
Mucosal Immune Responses to Respiratory Syncytial Virus


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Background
2. Methods
3. Results
3.1. Studies Included in the Review
3.2. The Response of the Respiratory Mucosa to RSV Infection
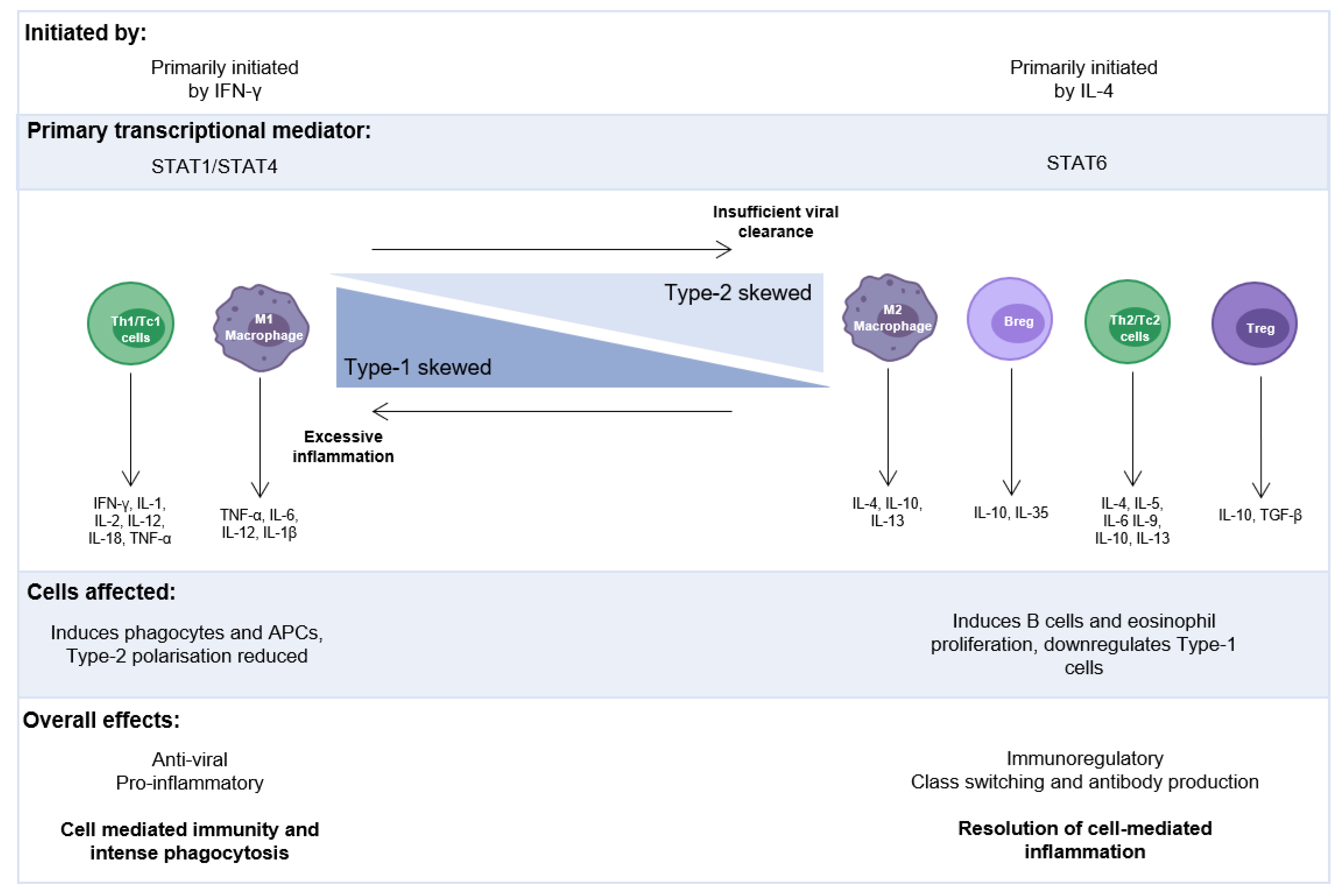
3.2.1. Evidence for Immunoregulation during RSV Infection
Alveolar Macrophages (AMφ)
Regulatory T Cells (Tregs)
Regulatory B Cells (Bregs)
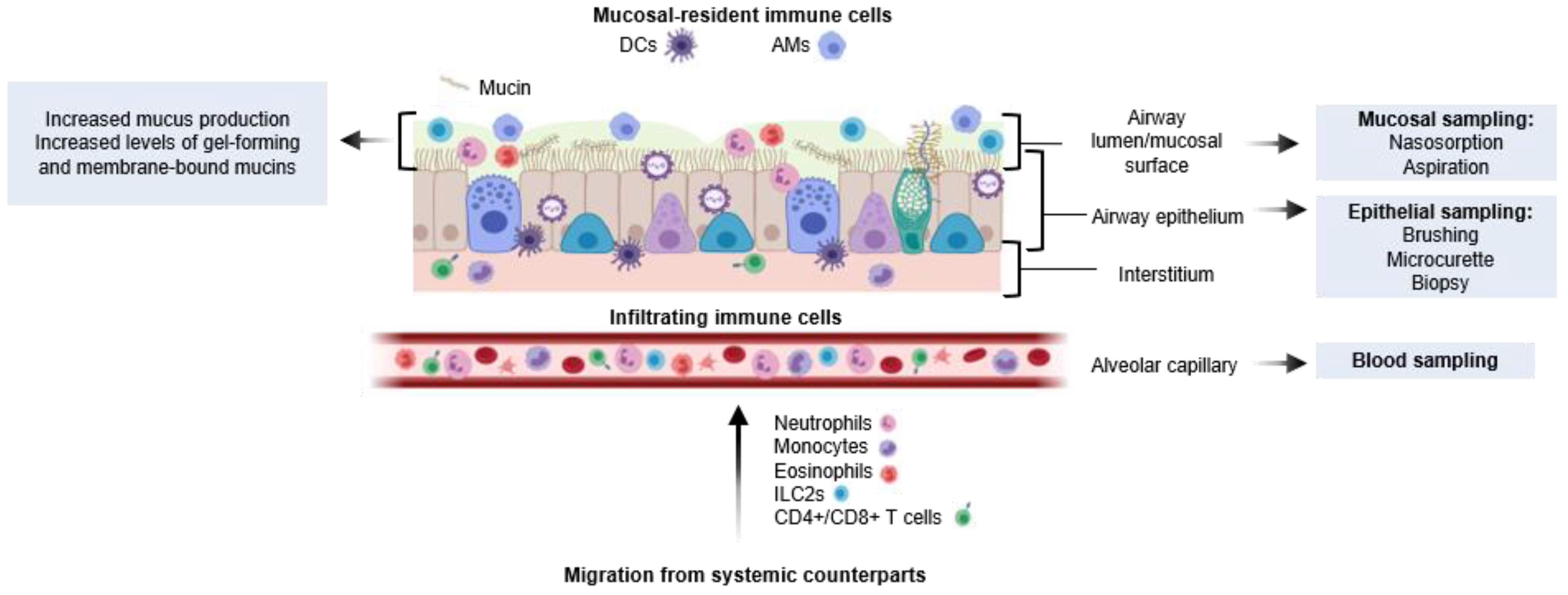
3.2.2. Resident and Infiltrating Immune Cells to the Respiratory Mucosa during Infection
Immune Cells Measured in Respiratory Samples of RSV-Infected Infants
3.2.3. Immune Cell Stimulation as a Measure of Functionality
4. Cytokine Production in Response to Early-Life RSV Infection
4.1. Upper Respiratory Sampling Using NPAs and Nasosorption
4.2. Differences in Cytokine Profiles in the Upper and Lower Airways
4.3. The Anti-Inflammatory Function of IL-10
4.4. Differential Immune Responses to RSV Compared to Other Respiratory Viruses
5. Epithelial Cells Mediating the Immune Response to RSV
Evidence for the Immunogenicity of Mucins
6. Genetic and Environmental Imprinting on Mucosal Immunity
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Collins, P.L.; Fearns, R.; Graham, B.S. Respiratory Syncytial Virus: Virology, Reverse Genetics, and Pathogenesis of Disease. Curr. Top. Microbiol. Immunol. 2013, 372, 3–38. [Google Scholar] [PubMed]
- Shi, T.; McAllister, D.A.; O’Brien, K.L.; Simoes, E.A.F.; Madhi, S.A.; Gessner, B.D.; Polack, F.P.; Balsells, E.; Acacio, S.; Aguayo, C.; et al. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in young children in 2015: A systematic review and modelling study. Lancet 2017, 390, 946–958. [Google Scholar] [CrossRef]
- Coultas, J.A.; Smyth, R.; Openshaw, P.J. Respiratory syncytial virus (RSV): A scourge from infancy to old age. Thorax 2019, 74, 986–993. [Google Scholar] [CrossRef] [PubMed]
- Fleming, D.M.; Taylor, R.J.; Lustig, R.L.; Schuck-Paim, C.; Haguinet, F.; Webb, D.J.; Logie, J.; Matias, G.; Taylor, S. Modelling estimates of the burden of Respiratory Syncytial virus infection in adults and the elderly in the United Kingdom. BMC Infect. Dis. 2015, 15, 443. [Google Scholar] [CrossRef]
- Mazur, N.I.; Higgins, D.; Nunes, M.C.; Melero, J.A.; Langedijk, A.C.; Horsley, N.; Buchholz, U.J.; Openshaw, P.J.; McLellan, J.S.; Englund, J.A.; et al. The respiratory syncytial virus vaccine landscape: Lessons from the graveyard and promising candidates. Lancet Infect. Dis. 2018, 18, e295–e311. [Google Scholar] [CrossRef]
- Melero, J.A.; Moore, M.L. Influence of Respiratory Syncytial Virus Strain Differences on Pathogenesis and Immunity. Curr. Top. Microbiol. Immunol. 2013, 372, 59–82. [Google Scholar]
- O’Brien, K.L.; Baggett, H.C.; Brooks, W.A.; Feikin, D.R.; Hammitt, L.L.; Higdon, M.M.; Howie, S.R.C.; Deloria Knoll, M.; Kotloff, K.L.; Levine, O.S.; et al. Causes of severe pneumonia requiring hospital admission in children without HIV infection from Africa and Asia: The PERCH multi-country case-control study. Lancet 2019, 394, 757–779. [Google Scholar] [CrossRef]
- Parikh, R.C.; McLaurin, K.K.; Margulis, A.V.; Mauskopf, J.; Ambrose, C.S.; Pavilack, M.; Candrilli, S.D. Chronologic Age at Hospitalization for Respiratory Syncytial Virus Among Preterm and Term Infants in the United States. Infect. Dis. Ther. 2017, 6, 477–486. [Google Scholar] [CrossRef]
- Berger, T.M.; Aebi, C.; Duppenthaler, A.; Stocker, M. Prospective population-based study of rsv-related intermediate care and intensive care unit admissions in Switzerland over a 4-year period (2001–2005). Infection 2009, 37, 109–116. [Google Scholar] [CrossRef]
- Resch, B.; Paes, B. Are late preterm infants as susceptible to RSV infection as full term infants? Early Hum. Dev. 2011, 87 (Suppl. 1), S47–S49. [Google Scholar] [CrossRef]
- Resch, B.; Resch, B.; Gusenleitner, W.; Müller, W. The Impact of Respiratory Syncytial Virus Infection: A Prospective Study in Hospitalized Infants Younger than 2 Years. Infection 2002, 30, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Torow, N.; Marsland, B.J.; Hornef, M.W.; Gollwitzer, E.S. Neonatal mucosal immunology. Mucosal Immunol. 2017, 10, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Suleiman-Martos, N.; Caballero-Vázquez, A.; Gómez-Urquiza, J.L.; Albendín-García, L.; Romero-Béjar, J.L.; Cañadas-De la Fuente, G.A. Prevalence and Risk Factors of Respiratory Syncytial Virus in Children under 5 Years of Age in the WHO European Region: A Systematic Review and Meta-Analysis. J. Pers. Med. 2021, 11, 416. [Google Scholar] [CrossRef] [PubMed]
