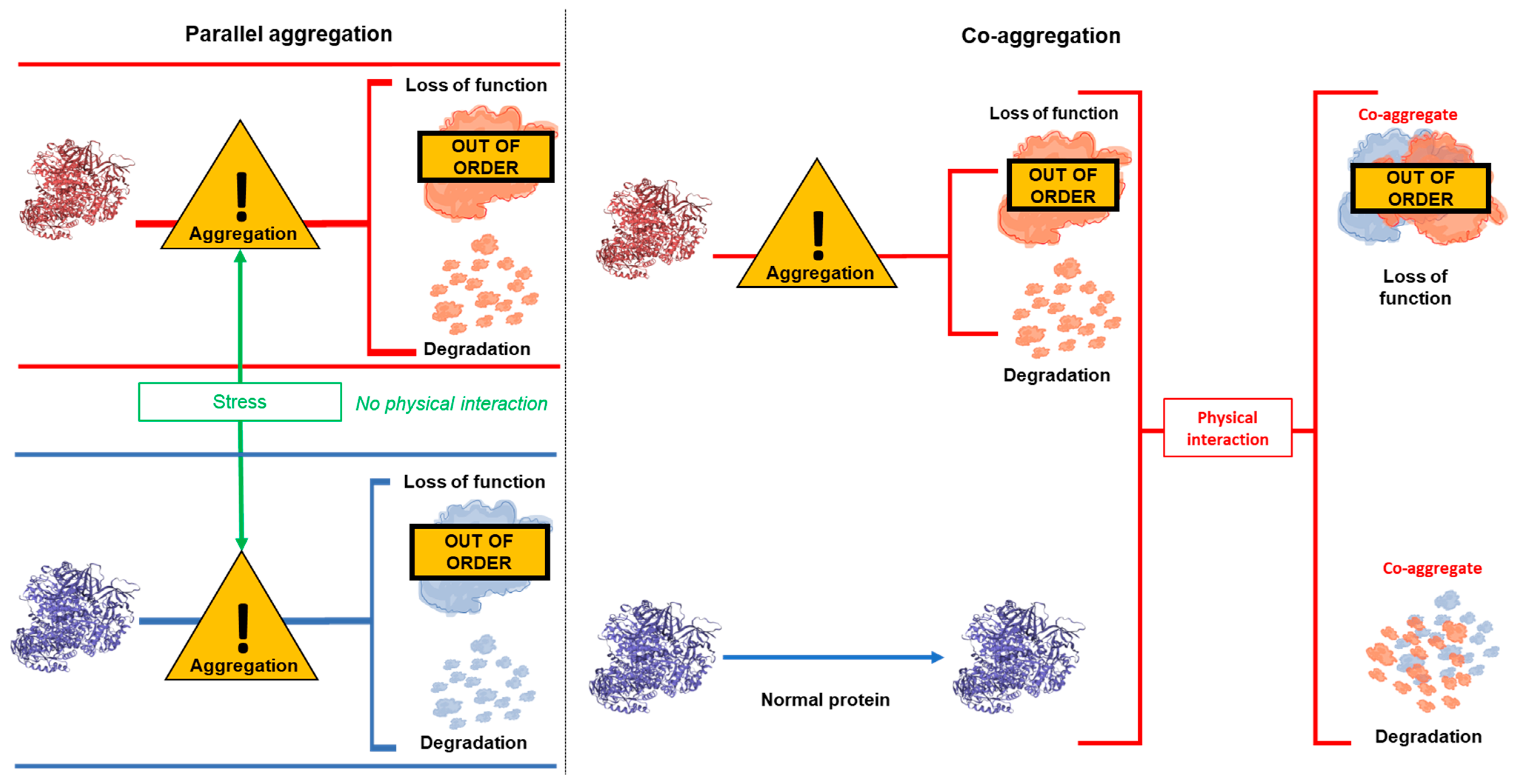
1. Introduction
2. Materials and Methods
2.1. Human Brain Tissue
2.2. Insolubility Assay
2.3. Antibodies
2.4. Plasmids
2.5. Cell Culture
2.6. Western Blotting
2.7. Immunocytochemistry and Microscopy
3. Results
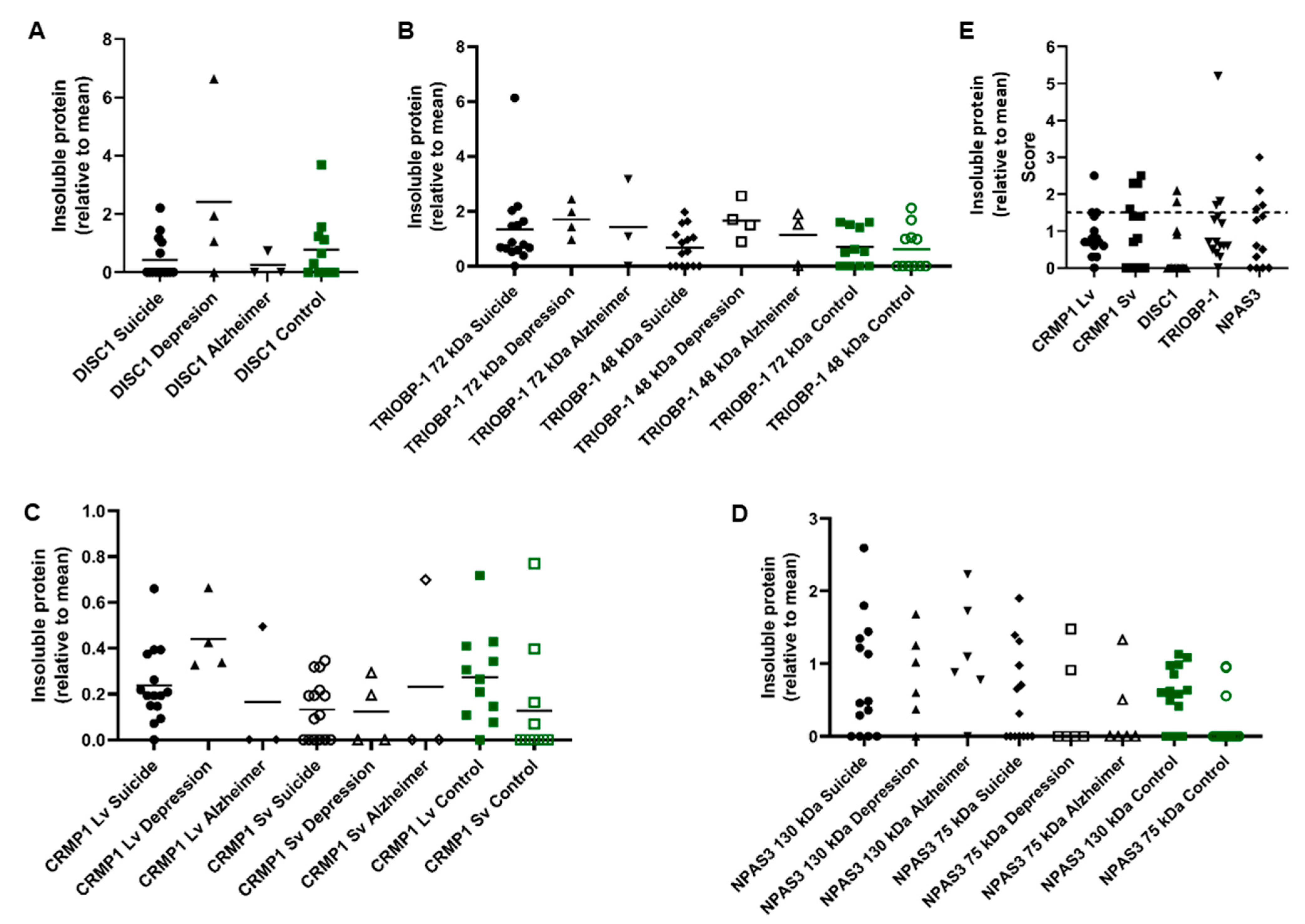
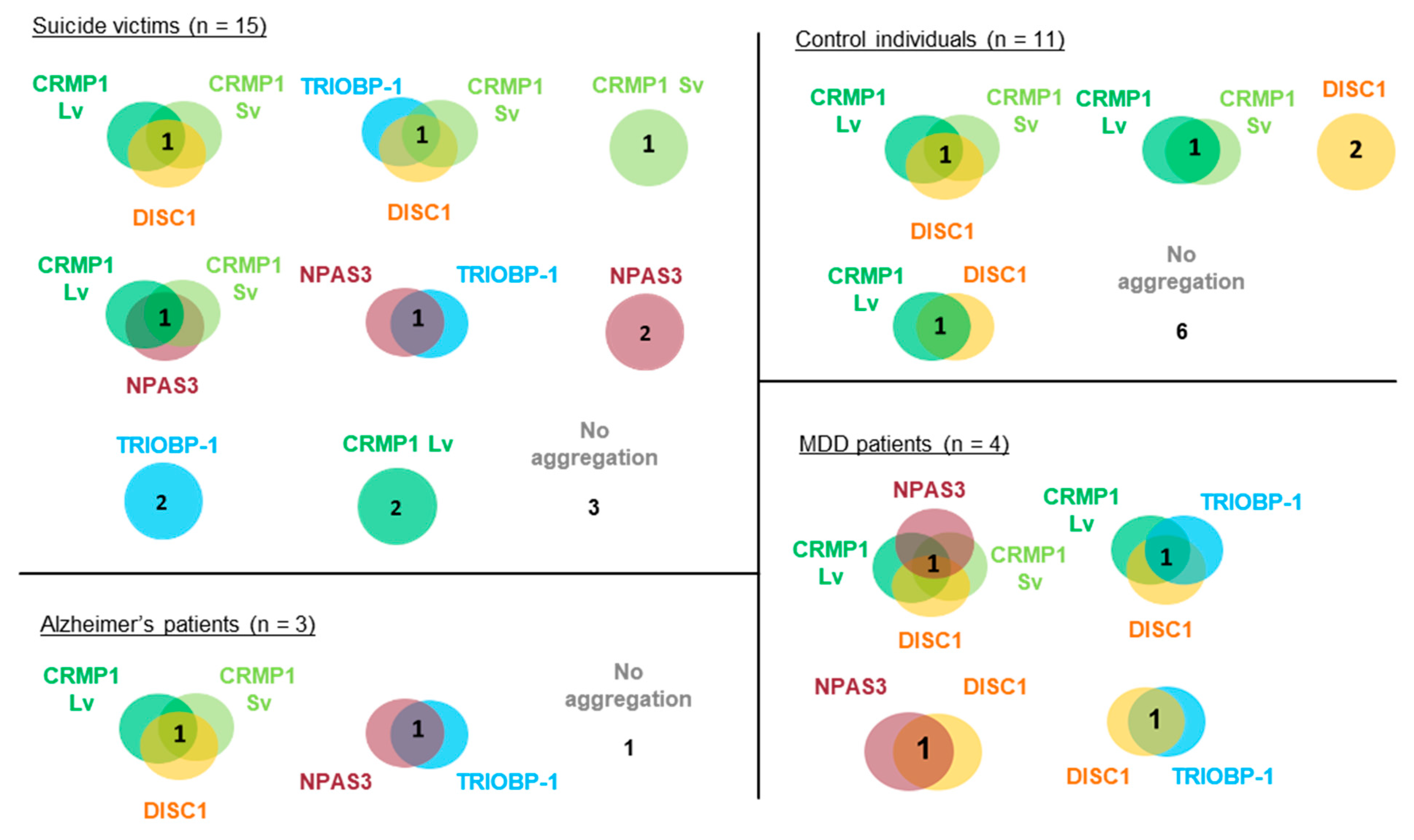
3.1. Presence of Multiple Insoluble Proteins in Individual Brain Samples
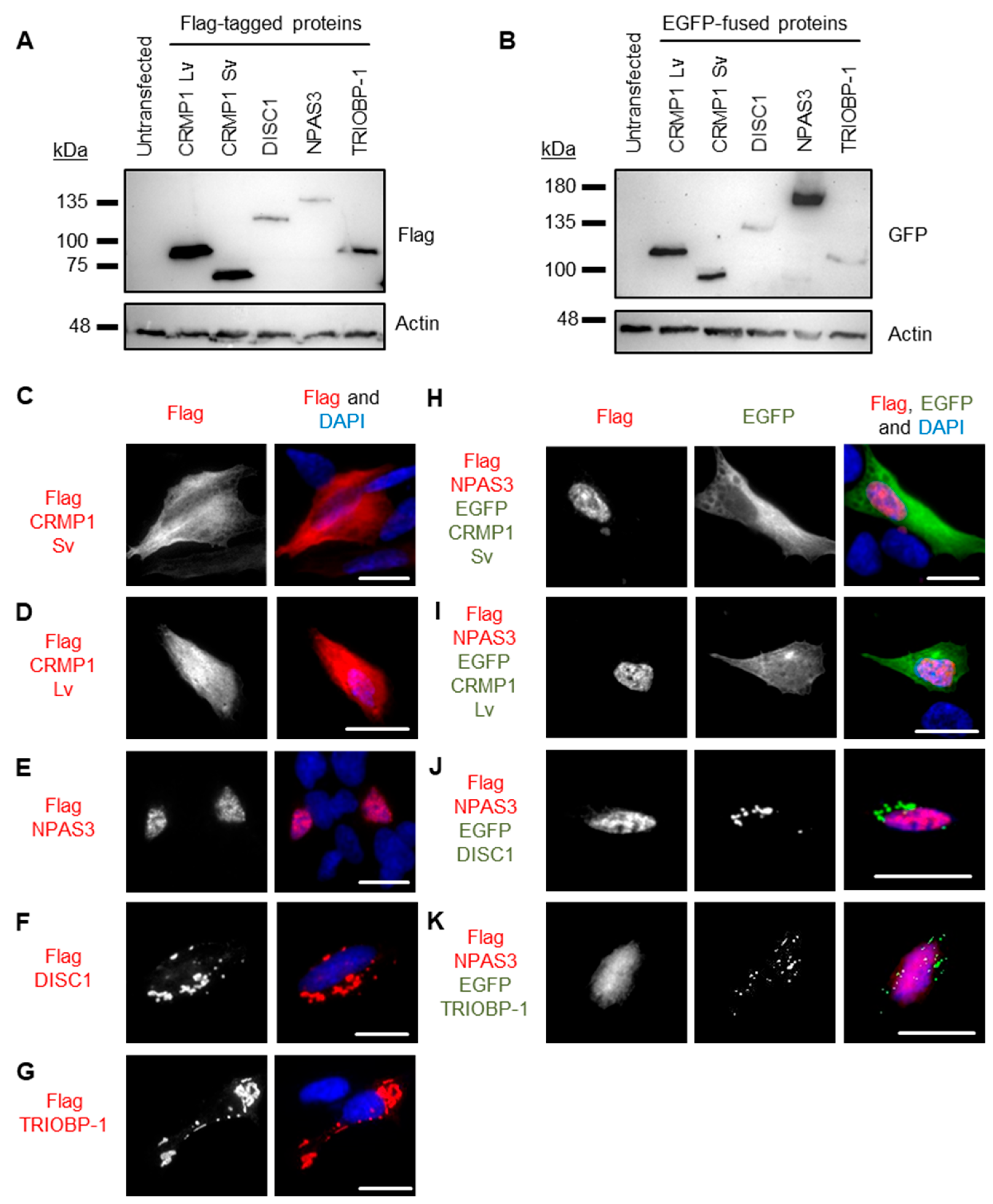
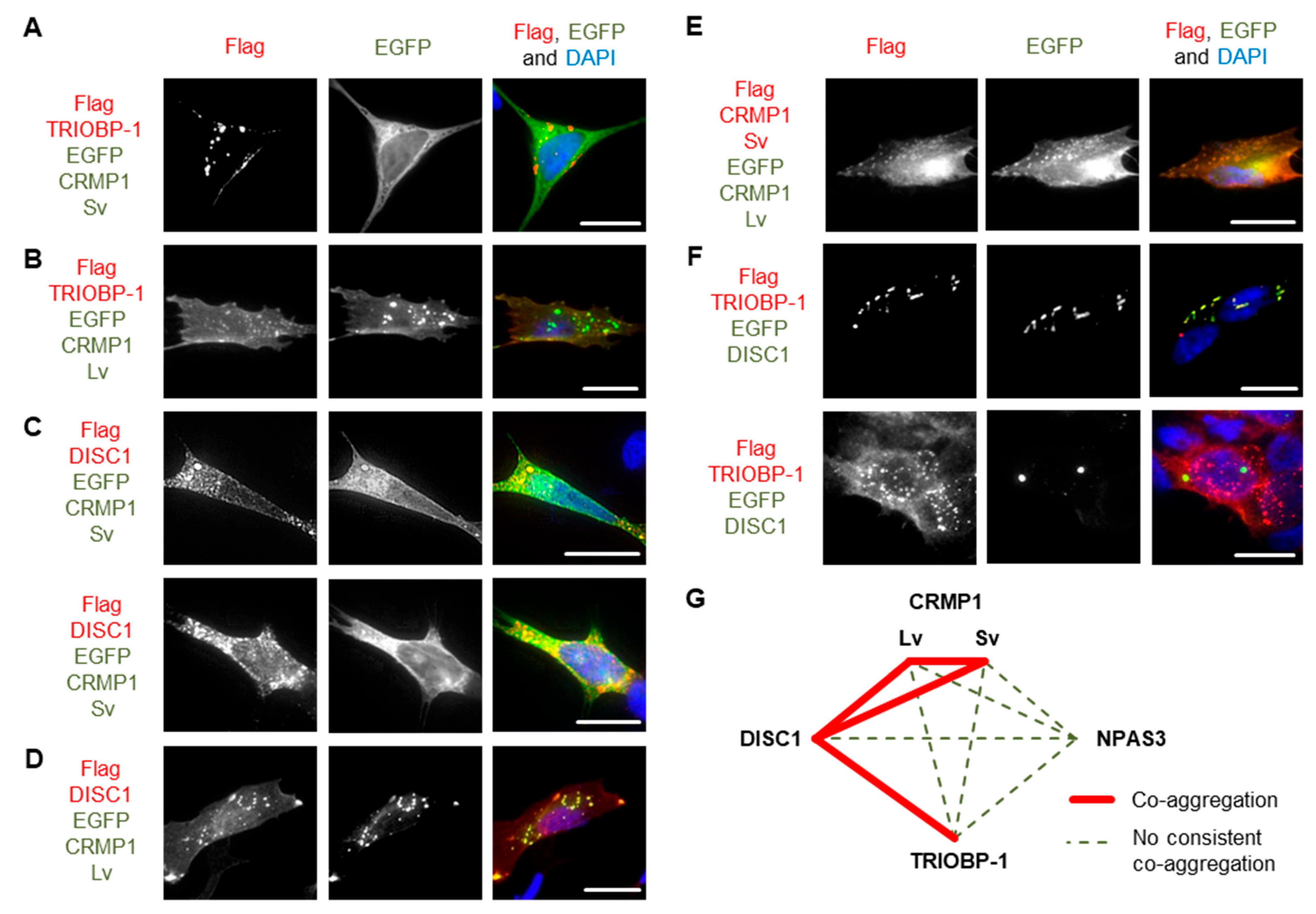
3.2. Pairwise Co-Expression Studies in Neuroblastoma Cells Show only DISC1 Readily Forms Co-Aggregates, with Both CRMP1 and TRIOBP-1
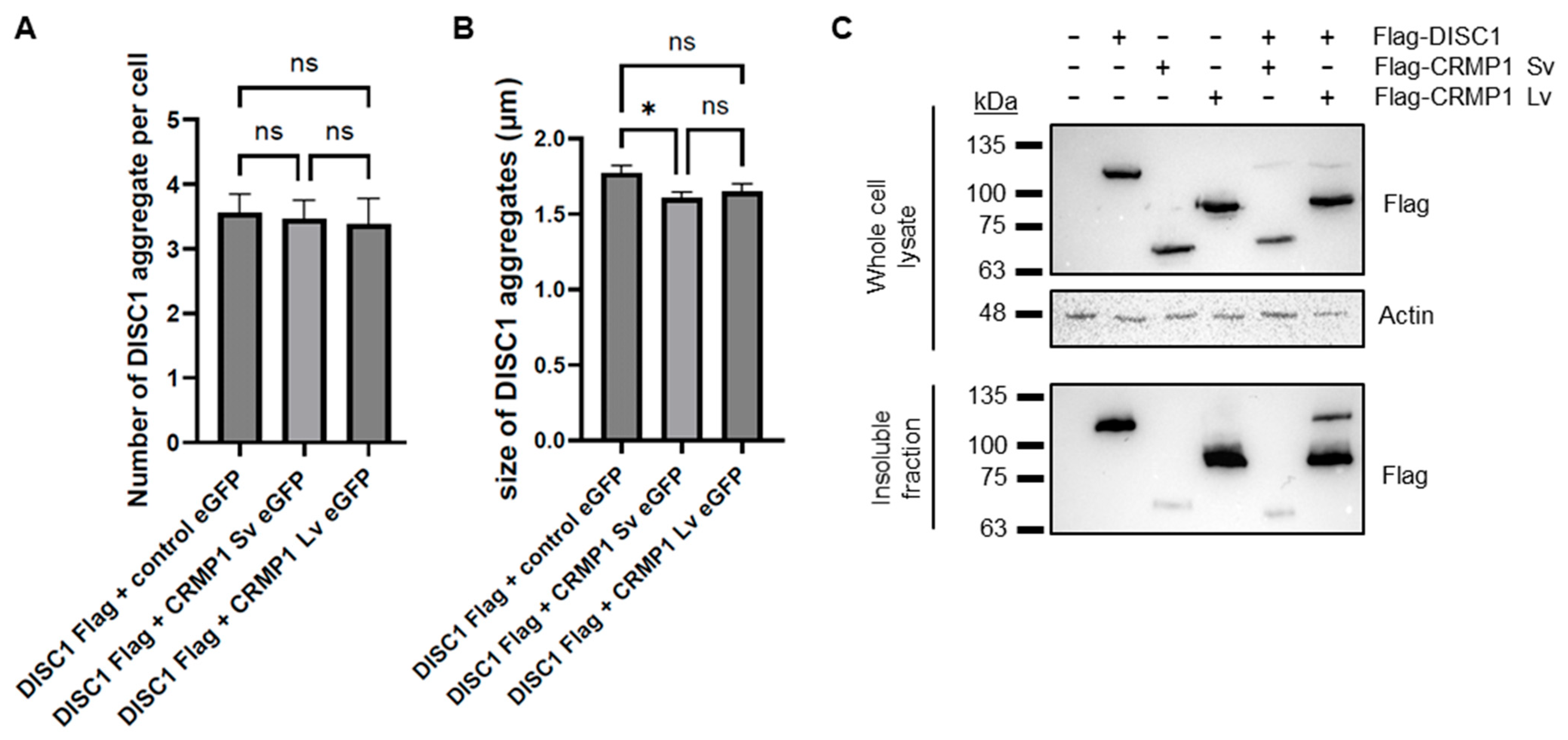
3.3. No Obvious Effect of CRMP1 on the Extent of DISC1 Aggregation
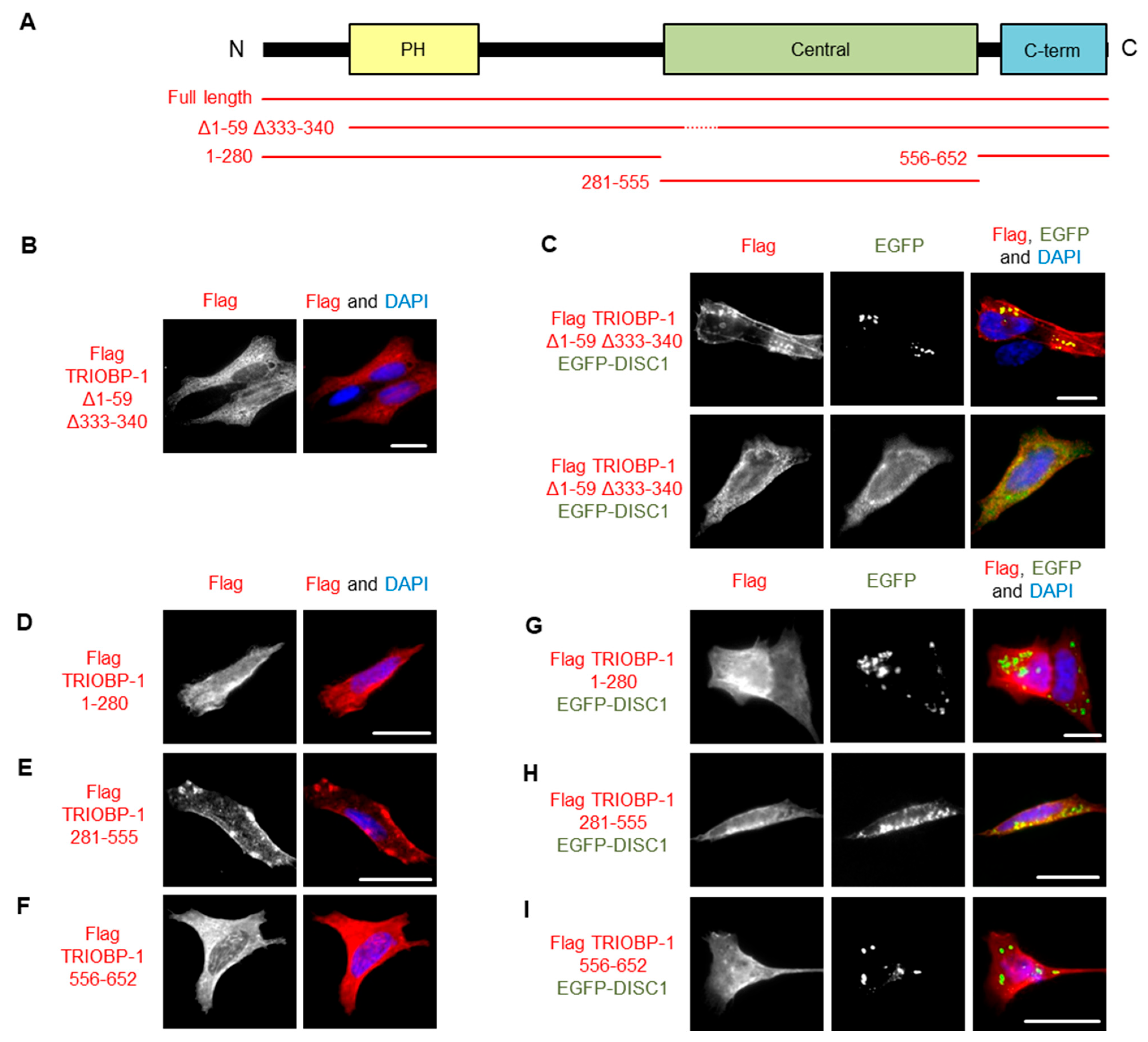
3.4. DISC1 Can Induce Aggregation of TRIOBP-1, with No Individual Domain of TRIOBP-1 Sufficient for This
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wray, N.R.; Ripke, S.; Mattheisen, M.; Trzaskowski, M.; Byrne, E.M.; Abdellaoui, A.; Adams, M.J.; Agerbo, E.; Air, T.M.; Andlauer, T.M.F.; et al. Genome-wide association analyses identify 44 risk variants and refine the genetic architecture of major depression. Nat. Genet. 2018, 50, 668–681. [Google Scholar] [CrossRef] [PubMed]
- Stahl, E.A.; Breen, G.; Forstner, A.J.; McQuillin, A.; Ripke, S.; Trubetskoy, V.; Mattheisen, M.; Wang, Y.; Coleman, J.R.I.; Gaspar, H.A.; et al. Genome-wide association study identifies 30 loci associated with bipolar disorder. Nat. Genet. 2019, 51, 793–803. [Google Scholar] [CrossRef]
