
Towards an Improved Understanding of the Effects of Elevated Progesterone Levels on Human Endometrial Receptivity and Oocyte/Embryo Quality during Assisted Reproductive Technologies

Abstract
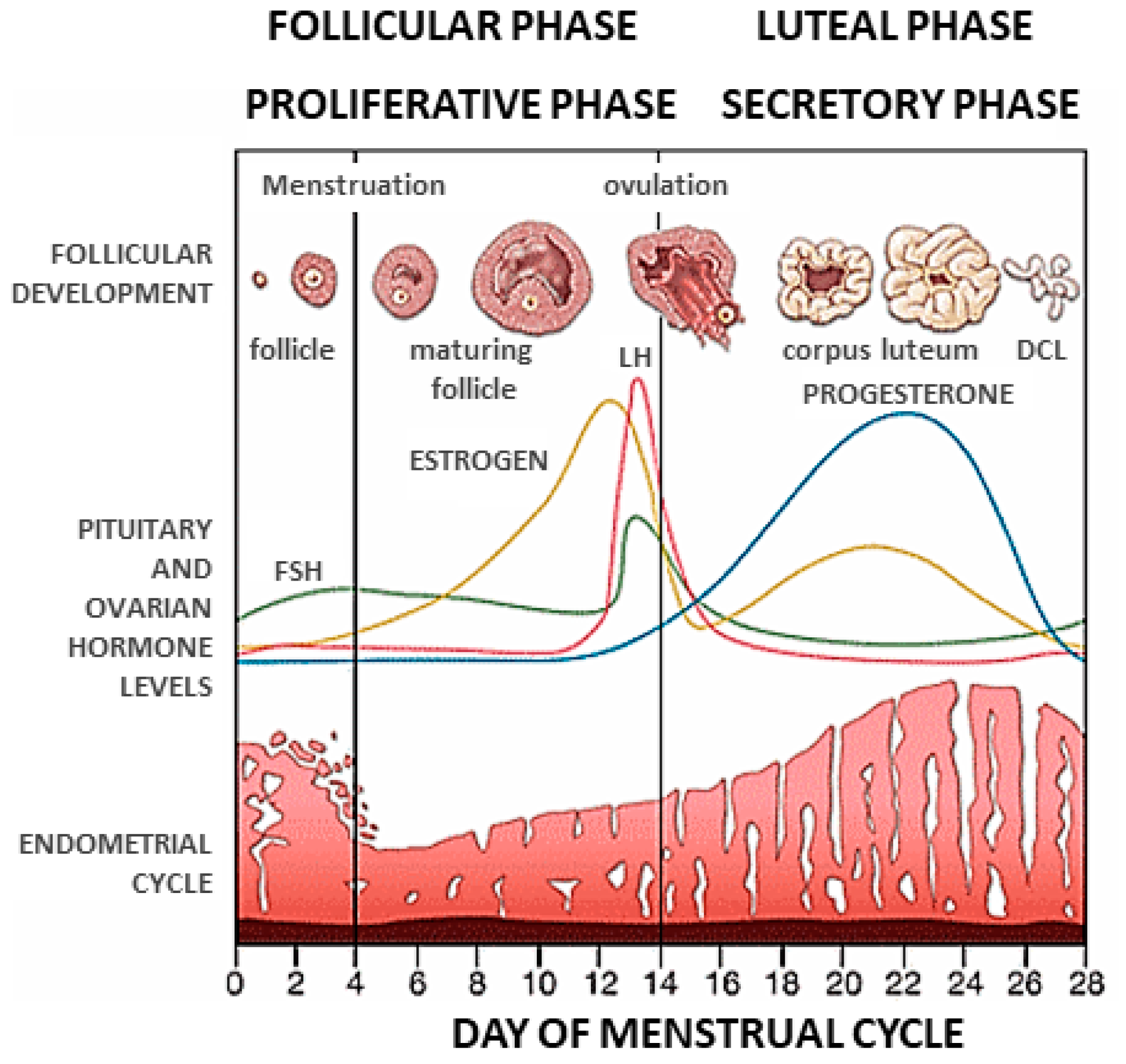
1. Introduction
2. The Impact of Progesterone Levels on Endometrial Histological Development and Endometrial Receptivity
2.1. Introduction
2.2. Assessing the Impact of Progesterone Levels on Endometrial Histological Development in an Ovarian Failure Model Using Endometrial Dating
2.3. Assessing the Impact of Progesterone Levels on Endometrial Histological Development in an IVF Donor Cycle Using Endometrial Dating
2.4. Assessing the Impact of Progesterone Levels on Endometrial Histological Development in an IVF/ET Cycle Using Noninvasive High-Resolution Transvaginal Ultrasonography
2.5. Assessing the Impact of Progesterone Levels on Endometrial Histological Development in an IVF/ET Cycle Using Endometrial Dating
2.6. Assessing the Impact of Progesterone Levels on Endometrial Histological Development in Healthy Women Undergoing Experimentally Modeled Endometrial Cycles Using Endometrial Dating
2.7. Conclusions
3. The Impact of Progesterone Levels on The Endometrial Transcriptome and Endometrial Receptivity
3.1. Introduction
3.2. Assessing the Impact of Randomized Serum Progesterone Concentrations on Endometrial Gene Expression
3.3. Elevated Progesterone Levels on the Day of hCG Trigger Alters Endometrial Gene Expression during Ovarian Stimulation
3.4. Microarray Analysis of Endometrial Gene Expression in OS Cycles with a Premature Elevation in Progesterone Levels Uncovers the Dysregulated Expression of Cell Cycle Genes in the Pre-Receptive Phase
3.5. Examining the Effects of Elevated Progesterone Levels on DNA Methylation and Endometrial Gene and Protein Expression during Ovarian Stimulation
3.6. Limitations
4. The Impact of Elevated Progesterone Levels on Oocyte and Embryo Quality
4.1. Introduction
4.2. Oocyte and Embryo Quality

{kind=link}
| Study/Study Type | ART Cycle | Inclusion Criteria | Exclusion Criteria | Pituitary Desensitization Protocol | Ovarian Stimulation Protocol | Trigger | Fertilization Method | ET/Luteal Support | Assay Coefficients of Variability | Serum Progesterone Level |
|---|---|---|---|---|---|---|---|---|---|---|
| Hofmann et al., 1993 [79]/retrospective | IVF/ET with donor oocyte in those with ovarian failure (n = 68) | Subjects undergoing OS as ovum donors and subjects with ovarian failure | Not described | GNRH agonist (leuprolide acetate) | hMG | hCG | Insemination | Oral estradiol, vaginal or IM P | Intra-assay: 4.7% Inter-assay: 7.9% | 0.9 ng/mL throughout follicular phase (n = 47) Experimental: >1.1 ng/mL on or day before hCG (n = 21) |
| Neves et al., 2021 [87]/retrospective | IVF cycles with frozen ET and PGT-A (n = 1495) | 18–40-y/o infertile subjects with ICSI cycles and PGT-A | Oocyte donation cycles, conventional IVF and ICSI performed in the same cycle, use of testicular spermatozoa, fresh ET cycles, known chromosomal rearrangements or monogenic diseases | GNRH antagonist | FHS, hMG | hCG | ICSI | Not described | Intra-assay: <7% | 1.5 ng/mL on day of trigger (n = 1328) Experimental: >1.5 ng/mL (n = 167) |
| Pardinas et al., 2021 [86]/retrospective | IVF cycles with PGT-A (n = 1597) | Subjects undergoing PGT-A | No exclusion criteria | GNRH antagonist | FSH, LH, hMG | hCG | ICSI | Not described | Intra-assay: 1.2–11.8% Inter-assay: 3.6–23.1% | Control: <1.5 ng/mL on day of hCG (n = 1465) Experimental: ≥1.5 ng/mL (n = 132) |
| Huang et al., 2016 [40]/retrospective | IVF (n = 4236) with fresh ET (n = 2639) | Not described | ICSI cycles, donor cycles | Long GNRH agonist (Decapeptyl and Diphereline) | rFSH (Gonal-F or Puregon) | rhCG (Ovidrel) | Insemination | Not described | Not described | Group 1: 1.00 ng/mL on day of hCG Group 2: 1.00–1.50 ng/mL Group 3: 1.50–2.00 ng/mL Group 4: 2.00–2.50 ng/mL Group 5: >2.50 ng/mL |
| Hernandez-Nieto et al., 2021 [83]/retrospective | IVF with PGT-A (n = 5141) | Only GNRH antagonist protocol | Not described | Flexible GNRH antagonist (cetrorelix acetate or ganirelix acetate) | rFSH, hMG | hCG, leuprolide acetate | ICSI | Oral estradiol, IM P | Not described | 2 ng/mL on day of trigger (n = 4925) Experimental: >2 ng/mL (n = 216) |
| Fanchin et al., 1997 [35]/retrospective | IVF with fresh ET (n = 153) | Not described | Abnormalities of uterine cavity, abnormal sperm analysis | GNRH agonist (leuprolide acetate) | hMG | hCG | Insemination | Not described | Intra-assay: 8% Inter-assay: 11% | 0.9 ng/mL on day of trigger (n = 112) Experimental: >1.1 ng/mL (n = 41) |
| Hill et al., 2015 [85]/retrospective | IVF with fresh ET (n = 1620) | Subjects undergoing fresh autologous ET cycles and measures serum P on day of hCG trigger | ICSI cycles, donor cycles | GNRH-antagonist (ganirelix) or GNRH-agonist (leuprolide acetate) | rFSH, hMG | hCG | Insemination or ICSI | Not described | Intra-assay: 6.7% Inter-assay: 7.2% | 1.5 ng/mL on day of trigger (n = 1466) Group 2: >1.5 ng/mL (n = 114) Group 3: >2.0 ng/mL (n = 40) |
