
Salvia miltiorrhiza Bunge as a Potential Natural Compound against COVID-19

 and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. The S. miltiorrhiza Root Extract
2.2. Analysis of the S. miltiorrhiza Root Extract (UHPLC)
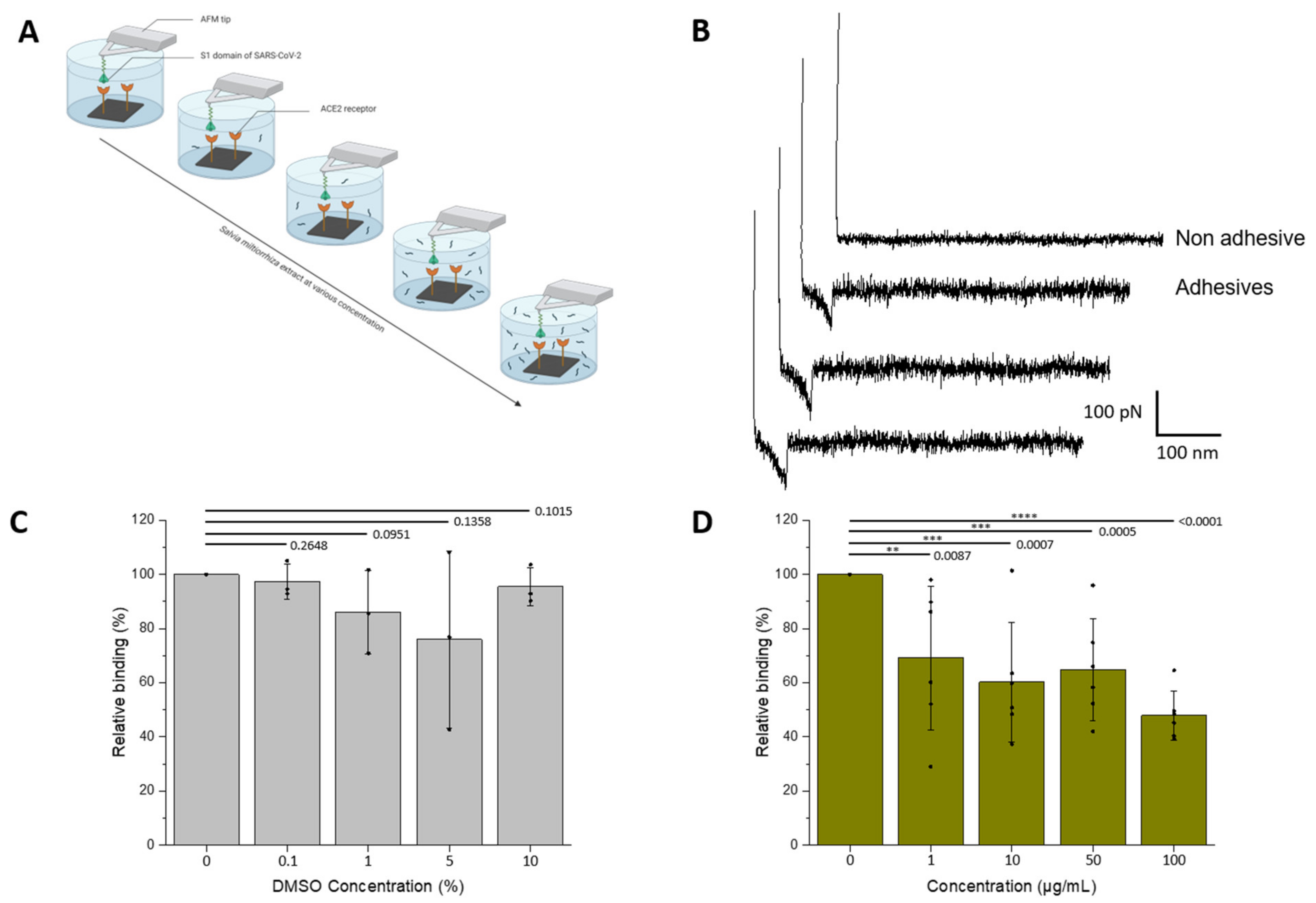
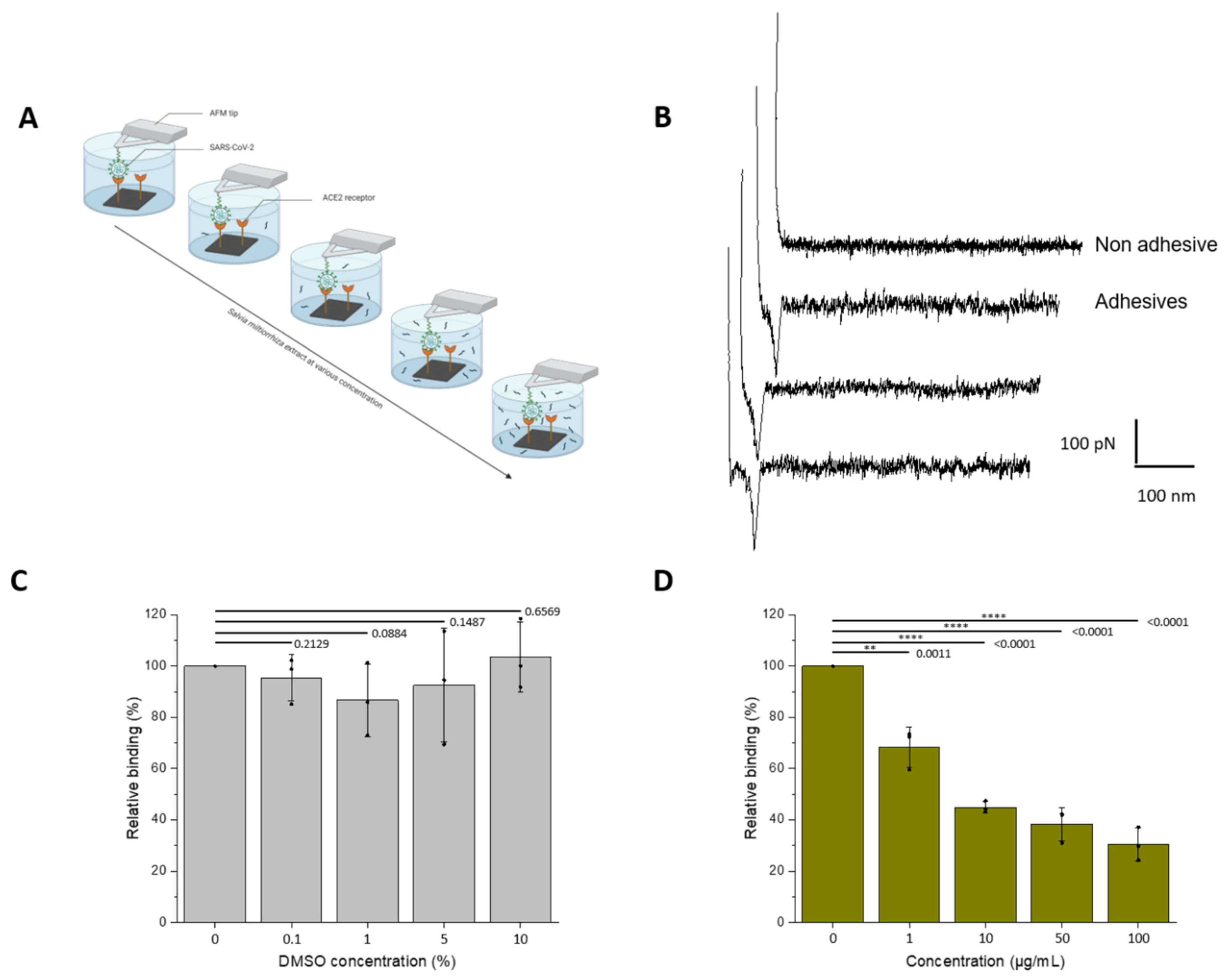
2.3. Preparation of ACE2-Coated Model Surfaces
2.4. Tip Functionalization
2.5. Preparation of S. miltiorrhiza Stock Solutions
2.6. FD Curve-Based Atomic Force Microscopy (AFM)
2.7. Isolation and Culture of Peripheral Blood Mononuclear Cells (PBMC)
2.8. Cytokine Release Assays
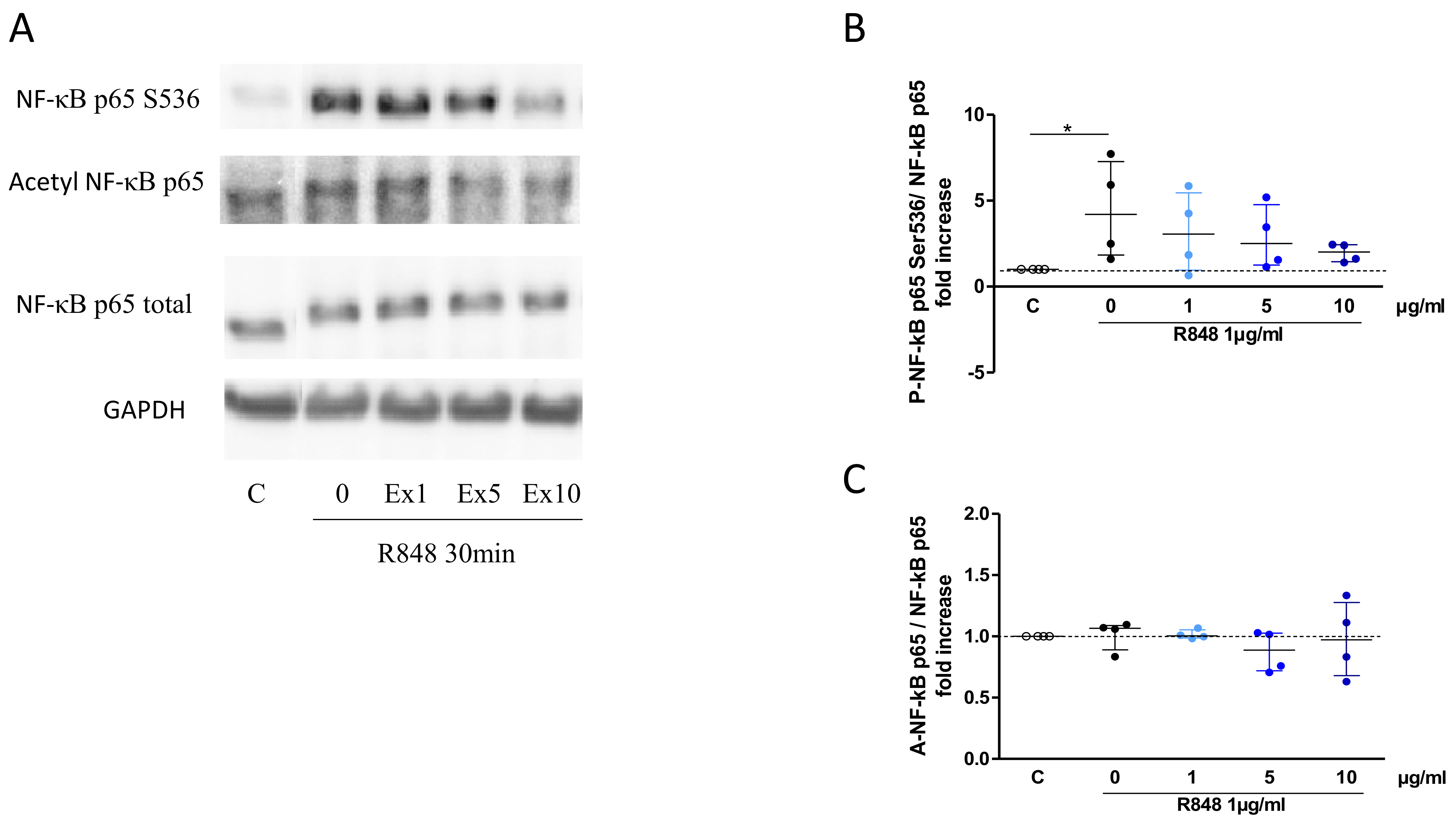
2.9. Western Blot for NF-κB Signalling
2.10. Statistics
3. Results
3.1. Characterization of the S. miltiorrhiza Root Powder Extract
3.2. Assessment of the Inhibitory Effect of the Interaction between the S1-Subunit and ACE2
3.3. Assessment of the Inhibitory Effect of the Interaction between the UV-Inactivated SARS-CoV-2 Virions and ACE2
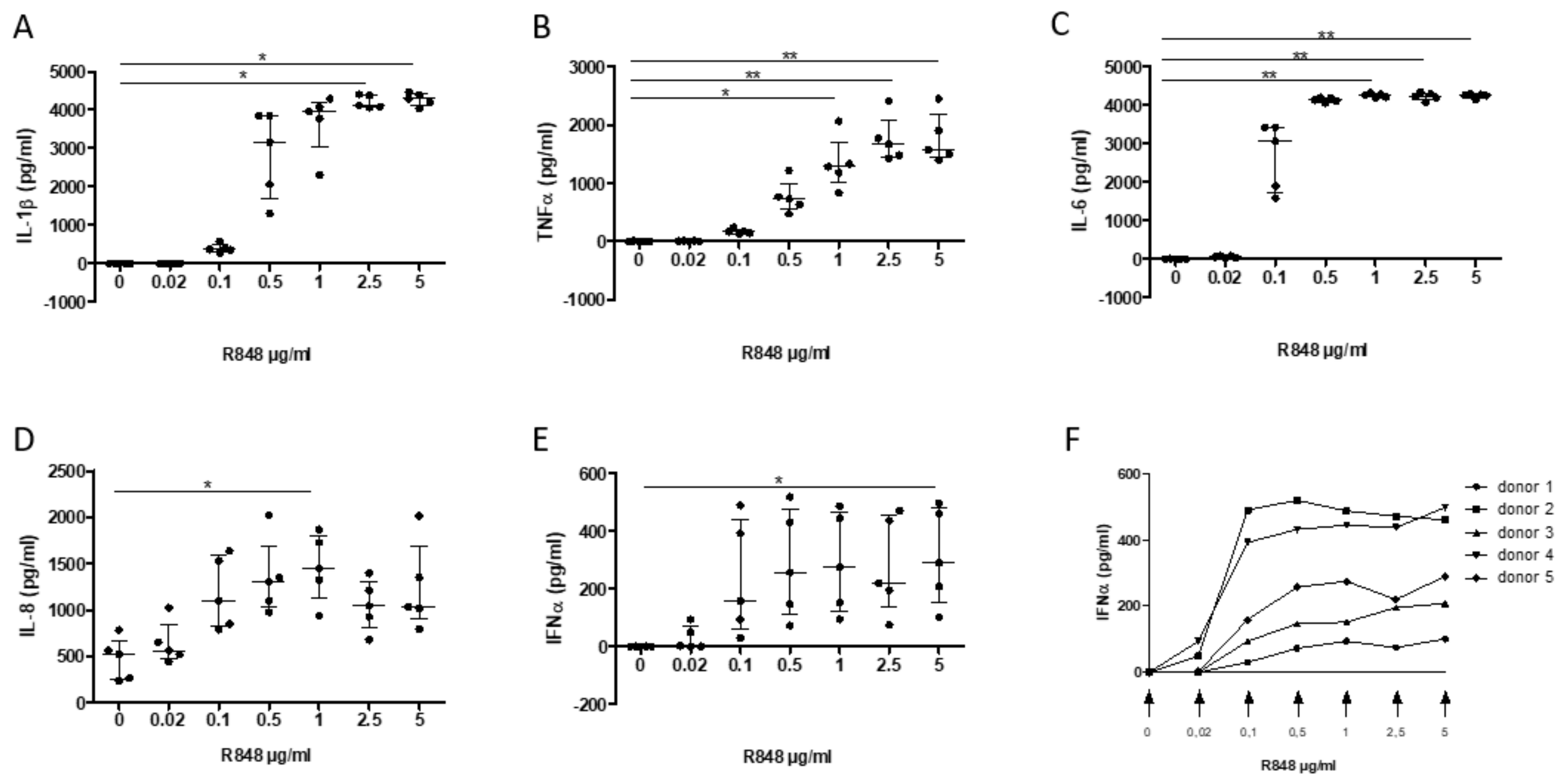
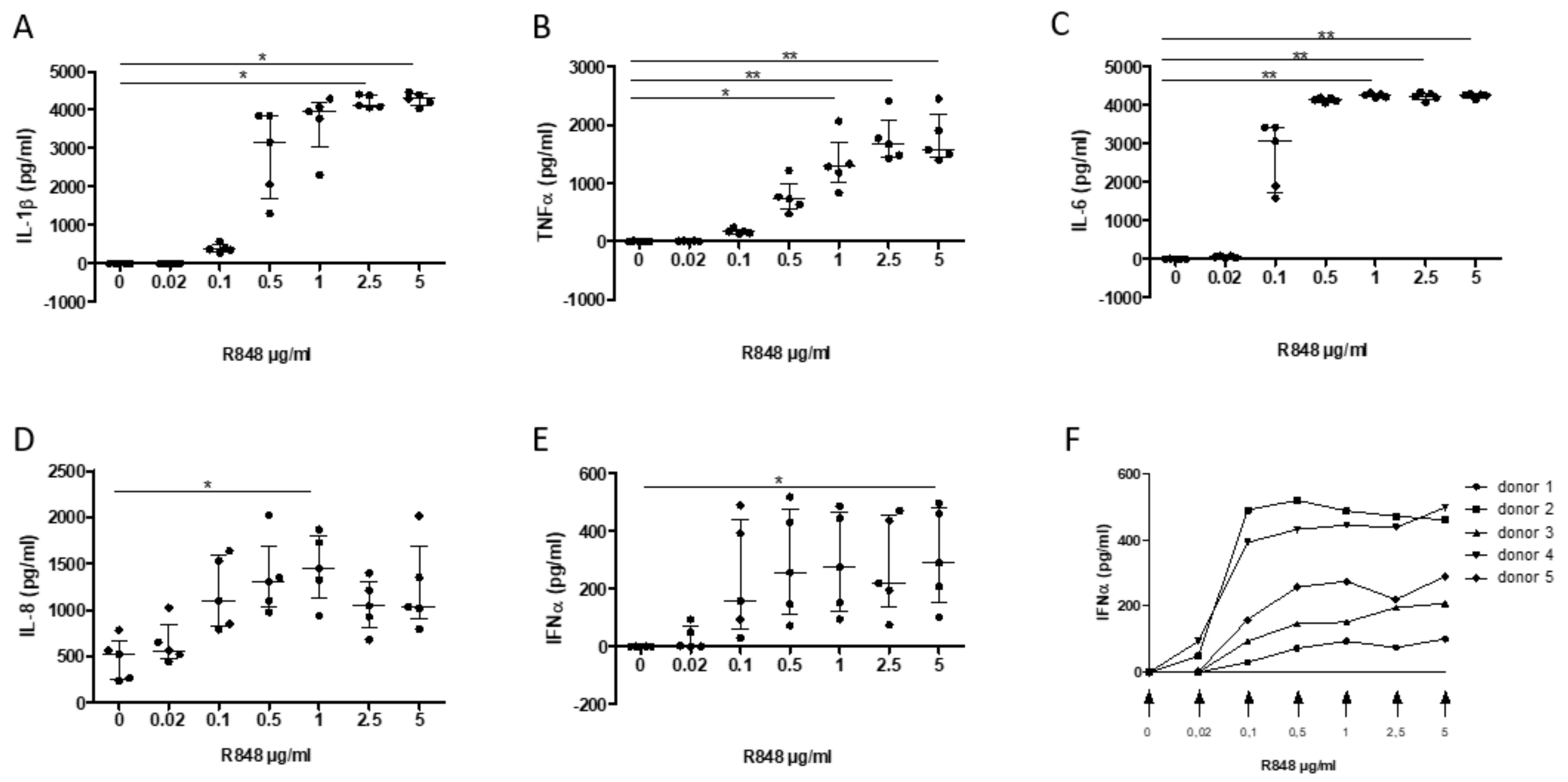
3.4. Establishment of an In Vitro Cell-Based Assay of TLR7/8 Activation of PBMC Mimicking the Inflammatory Response to SARS-CoV Infection
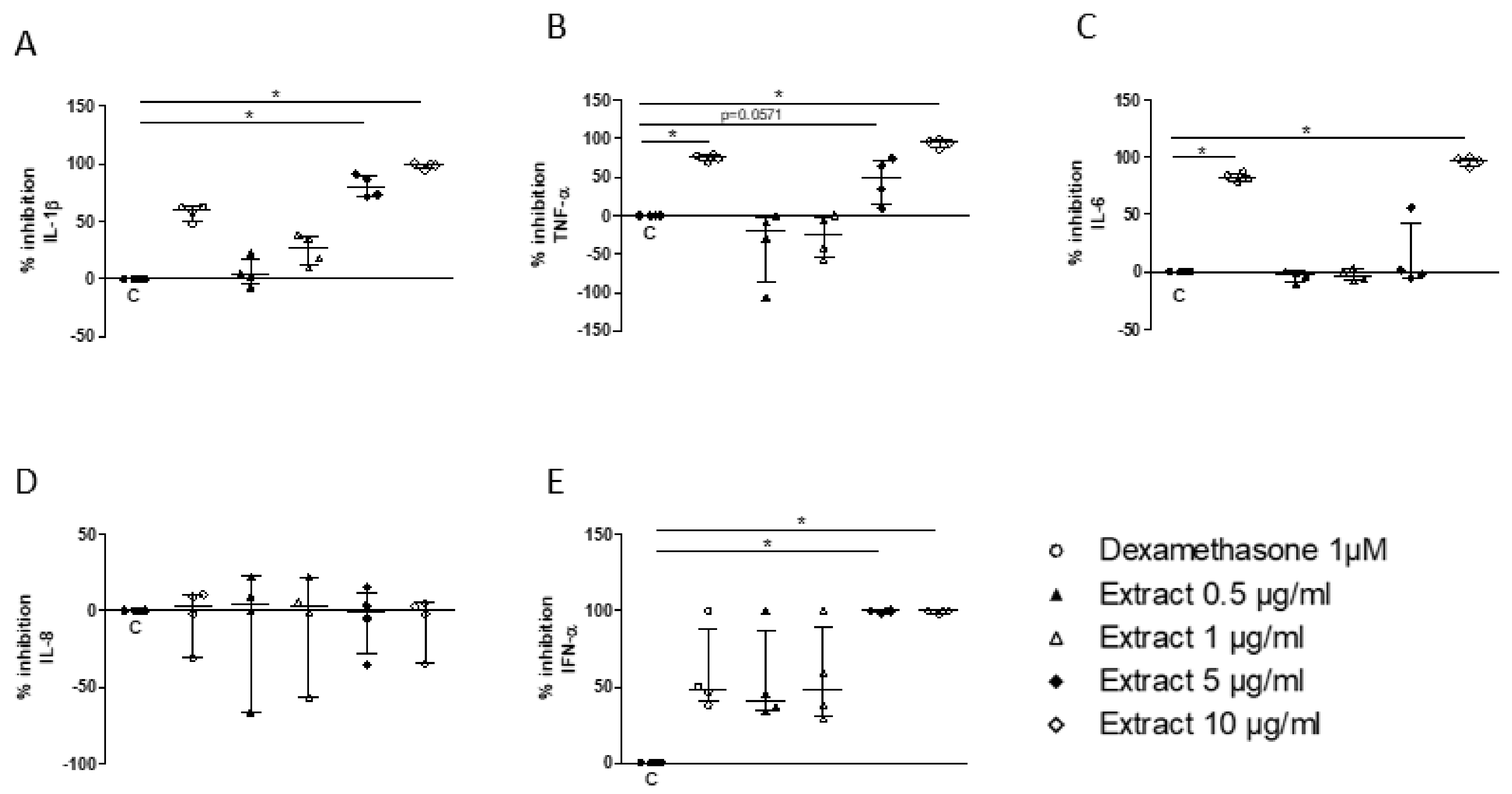
3.5. Assessment of the Inhibitory Effect of a S. miltiorrhiza Extract on the PBMC Response to TLR7/8 Activation
3.6. Assessment of the Mechanism of Inhibition of the Cell Response to TLR7/8 Activation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, J.L.; Li, W.X.; Li, Y.; Wong, M.S.; Wang, Y.J.; Zhang, Y. Therapeutic options of TCM for organ injuries associated with COVID-19 and the underlying mechanism. Phytomedicine 2021, 85, 153297. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Manheimer, E.; Shi, Y.; Gluud, C. Chinese herbal medicine for severe acute respiratory syndrome: A systematic review and meta-analysis. J. Altern. Complement. Med. 2004, 10, 1041–1051. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Tang, Q.L.; Shang, Y.X.; Liang, S.B.; Yang, M.; Robinson, N.; Liu, J.P. Can Chinese Medicine Be Used for Prevention of Corona Virus Disease 2019 (COVID-19)? A Review of Historical Classics, Research Evidence and Current Prevention Programs. Chin. J. Integr. Med. 2020, 26, 243–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, C.Y.; Ming, Q.L.; Rahman, K.; Han, T.; Qin, L.P. Salvia miltiorrhiza: Traditional medicinal uses, chemistry, and pharmacology. Chin. J. Nat. Med. 2015, 13, 163–182. [Google Scholar] [CrossRef]