- Atay, Ö.; Pekcan, S.; Göktürk, B.; Özdemir, M. Risk Factors and Clinical Determinants in Bronchiolitis of Infancy. Turkish Thorac. J. 2020, 21, 156–162. [Google Scholar]
- Esposito, S.; Scarselli, E.; Lelii, M.; Scala, A.; Vitelli, A.; Capone, S.; Fornili, M.; Biganzoli, E.; Orenti, A.; Nicosia, A.; et al. Antibody response to respiratory syncytial virus infection in children <18 months old. Hum. Vaccin. Immunother. 2016, 12, 1700–1706. [Google Scholar]
- López, E.L.; Ferolla, F.M.; Toledano, A.; Yfran, E.W.; Giordano, A.C.; Carrizo, B.; Feldman, F.; Talarico, L.B.; Caratozzolo, A.; Contrini, M.M.; et al. Genetic Susceptibility to Life-threatening Respiratory Syncytial Virus Infection in Previously Healthy Infants. Pediatr. Infect. Dis. J. 2020, 39, 1057–1061. [Google Scholar] [CrossRef]
- Pasanen, A.; Karjalainen, M.K.; Bont, L.; Piippo-Savolainen, E.; Ruotsalainen, M.; Goksör, E.; Kumawat, K.; Hodemaekers, H.; Nuolivirta, K.; Jartti, T.; et al. Genome-Wide Association Study of Polymorphisms Predisposing to Bronchiolitis. Sci. Rep. 2017, 7, 41653. [Google Scholar] [CrossRef]
- Canducci, F.; Debiaggi, M.; Sampaolo, M.; Marinozzi, M.C.; Berrè, S.; Terulla, C.; Gargantini, G.; Cambieri, P.; Romero, E.; Clementi, M. Two-year prospective study of single infections and co-infections by respiratory syncytial virus and viruses identified recently in infants with acute respiratory disease. J. Med. Virol. 2008, 80, 716–723. [Google Scholar] [CrossRef]
- Sigurs, N.; Gustafsson, P.M.; Bjarnason, R.; Lundberg, F.; Schmidt, S.; Sigurbergsson, F.; Kjellman, B. Severe respiratory syncytial virus bronchiolitis in infancy and asthma and allergy at age 13. Am. J. Respir. Crit. Care Med. 2005, 171, 137–141. [Google Scholar] [CrossRef]
- Sigurs, N.; Aljassim, F.; Kjellman, B.; Robinson, P.D.; Sigurbergsson, F.; Bjarnason, R.; Gustafsson, P.M. Asthma and allergy patterns over 18 years after severe RSV bronchiolitis in the first year of life. Thorax 2010, 65, 1045–1052. [Google Scholar] [CrossRef]
- Lambert, L.; Culley, F.J. Innate immunity to respiratory infection in early life. Front. Immunol. 2017, 8, 1570. [Google Scholar] [PubMed]
- Lemanske, R.F. The Childhood Origins of Asthma (COAST) study. Pediatr. Allergy Immunol. 2002, 13, 38–43. [Google Scholar] [PubMed]
- Granell, R.; Henderson, A.J.; Sterne, J.A. Associations of wheezing phenotypes with late asthma outcomes in the Avon Longitudinal Study of Parents and Children: A population-based birth cohort. J. Allergy Clin. Immunol. 2016, 138, 1060–1070. [Google Scholar]
- Openshaw, P.J.; Dean, G.S.; Culley, F.J. Links between respiratory syncytial virus bronchiolitis and childhood asthma: Clinical and research approaches. Pediatr. Infect. Dis. J. 2003, 22, S58–S65. [Google Scholar] [PubMed]
- Chu, C.-Y.; Qiu, X.; McCall, M.N.; Wang, L.; Corbett, A.; Holden-Wiltse, J.; Slaunwhite, C.; Grier, A.; Gill, S.R.; Pryhuber, G.S.; et al. Airway Gene Expression Correlates of Respiratory Syncytial Virus Disease Severity and Microbiome Composition in Infants. J. Infect. Dis. 2021, 223, 1639–1649. [Google Scholar]
- Saluzzo, S.; Gorki, A.D.; Rana, B.M.J.; Martins, R.; Scanlon, S.; Starkl, P.; Lakovits, K.; Hladik, A.; Korosec, A.; Sharif, O.; et al. First-Breath-Induced Type 2 Pathways Shape the Lung Immune Environment. Cell Rep. 2017, 18, 1893–1905. [Google Scholar]
- Zhivaki, D.; Lemoine, S.; Lim, A.; Morva, A.; Vidalain, P.O.; Schandene, L.; Casartelli, N.; Rameix-Welti, M.A.; Hervé, P.L.; Dériaud, E.; et al. Respiratory Syncytial Virus Infects Regulatory B Cells in Human Neonates via Chemokine Receptor CX3CR1 and Promotes Lung Disease Severity. Immunity 2017, 46, 301–314. [Google Scholar]
- Thwaites, R.S.; Coates, M.; Ito, K.; Ghazaly, M.; Feather, C.; Abdulla, F.; Tunstall, T.; Jain, P.; Cass, L.; Rapeport, G.; et al. Reduced Nasal Viral Load and IFN Responses in Infants with Respiratory Syncytial Virus Bronchiolitis and Respiratory Failure. Am. J. Respir. Crit. Care Med. 2018, 198, 1074–1084. [Google Scholar]
- Garcia-Mauriño, C.; Moore-Clingenpeel, M.; Thomas, J.; Mertz, S.; Cohen, D.M.; Ramilo, O.; Mejias, A. Viral Load Dynamics and Clinical Disease Severity in Infants with Respiratory Syncytial Virus Infection. J. Infect. Dis. 2018, 219, 1207–1215. [Google Scholar]
