
Exercise Affects the Formation and Recovery of Alcoholic Liver Disease through the IL-6–p47phox Oxidative–Stress Axis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
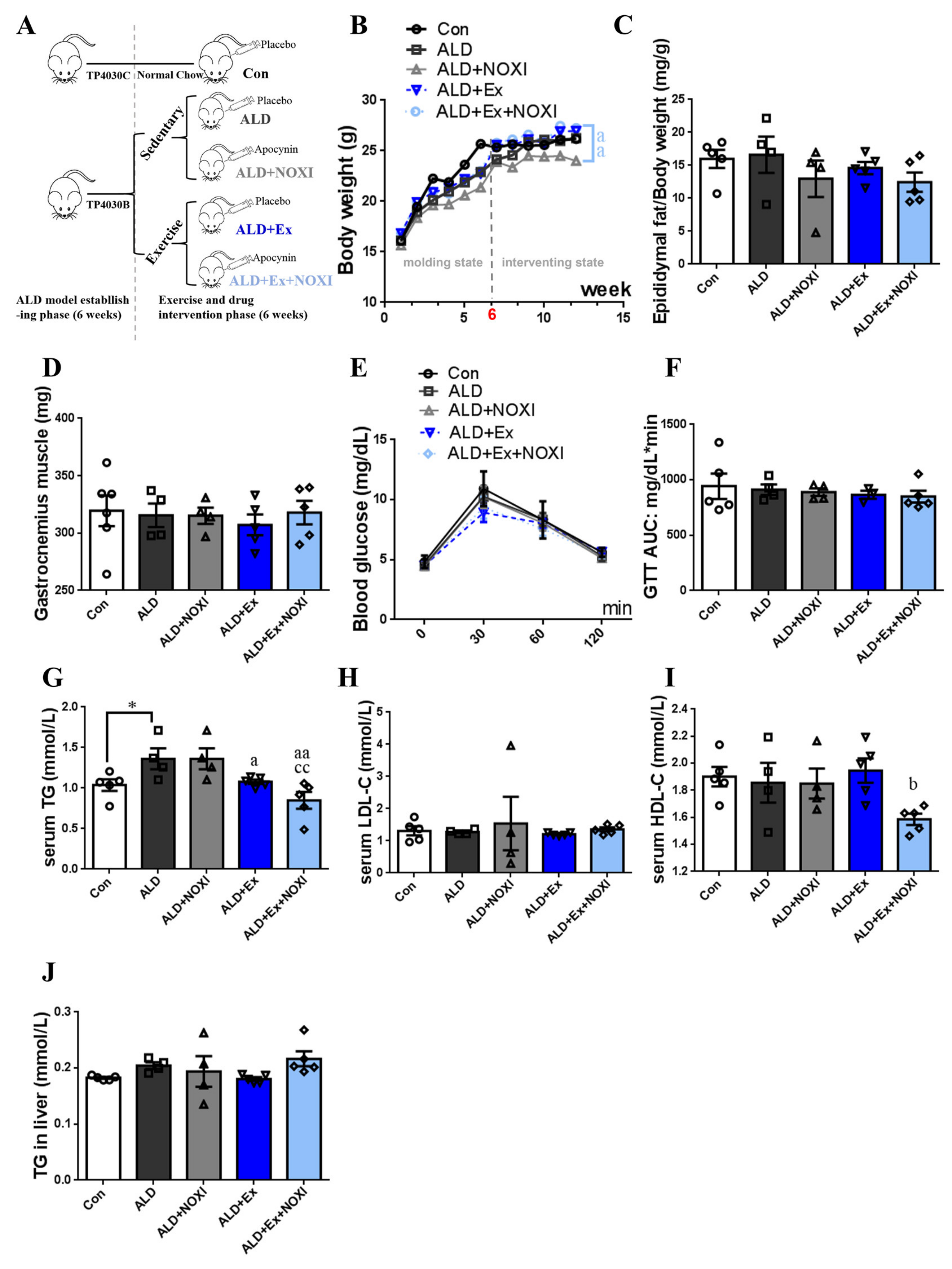
2.1. Animal Grouping and Model Preparation
2.2. Feed and Reagents
2.3. Mice Keeping and Harvesting
2.4. Glucose Tolerance Testing
2.5. Spectrophotometric Density Value Analysis
2.6. HE and Oil Red O Staining
2.7. Western Blotting
2.8. Statistical Analysis
3. Results
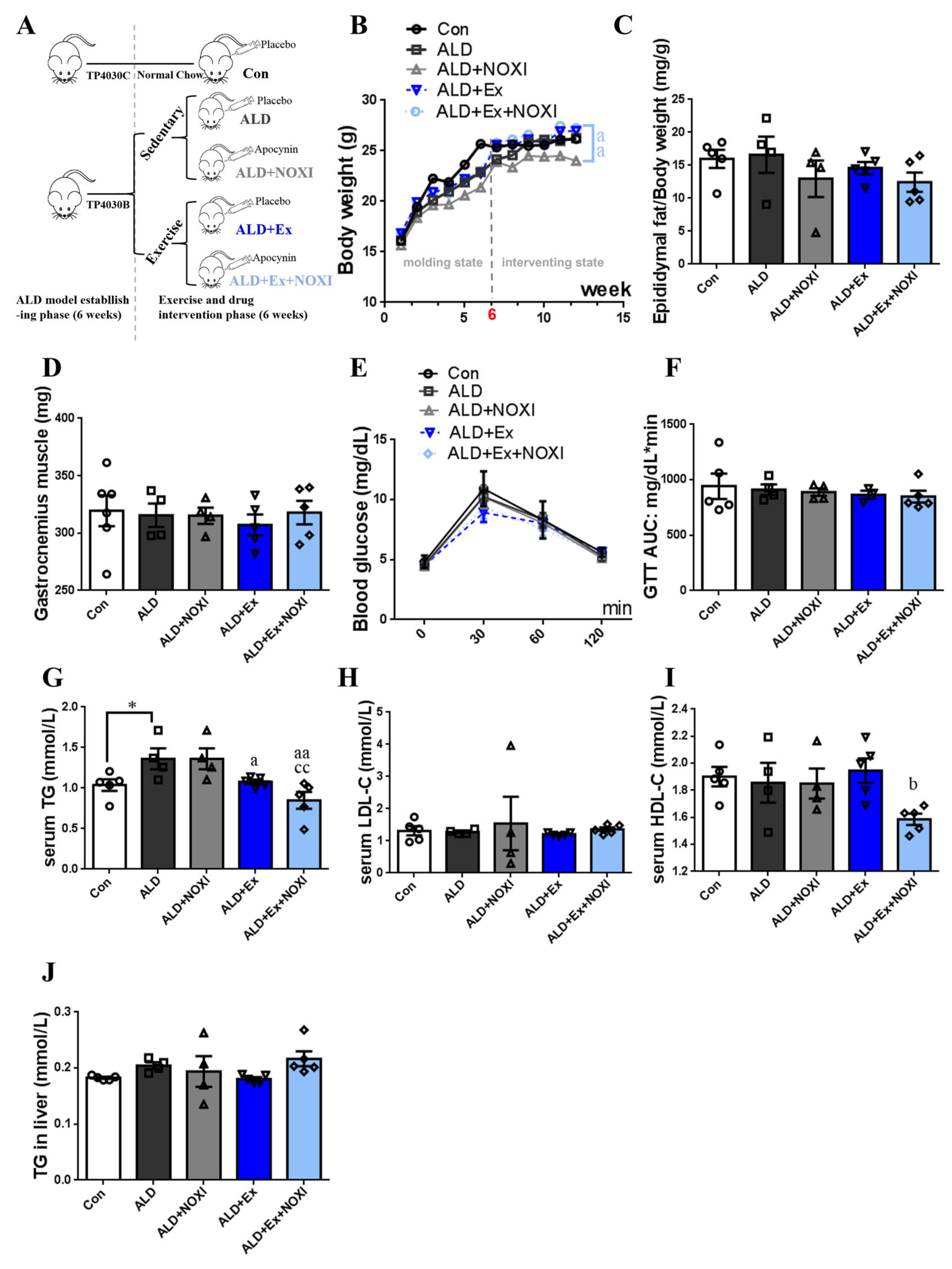
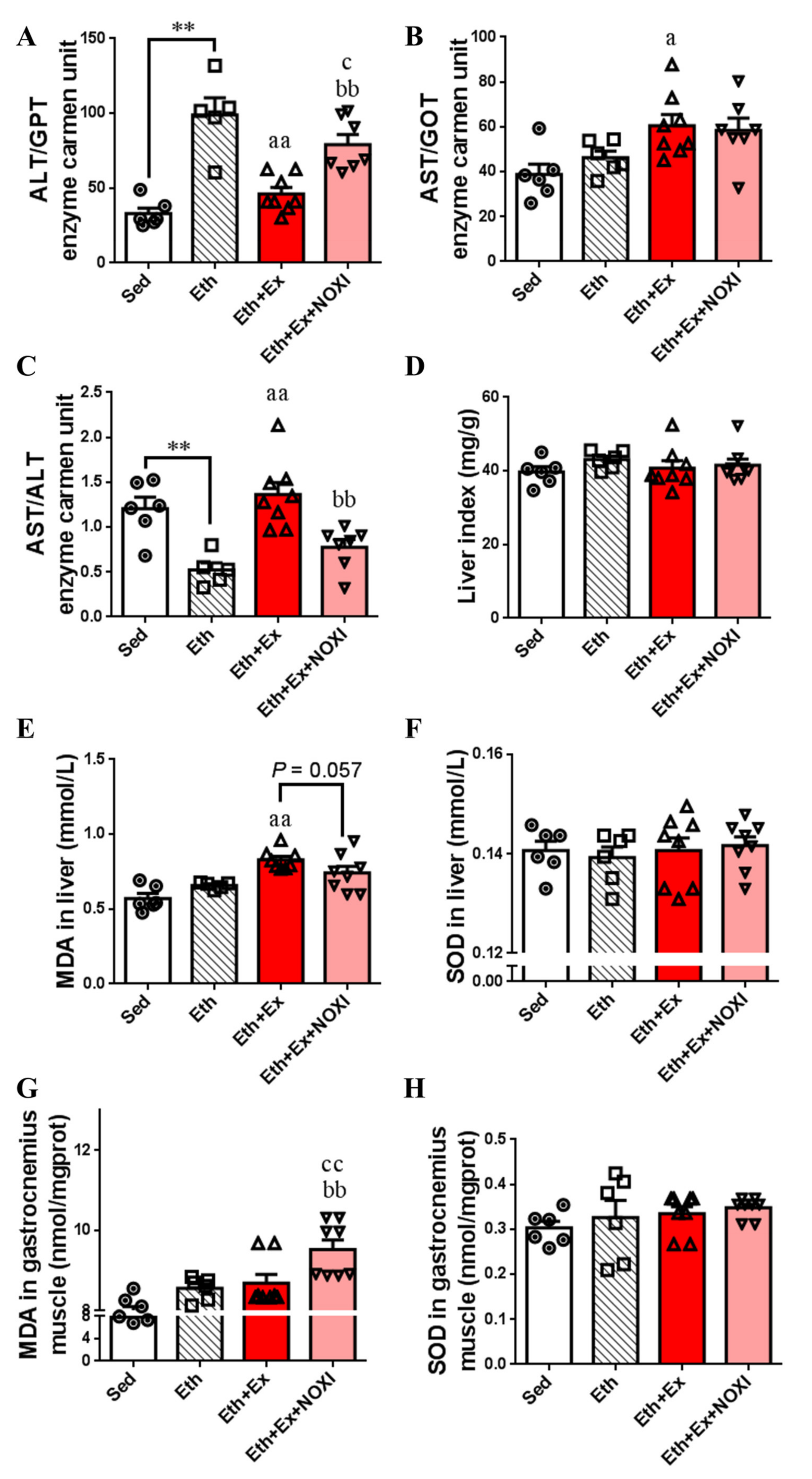
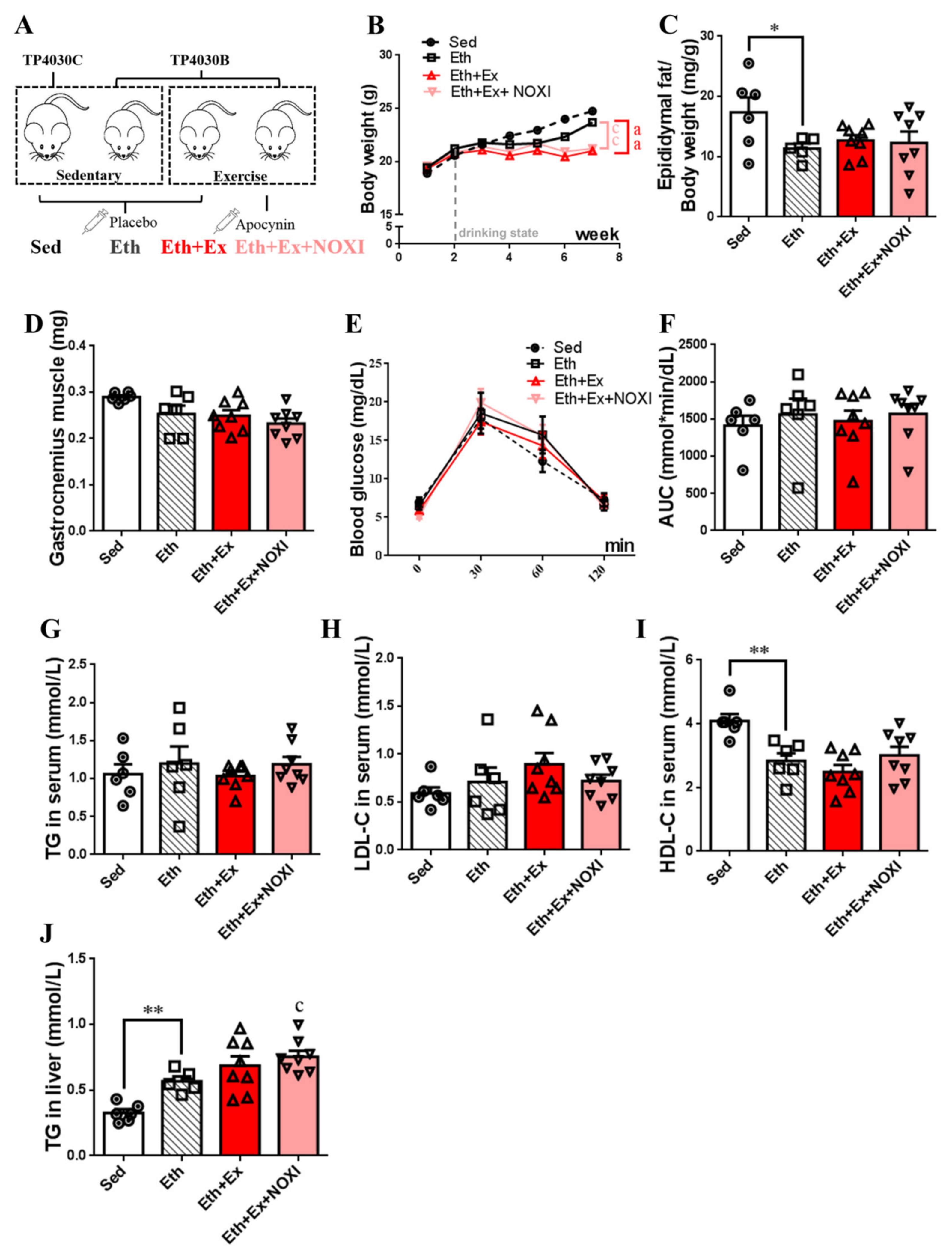
3.1. Exercise Training Ameliorates Dyslipidemia and Inflammation
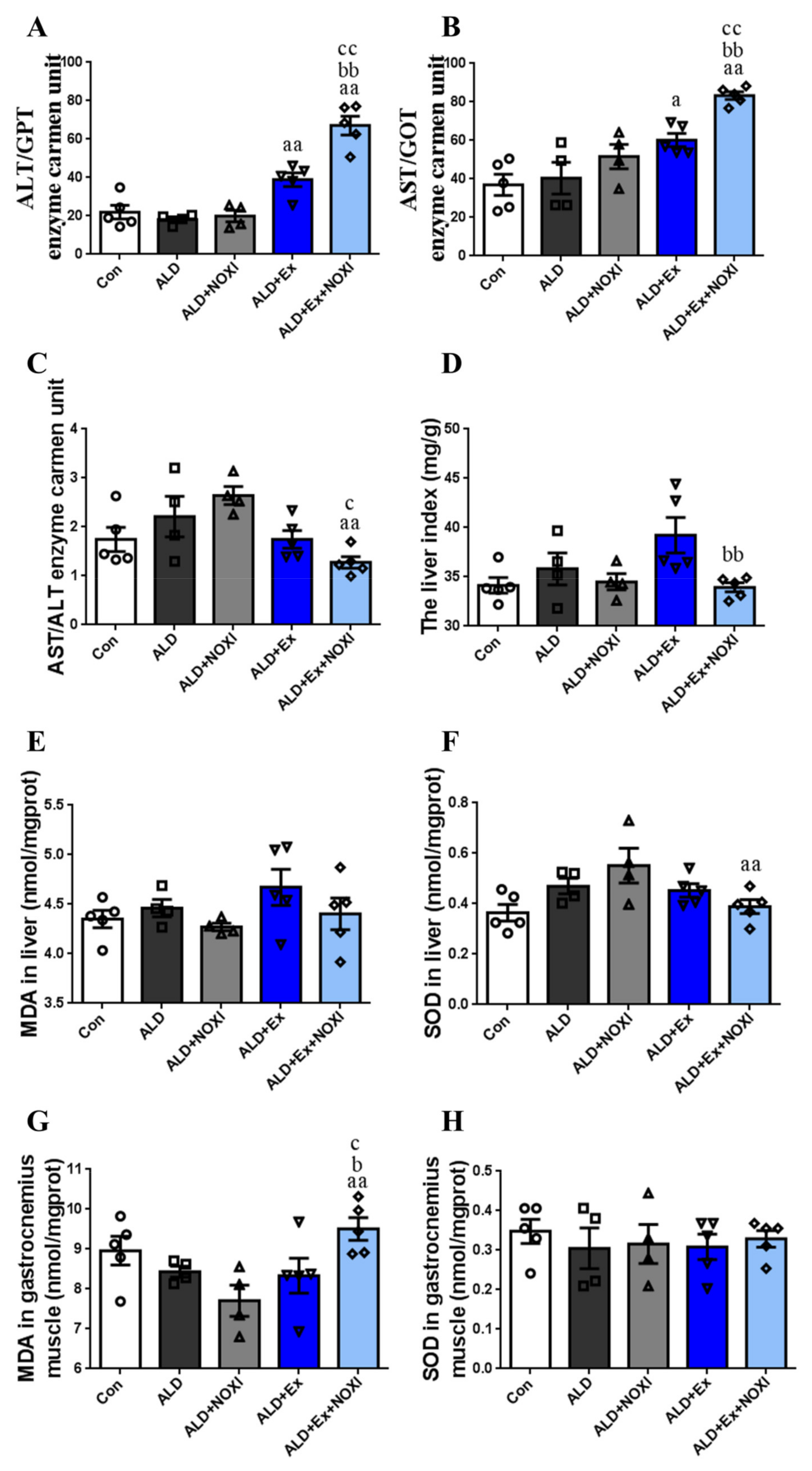
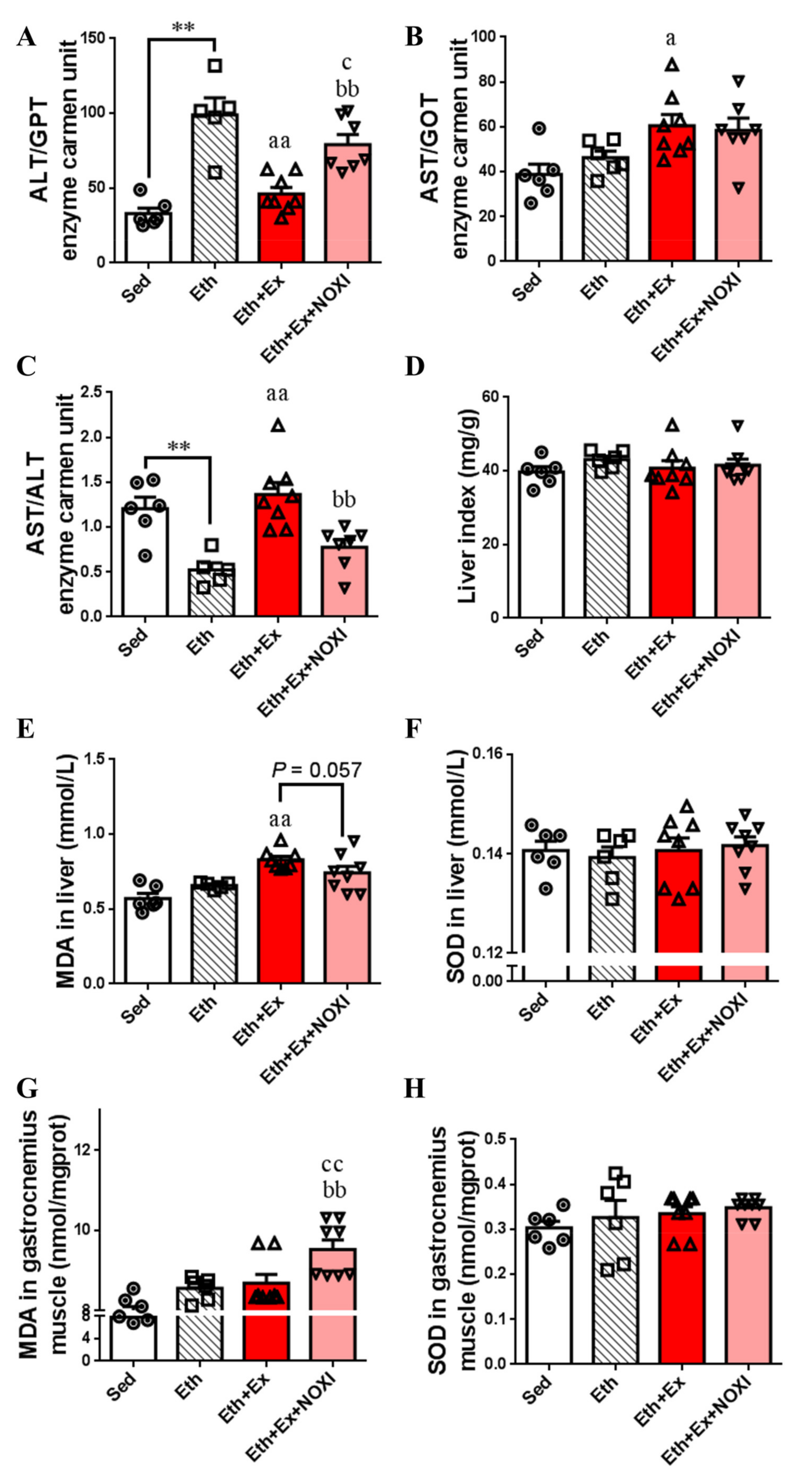
3.2. Exercise Reduces Oxidative Stress Levels in Liver Tissue
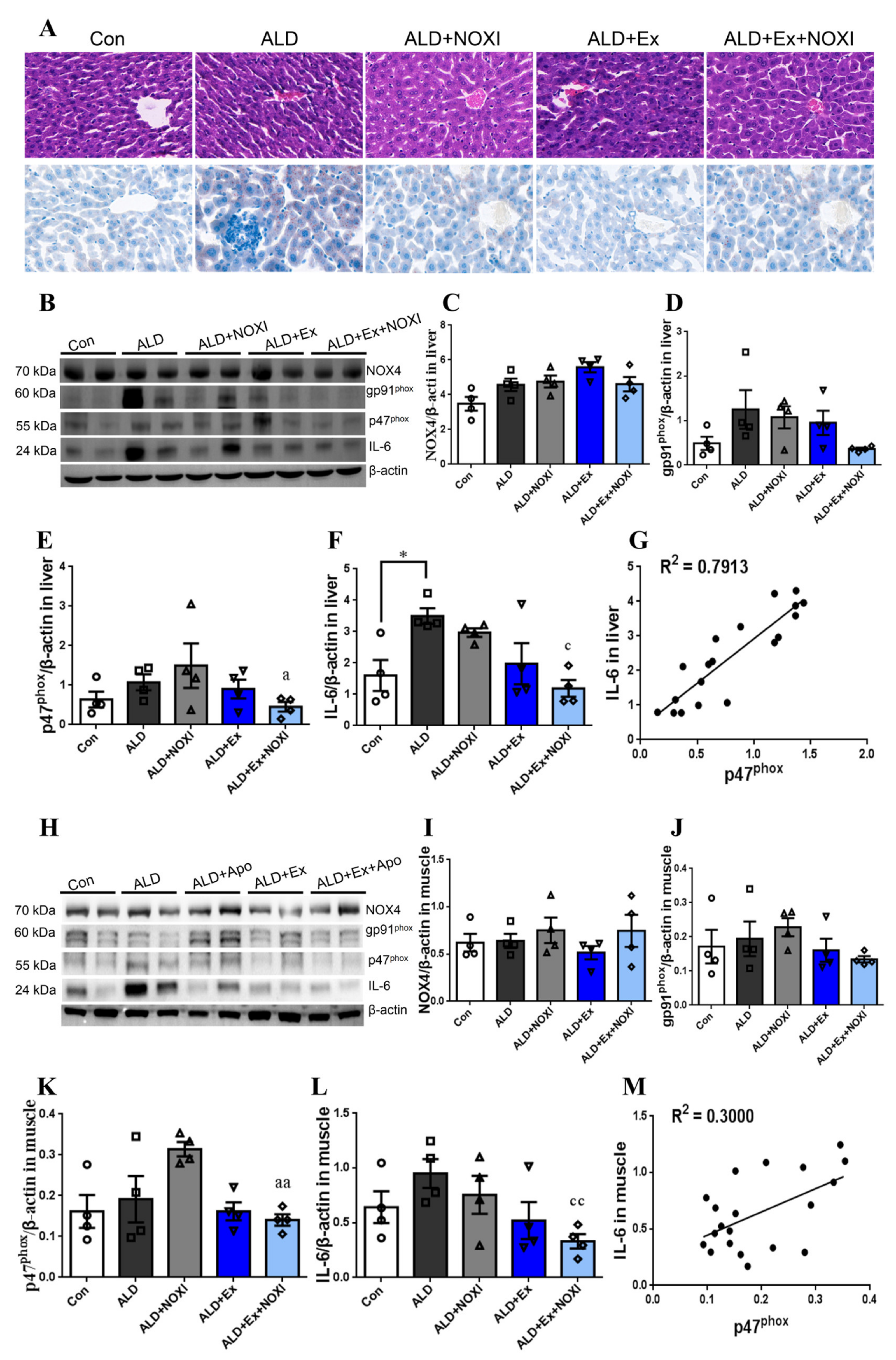
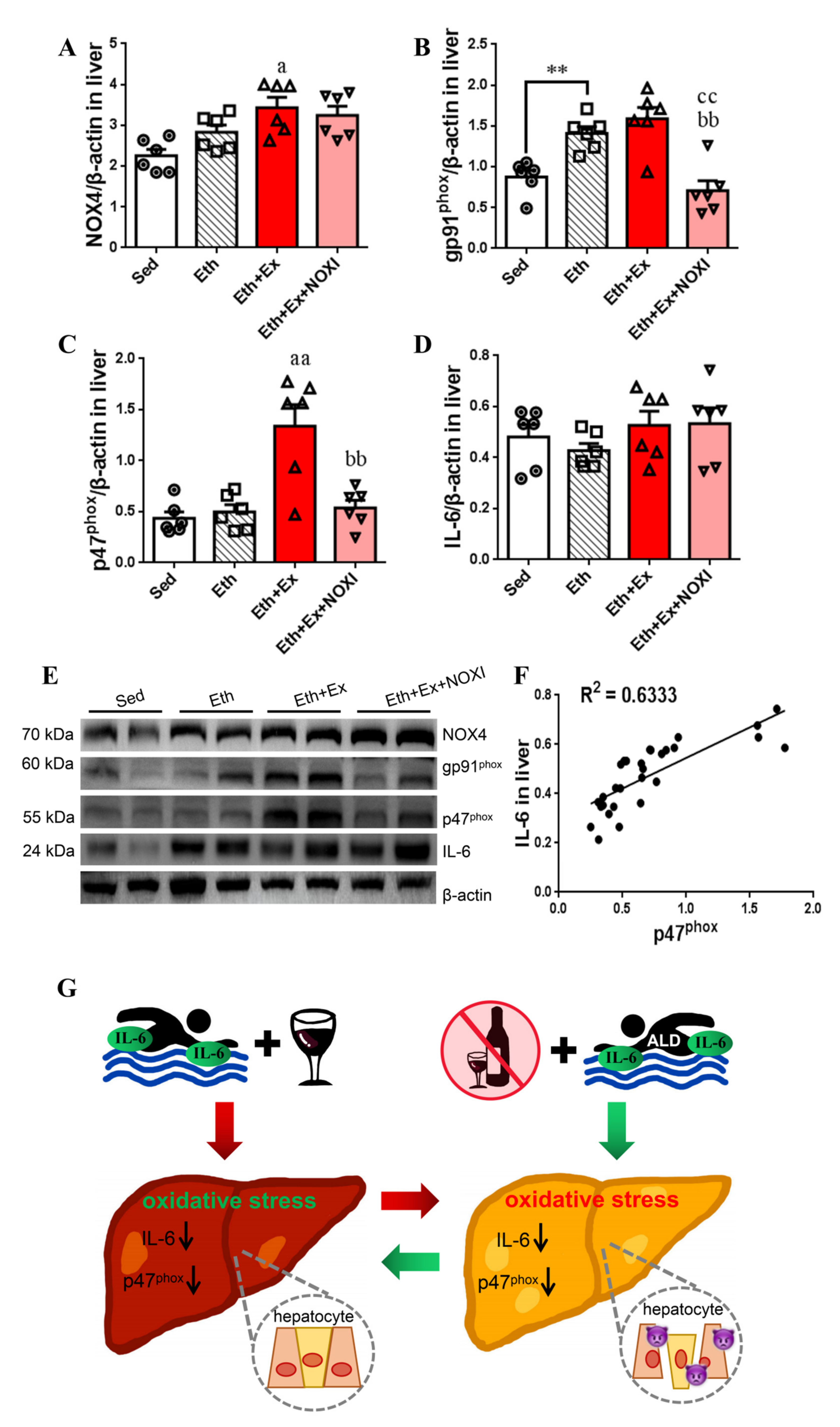
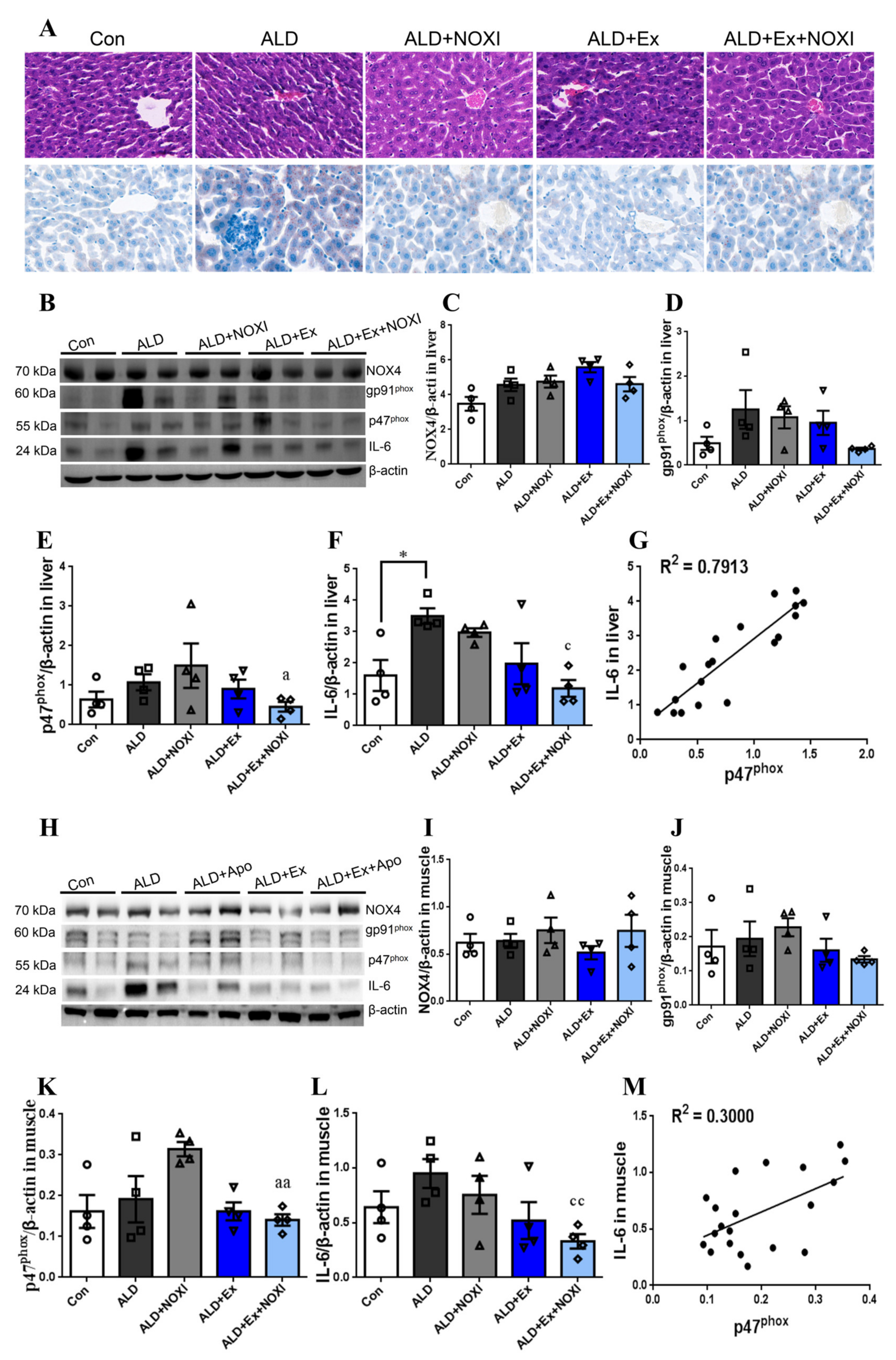
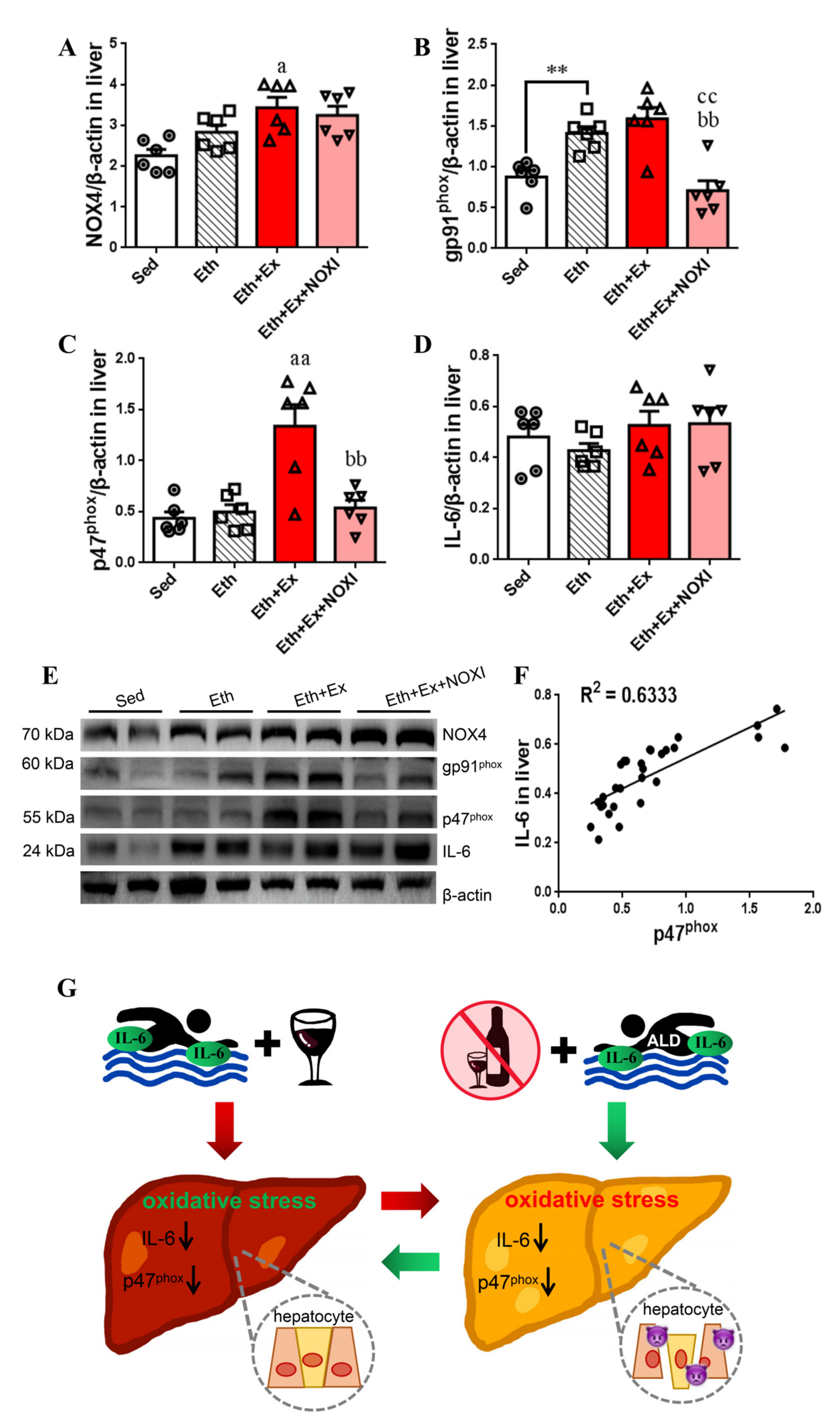
3.3. Exercise Significantly Improved Fat Accumulation and Inflammatory Injury in Liver Tissue, and Its Positive Effect Is Involved with the Liver IL-6–p47phox Axis
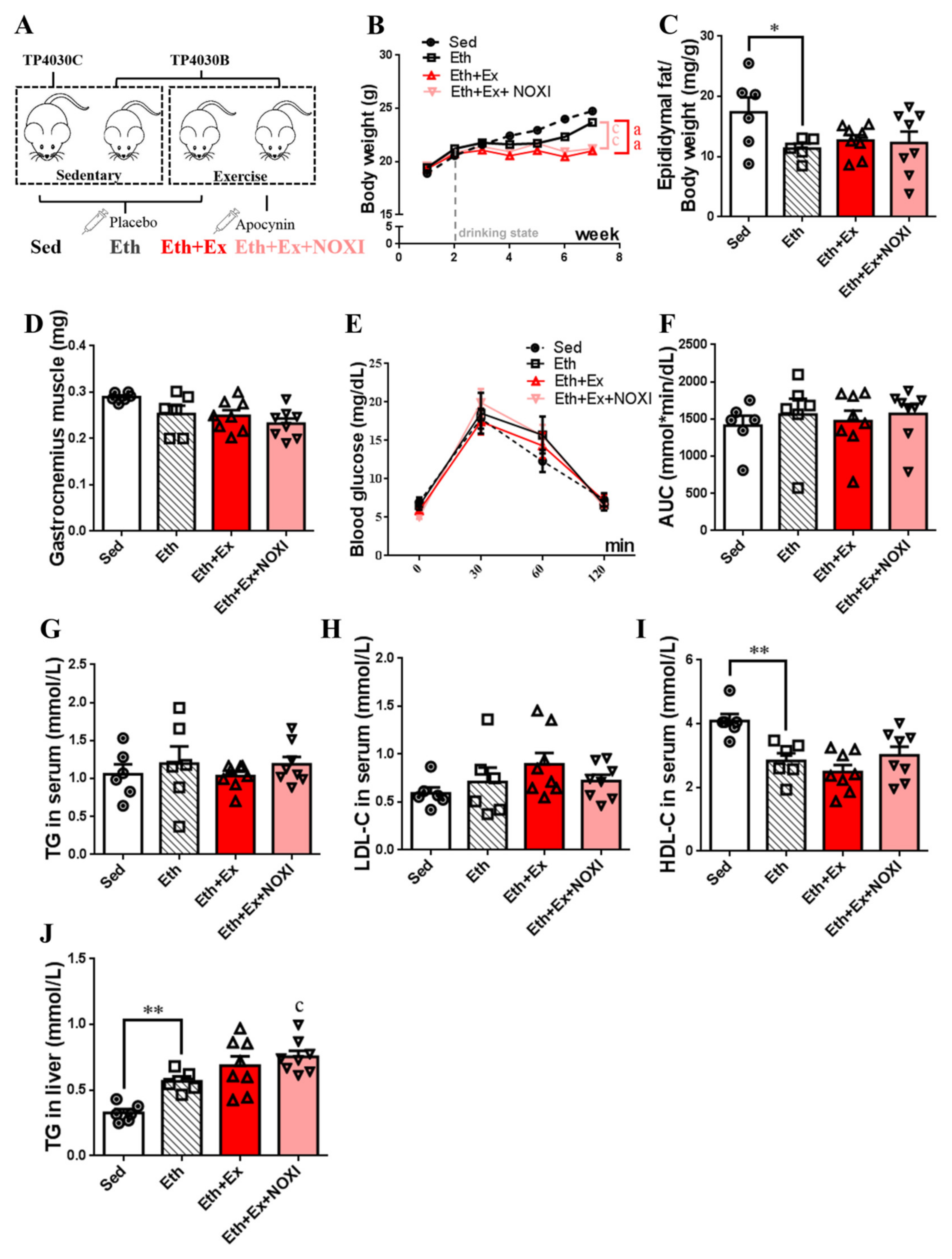
3.4. Drinking Alcohol during Exercise Aggravates Dyslipidemia, Which Had No Significant Effect after Apocynin Intervention
3.5. Drinking Alcohol during Exercise can Increase the Level of Inflammation and the Oxidative Stress in Tissues
3.6. Exercise-Mediated Protein Expression of IL-6 and p47phox in Liver Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cui, Y.; Jiang, L.; Shao, Y.; Mei, L.; Tao, Y. Anti-alcohol liver disease effect of Gentianae macrophyllae extract through MAPK/JNK/p38 pathway. J. Pharm. Pharmacol. 