- Trubetskoy, V.; Pardiñas, A.F.; Qi, T.; Panagiotaropoulou, G.; Awasthi, S.; Bigdeli, T.B.; Bryois, J.; Chen, C.-Y.; Dennison, C.A.; Hall, L.S.; et al. Mapping genomic loci implicates genes and synaptic biology in schizophrenia. Nature 2022, 604, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, N.J.; Korth, C. Protein misassembly and aggregation as potential convergence points for non-genetic causes of chronic mental illness. Mol. Psychiatry 2019, 24, 936–951. [Google Scholar] [CrossRef] [PubMed]
- Leliveld, S.R.; Bader, V.; Hendriks, P.; Prikulis, I.; Sajnani, G.; Requena, J.R.; Korth, C. Insolubility of Disrupted-in-Schizophrenia 1 disrupts oligomer-dependent interactions with Nuclear Distribution Element 1 and is associated with sporadic mental disease. J. Neurosci. 2008, 28, 3839–3845. [Google Scholar] [CrossRef]
- Zaharija, B.; Odorčić, M.; Hart, A.; Samardžija, B.; Marreiros, R.; Prikulis, I.; Juković, M.; Hyde, T.M.; Kleinman, J.E.; Korth, C.; et al. TRIOBP-1 protein aggregation exists in both major depressive disorder and schizophrenia, and can occur through two distinct regions of the protein. Int. J. Mol. Sci. 2022, 23, 11048. [Google Scholar] [CrossRef]
- Ottis, P.; Bader, V.; Trossbach, S.V.; Kretzschmar, H.; Michel, M.; Leliveld, S.R.; Korth, C. Convergence of two independent mental disease genes on the protein level: Recruitment of dysbindin to cell-invasive Disrupted-in-Schizophrenia 1 aggresomes. Biol. Psychiatry 2011, 70, 604–610. [Google Scholar] [CrossRef]
