
Effects of the Light/Dark Phase and Constant Light on Spatial Working Memory and Spine Plasticity in the Mouse Hippocampus
 , ,
, ,  and
and 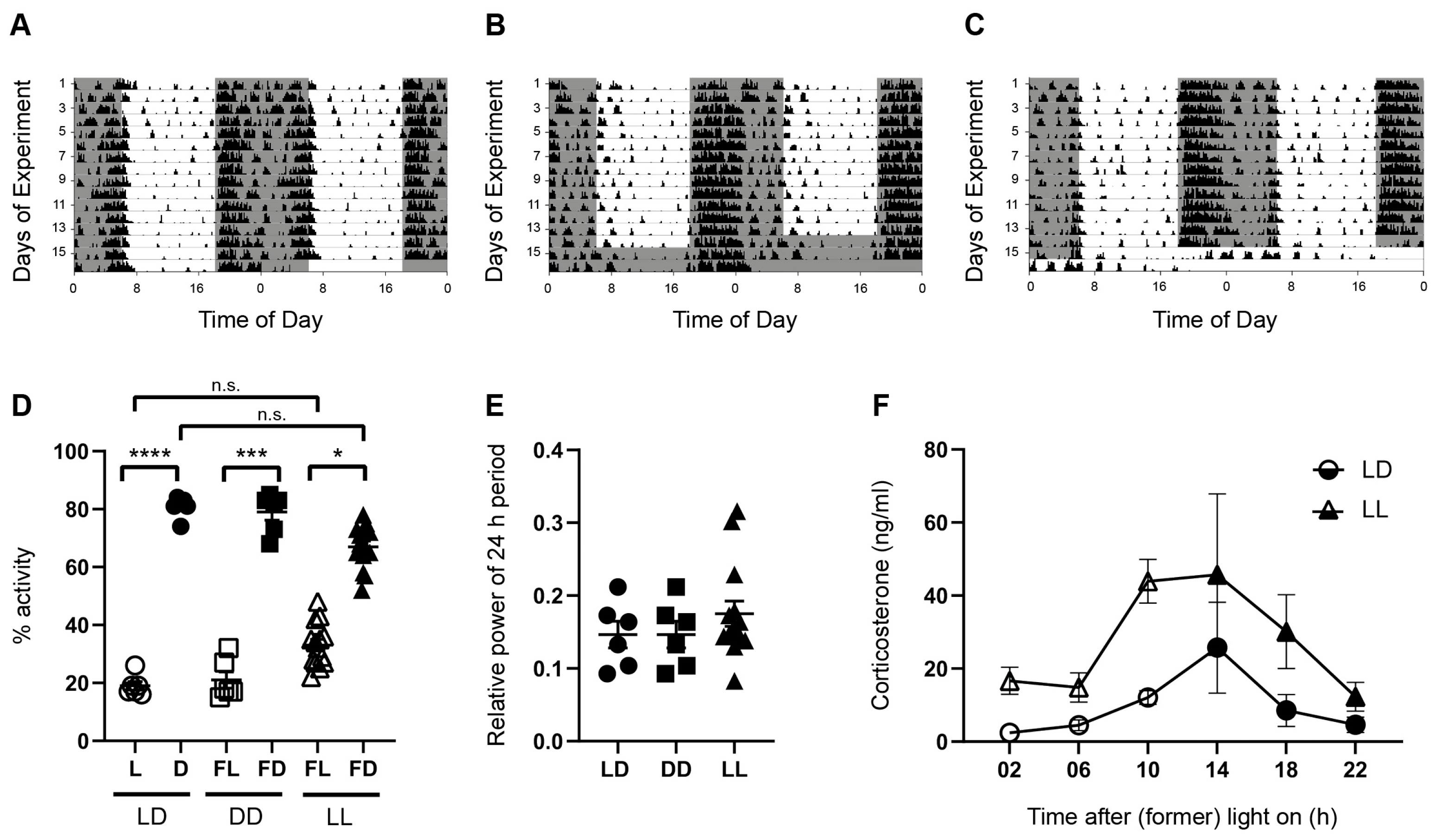{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Analysis of Spontaneous Locomotor Activity
2.3. Corticosterone Assay
2.4. Exploration Activity and Hippocampus-Dependent Spatial Working Memory
2.5. Immunofluorescence and Quantitative Analysis
2.6. Golgi Staining
2.7. Statistical Analysis
3. Results
3.1. Spontaneous Locomotor Activity Is Rhythmic under Acute Constant Darkness and Acute Constant Light
3.2. Hippocampus-Dependent Spatial Working Memory Is Affected by the Light Phase and by Acute Constant Light
3.3. Constant Light Affects Hippocampal Spine Morphology
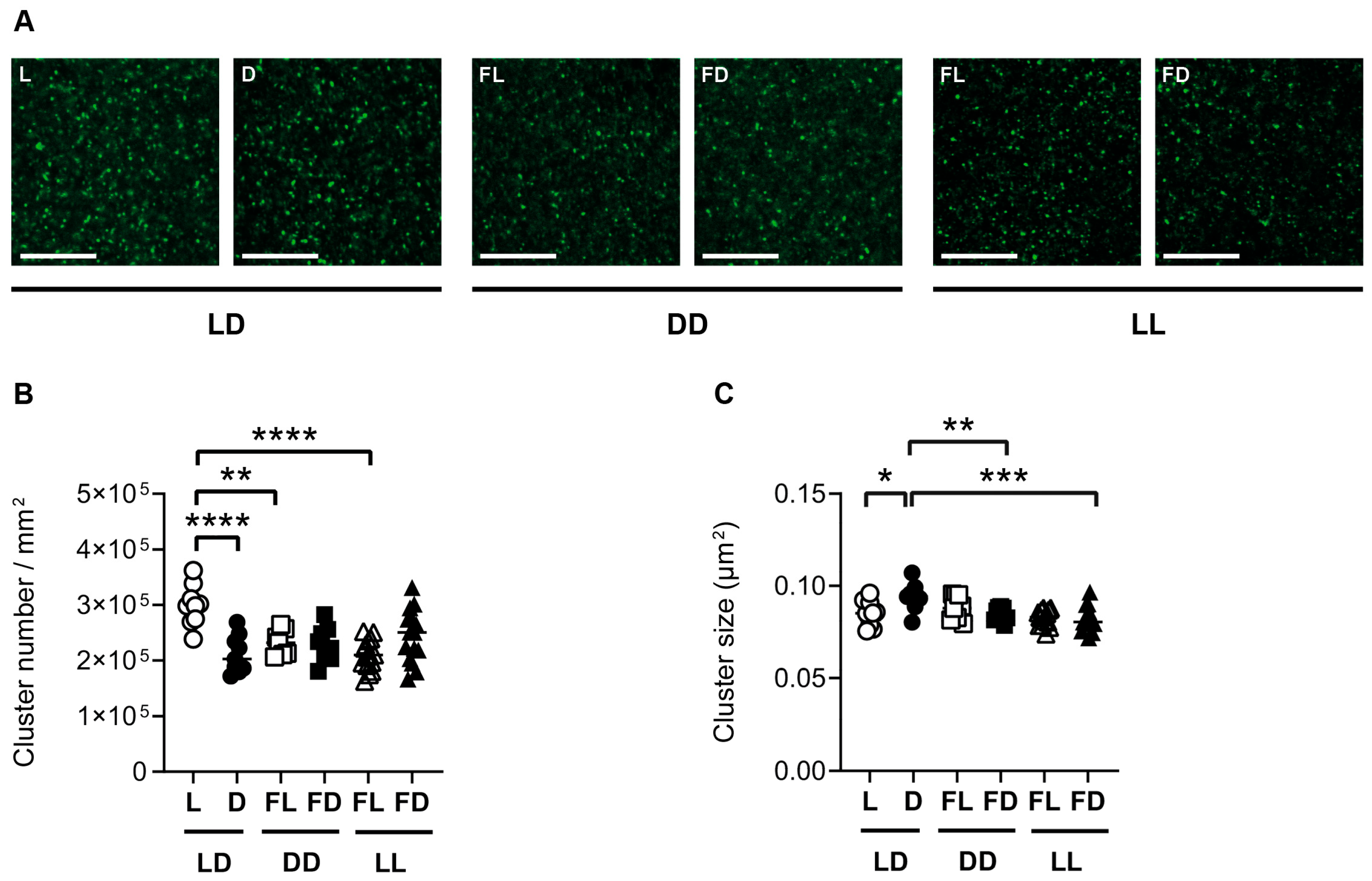
3.4. Hippocampal Synaptopodin Is Affected by the Light/Dark Phase and Acute Constant Light
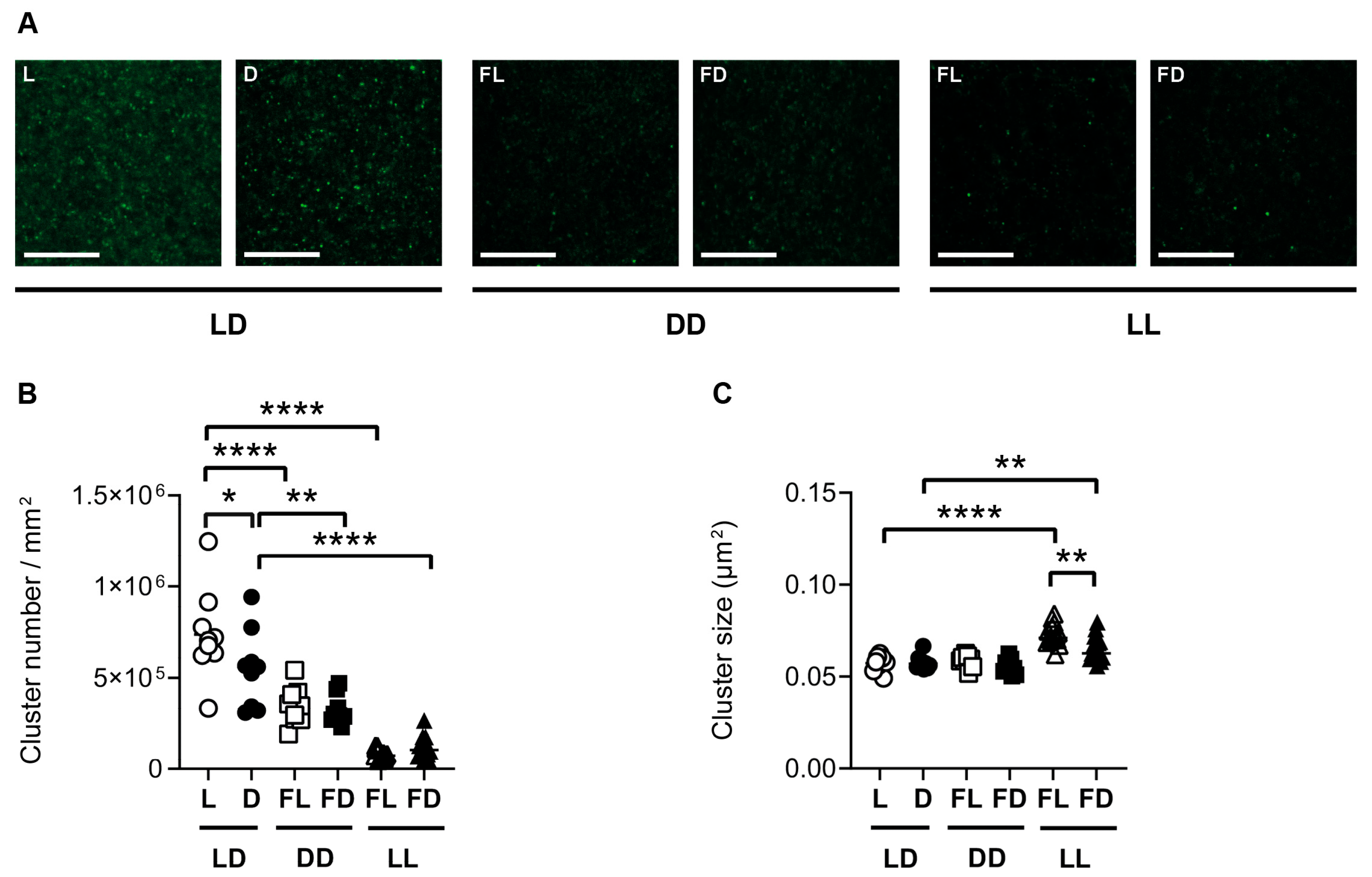
3.5. Hippocampal GluR1 Is Affected by the Light/Dark Phase and Acute Constant Light
4. Discussion
4.1. Effect of Light/Dark Cycle
4.2. Effect of Acute Constant Light
4.3. Limitations and Outlook
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Korf, H.-W.; von Gall, C. Circadian Physiology. In Neuroscience in the 21st Century, 3rd ed.; Pfaff, D.W., Volkow, N.D., Eds.; Springer Science + Business Media: Berlin/Heidelberg, Germany, 2021. [Google Scholar]
- Reppert, S.M.; Weaver, D.R. Coordination of circadian timing in mammals. Nature 2002, 418, 935–941. [Google Scholar] [CrossRef]
- Balsalobre, A.; Brown, S.A.; Marcacci, L.; Tronche, F.; Kellendonk, C.; Reichardt, H.M.; Schütz, G.; Schibler, U. Resetting of circadian time in peripheral tissues by glucocorticoid signaling. Science 2000, 289, 2344–2347. [Google Scholar] [CrossRef]
- Mrosovsky, N. Masking: History, definitions, and measurement. Chronobiol. Int. 1999, 16, 415–429. [Google Scholar] [CrossRef] [PubMed]
- von Gall, C. The Effects of Light and the Circadian System on Rhythmic Brain Function. Int. J. Mol. Sci. 2022, 23, 2778. [Google Scholar] [CrossRef] [PubMed]