2019, 71, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Luther, J.; Khan, S.; Gala, M.K.; Kedrin, D.; Sridharan, G.; Goodman, R.P.; Garber, J.J.; Masia, R.; Diagacomo, E.; Adams, D.; et al. Hepatic gap junctions amplify alcohol liver injury by propagating cGAS-mediated IRF3 activation. Proc. Natl. Acad. Sci. USA 2020, 117, 11667–11673. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhang, N.; Zhao, L.; Wu, W.; Zhang, L.; Zhou, F.; Li, J. Astragalus Polysaccharides and Saponins Alleviate Liver Injury and Regulate Gut Microbiota in Alcohol Liver Disease Mice. Foods 2021, 10, 2688. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Gu, J.; Liu, R.; Wei, S.; Wang, Q.; Shen, H.; Dai, Y.; Zhou, H.; Zhang, F.; Lu, L. Spermine Alleviates Acute Liver Injury by Inhibiting Liver-Resident Macrophage Pro-Inflammatory Response Through ATG5-Dependent Autophagy. Front. Immunol. 2018, 9, 948. [Google Scholar] [CrossRef]
- Wu, J.; Wu, D.; Ma, K.; Wang, T.; Shi, G.; Shao, J.; Wang, C.; Yan, G. Paeonol ameliorates murine alcohol liver disease via mycobiota-mediated Dectin-1/IL-1beta signaling pathway. J. Leukoc. Biol. 2020, 108, 199–214. [Google Scholar] [CrossRef]
- Rishi, P.; Arora, S.; Kaur, U.J.; Chopra, K.; Kaur, I.P. Better Management of Alcohol Liver Disease Using a ‘Microstructured Synbox’ System Comprising L. plantarum and EGCG. PLoS ONE 2017, 12, e0168459. [Google Scholar] [CrossRef]
- Tang, X.H.; Melis, M.; Mai, K.; Gudas, L.J.; Trasino, S.E. Fenretinide Improves Intestinal Barrier Function and Mitigates Alcohol Liver Disease. Front. Pharmacol. 2021, 12, 630557. [Google Scholar] [CrossRef]
- Cichoz-Lach, H.; Partycka, J.; Nesina, I.; Celinski, K.; Slomka, M.; Wojcierowski, J. Genetic polymorphism of alcohol dehydrogenase 3 in alcohol liver cirrhosis and in alcohol chronic pancreatitis. Alcohol Alcohol. 2006, 41, 14–17. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Zhang, Y.; Shi, H.; Feng, Y.; Zhang, Z.; Zhang, L. Proteomic profiling of hepatic stellate cells in alcohol liver fibrosis reveals proteins involved in collagen production. Alcohol 2020, 86, 81–91. [Google Scholar] [CrossRef]
- Schmidt-Arras, D.; Rose-John, S. IL-6 pathway in the liver: From physiopathology to therapy. J. Hepatol. 2016, 64, 1403–1415. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Gu, J.; Liu, R.; Zhou, H.; Lu, L.; Dai, X.; Qian, X. IL-2/IL-6 ratio correlates with liver function and recovery in acute liver injury patients. APMIS 2019, 127, 468–474. [Google Scholar] [CrossRef]
- Tao, Y.; Wang, M.; Chen, E.; Tang, H. Liver Regeneration: Analysis of the Main Relevant Signaling Molecules. Mediat. Inflamm. 2017, 2017, 4256352. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; He, Y.; Zhou, Z.; Ramirez, T.; Gao, Y.; Gao, Y.; Ross, R.A.; Cao, H.; Cai, Y.; Xu, M.; et al. MicroRNA-223 ameliorates alcoholic liver injury by inhibiting the IL-6–p47phox–oxidative stress pathway in neutrophils. Gut 2017, 66, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Hong, M.; Tan, H.-Y.; Wang, N.; Feng, Y. Insights into the Role and Interdependence of Oxidative Stress and Inflammation in Liver Diseases. Oxidative Med. Cell. Longev. 2016, 2016, 4234061. [Google Scholar] [CrossRef]
- Zhou, T.; Zhang, Y.J.; Xu, D.P.; Wang, F.; Zhou, Y.; Zheng, J.; Li, Y.; Zhang, J.J.; Li, H.B. Protective Effects of Lemon Juice on Alcohol-Induced Liver Injury in Mice. Biomed. Res. Int. 2017, 2017, 7463571. [Google Scholar] [CrossRef]
- Cui, W.; Cui, D.; Ouyang, T.; Li, X.; Wei, H.T.; Xue, W.Y.; Zhou, G.; Qiu, Y. NOX inhibition alleviates hepatocyte injury and lipid metabolism disorder in alcoholic liver model mice [J]. Chin. J. Tissue Eng. Res. 2022, 3, 1–7. [Google Scholar]
- Zheng, Y.; Ley, S.H.; Hu, F.B. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat. Rev. Endocrinol. 2018, 14, 88–98. [Google Scholar] [CrossRef]
- Petersen, M.C.; Vatner, D.F.; Shulman, G.I. Regulation of hepatic glucose metabolism in health and disease. Nat. Rev. Endocrinol. 2017, 13, 572–587. [Google Scholar] [CrossRef] [Green Version]
- Younossi, Z.M.; Golabi, P.; de Avila, L.; Paik, J.M.; Srishord, M.; Fukui, N.; Qiu, Y.; Burns, L.; Afendy, A.; Nader, F. The global epidemiology of NAFLD and NASH in patients with type 2 diabetes: A systematic review and meta-analysis. J. Hepatol. 2019, 71, 793–801. [Google Scholar] [CrossRef]
- Stavropoulos, K.; Imprialos, K.; Pittaras, A.; Faselis, C.; Narayan, P.; Kokkinos, P. Lifestyle Modifications in Non-Alcoholic Fatty Liver Disease and Non- Alcoholic Steatohepatitis. Curr. Vasc. Pharmacol. 2018, 16, 239–245. [Google Scholar] [CrossRef]
- Anagnostopoulou, A.; Camargo, L.L.; Rodrigues, D.; Montezano, A.C.; Touyz, R.M. Importance of cholesterol-rich microdomains in the regulation of Nox isoforms and redox signaling in human vascular smooth muscle cells. Sci. Rep. 2020, 10, 17818. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.J.; Min, H.S.; Kim, K.T.; Kim, J.E.; Ghee, J.Y.; Kim, H.W.; Lee, J.E.; Han, J.-Y.; Lee, G.; Ha, H.; et al. APX-115, a first-in-class pan-NADPH oxidase (Nox) inhibitor, protects db/db mice from renal injury. Lab. Investig. 2017, 97, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Zhao, J.F.; Hsu, C.P.; Kou, Y.R.; Lu, T.M.; Lee, T.S. The detrimental effect of asymmetric dimethylarginine on cholesterol efflux of macrophage foam cells: Role of the NOX/ROS signaling. Free Radic. Biol. Med. 2019, 143, 354–365. [Google Scholar] [CrossRef] [PubMed]
- Safdar, A.; Saleem, A.; Tarnopolsky, M.A. The potential of endurance exercise-derived exosomes to treat metabolic diseases. Nat. Rev. Endocrinol. 2016, 12, 504–517. [Google Scholar] [CrossRef]
- Thompson, K.J.; Nazari, S.S.; Jacobs, W.C.; Grahame, N.J.; McKillop, I.H. Use of a crossed high alcohol preferring (cHAP) mouse model with the NIAAA-model of chronic-binge ethanol intake to study liver injury. Alcohol Alcohol. 2017, 52, 629–637. [Google Scholar] [CrossRef] [Green Version]
- Bertola, A.; Mathews, S.; Ki, S.H.; Wang, H.; Gao, B. Mouse model of chronic and binge ethanol feeding (the NIAAA model). Nat. Protoc. 2013, 8, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Perry, R.J.; Samuel, V.T.; Petersen, K.F.; Shulman, G.I. The role of hepatic lipids in hepatic insulin resistance and type 2 diabetes. Nature 2014, 7503, 84–91. [Google Scholar] [CrossRef]
- Rajput, R.; Ahlawat, P. Prevalence and predictors of non-alcoholic fatty liver disease in prediabetes. Diabetes Metab. Syndr. 2019, 13, 2957–2960. [Google Scholar] [CrossRef]
- Draijer, L.G.; Feddouli, S.; Bohte, A.E.; Slootweg, O.V.B.; Rijcken, T.H.P.; Benninga, M.A.; Stoker, J.; Koot, B.G.P. Comparison of diagnostic accuracy of screening tests ALT and ultrasound for pediatric non-alcoholic fatty liver disease. Eur. J. Pediatr. 2019, 178, 863–870. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Wang, Q.; Yu, L.; Yin, X.; Yang, H.; Xu, X.; Xia, Y.; Luo, Y.; Peng, Y.; Yu, Q.; et al. Revision of serum ALT upper limits of normal facilitates assessment of mild liver injury in obese children with non-alcoholic fatty liver disease. J. Clin. Lab. Anal. 2020, 34, e23285. [Google Scholar] [CrossRef]
- Abhilash, K.; Gn, A. High Ast/Alt Is the Index of Advanced Alcoholic Liver Disease Rather than Heavy Alcohol Drinking: A Study in Tertiary Care Hospital. J. Assoc. Physicians India 2020, 68, 98. [Google Scholar]
- Kresnamurti, A.; Rakhma, D.N.; Damayanti, A.; Santoso, S.D.; Restryarto, E.; Hadinata, W.; Hamid, I.S. AST/ALT levels, MDA, and liver histopathology of Echinometra mathaei ethanol extract on paracetamol-induced hepatotoxicity in rats. J. Basic Clin. Physiol. Pharmacol. 2021, 32, 511–516. [Google Scholar] [CrossRef]
- Knudsen, J.G.; Bertholdt, L.; Gudiksen, A.; Gerbal-Chaloin, S.; Rasmussen, M.K. Skeletal Muscle Interleukin-6 Regulates Hepatic Cytochrome P450 Expression: Effects of 16-Week High-Fat Diet and Exercise. Toxicol. Sci. 2018, 162, 309–317. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Huang, Y.S.; Wang, L.Y.; Chang, C.H.; Perng, C.L.; Lin, H.C. Superoxide Dismutase 2 Genetic Variation as a Susceptibility Risk Factor for Alcoholic Cirrhosis. Alcohol Alcohol. 2016, 51, 633–637. [Google Scholar] [CrossRef]
- Barreto, T.O.; Cleto, L.S.; Gioda, C.R.; Silva, R.S.; Campi-Azevedo, A.C.; de Sousa-Franco, J.; de Magalhaes, J.C.; Penaforte, C.L.; Pinto, K.M.; Cruz Jdos, S.; et al. Swim training does not protect mice from skeletal muscle oxidative damage following a maximum exercise test. Eur. J. Appl. Physiol. 2012, 112, 2523–2530. [Google Scholar] [CrossRef]
- Nakanishi, T.; Tsujii, M.; Asano, T.; Iino, T.; Sudo, A. Protective Effect of Edaravone Against Oxidative Stress in C2C12 Myoblast and Impairment of Skeletal Muscle Regeneration Exposed to Ischemic Injury in Ob/ob Mice. Front. Physiol. 2019, 10, 1596. [Google Scholar] [CrossRef]
- De Castro, M.R.T.; Ferreira, A.P.O.; Busanello, G.L.; Da Silva, L.R.H.; Junior, M.E.P.D.S.; Fiorin, F.D.S.; Arrifano, G.; Crespo-Lopez, M.E.; Barcelos, R.P.; Cuevas, M.J.; et al. Previous physical exercise alters the hepatic profile of oxidative-inflammatory status and limits the secondary brain damage induced by severe traumatic brain injury in rats. J. Physiol. 2017, 595, 6023–6044. [Google Scholar] [CrossRef]
- Shen, F.; Wang, Z.; Liu, W.; Liang, Y. Ethyl pyruvate can alleviate alcoholic liver disease through inhibiting Nrf2 signaling pathway. Exp. Ther. Med. 2018, 15, 4223–4228. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, Y.; Dehnad, A.; Fish, S.; Sato, A.; Jiang, J.; Tian, J.; Schröder, K.; Brandes, R.; Török, N.J. NOX4 Regulates CCR2 and CCL2 mRNA Stability in Alcoholic Liver Disease. Sci. Rep. 2017, 7, srep46144. [Google Scholar] [CrossRef]
- Paik, Y.H.; Kim, J.; Aoyama, T.; De Minicis, S.; Bataller, R.; Brenner, D.A. Role of NADPH Oxidases in Liver Fibrosis. Antioxid. Redox Signal. 2014, 20, 2854–2872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, S.; Kisseleva, T.; Brenner, D.A. The Role of NADPH Oxidases (NOXs) in Liver Fibrosis and the Activation of Myofibroblasts. Front. Physiol. 2016, 7, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, W.; Cui, D. The role of interleukin-6 in alcoholic liver disease. Chn. J. Bio. Mol. Bio. 2021, 37, 467–474. [Google Scholar]