- Leliveld, S.R.; Hendriks, P.; Michel, M.; Sajnani, G.; Bader, V.; Trossbach, S.; Prikulis, I.; Hartmann, R.; Jonas, E.; Willbold, D.; et al. Oligomer assembly of the C-terminal DISC1 domain (640–854) is controlled by self-association motifs and disease-associated polymorphism S704C. Biochemistry 2009, 48, 7746–7755. [Google Scholar] [CrossRef]
- Bader, V.; Tomppo, L.; Trossbach, S.V.; Bradshaw, N.J.; Prikulis, I.; Leliveld, S.R.; Lin, C.-Y.; Ishizuka, K.; Sawa, A.; Ramos, A.; et al. Proteomic, genomic and translational approaches identify CRMP1 for a role in schizophrenia and its underlying traits. Hum. Mol. Genet. 2012, 21, 4406–4418. [Google Scholar] [CrossRef]
- Bradshaw, N.J.; Bader, V.; Prikulis, I.; Lueking, A.; Müllner, S.; Korth, C. Aggregation of the protein TRIOBP-1 and its potential relevance to schizophrenia. PLoS ONE 2014, 9, e111196. [Google Scholar] [CrossRef]
- Yu, L.; Arbez, N.; Nucifora, L.G.; Sell, G.L.; DeLisi, L.E.; Ross, C.A.; Margolis, R.L.; Nucifora, F.C. A mutation in NPAS3 segregates with mental illness in a small family. Mol. Psychiatry 2014, 19, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Nucifora, L.G.; Wu, Y.C.; Lee, B.J.; Sha, L.; Margolis, R.L.; Ross, C.A.; Sawa, A.; Nucifora, F.C., Jr. A mutation in NPAS3 that segregates with schizophrenia in a small family leads to protein aggregation. Mol. Neuropsychiatry 2016, 2, 133–144. [Google Scholar] [CrossRef]