4.3. Clinical Outcome
4.4. Limitations
4.5. Conclusions
5. General Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Assisted reproductive technology | (ART) |
| Corpus luteum | (CL) |
| Cyclic adenosine monophosphate | (cAMP) |
| DNA methyltransferases | (DNMTs) |
| Endometrial biopsy | (EMB) |
| Estrogen | (E) |
| Estradiol | (E2) |
| Follicle-stimulating hormone | (FSH) |
| Recombinant follicle-stimulating hormone | (rFSH) |
| Gonadotropin-releasing hormone | (GNRH) |
| Hormonal replacement therapy | (HRT) |
| Human chorionic gonadotropin | (hCG) |
| Recombinant chorionic gonadotrophin | (rCG) |
| Human menopausal gonadotropin | (hMG) |
| In vitro fertilization/embryo transfer | (IVF/ET) |
| Intracytoplasmic sperm injection | (ICSI) |
| Intramuscular | (IM) |
| Luteinizing hormone | (LH) |
| Ovarian hyperstimulation syndrome | (OHSS) |
| Ovarian stimulation | (OS) |
| Progesterone | (P) |
| Preimplantation genetic testing for aneuploidies | (PGT-A) |
| Significance analysis of microarray data | (SAM) |
| Analysis of variance | (ANOVA) |
| Top quality embryo | (TQE) |
| Window of implantation | (WOI) |
| Immunohistochemistry | (IHC) |
References
- Stocco, C.; Telleria, C.; Gibori, G. The molecular control of corpus luteum formation, function, and regression. Endocr. Rev. 2007, 28, 117–149. [Google Scholar] [CrossRef] [PubMed]
- Taraborrelli, S. Physiology, production and action of progesterone. Acta Obstet. Gynecol. Scand. 2015, 94, 8–16. [Google Scholar] [CrossRef]
- De Geyter, C.; De Geyter, M.; Huber, P.R.; Nieschlag, E.; Holzgreve, W. Progesterone serum levels during the follicular phase of the menstrual cycle originate from the crosstalk between the ovaries and the adrenal cortex. Hum. Reprod. 2002, 17, 933–939. [Google Scholar] [CrossRef]
- Filicori, M.; Butler, J.P.; Crowley, W.F., Jr. Neuroendocrine regulation of the corpus luteum in the human. Evidence for pulsatile progesterone secretion. J. Clin. Investig. 1984, 73, 1638–1647. [Google Scholar] [CrossRef] [PubMed]
- Grunfeld, L.; Walker, B.; Bergh, P.A.; Sandler, B.; Hofmann, G.; Navot, D. High-resolution endovaginal ultrasonography of the endometrium: A noninvasive test for endometrial adequacy. Obstet. Gynecol. 1991, 78, 200–204. [Google Scholar] [PubMed]
- Nadji, P.; Reyniak, J.V.; Sedlis, A.; Szarowski, D.H.; Bartosik, D. Endometrial dating correlated with progesterone levels. Obstet. Gynecol. 1975, 45, 193–194. [Google Scholar] [PubMed]
- Noyes, R.W.; Hertig, A.T.; Rock, J. Dating the endometrial biopsy. Obstet. Gynecol. Surv. 1950, 5, 561–564. [Google Scholar] [CrossRef]
- Santoro, N.; Goldsmith, L.T.; Heller, D.; Illsley, N.; McGovern, P.; Molina, C.; Peters, S.; Skurnick, J.H.; Forst, C.; Weiss, G. Luteal Progesterone Relates to Histological Endometrial Maturation in Fertile Women. J. Clin. Endocrinol. Metab. 2000, 85, 4207–4211. [Google Scholar] [CrossRef][Green Version]
- Rock, J.; Bartlett, M.K. Biopsy studies of human endometrium: Criteria of dating and information about amenorrhea, menorrhagia, and time of ovulation. J. Am. Med. Assoc. 1937, 108, 2022–2028. [Google Scholar] [CrossRef]
- Von Grothusen, C.; Lalitkumar, P.G.; Ruiz-Alonso, M.; Boggavarapu, N.R.; Navarro, R.; Miravet-Valenciano, J.; Gemzell-Danielsson, K.; Simon, C. Effect of mifepristone on the transcriptomic signature of endometrial receptivity. Hum. Reprod. 2018, 33, 1889–1897. [Google Scholar] [CrossRef]
- Domínguez, F.; Remohí, J.; Pellicer, A.; Simón, C. Human endometrial receptivity: A genomic approach. Reprod. Biomed. Online 2003, 6, 332–338. [Google Scholar] [CrossRef]
- Martín, J.; Domínguez, F.; Avila, S.; Castrillo, J.L.; Remohí, J.; Pellicer, A.; Simón, C. Human endometrial receptivity: Gene regulation. J. Reprod. Immunol. 2002, 55, 131–139. [Google Scholar] [CrossRef]
- Meseguer, M.; Aplin, J.D.; Caballero-Campo, P.; O’Connor, J.E.; Martín, J.C.; Remohí, J.; Pellicer, A.; Simón, C. Human endometrial mucin MUC1 is up-regulated by progesterone and down-regulated in vitro by the human blastocyst. Biol. Reprod. 2001, 64, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Gemzell-Danielsson, K.; Swahn, M.L.; Svalander, P.; Bygdeman, M. Early luteal phase treatment with mifepristone (RU 486) for fertility regulation. Hum. Reprod. 1993, 8, 870–873. [Google Scholar] [CrossRef]
- Gemzell-Danielsson, K.; Svalander, P.; Swahn, M.L.; Johannisson, E.; Bygdeman, M. Effects of a single post-ovulatory dose of RU486 on endometrial maturation in the implantation phase. Hum. Reprod. 1994, 9, 2398–2404. [Google Scholar] [CrossRef]
- Ruiz-Alonso, M.; Valbuena, D.; Gomez, C.; Cuzzi, J.; Simon, C. Endometrial Receptivity Analysis (ERA): Data versus opinions. Hum. Reprod. Open 2021, 2021, hoab011. [Google Scholar] [CrossRef]