- Wang, J.W.; Wu, J.Y. Tanshinone biosynthesis in Salvia miltiorrhiza and production in plant tissue cultures. Appl. Microbiol. Biotechnol. 2010, 88, 437–449. [Google Scholar] [CrossRef]
- Wang, C.H.; Zheng, L.P.; Tian, H.; Wang, J.W. Synergistic effects of ultraviolet-B and methyl jasmonate on tanshinone biosynthesis in Salvia miltiorrhiza hairy roots. J. Photochem. Photobiol. B 2016, 159, 93–100. [Google Scholar] [CrossRef]
- Zhang, D.H.; Wu, K.L.; Zhang, X.; Deng, S.Q.; Peng, B. In silico screening of Chinese herbal medicines with the potential to directly inhibit 2019 novel coronavirus. J. Integr. Med. 2020, 18, 152–158. [Google Scholar] [CrossRef]
- Park, J.Y.; Kim, J.H.; Kim, Y.M.; Jeong, H.J.; Kim, D.W.; Park, K.H.; Kwon, H.J.; Park, S.J.; Lee, W.S.; Ryu, Y.B. Tanshinones as selective and slow-binding inhibitors for SARS-CoV cysteine proteases. Bioorg. Med. Chem. 2012, 20, 5928–5935. [Google Scholar] [CrossRef]
- Hu, S.; Wang, J.; Zhang, Y.; Bai, H.; Wang, C.; Wang, N.; He, L. Three salvianolic acids inhibit 2019-nCoV spike pseudovirus viropexis by binding to both its RBD and receptor ACE2. J. Med. Virol. 2021, 93, 3143–3151. [Google Scholar] [CrossRef]
- Zhang, Q.; Gan, C.; Liu, H.; Wang, L.; Li, Y.; Tan, Z.; You, J.; Yao, Y.; Xie, Y.; Yin, W.; et al. Cryptotanshinone reverses the epithelial-mesenchymal transformation process and attenuates bleomycin-induced pulmonary fibrosis. Phytother. Res. 2020, 34, 2685–2696. [Google Scholar] [CrossRef]
- Wu, H.; Li, Y.; Wang, Y.; Xu, D.; Li, C.; Liu, M.; Sun, X.; Li, Z. Tanshinone IIA attenuates bleomycin-induced pulmonary fibrosis via modulating angiotensin-converting enzyme 2/angiotensin-(1-7) axis in rats. Int. J. Med. Sci. 2014, 11, 578–586. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Petitjean, S.J.; Koehler, M.; Zhang, Q.; Dumitru, A.C.; Chen, W.; Derclaye, S.; Vincent, S.P.; Soumillion, P.; Alsteens, D. Molecular interaction and inhibition of SARS-CoV-2 binding to the ACE2 receptor. Nat. Commun. 2020, 11, 4541. [Google Scholar] [CrossRef]
- Yang, C.; Cheng, C.; Wang, J.; Chen, K.; Zhan, J.; Pan, X.; Xu, X.; Xu, W.; Liu, S. [Salvianolic acid B and its magnesium salt inhibit SARS-CoV-2 infection of Vero-E6 cells by blocking spike protein-mediated membrane fusion]. Nan Fang Yi Ke Da Xue Xue Bao 2021, 41, 475–482. (In Chinese) [Google Scholar]
- Yang, C.; Pan, X.; Xu, X.; Cheng, C.; Huang, Y.; Li, L.; Jiang, S.; Xu, W.; Xiao, G.; Liu, S. Salvianolic acid C potently inhibits SARS-CoV-2 infection by blocking the formation of six-helix bundle core of spike protein. Signal Transduct. Target Ther. 2020, 5, 220. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, Y.I.; Park, S.J.; Kim, I.K.; Choi, Y.K.; Kim, S.H. Safe, high-throughput screening of natural compounds of MERS-CoV entry inhibitors using a pseudovirus expressing MERS-CoV spike protein. Int. J. Antimicrob. Agents 2018, 52, 730–732. [Google Scholar] [CrossRef]
- Wang, W.; Li, S.S.; Xu, X.F.; Yang, C.; Niu, X.G.; Yin, S.X.; Pan, X.Y.; Xu, W.; Hu, G.D.; Wang, C.; et al. Danshensu alleviates pseudo-typed SARS-CoV-2 induced mouse acute lung inflammation. Acta Pharmacol. Sin. 2022, 43, 771–780. [Google Scholar] [CrossRef]
- Li, M.; Zhang, L.; Cai, R.L.; Gao, Y.; Qi, Y. Lipid-soluble extracts from Salvia miltiorrhiza inhibit production of LPS-induced inflammatory mediators via NF-κB modulation in RAW 264.7 cells and perform antiinflammatory effects in vivo. Phytother. Res. 2012, 26, 1195–1204. [Google Scholar] [CrossRef]
- Zhang, W.; Suo, M.; Yu, G.; Zhang, M. Antinociceptive and anti-inflammatory effects of cryptotanshinone through PI3K/Akt signaling pathway in a rat model of neuropathic pain. Chem. Biol. Interact. 2019, 305, 127–133. [Google Scholar] [CrossRef]
- Yuan, R.; Huang, L.; Du, L.J.; Feng, J.F.; Li, J.; Luo, Y.Y.; Xu, Q.M.; Yang, S.L.; Gao, H.; Feng, Y.L. Dihydrotanshinone exhibits an anti-inflammatory effect in vitro and in vivo through blocking TLR4 dimerization. Pharmacol. Res. 2019, 142, 102–114. [Google Scholar] [CrossRef]
- Chen, Z.; Gao, X.; Jiao, Y.; Qiu, Y.; Wang, A.; Yu, M.; Che, F.; Li, S.; Liu, J.; Li, J.; et al. Tanshinone IIA Exerts Anti-Inflammatory and Immune-Regulating Effects on Vulnerable Atherosclerotic Plaque Partially via the TLR4/MyD88/NF-κB Signal Pathway. Front Pharmacol. 2019, 10, 850. [Google Scholar] [CrossRef] [Green Version]
- Ding, B.; Lin, C.; Liu, Q.; He, Y.; Ruganzu, J.B.; Jin, H.; Peng, X.; Ji, S.; Ma, Y.; Yang, W. Tanshinone IIA attenuates neuroinflammation via inhibiting RAGE/NF-κB signaling pathway in vivo and in vitro. J. Neuroinflamm. 2020, 17, 302. [Google Scholar] [CrossRef]