- Samardžija, B.; Pavešić Radonja, A.; Zaharija, B.; Bergman, M.; Renner, É.; Palkovits, M.; Rubeša, G.; Bradshaw, N.J. Protein aggregation of NPAS3, implicated in mental illness, is not limited to the V304I mutation. J. Pers. Med. 2021, 11, 1070. [Google Scholar] [CrossRef]
- Bousman, C.A.; Luza, S.; Mancuso, S.G.; Kang, D.; Opazo, C.M.; Mostaid, M.S.; Cropley, V.; McGorry, P.; Weickert, C.S.; Pantelis, C.; et al. Elevated ubiquitinated proteins in brain and blood of individuals with schizophrenia. Sci. Rep. 2019, 9, 2307. [Google Scholar] [CrossRef]
- Nucifora, L.G.; MacDonald, M.L.; Lee, B.J.; Peters, M.E.; Norris, A.L.; Orsburn, B.C.; Yang, K.; Gleason, K.; Margolis, R.L.; Pevsner, J.; et al. Increased protein insolubility in brains from a subset of patients with schizophrenia. Am. J. Psychiatry 2019, 176, 730–743. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.R.; Meador-Woodruff, J.H. Intracellular compartment-specific proteasome dysfunction in postmortem cortex in schizophrenia patients. Mol. Psychiatry 2020, 25, 776–790. [Google Scholar] [CrossRef]
- Palkovits, M. Isolated removal of hypothalamic or other brain nuclei of the rat. Brain Res. 1973, 59, 449–450. [Google Scholar] [CrossRef]
- Palkovits, M. Microdissection of individual brain nuclei and areas. In General Neurochemical Techniques; Neuromethods; Boulton, A.A., Baker, G.B., Eds.; Humana Press: Clifton, NJ, USA, 1985; pp. 1–17. [Google Scholar]
- Bradshaw, N.J.; Yerabham, A.S.K.; Marreiros, R.; Zhang, T.; Nagel-Steger, L.; Korth, C. An unpredicted aggregation-critical region of the actin-polymerizing protein TRIOBP-1/Tara, determined by elucidation of its domain structure. J. Biol. Chem. 2017, 292, 9583–9598. [Google Scholar] [CrossRef] [PubMed]
- Campeau, E.; Ruhl, V.E.; Rodier, F.; Smith, C.L.; Rahmberg, B.L.; Fuss, J.O.; Campisi, J.; Yaswen, P.; Cooper, P.K.; Kaufman, P.D. A versatile viral system for expression and depletion of proteins in mammalian cells. PLoS ONE 2009, 4, e6529. [Google Scholar] [CrossRef] [PubMed]
- Seiler, C.Y.; Park, J.G.; Sharma, A.; Hunter, P.; Surapaneni, P.; Sedillo, C.; Field, J.; Algar, R.; Price, A.; Steel, J.; et al. DNASU plasmid and PSI:Biology-Materials repositories: Resources to accelerate biological research. Nucleic Acids Res. 2013, 42, D1253–D1260. [Google Scholar] [CrossRef]
- Wiemann, S.; Pennacchio, C.; Hu, Y.; Hunter, P.; Harbers, M.; Amiet, A.; Bethel, G.; Busse, M.; Carninci, P.; Diekhans, M.; et al. The ORFeome Collaboration: A genome-scale human ORF-clone resource. Nat. Methods 2016, 13, 191–192. [Google Scholar] [CrossRef]
- Agrotis, A.; Pengo, N.; Burden, J.J.; Ketteler, R. Redundancy of human ATG4 protease isoforms in autophagy and LC3/GABARAP processing revealed in cells. Autophagy 2019, 15, 976–997. [Google Scholar] [CrossRef]
- Gogolla, N. The insular cortex. Curr. Biol. 2017, 27, R573–R591. [Google Scholar] [CrossRef] [PubMed]
- Stroedicke, M.; Bounab, Y.; Strempel, N.; Klockmeier, K.; Yigit, S.; Friedrich, R.P.; Chaurasia, G.; Li, S.; Hesse, F.; Riechers, S.-P.; et al. Systematic interaction network filtering identifies CRMP1 as a novel suppressor of huntingtin misfolding and neurotoxicity. Genome Res. 2015, 25, 701–713. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Ishizuka, K.; Nekooki-Machida, Y.; Endo, R.; Takashima, N.; Sasaki, H.; Komi, Y.; Gathercole, A.; Huston, E.; Ishii, K.; et al. Aggregation of scaffolding protein DISC1 dysregulates phosphodiesterase 4 in Huntington’s disease. J. Clin. Investig. 2017, 127, 1438–1450. [Google Scholar] [CrossRef]
- Endo, R.; Takashima, N.; Nekooki-Machida, Y.; Komi, Y.; Hui, K.K.-W.; Takao, M.; Akatsu, H.; Murayama, S.; Sawa, A.; Tanaka, M. TDP-43 and DISC1 co-aggregation disrupts dendritic local translation and mental function in FTLD. Biol. Psychiatry 2018, 84, 509–521. [Google Scholar] [CrossRef]
- Seipel, K.; O’Brien, S.P.; Iannotti, E.; Medley, Q.G.; Streuli, M. Tara, a novel F-actin binding protein, associates with the Trio guanine nucleotide exchange factor and regulates actin cytoskeletal organization. J. Cell Sci. 2001, 114, 389–399. [Google Scholar] [CrossRef]
- Enomoto, A.; Asai, N.; Namba, T.; Wang, Y.; Kato, T.; Tanaka, M.; Tatsumi, H.; Taya, S.; Tsuboi, D.; Kuroda, K.; et al. Roles of Disrupted-In-Schizophrenia 1-interacting protein Girdin in postnatal development of the dentate gyrus. Neuron 2009, 63, 774–787. [Google Scholar] [CrossRef]
- Kim, J.Y.; Duan, X.; Liu, C.Y.; Jang, M.-H.; Guo, J.U.; Pow-anpongkul, N.; Kang, E.; Song, H.; Ming, G.-l. DISC1 regulates new neuron development in the adult brain via modulation of AKT-mTOR signaling through KIAA1212. Neuron 2009, 63, 761–773. [Google Scholar] [CrossRef]