- Knudsen, J.G.; Joensen, E.; Bertholdt, L.; Jessen, H.; van Hauen, L.; Hidalgo, J.; Pilegaard, H. Skeletal muscle IL-6 and regulation of liver metabolism during high-fat diet and exercise training. Physiol. Rep. 2016, 4, e12788. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhou, G.; Mei, Y.; Xie, W.-J.; Li, P.-F.; Yang, F. Mechanism of oxidative stress in skeletal muscle of rats induced by one-off exhaustive exercise. Chin. J. Appl. Physiol. 2020, 36, 17–22. [Google Scholar]
- Li, P.F.; Zhou, G. Il-6 in exercise-induced skeletal muscle inflammation. J. Cult. Goods Technol. 2020, 3, 157–159. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, W.; Li, X.; Xue, W.; Wei, H.; Zhou, G.; Qiu, Y.; Cui, D. Exercise Affects the Formation and Recovery of Alcoholic Liver Disease through the IL-6–p47phox Oxidative–Stress Axis. Cells 2022, 11, 1305. https://doi.org/10.3390/cells11081305
Cui W, Li X, Xue W, Wei H, Zhou G, Qiu Y, Cui D. Exercise Affects the Formation and Recovery of Alcoholic Liver Disease through the IL-6–p47phox Oxidative–Stress Axis. Cells. 2022; 11(8):1305. https://doi.org/10.3390/cells11081305
Chicago/Turabian StyleCui, Wei, Xiang Li, Weiyue Xue, Huiting Wei, Gang Zhou, Ye Qiu, and Di Cui. 2022. "Exercise Affects the Formation and Recovery of Alcoholic Liver Disease through the IL-6–p47phox Oxidative–Stress Axis" Cells 11, no. 8: 1305. https://doi.org/10.3390/cells11081305
APA StyleCui, W., Li, X., Xue, W., Wei, H., Zhou, G., Qiu, Y., & Cui, D. (2022). Exercise Affects the Formation and Recovery of Alcoholic Liver Disease through the IL-6–p47phox Oxidative–Stress Axis. Cells, 11(8), 1305. https://doi.org/10.3390/cells11081305