- Simón, C.; Cano, F.; Valbuena, D.; Remohí, J.; Pellicer, A. Clinical evidence for a detrimental effect on uterine receptivity of high serum oestradiol concentrations in high and normal responder patients. Hum. Reprod. 1995, 10, 2432–2437. [Google Scholar] [CrossRef]
- Simón, C.; Garcia Velasco, J.J.; Valbuena, D.; Peinado, J.A.; Moreno, C.; Remohí, J.; Pellicer, A. Increasing uterine receptivity by decreasing estradiol levels during the preimplantation period in high responders with the use of a follicle-stimulating hormone step-down regimen. Fertil. Steril. 1998, 70, 234–239. [Google Scholar] [CrossRef]
- Simon, C.; Domínguez, F.; Valbuena, D.; Pellicer, A. The role of estrogen in uterine receptivity and blastocyst implantation. Trends Endocrinol. Metab. 2003, 14, 197–199. [Google Scholar] [CrossRef]
- Horcajadas, J.A.; Riesewijk, A.; Polman, J.; van Os, R.; Pellicer, A.; Mosselman, S.; Simón, C. Effect of controlled ovarian hyperstimulation in IVF on endometrial gene expression profiles. Mol. Hum. Reprod. 2005, 11, 195–205. [Google Scholar] [CrossRef]
- Bosch, E.; Labarta, E.; Crespo, J.; Simón, C.; Remohí, J.; Jenkins, J.; Pellicer, A. Circulating progesterone levels and ongoing pregnancy rates in controlled ovarian stimulation cycles for in vitro fertilization: Analysis of over 4000 cycles. Hum. Reprod. 2010, 25, 2092–2100. [Google Scholar] [CrossRef] [PubMed]
- Lawrenz, B.; Melado, L.; Fatemi, H. Premature progesterone rise in ART-cycles. Reprod. Biol. 2018, 18, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Hamori, M.; Stuckensen, J.A.; Rumpf, D.; Kniewald, T.; Kniewald, A.; Kurz, C.S. Premature luteinization of follicles during ovarian stimulation for in-vitro fertilization. Hum. Reprod. 1987, 2, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Schoolcraft, W.; Sinton, E.; Schlenker, T.; Huynh, D.; Hamilton, F.; Meldrum, D.R. Lower pregnancy rate with premature luteinization during pituitary suppression with leuprolide acetate. Fertil. Steril. 1991, 55, 563–566. [Google Scholar] [CrossRef]
- Silverberg, K.M.; Burns, W.N.; Olive, D.L.; Riehl, R.M.; Schenken, R.S. Serum progesterone levels predict success of in vitro fertilization/embryo transfer in patients stimulated with leuprolide acetate and human menopausal gonadotropins. J. Clin. Endocrinol. Metab. 1991, 73, 797–803. [Google Scholar] [CrossRef]
- Urbancsek, J.; Rabe, T.; Grunwald, K.; Kiesel, L.; Runnebaum, B. Analysis of hormonal changes during combined buserelin/HMG treatment. Hum. Reprod. 1990, 5, 675–681. [Google Scholar] [CrossRef]
- Fanchin, R.; Righini, C.; Olivennes, F.; de Ziegler, D.; Selva, J.; Frydman, R. Premature progesterone elevation does not alter oocyte quality in in vitro fertilization. Fertil. Steril. 1996, 65, 1178–1183. [Google Scholar] [CrossRef]
- Trounson, A.O.; Calabrese, R. Changes in plasma progesterone concentrations around the time of the luteinizing hormone surge in women superovulated for in vitro fertilization. J. Clin. Endocrinol. Metab. 1984, 59, 1075–1080. [Google Scholar] [CrossRef]
- Loumaye, E.; de Cooman, S.; Anoma, M.; Psalti, I.; Depreester, S.; Schmit, M.; Thomas, K. Short-term utilization of a gonadotropin-releasing hormone agonist (buserelin) for induction of ovulation in an in vitro fertilization program. Ann. N. Y. Acad. Sci. 1988, 541, 96–102. [Google Scholar] [CrossRef]
- Macnamee, M.C.; Howles, C.M.; Edwards, R.G. Pregnancies after IVF when high tonic LH is reduced by long-term treatment with GnRH agonists. Hum. Reprod. 1987, 2, 569–571. [Google Scholar] [CrossRef]
- Fanchin, R.; de Ziegler, D.; Castracane, V.D.; Taieb, J.; Olivennes, F.; Frydman, R. Physiopathology of premature progesterone elevation. Fertil. Steril. 1995, 64, 796–801. [Google Scholar] [CrossRef]
- Andersen, A.N.; Devroey, P.; Arce, J.C. Clinical outcome following stimulation with highly purified hMG or recombinant FSH in patients undergoing IVF: A randomized assessor-blind controlled trial. Hum. Reprod. 2006, 21, 3217–3227. [Google Scholar] [CrossRef] [PubMed]
- Smitz, J.; Andersen, A.N.; Devroey, P.; Arce, J.C. Endocrine profile in serum and follicular fluid differs after ovarian stimulation with HP-hMG or recombinant FSH in IVF patients. Hum. Reprod. 2007, 22, 676–687. [Google Scholar] [CrossRef] [PubMed]
- Bosch, E.; Vidal, C.; Labarta, E.; Simon, C.; Remohi, J.; Pellicer, A. Highly purified hMG versus recombinant FSH in ovarian hyperstimulation with GnRH antagonists—A randomized study. Hum. Reprod. 2008, 23, 2346–2351. [Google Scholar] [CrossRef]
- Fanchin, R.; Righini, C.; Olivennes, F.; Taieb, J.; de Ziegler, D.; Frydman, R. Premature plasma progesterone and androgen elevation are not prevented by adrenal suppression in in vitro fertilization. Fertil. Steril. 1997, 67, 115–119. [Google Scholar] [CrossRef]
- Mirkin, S.; Nikas, G.; Hsiu, J.G.; Díaz, J.; Oehninger, S. Gene expression profiles and structural/functional features of the peri-implantation endometrium in natural and gonadotropin-stimulated cycles. J. Clin. Endocrinol. Metab. 2004, 89, 5742–5752. [Google Scholar] [CrossRef]
- Horcajadas, J.A.; Mínguez, P.; Dopazo, J.; Esteban, F.J.; Domínguez, F.; Giudice, L.C.; Pellicer, A.; Simón, C. Controlled ovarian stimulation induces a functional genomic delay of the endometrium with potential clinical implications. J. Clin. Endocrinol. Metab. 2008, 93, 4500–4510. [Google Scholar] [CrossRef]
- Macklon, N.S.; Stouffer, R.L.; Giudice, L.C.; Fauser, B.C. The science behind 25 years of ovarian stimulation for in vitro fertilization. Endocr. Rev. 2006, 27, 170–207. [Google Scholar] [CrossRef]
- Bu, Z.; Zhao, F.; Wang, K.; Guo, Y.; Su, Y.; Zhai, J.; Sun, Y. Serum progesterone elevation adversely affects cumulative live birth rate in different ovarian responders during in vitro fertilization and embryo transfer: A large retrospective study. PLoS ONE 2014, 9, e100011. [Google Scholar] [CrossRef]
- Huang, B.; Ren, X.; Wu, L.; Zhu, L.; Xu, B.; Li, Y.; Ai, J.; Jin, L. Elevated Progesterone Levels on the Day of Oocyte Maturation May Affect Top Quality Embryo IVF Cycles. PLoS ONE 2016, 11, e0145895. [Google Scholar] [CrossRef]
- Adda-Herzog, E.; Poulain, M.; de Ziegler, D.; Ayoubi, J.M.; Fanchin, R. Premature progesterone elevation in controlled ovarian stimulation: To make a long story short. Fertil. Steril. 2018, 109, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Fanchin, R.; Hourvitz, A.; Olivennes, F.; Taieb, J.; Hazout, A.; Frydman, R. Premature progesterone elevation spares blastulation but not pregnancy rates in in vitro fertilization with coculture. Fertil. Steril. 1997, 68, 648–652. [Google Scholar] [CrossRef] [PubMed]
- Vanni, V.S.; Somigliana, E.; Reschini, M.; Pagliardini, L.; Marotta, E.; Faulisi, S.; Paffoni, A.; Vigano, P.; Vegetti, W.; Candiani, M.; et al. Top quality blastocyst formation rates in relation to progesterone levels on the day of oocyte maturation in GnRH antagonist IVF/ICSI cycles. PLoS ONE 2017, 12, e0176482. [Google Scholar] [CrossRef] [PubMed]
- Teh, W.T.; McBain, J.; Rogers, P. What is the contribution of embryo-endometrial asynchrony to implantation failure? J. Assist. Reprod. Genet. 2016, 33, 1419–1430. [Google Scholar] [CrossRef] [PubMed]
- Healy, M.W.; Yamasaki, M.; Patounakis, G.; Richter, K.S.; Devine, K.; DeCherney, A.H.; Hill, M.J. The slow growing embryo and premature progesterone elevation: Compounding factors for embryo-endometrial asynchrony. Hum. Reprod. 2017, 32, 362–367. [Google Scholar] [CrossRef]
- Tavaniotou, A.; Albano, C.; Smitz, J.; Devroey, P. Impact of ovarian stimulation on corpus luteum function and embryonic implantation. J. Reprod. Immunol. 2002, 55, 123–130. [Google Scholar] [CrossRef]
- Ubaldi, F.; Bourgain, C.; Tournaye, H.; Smitz, J.; Van Steirteghem, A.; Devroey, P. Endometrial evaluation by aspiration biopsy on the day of oocyte retrieval in the embryo transfer cycles in patients with serum progesterone rise during the follicular phase. Fertil. Steril. 1997, 67, 521–526. [Google Scholar] [CrossRef]
- Ezra, Y.; Simon, A.; Sherman, Y.; Benshushan, A.; Younis, J.S.; Laufer, N. The effect of progesterone administration in the follicular phase of an artificial cycle on endometrial morphology: A model of premature luteinization. Fertil. Steril. 1994, 62, 108–112. [Google Scholar] [CrossRef]
- Chetkowski, R.J.; Kiltz, R.J.; Salyer, W.R. In premature luteinization, progesterone induces secretory transformation of the endometrium without impairment of embryo viability. Fertil. Steril. 1997, 68, 292–297. [Google Scholar] [CrossRef]
- Fanchin, R.; Righini, C.; Olivennes, F.; Taieb, J.; de Ziegler, D.; Frydman, R. Computerized assessment of endometrial echogenicity: Clues to the endometrial effects of premature progesterone elevation. Fertil. Steril. 1999, 71, 174–181. [Google Scholar] [CrossRef]
- Liu, L.; Sailan, S.; Li, T.; Mariee, N.; Laird, S.; Jiang, Z.; Zhang, S. The effect of a high progesterone concentration before oocyte retrieval on the peri-implantation endometrium. Reprod. Biomed. Online 2015, 31, 739–746. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Young, S.L.; Savaris, R.F.; Lessey, B.A.; Sharkey, A.M.; Balthazar, U.; Zaino, R.J.; Sherwin, R.A.; Fritz, M.A. Effect of randomized serum progesterone concentration on secretory endometrial histologic development and gene expression. Hum. Reprod. 2017, 32, 1903–1914. [Google Scholar] [CrossRef]
- Usadi, R.S.; Groll, J.M.; Lessey, B.A.; Lininger, R.A.; Zaino, R.J.; Fritz, M.A.; Young, S.L. Endometrial development and function in experimentally induced luteal phase deficiency. J. Clin. Endocrinol. Metab. 2008, 93, 4058–4064. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, H.; Casper, R.F.; Esfandiari, N.; Bentov, Y. Non-synchronized endometrium and its correction in non-ovulatory cryopreserved embryo transfer cycles. Reprod. Biomed. Online 2015, 30, 378–384. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Navot, D.; Scott, R.T.; Droesch, K.; Veeck, L.L.; Liu, H.C.; Rosenwaks, Z. The window of embryo transfer and the efficiency of human conception in vitro. Fertil. Steril. 1991, 55, 114–118. [Google Scholar] [CrossRef]
- Kolibianakis, E.; Bourgain, C.; Albano, C.; Osmanagaoglu, K.; Smitz, J.; Van Steirteghem, A.; Devroey, P. Effect of ovarian stimulation with recombinant follicle-stimulating hormone, gonadotropin releasing hormone antagonists, and human chorionic gonadotropin on endometrial maturation on the day of oocyte pick-up. Fertil. Steril. 2002, 78, 1025–1029. [Google Scholar] [CrossRef]
- Van Vaerenbergh, I.; Van Lommel, L.; Ghislain, V.; In’t Veld, P.; Schuit, F.; Fatemi, H.M.; Devroey, P.; Bourgain, C. In GnRH antagonist/rec-FSH stimulated cycles, advanced endometrial maturation on the day of oocyte retrieval correlates with altered gene expression. Hum. Reprod. 2009, 24, 1085–1091. [Google Scholar] [CrossRef]
- Díaz-Gimeno, P.; Horcajadas, J.A.; Martínez-Conejero, J.A.; Esteban, F.J.; Alamá, P.; Pellicer, A.; Simón, C. A genomic diagnostic tool for human endometrial receptivity based on the transcriptomic signature. Fertil. Steril. 2011, 95, 50–60.e15. [Google Scholar] [CrossRef]
- Ruiz-Alonso, M.; Blesa, D.; Díaz-Gimeno, P.; Gómez, E.; Fernández-Sánchez, M.; Carranza, F.; Carrera, J.; Vilella, F.; Pellicer, A.; Simón, C. The endometrial receptivity array for diagnosis and personalized embryo transfer as a treatment for patients with repeated implantation failure. Fertil. Steril. 2013, 100, 818–824. [Google Scholar] [CrossRef]
- Ruiz-Alonso, M.; Galindo, N.; Pellicer, A.; Simón, C. What a difference two days make: “personalized” embryo transfer (pET) paradigm: A case report and pilot study. Hum. Reprod. 2014, 29, 1244–1247. [Google Scholar] [CrossRef]
- Xiong, Y.; Hu, L.; Zhang, T.; Wang, M.; Xu, H.; Li, T.C.; Sun, Y.; Wang, C.C. Effects of high progesterone in in-vitro fertilization cycle on DNA methylation and gene expression of adhesion molecules on endometrium during implantation window. J. Assist. Reprod. Genet 2020, 37, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.B.; Healy, M.W.; DeCherney, A.H.; Hill, M.J. Adverse effect of prematurely elevated progesterone in in vitro fertilization cycles: A literature review. Biol. Reprod. 2018, 99, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Roque, M.; Lattes, K.; Serra, S.; Sola, I.; Geber, S.; Carreras, R.; Checa, M.A. Fresh embryo transfer versus frozen embryo transfer in in vitro fertilization cycles: A systematic review and meta-analysis. Fertil. Steril. 2013, 99, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Yoshinaga, K. A historical review of blastocyst implantation research. Biol. Reprod. 2018, 99, 175–195. [Google Scholar] [CrossRef]
- Horcajadas, J.A.; Minguez, P.; Dopazo, J.; Esteban, F.J.; Pellicer, A.; Simon, C. Gene expression analysis of the endometrium reveals that controlled ovarian stimulation induces a genomic delay with potential clinical implications. Fertil. Steril. 2007, 88, S43–S44. [Google Scholar] [CrossRef]
- Labarta, E.; Martínez-Conejero, J.A.; Alamá, P.; Horcajadas, J.A.; Pellicer, A.; Simón, C.; Bosch, E. Endometrial receptivity is affected in women with high circulating progesterone levels at the end of the follicular phase: A functional genomics analysis. Hum. Reprod. 2011, 26, 1813–1825. [Google Scholar] [CrossRef]
- Haouzi, D.; Bissonnette, L.; Gala, A.; Assou, S.; Entezami, F.; Perrochia, H.; Dechaud, H.; Hugues, J.N.; Hamamah, S. Endometrial receptivity profile in patients with premature progesterone elevation on the day of HCG administration. BioMed Res. Int. 2014, 2014, 951937. [Google Scholar] [CrossRef]
- Altmae, S.; Martinez-Conejero, J.A.; Salumets, A.; Simon, C.; Horcajadas, J.A.; Stavreus-Evers, A. Endometrial gene expression analysis at the time of embryo implantation in women with unexplained infertility. Mol. Hum. Reprod. 2010, 16, 178–187. [Google Scholar] [CrossRef]
- Talbi, S.; Hamilton, A.E.; Vo, K.C.; Tulac, S.; Overgaard, M.T.; Dosiou, C.; Le Shay, N.; Nezhat, C.N.; Kempson, R.; Lessey, B.A.; et al. Molecular phenotyping of human endometrium distinguishes menstrual cycle phases and underlying biological processes in normo-ovulatory women. Endocrinology 2006, 147, 1097–1121. [Google Scholar] [CrossRef]
- Du, H.; Taylor, H.S. The Role of Hox Genes in Female Reproductive Tract Development, Adult Function, and Fertility. Cold Spring Harb. Perspect. Med. 2015, 6, a023002. [Google Scholar] [CrossRef]
- Dennis, G., Jr.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol. 2003, 4, R60. [Google Scholar] [CrossRef]
- Horcajadas, J.A.; Pellicer, A.; Simon, C. Wide genomic analysis of human endometrial receptivity: New times, new opportunities. Hum. Reprod. Update 2007, 13, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Wang, J.; Liu, L.; Chen, X.; Xu, H.; Li, T.C.; Wang, C.C.; Zhang, S. Effects of high progesterone level on the day of human chorionic gonadotrophin administration in in vitro fertilization cycles on epigenetic modification of endometrium in the peri-implantation period. Fertil. Steril. 2017, 108, 269–276.e1. [Google Scholar] [CrossRef] [PubMed]
- Bienert, M.; Habib, P.; Buck, V.; Classen-Linke, I.; Skoblo, R.; Rösing, B. Intrauterine hCG application increases expression of endothelial cell-cell adhesion molecules in human. Arch. Gynecol. Obstet. 2021, 304, 1587–1597. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Santos, L.; Dimitriadis, E. Characterization of the role for cadherin 6 in the regulation of human endometrial receptivity. Reprod. Biol. Endocrinol. 2020, 18, 66. [Google Scholar] [CrossRef]
- Rahnama, F.; Thompson, B.; Steiner, M.; Shafiei, F.; Lobie, P.E.; Mitchell, M.D. Epigenetic regulation of E-cadherin controls endometrial receptivity. Endocrinology 2009, 150, 1466–1472. [Google Scholar] [CrossRef]
- Zhou, X.; Xu, B.; Zhang, D.; Jiang, X.; Chang, H.M.; Leung, P.C.K.; Xia, X.; Zhang, A. Loss of CDYL Results in Suppression of CTNNB1 and Decreased Endometrial Receptivity. Front. Cell Dev. Biol. 2020, 8, 105. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, L.; Li, T.C.; Zhu, H.; Lin, X.; Jin, X.; Tong, X.; Zhang, S. Comparison of progesterone measurement on day of, and day after, HCG administration in IVF-embryo transfer cycles. Reprod. Biomed. Online 2015, 30, 157–165. [Google Scholar] [CrossRef]
- Hofmann, G.E.; Bentzien, F.; Bergh, P.A.; Garrisi, G.J.; Williams, M.C.; Guzman, I.; Navot, D. Premature luteinization in controlled ovarian hyperstimulation has no adverse effect on oocyte and embryo quality. Fertil. Steril. 1993, 60, 675–679. [Google Scholar] [CrossRef]
- Silverberg, K.M.; Martin, M.; Olive, D.L.; Burns, W.N.; Schenken, R.S. Elevated serum progesterone levels on the day of human chorionic gonadotropin administration in in vitro fertilization cycles do not adversely affect embryo quality. Fertil. Steril. 1994, 61, 508–513. [Google Scholar] [CrossRef]
- Legro, R.S.; Ary, B.A.; Paulson, R.J.; Stanczyk, F.Z.; Sauer, M.V. Premature luteinization as detected by elevated serum progesterone is associated with a higher pregnancy rate in donor oocyte in-vitro fertilization. Hum. Reprod. 1993, 8, 1506–1511. [Google Scholar] [CrossRef] [PubMed]
- Baldini, D.; Savoia, M.V.; Sciancalepore, A.G.; Malvasi, A.; Vizziello, D.; Beck, R.; Vizziello, G. High progesterone levels on the day of HCG administration do not affect the embryo quality and the reproductive outcomes of frozen embryo transfers. Clin. Ter. 2018, 169, e91–e95. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Nieto, C.; Lee, J.A.; Alkon-Meadows, T.; Luna-Rojas, M.; Mukherjee, T.; Copperman, A.B.; Sandler, B. Late follicular phase progesterone elevation during ovarian stimulation is not associated with decreased implantation of chromosomally screened embryos in thaw cycles. Hum. Reprod. 2020, 35, 1889–1899. [Google Scholar] [CrossRef] [PubMed]
- Melo, M.A.; Meseguer, M.; Garrido, N.; Bosch, E.; Pellicer, A.; Remohi, J. The significance of premature luteinization in an oocyte-donation programme. Hum. Reprod. 2006, 21, 1503–1507. [Google Scholar] [CrossRef]
- Hill, M.J.; Royster, G.D.T.; Healy, M.W.; Richter, K.S.; Levy, G.; DeCherney, A.H.; Levens, E.D.; Suthar, G.; Widra, E.; Levy, M.J. Are good patient and embryo characteristics protective against the negative effect of elevated progesterone level on the day of oocyte maturation? Fertil. Steril. 2015, 103, 1477–1484.e5. [Google Scholar] [CrossRef]
- Pardinas, M.L.; Nohales, M.; Labarta, E.; De Los Santos, J.M.; Mercader, A.; Remohi, J.; Bosch, E.; De Los Santos, M.J. Elevated serum progesterone does not impact euploidy rates in PGT-A patients. J. Assist. Reprod. Genet. 2021, 38, 1819–1826. [Google Scholar] [CrossRef]
- Neves, A.R.; Santos-Ribeiro, S.; Garcia-Martinez, S.; Devesa, M.; Soares, S.R.; Garcia-Velasco, J.A.; Garrido, N.; Polyzos, N.P. The effect of late-follicular phase progesterone elevation on embryo ploidy and cumulative live birth rates. Reprod. Biomed. Online 2021, 43, 1063–1069. [Google Scholar] [CrossRef]
- Xu, B.; Li, Z.; Zhang, H.; Jin, L.; Li, Y.; Ai, J.; Zhu, G. Serum progesterone level effects on the outcome of in vitro fertilization in patients with different ovarian response: An analysis of more than 10,000 cycles. Fertil. Steril. 2012, 97, 1321–1327. [Google Scholar] [CrossRef]

| Study/Study Type | ART Cycle | Inclusion Criteria | Exclusion Criteria | Pituitary Desensitization Protocol | Ovarian Stimulation Protocol | Trigger | ET/luteal Support | Estrogen Administration | Study Groups: Progesterone Treatment or Blood Levels | Control Groups: Progesterone Treatment or Blood Levels | Day of Estrogen and Progesterone Measurements | Day(s) of Endometrial Biopsy or Ultrasound/Method Used for Assessing Endometrial Histologic Development |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ezra et al., 1994 [48]/ prospective or retrospective was not stated | 26-day artificial cycle (follicular and luteal phase) in the setting of ovarian failure | 27–47-y/o women with primary or secondary premature ovarian failure, FSH and LH levels > 50 IU/L and low E levels (<25 pg/mL) | None listed | n.a. | n.a. | n.a. | n.a. | Oral micronized E: 4 mg/day over 26 days | Study Group A (n = 8): administered 12.5 mg P (IM in oil) on days 2 and 7 followed by 50 mg/day from day 15–26. Study Group B (n = 8): administered 6.25 mg P (IM in oil) on days 3, 4, and 5 followed by 50 mg/day from day 15–26 | Subjects had standard preparatory cycles without follicular P supplementation (n = 16). Subjects only administered P (IM in oil) (50 mg/day) from luteal day 15–26 | Serum E and P: days 14 and 26 | 14, 26/endometrial dating |
| Chetkowski et al., 1997 [49]/ prospective | IVF donor cycle | Healthy parous women and infertile women with functioning ovaries | None listed | GNRH agonist (leuprolide acetate) long protocol | hMG | hCG | n.a. | n.a. | n.a. | n.a. | Serum P: 1 and 2 days before hCG administration and on day of hCG administration | 36 h after trigger at the time of oocyte retrieval/endometrial dating |
| Fanchin et al., 1999 [50]/ prospective | IVF/ET | ≤38-y/o women with morphologically healthy uterus and had at least three good-quality embryos | Women whose uterine position did not allow adequate visualization of the endometrial texture at ultrasound examination and those with grossly irregular ultrasonographic appearance of the myometrium | GNRH agonist (leuprolide acetate) long protocol | hMG | hCG | ET performed 2 days after oocyte retrieval/300 mg micronized P administered daily (100 mg in the morning and 200 mg in the evening) starting on the evening of the day of ET | n.a. | Study Group (n = 26): plasma P > 0.9 ng/mL | Control Group (n = 33): plasma P ≤ 0.9 ng/mL | Plasma E and P: day of hCG administration, day of oocyte retrieval, and day of ET | Ultrasound conducted on day of hCG administration, oocyte retrieval and ET/ultrasound-based endometrial echogenicity |
| Liu et al., 2015 [51]/prospective | IVF without ET | 23–40-y/o women with tubal or male infertility | None listed | GNRH agonist (Triptorelin) long protocol | FSH | hCG | P: 40 mg/day for 1 day, starting from the night of oocyte retrieval, and then at 60 mg/day for 2 days, and then at 80 mg/day for 3 days | n.a. | High P Group (n = 58): P = 1.7 ng/mL on the day of hCG administration and 9.5 ng/mL on hCG + 1 | Normal P Group (n = 48): P <1.7 ng/mL on the day of hCG administration and <9.5 ng/mL on hCG + 1 | E and P: 12–14 h before hCG administration and 12–14 h after hCG administration | 7 days after hCG administration/endometrial dating |
| Young et al., 2017 [52]/prospective | Healthy women with induced experimental modeled cycles | 19–34-y/o healthy women with regular intermenstrual interval between 25–35 days and no history of infertility or pelvic disease | An intermenstrual interval that varied by >3 days, use of medication that affects reproductive hormones or fertility within 60 days prior to enrollment, chronic disease, a body mass index >29.9 or <18.5, and history of infertility | GNRH agonist (leuprolide acetate) long protocol | n.a. | n.a. | n.a. | Transdermal 0.2 mg/day for 20 days following pituitary–ovary desensitization | P administered (IM in oil) daily after 10 days of E treatment. Group A (n = 6): P = 2.5 mg/day; Group B (n = 6): P = 5.0 mg/day; Group C (n = 12): 10.0 mg/day; Group D (n = 12): P = 40.0 mg/day | Control group (n = 10): natural cycles exhibiting normal reported levels of P (Nadji et al., 1975) [6] | Serum P in modeled cycles: 2–3 h after injection (peak) and 1–2 h before injection (trough) on two separate occasions between 3 and 10 days of P treatment | Control group: 10 days after the mid-cycle urinary LH surge. Modeled cycles: on P day 10 in subjects receiving 2.5 or 5 mg of P daily and from subjects receiving 10 or 40 mg of P daily/endometrial dating |
| Study/Study Type | ART Cycle | Inclusion Criteria | Exclusion Criteria | Pituitary Desensitization Protocol | Ovarian Stimulation Protocol | Trigger | ET/Luteal Support | Estrogen Administration | Study Groups: Progesterone Treatment or Blood Levels | Control Groups: Progesterone Treatment or Blood Levels | Day of Estrogen and Progesterone Measurements | Day(s) of Endometrial Biopsy or Ultrasound/Method Used for Assessing Endometrial Transcriptome |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Young et al., 2017 [52]/prospective | Healthy women with induced experimental modeled cycles | 19–34-y/o healthy women with regular intermenstrual interval between 25–35 days and no history of infertility or pelvic disease | Intermenstrual interval that varied by >3 days, use of medication that affect reproductive hormones or fertility within 60 days prior to enrollment, chronic disease, a body mass index >29.9 or <18.5, and history of infertility | GNRH agonist (leuprolide acetate) long protocol | n.a. | n.a. | n.a. | Transdermal 0.2 mg/day for 20 days following pituitary–ovary desensitization | P administered (IM in oil) daily after 10 days of E treatment. Group A (n = 6): P = 2.5 mg/day; Group B (n = 6): P = 5.0 mg/day; Group C (n = 12): 10.0 mg/day; Group D (n = 12): P = 40.0 mg/day | Control group (n = 10): natural cycles exhibiting normal reported levels of P (Nadji et al., 1975) [6] | Serum P in modeled cycles: 2–3 h after injection (peak) and 1–2 h before injection (trough) on two separate occasions between 3 and 10 days of P treatment | Control group: 10 days after the mid-cycle urinary LH surge (cycle day 23 in the mid-luteal phase). Modeled cycles: on P day 10 in subjects receiving 2.5 or 5 mg of P daily and from subjects receiving 10 or 40 mg of P daily/microarray hybridization with RT- PCR and in silico comparison to previous studies |
| Labarta et al., 2011 [66]/ prospective | OS without IVF (oocyte donors) | 18–350-y/o women, body mass index 18–25, 25–35-day menstrual cycles, normal basal serum hormone levels on day 3 of the menstrual cycle (FSH < 10 IU/L, LH < 10 IU/L, and E2 < 60 pg/mL, normal karyotype | Endometriosis, polycystic ovarian syndrome | GNRH agonist (leuprolide acetate) long protocol or GnRH antagonist (Cetrotide) | rFSH | hCG | No ET performed luteal support with 400 mg/day of micronized P administered 1 day after oocyte retrieval to simulate ET cycles | n.a. | High P Group (n = 6): P > 1.5 ng/mL on the day of hCG trigger | High P Group (n = 6): P > 1.5 ng/mL on the day of hCG trigger | Serum E2 on day 3 of menstrual cycle Serum P on day of hCG administration | 7 days after hCG administration (hCG + 7)/microarray hybridization |
| Haouzi, et al., 2014 [67]/ prospective | IVF/ET | 28–34-y/o women, male factor infertility, normal serum FSH, LH, and E2 on day 3 of OS and on the day of hCG administration | No specific criteria listed | GNRH agonist long protocol or GnRH antagonist | hMG | hCG | ET performed 3 days after oocyte retrieval/luteal support not specified | n.a. | High P Group (n = 8): P > 1.5 ng/mL on the day of hCG trigger | Control Group (n = 7): P < 1.5 ng/mL on the day of hCG trigger | Serum E2 on day 3 of OS Serum P on day of hCG administration | Pre-receptive (hCG + 2) and receptive (hCG + 5) secretory stages/microarray hybridization with RT- qPCR |
| Xiong, et al., 2020 [61]/ prospective | IVF without ET | 24–40-y/o women, body mass index 18–25, 25–35-day menstrual cycles, normal basal serum hormone levels on days 2–4 of the menstrual cycle (FSH < 10 IU/L and E2 < 60 pg/mL, tubal or male factor infertility, normal karyotype in both partners | Polycystic ovarian syndrome, hydrosalpinx, uterine abnormalities, thyroid dysfunction, recurrent miscarriage | GNRH agonist super-long or long protocol | FSH | hCG | No ET performed/luteal support not received | n.a. | High P Group (n = 20): P ≥ 1.7 ng/mL on day of hCG administration | Normal P Group (n = 20): P < 1.7 ng/mL on the day of hCG administration | Serum E2 on days 2–4 of menstrual cycle Serum P every 2–4 days during OS and on day of hCG administration | 7 days after hCG administration/Sequenom MALDI-TOF mass spectrometry or bisulfate sequencing PCR, and immunohistochemistry |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalakota, N.R.; George, L.C.; Morelli, S.S.; Douglas, N.C.; Babwah, A.V. Towards an Improved Understanding of the Effects of Elevated Progesterone Levels on Human Endometrial Receptivity and Oocyte/Embryo Quality during Assisted Reproductive Technologies. Cells 2022, 11, 1405. https://doi.org/10.3390/cells11091405
Kalakota NR, George LC, Morelli SS, Douglas NC, Babwah AV. Towards an Improved Understanding of the Effects of Elevated Progesterone Levels on Human Endometrial Receptivity and Oocyte/Embryo Quality during Assisted Reproductive Technologies. Cells. 2022; 11(9):1405. https://doi.org/10.3390/cells11091405
Chicago/Turabian StyleKalakota, Nischelle R., Lea C. George, Sara S. Morelli, Nataki C. Douglas, and Andy V. Babwah. 2022. "Towards an Improved Understanding of the Effects of Elevated Progesterone Levels on Human Endometrial Receptivity and Oocyte/Embryo Quality during Assisted Reproductive Technologies" Cells 11, no. 9: 1405. https://doi.org/10.3390/cells11091405
APA StyleKalakota, N. R., George, L. C., Morelli, S. S., Douglas, N. C., & Babwah, A. V. (2022). Towards an Improved Understanding of the Effects of Elevated Progesterone Levels on Human Endometrial Receptivity and Oocyte/Embryo Quality during Assisted Reproductive Technologies. Cells, 11(9), 1405. https://doi.org/10.3390/cells11091405