- Vu, L.; Siefker, D.; Jones, T.; You, D.; Taylor, R.; DeVincenzo, J.; Cormier, S. Elevated Levels of Type 2 Respiratory Innate Lymphoid Cells in Human Infants with Severe Respiratory Syncytial Virus Bronchiolitis. Am. J. Respir. Crit. Care Med. 2019, 200, 1414–1423. [Google Scholar]
- Torres, J.P.; Gomez, A.M.; Khokhar, S.; Bhoj, V.G.; Tagliabue, C.; Chang, M.L.; Kiener, P.A.; Revell, P.A.; Ramilo, O.; Mejias, A. Respiratory Syncytial Virus (RSV) RNA loads in peripheral blood correlates with disease severity in mice. Respir. Res. 2010, 11, 125. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mella, C.; Suarez-Arrabal, M.C.; Lopez, S.; Stephens, J.; Fernandez, S.; Hall, M.W.; Ramilo, O.; Mejias, A. Innate immune dysfunction is associated with enhanced disease severity in infants with severe respiratory syncytial virus bronchiolitis. J. Infect. Dis. 2013, 207, 564–573. [Google Scholar] [PubMed]
- Vieira, R.A.; de Diniz, E.M.A.; Ceccon, M.E.J.R. Correlation between inflammatory mediators in the nasopharyngeal secretion and in the serum of children with lower respiratory tract infection caused by respiratory syncytial virus and disease severity. J. Bras. Pneumol. 2010, 36, 59–66. [Google Scholar] [PubMed]
- Pitrez, P.M.C.; Brennan, S.; Sly, P.D. Inflammatory profile in nasal secretions of infants hospitalized with acute lower airway tract infections. Respirology 2005, 10, 365–370. [Google Scholar] [CrossRef]
- McNamara, P.S.; Flanagan, B.F.; Selby, A.M.; Hart, C.A.; Smyth, R.L. Pro- and anti-inflammatory responses in respiratory syncytial virus bronchiolitis. Eur. Respir. J. 2004, 23, 106–112. [Google Scholar] [PubMed]
- Legg, J.P.; Hussain, I.R.; Warner, J.A.; Johnston, S.L.; Warner, J.O. Type 1 and type 2 cytokine imbalance in acute respiratory syncytial virus bronchiolitis. Am. J. Respir. Crit. Care Med. 2003, 168, 633–639. [Google Scholar]
- Hornsleth, A.; Loland, L.; Larsen, L.B. Cytokines and chemokines in respiratory secretion and severity of disease in infants with respiratory syncytial virus (RSV) infection. J. Clin. Virol. 2001, 21, 163–170. [Google Scholar]
- Thwaites, R.S.; Ito, K.; Chingono, J.M.S.; Coates, M.; Jarvis, H.C.; Tunstall, T.; Anderson-Dring, L.; Cass, L.; Rapeport, G.; Openshaw, P.J.; et al. Nasosorption as a minimally invasive sampling procedure: Mucosal viral load and inflammation in primary RSV bronchiolitis. J. Infect. Dis. 2017, 215, 1240–1244. [Google Scholar]
- Fan, R.; Wen, B.; Liu, W.; Zhang, J.; Liu, C.; Fan, C.; Qu, X. Altered regulatory cytokine profiles in cases of pediatric respiratory syncytial virus infection. Cytokine 2018, 103, 57–62. [Google Scholar] [CrossRef]
- Caserta, M.T.; Qiu, X.; Tesini, B.; Wang, L.; Murphy, A.; Corbett, A.; Topham, D.J.; Falsey, A.R.; Holden-Wiltse, J.; Walsh, E.E. Development of a Global Respiratory Severity Score for Respiratory Syncytial Virus Infection in Infants. J. Infect. Dis. 2017, 215, 750–756. [Google Scholar] [CrossRef]
- Shi, T.; Balsells, E.; Wastnedge, E.; Singleton, R.; Rasmussen, Z.A.; Zar, H.J.; Rath, B.A.; Madhi, S.A.; Campbell, S.; Vaccari, L.C.; et al. Risk factors for respiratory syncytial virus associated with acute lower respiratory infection in children under five years: Systematic review and meta-analysis. J. Glob. Health 2015, 5, 020416. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, C.M.; Snelgrove, R.J. Type 2 immunity: Expanding our view. Sci. Immunol. 2018, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Geginat, J.; Paroni, M.; Maglie, S.; Alfen, J.S.; Kastirr, I.; Gruarin, P.; de Simone, M.; Pagani, M.; Abrignani, S. Plasticity of human CD4 T cell subsets. Front. Immunol. 2014, 5, 630. [Google Scholar] [CrossRef] [PubMed]
- Tuzlak, S.; Dejean, A.S.; Iannacone, M.; Quintana, F.J.; Waisman, A.; Ginhoux, F.; Korn, T.; Becher, B. Repositioning TH cell polarization from single cytokines to complex help. Nat. Immunol. 2021, 22, 1210–1217. [Google Scholar] [CrossRef]
- Pinto, R.A.; Arredondo, S.M.; Bono, M.R.; Gaggero, A.A.; Díaz, P.V. T helper 1/T helper 2 cytokine imbalance in respiratory syncytial virus infection is associated with increased endogenous plasma cortisol. Pediatrics 2006, 117, e878–e886. [Google Scholar] [CrossRef] [PubMed]