- Hattar, S.; Liao, H.W.; Takao, M.; Berson, D.M.; Yau, K.W. Melanopsin-containing retinal ganglion cells: Architecture, projections, and intrinsic photosensitivity. Science 2002, 295, 1065–1070. [Google Scholar] [CrossRef] [PubMed]
- Provencio, I.; Rodriguez, I.R.; Jiang, G.; Hayes, W.P.; Moreira, E.F.; Rollag, M.D. A novel human opsin in the inner retina. J. Neurosci. 2000, 20, 600–605. [Google Scholar] [CrossRef]
- Hattar, S.; Kumar, M.; Park, A.; Tong, P.; Tung, J.; Yau, K.-W.; Berson, D.M. Central projections of melanopsin-expressing retinal ganglion cells in the mouse. J. Comp. Neurol. 2006, 497, 326–349. [Google Scholar] [CrossRef]
- Rumanova, V.S.; Okuliarova, M.; Zeman, M. Differential Effects of Constant Light and Dim Light at Night on the Circadian Control of Metabolism and Behavior. Int. J. Mol. Sci. 2020, 21, 5478. [Google Scholar] [CrossRef] [PubMed]
- Walker, W.H., 2nd; Walton, J.C.; DeVries, A.C.; Nelson, R.J. Circadian rhythm disruption and mental health. Transl. Psychiatry 2020, 10, 28. [Google Scholar] [CrossRef]
- Chinoy, E.D.; Duffy, J.F.; Czeisler, C.A. Unrestricted evening use of light-emitting tablet computers delays self-selected bedtime and disrupts circadian timing and alertness. Physiol. Rep. 2018, 6, e13692. [Google Scholar] [CrossRef]
- González, M.M.C. Dim Light at Night and Constant Darkness: Two Frequently Used Lighting Conditions That Jeopardize the Health and Well-being of Laboratory Rodents. Front. Neurol. 2018, 9, 609. [Google Scholar] [CrossRef]
- Claustrat, B.; Valatx, J.L.; Harthe, C.; Brun, J. Effect of constant light on prolactin and corticosterone rhythms evaluated using a noninvasive urine sampling protocol in the rat. Horm. Metab. Res. 2008, 40, 398–403. [Google Scholar] [CrossRef]
- Mrosovsky, N. Aschoff’s rule in retinally degenerate mice. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2003, 189, 75–78. [Google Scholar] [CrossRef]
- Raven, F.; Van der Zee, E.A.; Meerlo, P.; Havekes, R. The role of sleep in regulating structural plasticity and synaptic strength: Implications for memory and cognitive function. Sleep Med. Rev. 2018, 39, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Yuste, R.; Denk, W. Dendritic spines as basic functional units of neuronal integration. Nature 1995, 375, 682–684. [Google Scholar] [CrossRef]