- Yang, L.; Zhou, G.; Liu, J.; Song, J.; Zhang, Z.; Huang, Q.; Wei, F. Tanshinone I and Tanshinone IIA/B attenuate LPS-induced mastitis via regulating the NF-κB. Biomed. Pharmacother. 2021, 137, 111353. [Google Scholar] [CrossRef]
- Bonnet, B.; Cosme, J.; Dupuis, C.; Coupez, E.; Adda, M.; Calvet, L.; Fabre, L.; Saint-Sardos, P.; Bereiziat, M.; Vidal, M.; et al. Severe COVID-19 is characterized by the co-occurrence of moderate cytokine inflammation and severe monocyte dysregulation. EBioMedicine 2021, 73, 103622. [Google Scholar] [CrossRef]
- Ziegler, C.G.; Allon, S.J.; Nyquist, S.K.; Mbano, I.M.; Miao, V.N.; Tzouanas, C.N.; Cao, Y.; Yousif, A.S.; Bals, J.; Hauser, B.M.; et al. SARS-CoV-2 Receptor ACE2 Is an Interferon-Stimulated Gene in Human Airway Epithelial Cells and Is Detected in Specific Cell Subsets across Tissues. Cell 2020, 181, 1016–1035.e19. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Active Compound | S. miltiorrhiza e Root Powder Extract (mg/g Dry Mater) |
|---|---|
| Danshensu/Salvianic acid | 0.05 |
| Rosmarinic acid | 1.61 |
| Salvianolic acid A | 0.31 |
| Salvianolic acid B | 0.07 |
| Ursolic acid | 1.64 |
| Tanshinone I | 0.90 |
| Tanshinone IIA | 10.80 |
| Tanshinone IIB | 10.22 |
| Tanshinone VI | 5.53 |
| Dihydrotanshinone I | 15.35 |