- Millar, J.K.; Christie, S.; Porteous, D.J. Yeast two-hybrid screens implicate DISC1 in brain development and function. Biochem. Biophys. Res. Comm. 2003, 311, 1019–1025. [Google Scholar] [CrossRef]
- Morris, J.A.; Kandpal, G.; Ma, L.; Austin, C.P. DISC1 (Disrupted-In-Schizophrenia 1) is a centrosome-associated protein that interacts with MAP1A, MIPT3, ATF4/5 and NUDEL: Regulation and loss of interaction with mutation. Hum. Mol. Genet. 2003, 12, 1591–1608. [Google Scholar] [CrossRef] [PubMed]
- Ozeki, Y.; Tomoda, T.; Kleiderlein, J.; Kamiya, A.; Bord, L.; Fujii, K.; Okawa, M.; Yamada, N.; Hatten, M.E.; Snyder, S.H.; et al. Disrupted-in-Schizophrenia-1 (DISC-1): Mutant truncation prevents binding to NudE-like (NUDEL) and inhibits neurite outgrowth. Proc. Natl. Acad. Sci. USA 2003, 100, 289–294. [Google Scholar] [CrossRef]
- Brandon, N.J.; Handford, E.J.; Schurov, I.; Rain, J.-C.; Pelling, M.; Duran-Jimeriz, B.; Camargo, L.M.; Oliver, K.R.; Beher, D.; Shearman, M.S.; et al. Disrupted in Schizophrenia 1 and Nudel form a neurodevelopmentally regulated protein complex: Implications for schizophrenia and other major neurological disorders. Mol. Cell. Neurosci. 2004, 25, 42–55. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.-H.; Kwak, Y.; Woo, Y.; Park, C.; Lee, S.-A.; Lee, H.; Park, S.J.; Suh, Y.; Suh, B.K.; Goo, B.S.; et al. Regulation of the actin cytoskeleton by the Ndel1-Tara complex is critical for cell migration. Sci. Rep. 2016, 6, 31827. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-Y.; Huang, P.-H.; Cheng, H.-J. Disrupted-in-Schizophrenia 1-mediated axon guidance involves TRIO-RAC-PAK small GTPase pathway signaling. Proc. Natl. Acad. Sci. USA 2011, 108, 5861–5866. [Google Scholar] [CrossRef] [PubMed]
- Yerabham, A.S.K.; Weiergräber, O.H.; Bradshaw, N.J.; Korth, C. Revisiting Disrupted in Schizophrenia 1 as a scaffold protein. Biol. Chem. 2013, 394, 1425–1437. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samardžija, B.; Juković, M.; Zaharija, B.; Renner, É.; Palkovits, M.; Bradshaw, N.J. Co-Aggregation and Parallel Aggregation of Specific Proteins in Major Mental Illness. Cells 2023, 12, 1848. https://doi.org/10.3390/cells12141848
Samardžija B, Juković M, Zaharija B, Renner É, Palkovits M, Bradshaw NJ. Co-Aggregation and Parallel Aggregation of Specific Proteins in Major Mental Illness. Cells. 2023; 12(14):1848. https://doi.org/10.3390/cells12141848
Chicago/Turabian StyleSamardžija, Bobana, Maja Juković, Beti Zaharija, Éva Renner, Miklós Palkovits, and Nicholas J. Bradshaw. 2023. "Co-Aggregation and Parallel Aggregation of Specific Proteins in Major Mental Illness" Cells 12, no. 14: 1848. https://doi.org/10.3390/cells12141848
APA StyleSamardžija, B., Juković, M., Zaharija, B., Renner, É., Palkovits, M., & Bradshaw, N. J. (2023). Co-Aggregation and Parallel Aggregation of Specific Proteins in Major Mental Illness. Cells, 12(14), 1848. https://doi.org/10.3390/cells12141848