- Vlachos, A.; Ikenberg, B.; Lenz, M.; Becker, D.; Reifenberg, K.; Bas-Orth, C.; Deller, T. Synaptopodin regulates denervation-induced homeostatic synaptic plasticity. Proc. Natl. Acad. Sci. USA 2013, 110, 8242–8247. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, A.; Becker, D.; Jedlicka, P.; Winkels, R.; Roeper, J.; Deller, T. Entorhinal denervation induces homeostatic synaptic scaling of excitatory postsynapses of dentate granule cells in mouse organotypic slice cultures. PLoS ONE 2012, 7, e32883. [Google Scholar] [CrossRef] [PubMed]
- Turrigiano, G. Homeostatic synaptic plasticity: Local and global mechanisms for stabilizing neuronal function. Cold Spring Harb. Perspect. Biol. 2012, 4, a005736. [Google Scholar] [CrossRef]
- O’Brien, R.J.; Kamboj, S.; Ehlers, M.D.; Rosen, K.R.; Fischbach, G.D.; Huganir, R.L. Activity-dependent modulation of synaptic AMPA receptor accumulation. Neuron 1998, 21, 1067–1078. [Google Scholar] [CrossRef]
- Sutton, M.A.; Ito, H.T.; Cressy, P.; Kempf, C.; Woo, J.C.; Schuman, E.M. Miniature neurotransmission stabilizes synaptic function via tonic suppression of local dendritic protein synthesis. Cell 2006, 125, 785–799. [Google Scholar] [CrossRef]
- Pozo, K.; Goda, Y. Unraveling mechanisms of homeostatic synaptic plasticity. Neuron 2010, 66, 337–351. [Google Scholar] [CrossRef]
- Kopec, C.D.; Real, E.; Kessels, H.W.; Malinow, R. GluR1 links structural and functional plasticity at excitatory synapses. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 13706–13718. [Google Scholar] [CrossRef]
- Mundel, P. Synaptopodin: An Actin-associated Protein in Telencephalic Dendrites and Renal Podocytes. J. Cell Biol. 1997, 139, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Deller, T.; Korte, M.; Chabanis, S.; Drakew, A.; Schwegler, H.; Stefani, G.G.; Zuniga, A.; Schwarz, K.; Bonhoeffer, T.; Zeller, R.; et al. Synaptopodin-deficient mice lack a spine apparatus and show deficits in synaptic plasticity. Proc. Natl. Acad. Sci. USA 2003, 100, 10494–10499. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, A.; Korkotian, E.; Schonfeld, E.; Copanaki, E.; Deller, T.; Segal, M. Synaptopodin regulates plasticity of dendritic spines in hippocampal neurons. J. Neurosci. 2009, 29, 1017–1033. [Google Scholar] [CrossRef]
- Bas Orth, C.; Vlachos, A.; Del Turco, D.; Burbach, G.J.; Haas, C.A.; Mundel, P.; Feng, G.; Frotscher, M.; Deller, T. Lamina-specific distribution of Synaptopodin, an actin-associated molecule essential for the spine apparatus, in identified principal cell dendrites of the mouse hippocampus. J. Comp. Neurol. 2005, 487, 227–239. [Google Scholar] [CrossRef]
- Deller, T. Actin-associated protein Synaptopodin. J. Comp. Neurol. 2000, 418, 164–181. [Google Scholar] [CrossRef]
- Deller, T. Potential Role of Synaptopodin in Spine Motility by Coupling Actin to the Spine Apparatus. Hippocampus 2000, 10, 569–581. [Google Scholar] [CrossRef]
- Yap, K.; Drakew, A.; Smilovic, D.; Rietsche, M.; Paul, M.H.; Vuksic, M.; Del Turco, D.; Deller, T. The actin-modulating protein synaptopodin mediates long-term survival of dendritic spines. eLife 2020, 9, e62944. [Google Scholar] [CrossRef]
- Matsuzaki, M.; Ellis-Davies, G.C.; Nemoto, T.; Miyashita, Y.; Iino, M.; Kasai, H. Dendritic spine geometry is critical for AMPA receptor expression in hippocampal CA1 pyramidal neurons. Nat. Neurosci. 2001, 4, 1086–1092. [Google Scholar] [CrossRef]
- Matsuzaki, M.; Honkura, N.; Ellis-Davies, G.C.; Kasai, H. Structural basis of long-term potentiation in single dendritic spines. Nature 2004, 429, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Hojo, Y.; Komatsuzaki, Y.; Okamoto, M.; Kato, A.; Takeda, T.; Kawato, S. Hippocampal spine changes across the sleep-wake cycle: Corticosterone and kinases. J. Endocrinol. 2015, 226, M13–M27. [Google Scholar] [CrossRef] [PubMed]
- Jasinska, M.; Grzegorczyk, A.; Woznicka, O.; Jasek, E.; Kossut, M.; Barbacka-Surowiak, G.; Litwin, J.A.; Pyza, E. Circadian rhythmicity of synapses in mouse somatosensory cortex. Eur. J. Neurosci. 2015, 42, 2585–2594. [Google Scholar] [CrossRef] [PubMed]
- Debski, K.J.; Ceglia, N.; Ghestem, A.; Ivanov, A.I.; Brancati, G.E.; Bröer, S.; Bot, A.M.; Müller, J.A.; Schoch, S.; Becker, A.; et al. The circadian hippocampus and its reprogramming in epilepsy: Impact for chronotherapeutics. bioRxiv 2020. [Google Scholar] [CrossRef]
- Kilkenny, C.; Browne, W.J.; Cuthi, I.; Emerson, M.; Altman, D.G. Improving bioscience research reporting: The ARRIVE guidelines for reporting animal research. Vet. Clin. Pathol. 2012, 41, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, R. The neurobiological basis of spontaneous alternation. Neurosci. Biobehav. Rev. 2002, 26, 91–104. [Google Scholar] [CrossRef]
- Naert, A.; Gantois, I.; Laeremans, A.; Vreysen, S.; Van den Bergh, G.; Arckens, L.; Callaerts-Vegh, Z.; D’Hooge, R. Behavioural alterations relevant to developmental brain disorders in mice with neonatally induced ventral hippocampal lesions. Brain Res. Bull. 2013, 94, 71–81. [Google Scholar] [CrossRef]
- Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates/Keith B.J. Franklin, George Paxinos; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Strehl, A.; Lenz, M.; Itsekson-Hayosh, Z.; Becker, D.; Chapman, J.; Deller, T.; Maggio, N.; Vlachos, A. Systemic inflammation is associated with a reduction in Synaptopodin expression in the mouse hippocampus. Exp. Neurol. 2014, 261, 230–235. [Google Scholar] [CrossRef]
- Lenz, M.; Ben Shimon, M.; Deller, T.; Vlachos, A.; Maggio, N. Pilocarpine-Induced Status Epilepticus Is Associated with Changes in the Actin-Modulating Protein Synaptopodin and Alterations in Long-Term Potentiation in the Mouse Hippocampus. Neural Plast. 2017, 2017, 2652560. [Google Scholar] [CrossRef]
- Richetto, J.; Polesel, M.; Weber-Stadlbauer, U. Effects of light and dark phase testing on the investigation of behavioural paradigms in mice: Relevance for behavioural neuroscience. Pharmacol. Biochem. Behav. 2019, 178, 19–29. [Google Scholar] [CrossRef]
- Tsao, C.H.; Flint, J.; Huang, G.J. Influence of diurnal phase on behavioral tests of sensorimotor performance, anxiety, learning and memory in mice. Sci. Rep. 2022, 12, 432. [Google Scholar] [CrossRef]
- Roedel, A.; Storch, C.; Holsboer, F.; Ohl, F. Effects of light or dark phase testing on behavioural and cognitive performance in DBA mice. Lab. Anim. 2006, 40, 371–381. [Google Scholar] [CrossRef]
- Jasinska, M.; Jasek-Gajda, E.; Woznicka, O.; Lis, G.J.; Pyza, E.; Litwin, J.A. Circadian clock regulates the shape and content of dendritic spines in mouse barrel cortex. PLoS ONE 2019, 14, e0225394. [Google Scholar] [CrossRef] [PubMed]
- de Vivo, L.; Bellesi, M.; Marshall, W.; Bushong, E.A.; Ellisman, M.H.; Tononi, G.; Cirelli, C. Ultrastructural evidence for synaptic scaling across the wake/sleep cycle. Science 2017, 355, 507–510. [Google Scholar] [CrossRef] [PubMed]
- Chaudhury, D.; Wang, L.M.; Colwell, C.S. Circadian regulation of hippocampal long-term potentiation. J. Biol. Rhythm. 2005, 20, 225–236. [Google Scholar] [CrossRef]
- Wang, L.M.; Suthana, N.A.; Chaudhury, D.; Weaver, D.R.; Colwell, C.S. Melatonin inhibits hippocampal long-term potentiation. Eur. J. Neurosci. 2005, 22, 2231–2237. [Google Scholar] [CrossRef] [PubMed]
- Fukazawa, Y. Hippocampal LTP Is Accompanied by Enhanced F-Actin Content within the Dendritic Spine that Is Essential for Late LTP Maintenance In Vivo. Neuron 2003, 38, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, M.; Matsuo, R.; Fukazawa, Y.; Ozawa, F.; Inokuchi, K. Regulated expression of an actin-associated protein, synaptopodin, during long-term potentiation. J. Neurochem. 2001, 79, 192–199. [Google Scholar] [CrossRef]
- Vlachos, A.; Maggio, N.; Segal, M. Lack of correlation between synaptopodin expression and the ability to induce LTP in the rat dorsal and ventral hippocampus. Hippocampus 2008, 18, 1–4. [Google Scholar] [CrossRef]
- Filipski, E.; King, V.M.; Li, X.; Granda, T.G.; Mormont, M.C.; Liu, X.; Claustrat, B.; Hastings, M.H.; Lévi, F. Host circadian clock as a control point in tumor progression. J. Natl. Cancer Inst. 2002, 94, 690–697. [Google Scholar] [CrossRef]
- Tsinkalovsky, O.; Filipski, E.; Rosenlund, B.; Sothern, R.; Eiken, H.; Wu, M.; Claustrat, B.; Bayer, J.; Lévi, F.; Laerum, O. Circadian expression of clock genes in purified hematopoietic stem cells is developmentally regulated in mouse bone marrow. Exp. Hematol. 2006, 34, 1249–1261. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Hieber, C.; Häusser, M. Cellular mechanisms of spatial navigation in the medial entorhinal cortex. Nat. Neurosci. 2013, 16, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Kalén, P.; Rosegren, E.; Lindvall, O.; Björklund, A. Hippocampal Noradrenaline and Serotonin Release over 24 Hours as Measured by the Dialysis Technique in Freely Moving Rats: Correlation to Behavioural Activity State, Effect of Handling and Tail-Pinch. Eur. J. Neurosci. 1989, 1, 181–188. [Google Scholar] [CrossRef]
- Kasai, H.; Fukuda, M.; Watanabe, S.; Hayashi-Takagi, A.; Noguchi, J. Structural dynamics of dendritic spines in memory and cognition. Trends Neurosci. 2010, 33, 121–129. [Google Scholar] [CrossRef]
- Jasinska, M.; Woznicka, O.; Jasek-Gajda, E.; Lis, G.J.; Pyza, E.; Litwin, J.A. Circadian Changes of Dendritic Spine Geometry in Mouse Barrel Cortex. Front. Neurosci. 2020, 14, 578881. [Google Scholar] [CrossRef]
- Araya, R.; Vogels, T.P.; Yuste, R. Activity-dependent dendritic spine neck changes are correlated with synaptic strength. Proc. Natl. Acad. Sci. USA 2014, 111, E2895–E2904. [Google Scholar] [CrossRef] [PubMed]
- Chai, A.P.; Ma, W.P.; Wang, L.P.; Cao, J.; Xu, L.; Yang, Y.X.; Mao, R.R. Chronic constant light-induced hippocampal late-phase long-term potentiation impairment in vitro is attenuated by antagonist of D1/D5 receptors. Brain Res. 2015, 1622, 72–80. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, H.K.; Liu, F.; Lei, G.; Liu, P.; Jiao, T.; Dang, Y.H. Altered Light Conditions Contribute to Abnormalities in Emotion and Cognition through HINT1 Dysfunction in C57BL/6 Mice. Front. Behav. Neurosci. 2018, 12, 110. [Google Scholar] [CrossRef]
- LeGates, T.A.; Altimus, C.M.; Wang, H.; Lee, H.K.; Yang, S.; Zhao, H.; Kirkwood, A.; Weber, E.T.; Hattar, S. Aberrant light directly impairs mood and learning through melanopsin-expressing neurons. Nature 2012, 491, 594–598. [Google Scholar] [CrossRef]