| Cryptotanshinone | 77.59 |
| Tetrahydrotanshinone | 3.61 |
| Total (%) | 12.8% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petitjean, S.J.L.; Lecocq, M.; Lelong, C.; Denis, R.; Defrère, S.; Mariage, P.-A.; Alsteens, D.; Pilette, C. Salvia miltiorrhiza Bunge as a Potential Natural Compound against COVID-19. Cells 2022, 11, 1311. https://doi.org/10.3390/cells11081311
Petitjean SJL, Lecocq M, Lelong C, Denis R, Defrère S, Mariage P-A, Alsteens D, Pilette C. Salvia miltiorrhiza Bunge as a Potential Natural Compound against COVID-19. Cells. 2022; 11(8):1311. https://doi.org/10.3390/cells11081311
Chicago/Turabian StylePetitjean, Simon J. L., Marylène Lecocq, Camille Lelong, Robin Denis, Sylvie Defrère, Pierre-Antoine Mariage, David Alsteens, and Charles Pilette. 2022. "Salvia miltiorrhiza Bunge as a Potential Natural Compound against COVID-19" Cells 11, no. 8: 1311. https://doi.org/10.3390/cells11081311
APA StylePetitjean, S. J. L., Lecocq, M., Lelong, C., Denis, R., Defrère, S., Mariage, P.-A., Alsteens, D., & Pilette, C. (2022). Salvia miltiorrhiza Bunge as a Potential Natural Compound against COVID-19. Cells, 11(8), 1311. https://doi.org/10.3390/cells11081311