- Van Benten, I.J.; Van Drunen, C.M.; Koopman, L.P.; Van Middelkoop, B.C.; Hop, W.C.J.; Osterhaus, A.D.M.E.; Neijens, H.J.; Fokkens, W.J. Age- and infection-related maturation of the nasal immune response in 0–2-year-old children. Allergy Eur. J. Allergy Clin. Immunol. 2005, 60, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Stokholm, J.; Brejnrod, A.; Vestergaard, G.A.; Russel, J.; Trivedi, U.; Thorsen, J.; Gupta, S.; Hjelmsø, M.H.; Shah, S.A.; et al. The infant gut resistome associates with E. coli, environmental exposures, gut microbiome maturity, and asthma-associated bacterial composition. Cell Host Microbe 2021, 29, 975–987.e4. [Google Scholar] [CrossRef]
- Grier, A.; Gill, A.L.; Kessler, H.A.; Corbett, A.; Bandyopadhyay, S.; Java, J.; Holden-Wiltse, J.; Falsey, A.R.; Topham, D.J.; Mariani, T.J.; et al. Temporal Dysbiosis of Infant Nasal Microbiota Relative to Respiratory Syncytial Virus Infection. J. Infect. Dis. 2021, 223, 1650–1658. [Google Scholar] [CrossRef]
- Verwey, C.; Nunes, M.C. RSV lower respiratory tract infection and lung health in the first 2 years of life. Lancet Glob. Health 2020, 8, e1247–e1248. [Google Scholar] [CrossRef]
- Habibi, M.S.; Thwaites, R.S.; Chang, M.; Jozwik, A.; Paras, A.; Kirsebom, F.; Varese, A.; Owen, A.; Cuthbertson, L.; James, P.; et al. Neutrophilic inflammation in the respiratory mucosa predisposes to RSV infection. Science 2020, 370, aba9301. [Google Scholar] [CrossRef]
- Hu, G.; Christman, J.W. Editorial: Alveolar Macrophages in Lung Inflammation and Resolution. Front. Immunol. 2019, 10, 2275. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.Y.S.; Krasnow, M.A. Developmental origin of lung macrophage diversity. Development 2016, 143, 1318–1327. [Google Scholar] [CrossRef] [PubMed]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage polarization: Different gene signatures in M1(Lps+) vs. Classically and M2(LPS-) vs. Alternatively activated macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef] [PubMed]
- Mitsi, E.; Kamng’ona, R.; Rylance, J.; Solórzano, C.; Jesus Reiné, J.; Mwandumba, H.C.; Ferreira, D.M.; Jambo, K.C. Human alveolar macrophages predominately express combined classical M1 and M2 surface markers in steady state. Respir. Res. 2018, 19, 66. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef]
- Viola, A.; Munari, F.; Sánchez-Rodríguez, R.; Scolaro, T.; Castegna, A. The metabolic signature of macrophage responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef]
- Makita, N.; Hizukuri, Y.; Yamashiro, K.; Murakawa, M.; Hayashi, Y. IL-10 enhances the phenotype of M2 macrophages induced by IL-4 and confers the ability to increase eosinophil migration. Int. Immunol. 2015, 27, 131–141. [Google Scholar] [CrossRef]
- Jones, C.V.; Williams, T.M.; Walker, K.A.; Dickinson, H.; Sakkal, S.; Rumballe, B.A.; Little, M.H.; Jenkin, G.; Ricardo, S.D. M2 macrophage polarisation is associated with alveolar formation during postnatal lung development. Respir. Res. 2013, 14, 41. [Google Scholar] [CrossRef]
- Makris, S.; Bajorek, M.; Culley, F.J.; Goritzka, M.; Johansson, C. Alveolar macrophages can control respiratory syncytial virus infection in the absence of Type I interferons. J. Innate Immun. 2016, 8, 452–463. [Google Scholar] [CrossRef]
- Goenka, A.; Prise, I.E.; Connolly, E.; Fernandez-Soto, P.; Morgan, D.; Cavet, J.S.; Grainger, J.R.; Nichani, J.; Arkwright, P.D.; Hussell, T. Infant Alveolar Macrophages Are Unable to Effectively Contain Mycobacterium tuberculosis. Front. Immunol. 2020, 11, 486. [Google Scholar] [CrossRef]
- Campbell, D.J.; Ziegler, S.F. Opinion: FOXP3 modifies the phenotypic and functional properties of regulatory T cells. Nat. Rev. Immunol. 2007, 7, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Miyahara, Y.; Guo, Z.; Khattar, M.; Stepkowski, S.M.; Chen, W. “Default” Generation of Neonatal Regulatory T Cells. J. Immunol. 2010, 185, 71–78. [Google Scholar] [CrossRef]
- Kim, C.H. FOXP3 and its role in the immune system. Adv. Exp. Med. Biol. 2009, 665, 17–29. [Google Scholar] [PubMed]
- Christiaansen, A.F.; Syed, M.A.; Ten Eyck, P.P.; Hartwig, S.M.; Durairaj, L.; Kamath, S.S.; Varga, S.M. Altered Treg and cytokine responses in RSV-infected infants. Pediatr. Res. 2016, 80, 702–709. [Google Scholar] [CrossRef] [PubMed]
- Miyara, M.; Yoshioka, Y.; Kitoh, A.; Shima, T.; Wing, K.; Niwa, A.; Parizot, C.; Taflin, C.; Heike, T.; Valeyre, D.; et al. Functional Delineation and Differentiation Dynamics of Human CD4+ T Cells Expressing the FoxP3 Transcription Factor. Immunity 2009, 30, 899–911. [Google Scholar] [CrossRef]
- Aluvihare, V.R.; Kallikourdis, M.; Betz, A.G. Regulatory T cells mediate maternal tolerance to the fetus. Nat. Immunol. 2004, 5, 266–271. [Google Scholar] [CrossRef]
- Zahran, A.M.; Saad, K.; Abdel-Raheem, Y.F.; Elsayh, K.I.; El-Houfey, A.A.; Aboul-Khair, M.D.; Alblihed, M.A. Characterization of Regulatory T Cells in Preterm and Term Infants. Arch. Immunol. Ther. Exp. 2019, 67, 49–54. [Google Scholar] [CrossRef]
- Kumar, P.; Saini, S.; Khan, S.; Surendra Lele, S.; Prabhakar, B.S. Restoring self-tolerance in autoimmune diseases by enhancing regulatory T-cells. Cell. Immunol. 2019, 339, 41–49. [Google Scholar] [CrossRef]
- Durant, L.R.; Makris, S.; Voorburg, C.M.; Loebbermann, J.; Johansson, C.; Openshaw, P.J.M. Regulatory T cells prevent Th2 immune responses and pulmonary eosinophilia during respiratory syncytial virus infection in mice. J. Virol. 2013, 87, 10946–10954. [Google Scholar] [CrossRef]
- Raiden, S.; Pandolfi, J.; Payasliàn, F.; Anderson, M.; Rivarola, N.; Ferrero, F.; Urtasun, M.; Fainboim, L.; Geffner, J.; Arruvito, L. Depletion of circulating regulatory T cells during severe respiratory syncytial virus infection in young children. Am. J. Respir. Crit. Care Med. 2014, 189, 865–868. [Google Scholar] [CrossRef]
- Mubarak, A.; Ahmed, M.S.; Upile, N.; Vaughan, C.; Xie, C.; Sharma, R.; Acar, P.; McCormick, M.S.; Paton, J.C.; Mitchell, T.; et al. A dynamic relationship between mucosal T helper type 17 and regulatory T-cell populations in nasopharynx evolves with age and associates with the clearance of pneumococcal carriage in humans. Clin. Microbiol. Infect. 2016, 22, 736.e1–736.e7. [Google Scholar] [CrossRef]
- Fulton, R.B.; Meyerholz, D.K.; Varga, S.M. Foxp3+ CD4 regulatory T cells limit pulmonary immunopathology by modulating the CD8 T cell response during respiratory syncytial virus infection. J. Immunol. 2010, 185, 2382–2392. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Baba, A.; Yokota, T.; Nishikawa, H.; Ohkawa, Y.; Kayama, H.; Kallies, A.; Nutt, S.L.; Sakaguchi, S.; Takeda, K.; et al. Interleukin-10-producing plasmablasts exert regulatory function in autoimmune inflammation. Immunity 2014, 41, 1040–1051. [Google Scholar] [CrossRef] [PubMed]
- Rosser, E.C.; Mauri, C. Regulatory B Cells: Origin, Phenotype, and Function. Immunity 2015, 42, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Laubreton, D.; Drajac, C.; Eléou t, J.F.; Rameix-Welti, M.A.; Lo-Man, R.; Riffault, S.; Descamps, D. Regulatory B lymphocytes colonize the respiratory tract of neonatal mice and modulate immune responses of alveolar macrophages to RSV infection in IL-10-dependant manner. Viruses 2020, 12, 822. [Google Scholar] [CrossRef]
- Openshaw, P.J.M. RSV Takes Control of Neonatal Breg Cells: Two Hands on the Wheel. Immunity 2017, 46, 171–173. [Google Scholar] [CrossRef]
- Kinnear, E.; Lambert, L.; McDonald, J.; Cheeseman, H.; Caproni, L.; Tregoning, J. Airway T cells protect against RSV infection in the absence of antibody. Mucosal Immunol. 2018, 11, 249–256. [Google Scholar] [CrossRef]
- Zens, K.D.; Chen, J.K.; Guyer, R.S.; Wu, F.L.; Cvetkovski, F.; Miron, M.; Farber, D.L. Reduced generation of lung tissue–resident memory T cells during infancy. J. Exp. Med. 2017, 214, 2915. [Google Scholar] [CrossRef]
- Connors, T.J.; Baird, S.J.; Yopes, M.C.; Zens, K.D.; Pethe, K.; Ravindranath, T.M.; Ho, S.; Farber, D.L. Developmental Regulation of Effector and Resident Memory T Cell Generation during Pediatric Viral Respiratory Tract Infection. J. Immunol. 2018, 201, 432–439. [Google Scholar] [CrossRef]
- Gill, M.A.; Long, K.; Kwon, T.; Muniz, L.; Mejias, A.; Connolly, J.; Roy, L.; Banchereau, J.; Ramilo, O. Differential Recruitment of Dendritic Cells and Monocytes to Respiratory Mucosal Sites in Children with Influenza Virus or Respiratory Syncytial Virus Infection. J. Infect. Dis. 2008, 198, 1667–1776. [Google Scholar] [CrossRef]
- Gurram, R.K.; Zhu, J. Orchestration between ILC2s and Th2 cells in shaping type 2 immune responses. Cell. Mol. Immunol. 2019, 16, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Siefker, D.T.; Vu, L.; You, D.; McBride, A.; Taylor, R.; Jones, T.L.; DeVincenzo, J.; Cormier, S.A. Respiratory syncytial virus disease severity is associated with distinct CD8+ T-cell profiles. Am. J. Respir. Crit. Care Med. 2020, 201, 325–334. [Google Scholar] [CrossRef]
- Van Benten, I.J.; Van Drunen, C.M.; Koopman, L.P.; KleinJan, A.; Van Middelkoop, B.C.; De Waal, L.; Osterhaus, A.D.M.E.; Neijens, H.J.; Fokkens, W.J. RSV-induced bronchiolitis but not upper respiratory tract infection is accompanied by an increased nasal IL-18 response. J. Med. Virol. 2003, 71, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Deprez, M.; Zaragosi, L.-E.; Truchi, M.; Becavin, C.; García, S.R.; Arguel, M.-J.; Plaisant, M.; Magnone, V.; Lebrigand, K.; Abelanet, S.; et al. A Single-Cell Atlas of the Human Healthy Airways. Am. J. Respir. Crit. Care Med. 2020, 202, 1636–1645. [Google Scholar] [CrossRef]
- Chu, C.Y.; Qiu, X.; Wang, L.; Bhattacharya, S.; Lofthus, G.; Corbett, A.; Holden-Wiltse, J.; Grier, A.; Tesini, B.; Gill, S.R.; et al. The Healthy Infant Nasal Transcriptome: A Benchmark Study. Sci. Rep. 2016, 6, 33994. [Google Scholar] [CrossRef]
- Yu, J.; Peterson, D.R.; Baran, A.M.; Bhattacharya, S.; Wylie, T.N.; Falsey, A.R.; Mariani, T.J.; Storch, G.A. Host Gene Expression in Nose and Blood for the Diagnosis of Viral Respiratory Infection. J. Infect. Dis. 2019, 219, 1151–1161. [Google Scholar] [CrossRef]
- Antunes, K.H.; Becker, A.; Franceschina, C.; de Freitas, D.N.; Lape, I.; da Cunha, M.D.; Leitão, L.; Rigo, M.M.; Pinto, L.A.; Stein, R.T.; et al. Respiratory syncytial virus reduces STAT3 phosphorylation in human memory CD8 T cells stimulated with IL-21. Sci. Rep. 2019, 9, 17766. [Google Scholar] [CrossRef] [PubMed]
- Deenick, E.K.; Pelham, S.J.; Kane, A.; Ma, C.S. Signal transducer and activator of transcription 3 control of human T and B cell responses. Front. Immunol. 2018, 9, 168. [Google Scholar] [CrossRef]
- Connors, T.J.; Ravindranath, T.M.; Bickham, K.L.; Gordon, C.L.; Zhang, F.; Levin, B.; Baird, J.S.; Farber, D.L. Airway CD8 1 T Cells Are Associated with Lung Injury during Infant Viral Respiratory Tract Infection. Am. J. Respir. Cell Mol. Biol. 2016, 54, 822–830. [Google Scholar] [CrossRef]
- Munir, S.; Le Nouen, C.; Luongo, C.; Buchholz, U.J.; Collins, P.L.; Bukreyev, A. Nonstructural proteins 1 and 2 of respiratory syncytial virus suppress maturation of human dendritic cells. J. Virol. 2008, 82, 8780–8796. [Google Scholar] [CrossRef]
- Bont, L.; Heijnen, C.J.; Kavelaars, A.; Van Aalderen, W.M.C.; Brus, F.; Draaisma, J.T.M.; Geelen, S.M.; Kimpen, J.L.L. Monocyte IL-10 production during respiratory syncytial virus bronchiolitis is associated with recurrent wheezing in a one-year follow-up study. Am. J. Respir. Crit. Care Med. 2000, 161, 1518–1523. [Google Scholar] [CrossRef] [PubMed]
- Ahout, I.M.L.; Jans, J.; Haroutiounian, L.; Simonetti, E.R.; Van Der Gaast-De Jongh, C.; Diavatopoulos, D.A.; De Jonge, M.I.; De Groot, R.; Ferwerda, G. Reduced expression of HLA-DR on monocytes during severe respiratory syncytial virus infections. Pediatr. Infect. Dis. J. 2016, 35, e89–e96. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Quirós, A.; González, H.; Arranz, E.; Lapeña, S. Decreased interleukin-12 levels in umbilical cord blood in children who developed acute bronchiolitis. Pediatr. Pulmonol. 1999, 28, 175–180. [Google Scholar] [CrossRef]
- Juntti, H.; Österlund, P.; Kokkonen, J.; Dunder, T.; Renko, M.; Pokka, T.; Julkunen, I.; Uhari, M. Cytokine responses in cord blood predict the severity of later respiratory syncytial virus infection. J. Allergy Clin. Immunol. 2009, 124, 52–58. [Google Scholar] [CrossRef]
- Schuurhof, A.; Janssen, R.; de Groot, H.; Hodemaekers, H.M.; de Klerk, A.; Kimpen, J.L.L.; Bont, L. Local interleukin-10 production during respiratory syncytial virus bronchiolitis is associated with post-bronchiolitis wheeze. Respir. Res. 2011, 12, 121. [Google Scholar] [CrossRef]
- Thwaites, R.S.; Jarvis, H.C.; Singh, N.; Jha, A.; Pritchard, A.; Fan, H.; Tunstall, T.; Nanan, J.; Nadel, S.; Kon, O.M.; et al. Absorption of Nasal and Bronchial Fluids: Precision Sampling of the Human Respiratory Mucosa and Laboratory Processing of Samples. JoVE J. Vis. Exp. 2018, 2018, e56413. [Google Scholar] [CrossRef]
- Chung, J.Y.; Han, T.H.; Kim, J.S.; Kim, S.W.; Park, C.G.; Hwang, E.S. Th1 and Th2 cytokine levels in nasopharyngeal aspirates from children with human bocavirus bronchiolitis. J. Clin. Virol. 2008, 43, 223–225. [Google Scholar] [CrossRef]
- Pancham, K.; Perez, G.F.; Huseni, S.; Jain, A.; Kurdi, B.; Rodriguez-Martinez, C.E.; Preciado, D.; Rose, M.C.; Nino, G. Premature infants have impaired airway antiviral IFNγ responses to human metapneumovirus compared to respiratory syncytial virus. Pediatr. Res. 2015, 78, 389–394. [Google Scholar] [CrossRef]
- Murai, H.; Terada, A.; Mizuno, M.; Asai, M.; Hirabayashi, Y.; Shimizu, S.; Morishita, T.; Kakita, H.; Hussein, M.H.; Ito, T.; et al. IL-10 and RANTES are elevated in nasopharyngeal secretions of children with respiratory syncytial virus infection. Allergol. Int. 2007, 56, 157–163. [Google Scholar] [CrossRef]
- Chung, H.L.; Park, H.J.; Kim, S.Y.; Kim, S.G. Age-related difference in immune responses to respiratory syncytial virus infection in young children. Pediatr. Allergy Immunol. 2007, 18, 94–99. [Google Scholar] [CrossRef]
- Schuler, C.F.; Malinczak, C.A.; Best, S.K.K.; Morris, S.B.; Rasky, A.J.; Ptaschinski, C.; Lukacs, N.W.; Fonseca, W. Inhibition of uric acid or IL-1β ameliorates respiratory syncytial virus immunopathology and development of asthma. Allergy Eur. J. Allergy Clin. Immunol. 2020, 75, 2279–2293. [Google Scholar] [CrossRef] [PubMed]
- Pino, M.; Kelvin, D.J.; Bermejo-Martin, J.F.; Alonso, A.; Matías, V.; Tenorio, A.; Rico, L.; Eiros, J.M.; Castrodeza, J.; Blanco-Quiros, A.; et al. Nasopharyngeal aspirate cytokine levels 1 yr after severe respiratory syncytial virus infection. Pediatr. Allergy Immunol. 2009, 20, 791–795. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.Y.; Cheng, T.Y.; Yang, L.; Huang, Y.H.; Li, C.; Han, J.Z.; Li, X.H.; Fang, L.J.; Feng, D.D.; Tang, Y.T.; et al. G-CSF Inhibits Pulmonary Fibrosis by Promoting BMSC Homing to the Lungs via SDF-1/CXCR4 Chemotaxis. Sci. Rep. 2020, 10, 10515. [Google Scholar] [CrossRef] [PubMed]
- Saravia, J.; You, D.; Shrestha, B.; Jaligama, S.; Siefker, D.; Lee, G.I.; Harding, J.N.; Jones, T.L.; Rovnaghi, C.; Bagga, B.; et al. Respiratory Syncytial Virus Disease Is Mediated by Age-Variable IL-33. PLoS Pathog. 2015, 11, e1005217. [Google Scholar] [CrossRef]
- Mejias, A.; Dimo, B.; Suarez, N.M.; Garcia, C.; Suarez-Arrabal, M.C.; Jartti, T.; Blankenship, D.; Jordan-Villegas, A.; Ardura, M.I.; Xu, Z.; et al. Whole Blood Gene Expression Profiles to Assess Pathogenesis and Disease Severity in Infants with Respiratory Syncytial Virus Infection. PLoS Med. 2013, 10, e1001549. [Google Scholar] [CrossRef]
- Bertrand, P.; Lay, M.K.; Piedimonte, G.; Brockmann, P.E.; Palavecino, C.E.; Hernández, J.; León, M.A.; Kalergis, A.M.; Bueno, S.M. Elevated IL-3 and IL-12p40 levels in the lower airway of infants with RSV-induced bronchiolitis correlate with recurrent wheezing. Cytokine 2015, 76, 417–423. [Google Scholar] [CrossRef]
- Good, M.; Kolls, J.K.; Empey, K.M. Neonatal Pulmonary Host Defense. Fetal Neonatal Physiol. 2017, 2017, 1262–1293.e12. [Google Scholar]
- Chung, H.L.; Kim, W.T.; Kim, J.K.; Choi, E.J.; Lee, J.H.; Lee, G.H.; Kim, S.G. Relationship between atopic status and nasal interleukin 10 and 11 levels in infants with respiratory syncytial virus bronchiolitis. Ann. Allergy Asthma Immunol. 2005, 94, 267–272. [Google Scholar] [CrossRef]
- Díaz, P.V.; Valdivia, G.; Gaggero, A.A.; Bono, M.R.; Zepeda, G.; Rivas, M.; Uasapud, P.; Pinto, R.A.; Boza, M.L.; Guerrero, J. Pro-inflammatory cytokines in nasopharyngeal aspirate from hospitalized children with respiratory syncytial virus infection with or without rhinovirus bronchiolitis, and use of the cytokines as predictors of illness severity. Medicine 2015, 94, e1512. [Google Scholar] [CrossRef]
- Welliver, R.C. Respiratory syncytial virus and other respiratory viruses. Pediatr. Infect. Dis. J. 2003, 22, S6–S12. [Google Scholar] [CrossRef]
- Boncristiani, H.F.; Criado, M.F.; Arruda, E. Respiratory Viruses. In Encyclopedia of Microbiology; Elsevier Inc.: Amsterdam, The Netherlands, 2009; pp. 500–518. ISBN 9780123739445. [Google Scholar]
- Hasegawa, K.; Pérez-Losada, M.; Hoptay, C.E.; Epstein, S.; Mansbach, J.M.; Teach, S.J.; Piedra, P.A.; Camargo, C.A.; Freishtat, R.J. RSV vs. rhinovirus bronchiolitis: Difference in nasal airway microRNA profiles and NF B signaling. Pediatr. Res. 2018, 83, 606–614. [Google Scholar] [CrossRef]
- Turi, K.N.; Shankar, J.; Anderson, L.J.; Rajan, D.; Gaston, K.; Gebretsadik, T.; Das, S.R.; Stone, C.; Larkin, E.K.; Rosas-Salazar, C.; et al. Infant Viral Respiratory Infection Nasal Immune-Response Patterns and Their Association with Subsequent Childhood Recurrent Wheeze. Am. J. Respir. Crit. Care Med. 2018, 198, 1064–1073. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Xu, L.; Zhu, Y.; Xu, B.; Chen, X.; Duan, Y.; Xie, Z.; Shen, K. Comparison of clinical features of acute lower respiratory tract infections in infants with RSV/HRV infection, and incidences of subsequent wheezing or asthma in childhood. BMC Infect. Dis. 2020, 20, 387. [Google Scholar] [CrossRef] [PubMed]
- Pezzulo, A.A.; Starner, T.D.; Scheetz, T.E.; Traver, G.L.; Tilley, A.E.; Harvey, B.G.; Crystal, R.G.; McCray, P.B.; Zabner, J. The air-liquid interface and use of primary cell cultures are important to recapitulate the transcriptional profile of in vivo airway epithelia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 300, L25–L31. [Google Scholar] [CrossRef] [PubMed]
- Groves Id, H.E.; Guo, H.; Id, P.; Broadbent, L.; Shields, M.D.; Power, U.F. Characterisation of morphological differences in well-differentiated nasal epithelial cell cultures from preterm and term infants at birth and one-year. PLoS ONE 2018, 13, e0201328. [Google Scholar] [CrossRef] [PubMed]
- Broadbent, L.; Manzoor, S.; Zarcone, M.C.; Barabas, J.; Shields, M.D.; Saglani, S.; Lloyd, C.M.; Bush, A.; Custovic, A.; Ghazal, P.; et al. Comparative primary paediatric nasal epithelial cell culture differentiation and RSV-induced cytopathogenesis following culture in two commercial media. PLoS ONE 2020, 15, e0228229. [Google Scholar] [CrossRef]
- Coulter, P.J.; Groves, H.; Broadbent, L.; Coey, J.D.; Shields, M.D.; Power, U.F. Early life innate immune responses to RSV in cystic fibrosis airway epithelium. Access Microbiol. 2019, 1, 887. [Google Scholar] [CrossRef]
- Levardon, H.; Yonker, L.M.; Hurley, B.P.; Mou, H. Expansion of Airway Basal Cells and Generation of Polarized Epithelium. Bio-Protocol 2018, 8, e2877. [Google Scholar] [CrossRef]
- Chirkova, T.; Lin, S.; Oomens, A.G.P.; Gaston, K.A.; Boyoglu-Barnum, S.; Meng, J.; Stobart, C.C.; Cotton, C.U.; Hartert, T.V.; Moore, M.L.; et al. CX3CR1 is an important surface molecule for respiratory syncytial virus infection in human airway epithelial cells. J. Gen. Virol. 2015, 96, 2543–2556. [Google Scholar] [CrossRef]
- Villenave, R.; Broadbent, L.; Douglas, I.; Lyons, J.D.; Coyle, P.V.; Teng, M.N.; Tripp, R.A.; Heaney, L.G.; Shields, M.D.; Power, U.F. Induction and Antagonism of Antiviral Responses in Respiratory Syncytial Virus-Infected Pediatric Airway Epithelium. J. Virol. 2015, 89, 12309. [Google Scholar] [CrossRef]
- Persson, B.D.; Jaffe, A.B.; Fearns, R.; Danahay, H. Respiratory syncytial virus can infect basal cells and alter human airway epithelial differentiation. PLoS ONE 2014, 9, e102368. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.; Carnoy, C.; Lamblin, G.; Roussel, P.; Kuik, J.A.; Vliegenthart, J.F.G. Isolation and structural characterization of novel sialylated oligosaccharide-alditols from respiratory-mucus glycoproteins of a patient suffering from bronchiectasis. Eur. J. Biochem. 1993, 211, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Linden, S.K.; Sutton, P.; Karlsson, N.G.; Korolik, V.; McGuckin, M.A. Mucins in the mucosal barrier to infection. Mucosal Immunol. 2008, 1, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Zanin, M.; Baviskar, P.; Webster, R.; Webby, R. The interaction between respiratory pathogens and mucus. Cell Host Microbe 2016, 19, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Denneny, E.; Sahota, J.; Beatson, R.; Thornton, D.; Burchell, J.; Porter, J. Mucins and their receptors in chronic lung disease. Clin. Transl. Immunol. 2020, 9, e01120. [Google Scholar] [CrossRef]
- Baños-Lara, M.D.R.; Piao, B.; Guerrero-Plata, A. Differential mucin expression by respiratory syncytial virus and human metapneumovirus infection in human epithelial cells. Mediators Inflamm. 2015, 2015, 347292. [Google Scholar] [CrossRef]
- Stokes, K.L.; Chi, M.H.; Sakamoto, K.; Newcomb, D.C.; Currier, M.G.; Huckabee, M.M.; Lee, S.; Goleniewska, K.; Pretto, C.; Williams, J.V.; et al. Differential Pathogenesis of Respiratory Syncytial Virus Clinical Isolates in BALB/c Mice. J. Virol. 2011, 85, 5782–5793. [Google Scholar] [CrossRef]
- Roy, M.G.; Livraghi-Butrico, A.; Fletcher, A.A.; McElwee, M.M.; Evans, S.E.; Boerner, R.M.; Alexander, S.N.; Bellinghausen, L.K.; Song, A.S.; Petrova, Y.M.; et al. Muc5b is required for airway defence. Nature 2014, 505, 412–416. [Google Scholar] [CrossRef]
- Li, Y.; Dinwiddie, D.L.; Harrod, K.S.; Jiang, Y.; Kim, K.C. Anti-inflammatory effect of MUC1 during respiratory syncytial virus infection of lung epithelial cells in vitro. