
α7 Nicotinic Acetylcholine Receptors May Improve Schwann Cell Regenerating Potential via Metabotropic Signaling Pathways

 ,
,  ,
,  ,
,  , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Statements for Animal Use
2.2. Cell Cultures
2.3. Pharmacological Treatment
2.4. [Ca2+]i Measurements in Cultured SCs
2.5. Total RNA Extraction and RT-PCR Analysis
2.6. Protein Extraction and Western Blot Analysis
2.7. Wound Healing Assay
2.8. Phalloidin Staining
2.9. Immunocytochemistry
2.10. Nucleus–Cytoplasm Extraction
2.11. Statistical Analysis
3. Results
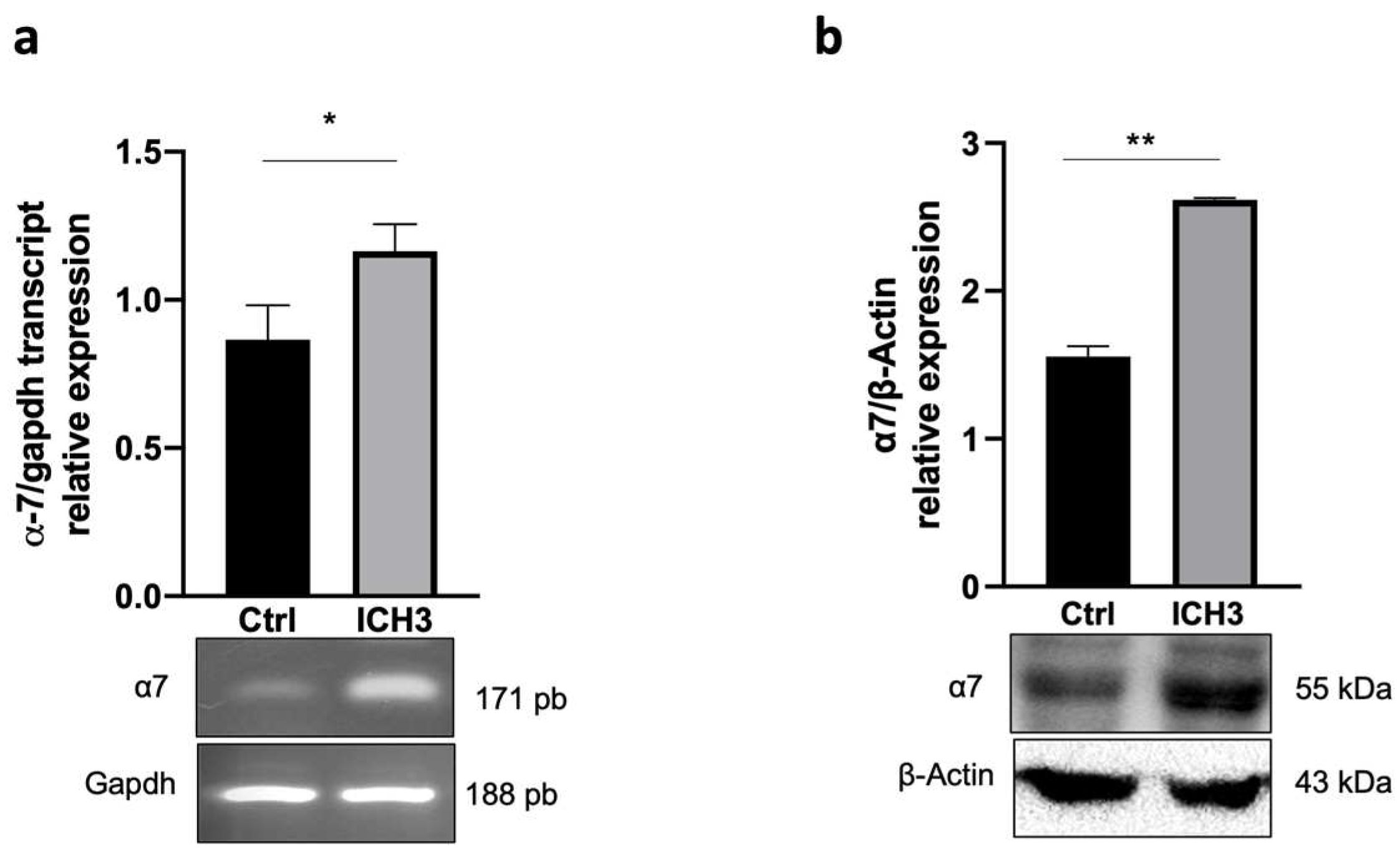
3.1. Analysis of α7 nAChR Expression
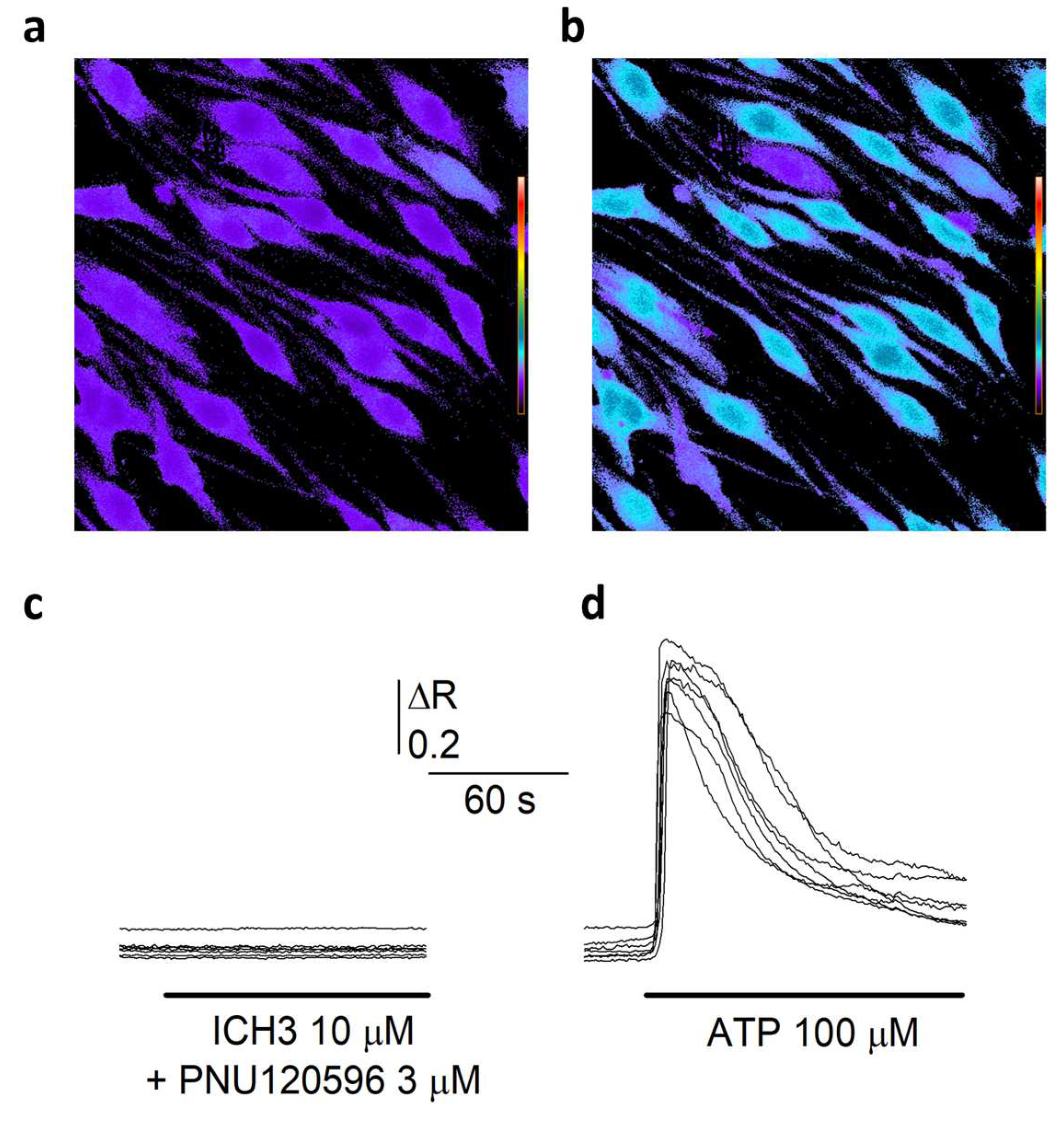
3.2. [Ca2+]i Measurements in Schwann Cells
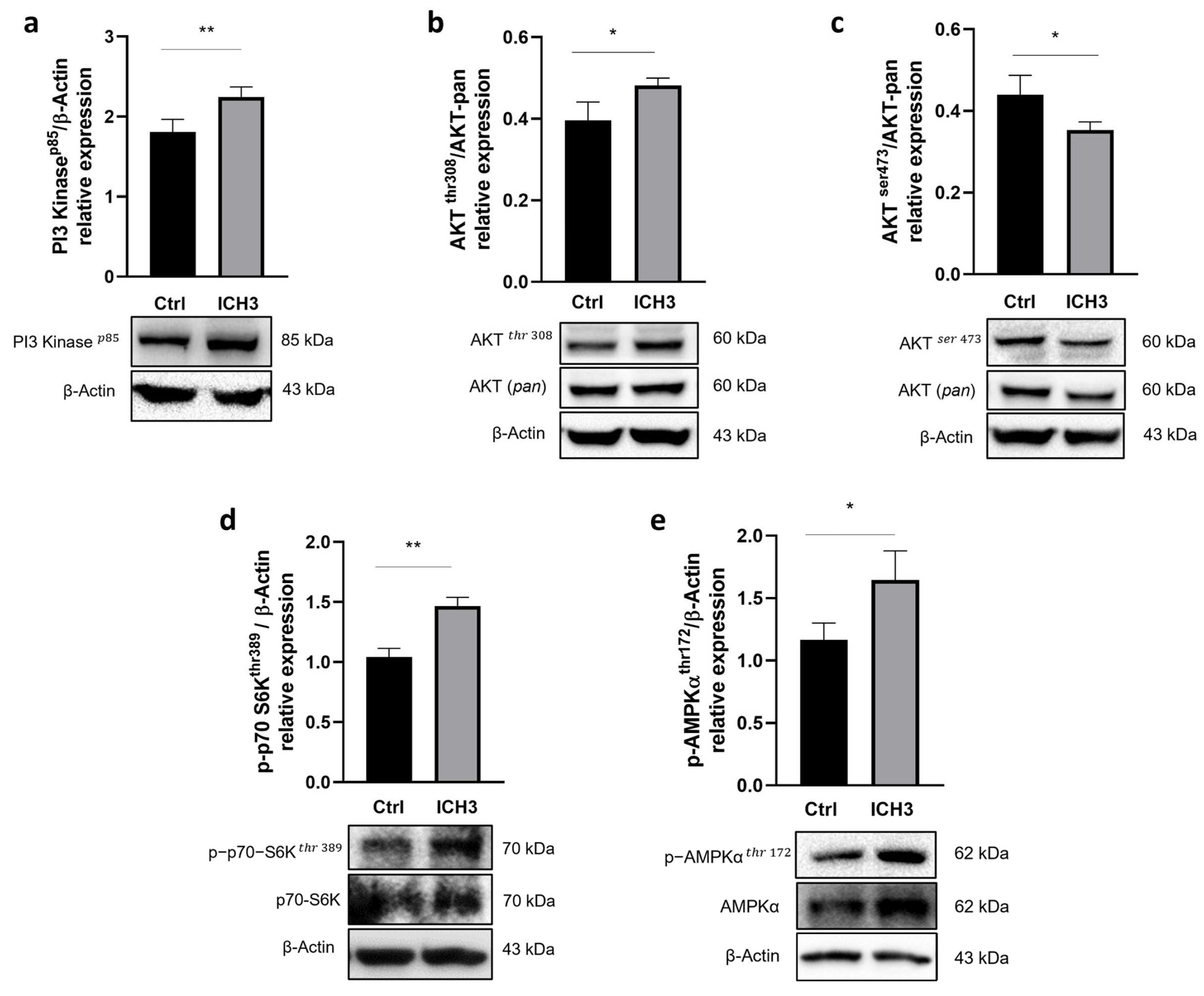
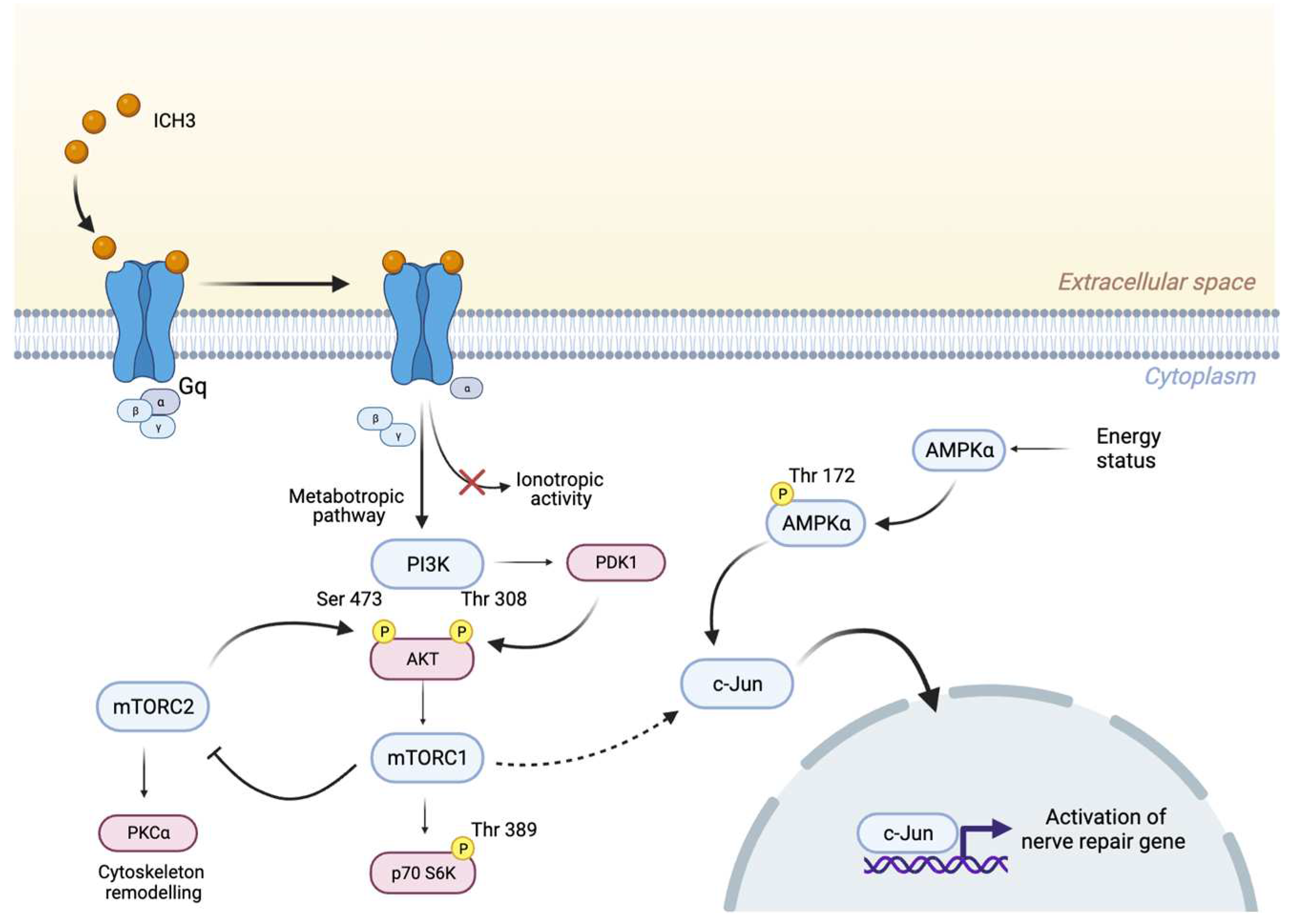
3.3. Analysis of the PI3K/AKT/mTORC1 Pathway
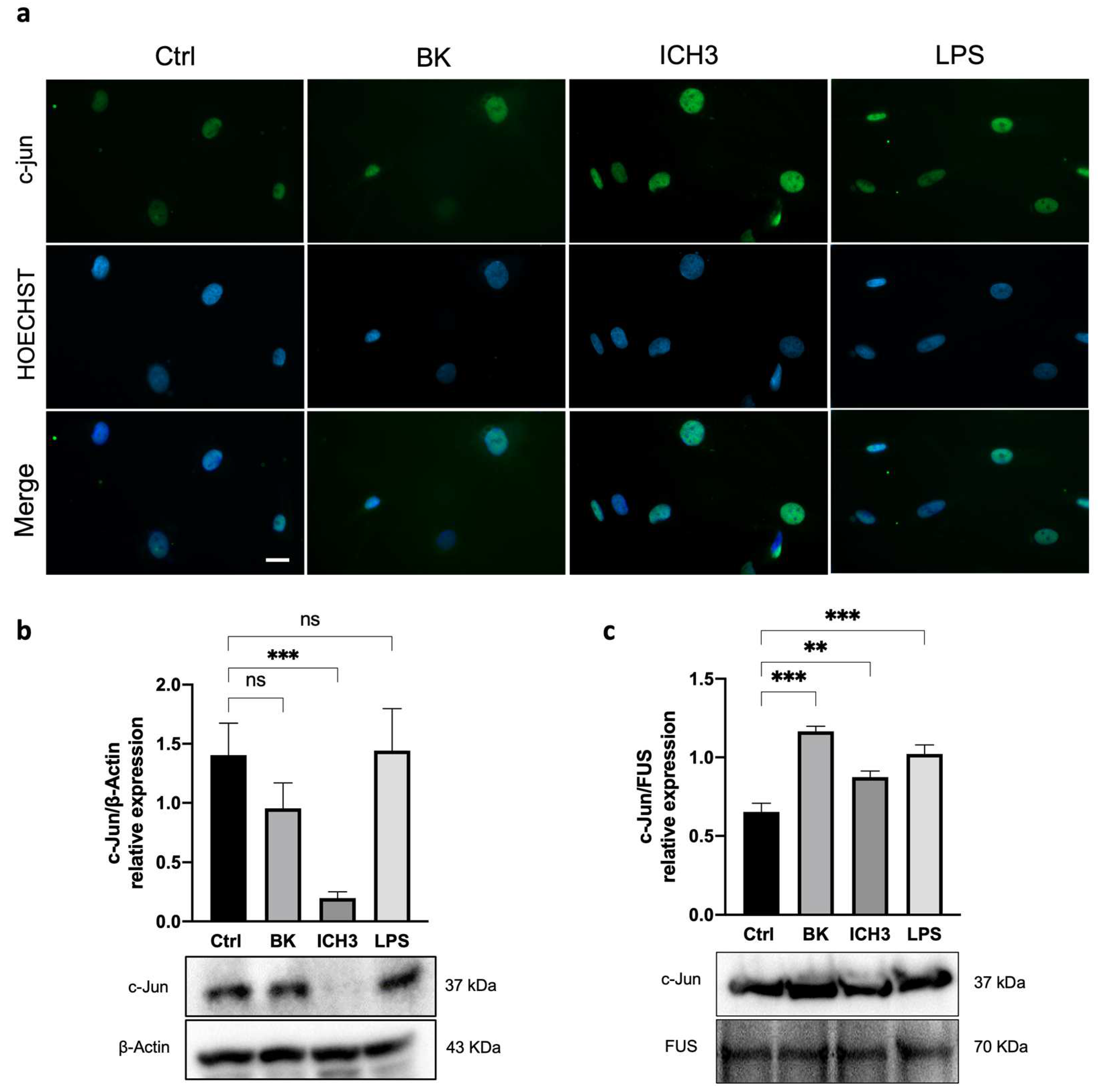
3.4. C-Jun Expression in SCs after α7 nAChR Activation
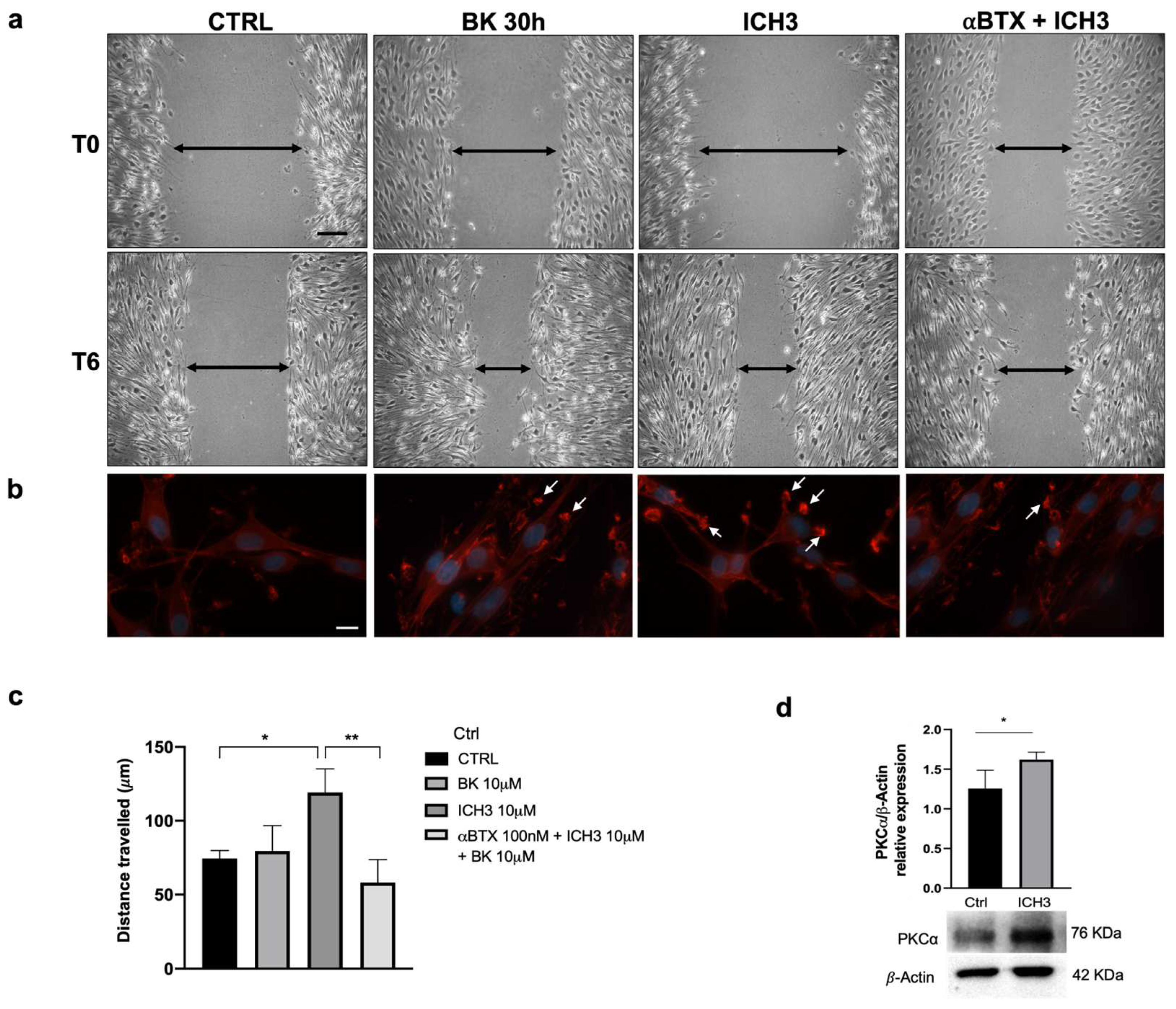
3.5. Analysis of SC Migration after α7 nAChR Activation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jessen, K.R.; Mirsky, R.; Lloyd, A.C. Schwann Cells: Development and Role in Nerve Repair. Cold Spring Harb. Perspect. Biol. 2015, 7, a020487. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Sinanan, A.; Parkinson, D.; Parmantier, E.; Mirsky, R.; Jessen, K.R. Schwann Cell Development in Embryonic Mouse Nerves. J. Neurosci. Res. 1999, 56, 334–348. [Google Scholar] [CrossRef]
- Gomez-Sanchez, J.A.; Pilch, K.S.; van der Lans, M.; Fazal, S.V.; Benito, C.; Wagstaff, L.J.; Mirsky, R.; Jessen, K.R. After Nerve Injury, Lineage Tracing Shows That Myelin and Remak Schwann Cells Elongate Extensively and Branch to Form Repair Schwann Cells, Which Shorten Radically on Remyelination. J. Neurosci. 2017, 37, 9086–9099. [Google Scholar] [CrossRef] [PubMed]
- Jessen, K.R.; Mirsky, R. The Success and Failure of the Schwann Cell Response to Nerve Injury. Front. Cell. Neurosci. 2019, 13, 33. [Google Scholar] [CrossRef]
- Loreti, S.; Ricordy, R.; Egle De Stefano, M.; Augusti-Tocco, G.; Maria Tata, A. Acetylcholine Inhibits Cell Cycle Progression in Rat Schwann Cells by Activation of the M2 Receptor Subtype. Neuron Glia Biol. 2007, 3, 269–279. [Google Scholar] [CrossRef]
- Loreti, S.; Vilaró, M.T.; Visentin, S.; Rees, H.; Levey, A.I.; Tata, A.M. Rat Schwann Cells Express M1–M4 Muscarinic Receptor Subtypes. J. Neurosci. Res. 2006, 84, 97–105. [Google Scholar] [CrossRef]
- Uggenti, C.; De Stefano, M.E.; Costantino, M.; Loreti, S.; Pisano, A.; Avallone, B.; Talora, C.; Magnaghi, V.; Tata, A.M. M2 Muscarinic Receptor Activation Regulates Schwann Cell Differentiation and Myelin Organization: ACh Modulation of Schwann Cell Development. Dev. Neurobiol. 2014, 74, 676–691. [Google Scholar] [CrossRef]
- Piovesana, R.; Faroni, A.; Tata, A.M.; Reid, A.J. Functional Characterization of Muscarinic Receptors in Human Schwann Cells. Int. J. Mol. Sci. 2020, 21, 6666. [Google Scholar] [CrossRef]
- Botticelli, E.; Salazar Intriago, M.S.; Piovesana, R.; Tata, A.M. Analysis of Signal Transduction Pathways Downstream M2 Receptor Activation: Effects on Schwann Cell Migration and Morphology. Life 2022, 12, 211. [Google Scholar] [CrossRef]
- Dallanoce, C.; Magrone, P.; Matera, C.; Frigerio, F.; Grazioso, G.; De Amici, M.; Fucile, S.; Piccari, V.; Frydenvang, K.; Pucci, L.; et al. Design, Synthesis, and Pharmacological Characterization of Novel Spirocyclic Quinuclidinyl-Δ2-Isoxazoline Derivatives as Potent and Selective Agonists of A7 Nicotinic Acetylcholine Receptors. ChemMedChem 2011, 6, 889–903. [Google Scholar] [CrossRef]
- Salazar Intriago, M.S.; Piovesana, R.; Matera, A.; Taggi, M.; Canipari, R.; Fabrizi, C.; Papotto, C.; Matera, C.; De Amici, M.; Dallanoce, C.; et al. The Mechanisms Mediated by A7 Acetylcholine Nicotinic Receptors May Contribute to Peripheral Nerve Regeneration. Molecules 2021, 26, 7668. [Google Scholar] [CrossRef] [PubMed]
- Tracey, K.J. Physiology and Immunology of the Cholinergic Antiinflammatory Pathway. J. Clin. Investig. 2007, 117, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, M.S.; Mikkelsen, J.D. The A7 Nicotinic Acetylcholine Receptor Ligands Methyllycaconitine, NS6740 and GTS-21 Reduce Lipopolysaccharide-Induced TNF-α Release from Microglia. J. Neuroimmunol. 2012, 251, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Kabbani, N.; Nichols, R.A. Beyond the Channel: Metabotropic Signaling by Nicotinic Receptors. Trends Pharmacol. Sci. 2018, 39, 354–366. [Google Scholar] [CrossRef]
- Figlia, G.; Norrmén, C.; Pereira, J.A.; Gerber, D.; Suter, U. Dual Function of the PI3K-Akt-MTORC1 Axis in Myelination of the Peripheral Nervous System. eLife 2017, 6, e29241. [Google Scholar] [CrossRef]
- Norrmén, C.; Figlia, G.; Pfistner, P.; Pereira, J.A.; Bachofner, S.; Suter, U. MTORC1 Is Transiently Reactivated in Injured Nerves to Promote C-Jun Elevation and Schwann Cell Dedifferentiation. J. Neurosci. 2018, 38, 4811–4828. [Google Scholar] [CrossRef]
- Liu, X.; Peng, S.; Zhao, Y.; Zhao, T.; Wang, M.; Luo, L.; Yang, Y.; Sun, C. AMPK Negatively Regulates Peripheral Myelination via Activation of C-Jun. Mol. Neurobiol. 2017, 54, 3554–3564. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Z.; Wang, J.; Du, S.; Yao, D.; Cao, R.; Cui, S. Protein Kinase Cα Promotes Proliferation and Migration of Schwann Cells by Activating ERK Signaling Pathway. Neuroscience 2020, 433, 94–107. [Google Scholar] [CrossRef]
- Matera, C.; Dondio, G.; Braida, D.; Ponzoni, L.; De Amici, M.; Sala, M.; Dallanoce, C. In Vivo and in Vitro ADMET Profiling and in Vivo Pharmacodynamic Investigations of a Selective A7 Nicotinic Acetylcholine Receptor Agonist with a Spirocyclic Δ 2 -Isoxazoline Molecular Skeleton. Eur. J. Pharmacol. 2018, 820, 265–273. [Google Scholar] [CrossRef]
- Scabia, G.; Cancello, R.; Dallanoce, C.; Berger, S.; Matera, C.; Dattilo, A.; Zulian, A.; Barone, I.; Ceccarini, G.; Santini, F.; et al. ICH3, a Selective Alpha7 Nicotinic Acetylcholine Receptor Agonist, Modulates Adipocyte Inflammation Associated with Obesity. J. Endocrinol. Investig. 2020, 43, 983–993. [Google Scholar] [CrossRef]
- Fucile, S. The Distribution of Charged Amino Acid Residues and the Ca2+ Permeability of Nicotinic Acetylcholine Receptors: A Predictive Model. Front. Mol. Neurosci. 2017, 10, 155. [Google Scholar] [CrossRef] [PubMed]
- Hurst, R.S. A Novel Positive Allosteric Modulator of the 7 Neuronal Nicotinic Acetylcholine Receptor: In Vitro and In Vivo Characterization. J. Neurosci. 2005, 25, 4396–4405. [Google Scholar] [CrossRef] [PubMed]
- Valbuena, S.; Lerma, J. Non-Canonical Signaling, the Hidden Life of Ligand-Gated Ion Channels. Neuron 2016, 92, 316–329. [Google Scholar] [CrossRef] [PubMed]
- Horenstein, N.A.; Papke, R.L. Anti-Inflammatory Silent Agonists. ACS Med. Chem. Lett. 2017, 8, 989–991. [Google Scholar] [CrossRef] [PubMed]
- Ishii, A.; Furusho, M.; Bansal, R. Mek/ERK1/2-MAPK and PI3K/Akt/mTOR Signaling Plays Both Independent and Cooperative Roles in Schwann Cell Differentiation, Myelination and Dysmyelination. Glia 2021, 69, 2429–2446. [Google Scholar] [CrossRef]
- Domenech-Estevez, E.; Baloui, H.; Meng, X.; Zhang, Y.; Deinhardt, K.; Dupree, J.L.; Einheber, S.; Chrast, R.; Salzer, J.L. Akt Regulates Axon Wrapping and Myelin Sheath Thickness in the PNS. J. Neurosci. 2016, 36, 4506–4521. [Google Scholar] [CrossRef]
- Norrmén, C.; Suter, U. Akt/MTOR Signalling in Myelination. Biochem. Soc. Trans. 2013, 41, 944–950. [Google Scholar] [CrossRef] [PubMed]
- Dibble, C.C.; Asara, J.M.; Manning, B.D. Characterization of Rictor Phosphorylation Sites Reveals Direct Regulation of MTOR Complex 2 by S6K1. Mol. Cell. Biol. 2009, 29, 5657–5670. [Google Scholar] [CrossRef]
- Boerboom, A.; Dion, V.; Chariot, A.; Franzen, R. Molecular Mechanisms Involved in Schwann Cell Plasticity. Front. Mol. Neurosci. 2017, 10, 38. [Google Scholar] [CrossRef]
- Fontana, X.; Hristova, M.; Da Costa, C.; Patodia, S.; Thei, L.; Makwana, M.; Spencer-Dene, B.; Latouche, M.; Mirsky, R.; Jessen, K.R.; et al. C-Jun in Schwann Cells Promotes Axonal Regeneration and Motoneuron Survival via Paracrine Signaling. J. Cell. Biol. 2012, 198, 127–141. [Google Scholar] [CrossRef]
- Parkinson, D.B.; Bhaskaran, A.; Arthur-Farraj, P.; Noon, L.A.; Woodhoo, A.; Lloyd, A.C.; Feltri, M.L.; Wrabetz, L.; Behrens, A.; Mirsky, R.; et al. C-Jun Is a Negative Regulator of Myelination. J. Cell. Biol. 2008, 181, 625–637. [Google Scholar] [CrossRef]
- Shy, M.E.; Shi, Y.; Wrabetz, L.; Kamholz, J.; Scherer, S.S. Axon-Schwann Cell Interactions Regulate the Expression of c-Jun in Schwann Cells. J. Neurosci. Res. 1996, 43, 511–525. [Google Scholar] [CrossRef]
- Shen, A.; Yang, J.; Gu, Y.; Zhou, D.; Sun, L.; Qin, Y.; Chen, J.; Wang, P.; Xiao, F.; Zhang, L.; et al. Lipopolysaccharide-Evoked Activation of P38 and JNK Leads to an Increase in ICAM-1 Expression in Schwann Cells of Sciatic Nerves: LPS Increases ICAM-1 Expression in Schwann Cells of Sciatic Nerves. FEBS J. 2008, 275, 4343–4353. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.T.; Hawrot, E.; Lentz, T.L. Distribution of Alpha-Bungarotoxin Binding Sites over Residues 173-204 of the Alpha Subunit of the Acetylcholine Receptor. Mol. Pharmacol. 1988, 34, 643–650. [Google Scholar] [PubMed]
- Trepat, X.; Chen, Z.; Jacobson, K. Cell Migration. In Comprehensive Physiology; Terjung, R., Ed.; Wiley: Hoboken, NJ, USA, 2012; pp. 2369–2392. ISBN 978-0-470-65071-4. [Google Scholar]
- Singh, R.K.; Kumar, S.; Gautam, P.K.; Tomar, M.S.; Verma, P.K.; Singh, S.P.; Kumar, S.; Acharya, A. Protein Kinase C-α and the Regulation of Diverse Cell Responses. Biomol. Concepts 2017, 8, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Jessen, K.R.; Mirsky, R. The Repair Schwann Cell and Its Function in Regenerating Nerves: Repair Schwann Cell and Its Function in Regenerating Nerves. J. Physiol. 2016, 594, 3521–3531. [Google Scholar] [CrossRef]
- Namgung, U. The Role of Schwann Cell-Axon Interaction in Peripheral Nerve Regeneration. Cells Tissues Organs 2014, 200, 6–12. [Google Scholar] [CrossRef]
- Piovesana, R.; Salazar Intriago, M.S.; Dini, L.; Tata, A.M. Cholinergic Modulation of Neuroinflammation: Focus on A7 Nicotinic Receptor. Int. J. Mol. Sci. 2021, 22, 4912. [Google Scholar] [CrossRef]
- King, J.R.; Kabbani, N. Alpha 7 Nicotinic Receptor Coupling to Heterotrimeric G Proteins Modulates RhoA Activation, Cytoskeletal Motility, and Structural Growth. J. Neurochem. 2016, 138, 532–545. [Google Scholar] [CrossRef] [PubMed]
- de Jonge, W.J.; van der Zanden, E.P.; The, F.O.; Bijlsma, M.F.; van Westerloo, D.J.; Bennink, R.J.; Berthoud, H.-R.; Uematsu, S.; Akira, S.; van den Wijngaard, R.M.; et al. Stimulation of the Vagus Nerve Attenuates Macrophage Activation by Activating the Jak2-STAT3 Signaling Pathway. Nat. Immunol. 2005, 6, 844–851. [Google Scholar] [CrossRef]
- Toborek, M.; Son, K.W.; Pudelko, A.; King-Pospisil, K.; Wylegala, E.; Malecki, A. ERK 1/2 Signaling Pathway Is Involved in Nicotine-Mediated Neuroprotection in Spinal Cord Neurons. J. Cell. Biochem. 2007, 100, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Vijayaraghavan, S. Nicotinic Cholinergic Signaling in Hippocampal Astrocytes Involves Calcium-Induced Calcium Release from Intracellular Stores. Proc. Natl. Acad. Sci. USA 2001, 98, 4148–4153. [Google Scholar] [CrossRef] [PubMed]
- Min, Q.; Parkinson, D.B.; Dun, X. Migrating Schwann Cells Direct Axon Regeneration within the Peripheral Nerve Bridge. Glia 2021, 69, 235–254. [Google Scholar] [CrossRef] [PubMed]
- Tata, A.M.; (Sapienza University of Rome, Rome, Italy). Unpublished work. 2023.
- Grada, A.; Otero-Vinas, M.; Prieto-Castrillo, F.; Obagi, Z.; Falanga, V. Research Techniques Made Simple: Analysis of Collective Cell Migration Using the Wound Healing Assay. J. Investig. Dermatol. 2017, 137, e11–e16. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, S.; Vacca, V.; Ricordy, R.; Uggenti, C.; Tata, A.M.; Luvisetto, S.; Pavone, F. The Analgesic Effect on Neuropathic Pain of Retrogradely Transported Botulinum Neurotoxin A Involves Schwann Cells and Astrocytes. PLoS ONE 2012, 7, e47977. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Botticelli, E.; Guerriero, C.; Fucile, S.; De Stefano, M.E.; Matera, C.; Dallanoce, C.; De Amici, M.; Tata, A.M. α7 Nicotinic Acetylcholine Receptors May Improve Schwann Cell Regenerating Potential via Metabotropic Signaling Pathways. Cells 2023, 12, 1494. https://doi.org/10.3390/cells12111494
Botticelli E, Guerriero C, Fucile S, De Stefano ME, Matera C, Dallanoce C, De Amici M, Tata AM. α7 Nicotinic Acetylcholine Receptors May Improve Schwann Cell Regenerating Potential via Metabotropic Signaling Pathways. Cells. 2023; 12(11):1494. https://doi.org/10.3390/cells12111494
Chicago/Turabian StyleBotticelli, Elisabetta, Claudia Guerriero, Sergio Fucile, Maria Egle De Stefano, Carlo Matera, Clelia Dallanoce, Marco De Amici, and Ada Maria Tata. 2023. "α7 Nicotinic Acetylcholine Receptors May Improve Schwann Cell Regenerating Potential via Metabotropic Signaling Pathways" Cells 12, no. 11: 1494. https://doi.org/10.3390/cells12111494
APA StyleBotticelli, E., Guerriero, C., Fucile, S., De Stefano, M. E., Matera, C., Dallanoce, C., De Amici, M., & Tata, A. M. (2023). α7 Nicotinic Acetylcholine Receptors May Improve Schwann Cell Regenerating Potential via Metabotropic Signaling Pathways. Cells, 12(11), 1494. https://doi.org/10.3390/cells12111494