- Krugers, H.J.; Douma, B.R.; Andringa, G.; Bohus, B.; Korf, J.; Luiten, P.G. Exposure to chronic psychosocial stress and corticosterone in the rat: Effects on spatial discrimination learning and hippocampal protein kinase Cgamma immunoreactivity. Hippocampus 1997, 7, 427–436. [Google Scholar] [CrossRef]
- Babayan, A.H.; Kramár, E.A. Rapid effects of oestrogen on synaptic plasticity: Interactions with actin and its signalling proteins. J. Neuroendocr. 2013, 25, 1163–1172. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schröder, J.K.; Abdel-Hafiz, L.; Ali, A.A.H.; Cousin, T.C.; Hallenberger, J.; Rodrigues Almeida, F.; Anstötz, M.; Lenz, M.; Vlachos, A.; von Gall, C.; et al. Effects of the Light/Dark Phase and Constant Light on Spatial Working Memory and Spine Plasticity in the Mouse Hippocampus. Cells 2023, 12, 1758. https://doi.org/10.3390/cells12131758
Schröder JK, Abdel-Hafiz L, Ali AAH, Cousin TC, Hallenberger J, Rodrigues Almeida F, Anstötz M, Lenz M, Vlachos A, von Gall C, et al. Effects of the Light/Dark Phase and Constant Light on Spatial Working Memory and Spine Plasticity in the Mouse Hippocampus. Cells. 2023; 12(13):1758. https://doi.org/10.3390/cells12131758
Chicago/Turabian StyleSchröder, Jane K., Laila Abdel-Hafiz, Amira A. H. Ali, Teresa C. Cousin, Johanna Hallenberger, Filipe Rodrigues Almeida, Max Anstötz, Maximilian Lenz, Andreas Vlachos, Charlotte von Gall, and et al. 2023. "Effects of the Light/Dark Phase and Constant Light on Spatial Working Memory and Spine Plasticity in the Mouse Hippocampus" Cells 12, no. 13: 1758. https://doi.org/10.3390/cells12131758
APA StyleSchröder, J. K., Abdel-Hafiz, L., Ali, A. A. H., Cousin, T. C., Hallenberger, J., Rodrigues Almeida, F., Anstötz, M., Lenz, M., Vlachos, A., von Gall, C., & Tundo-Lavalle, F. (2023). Effects of the Light/Dark Phase and Constant Light on Spatial Working Memory and Spine Plasticity in the Mouse Hippocampus. Cells, 12(13), 1758. https://doi.org/10.3390/cells12131758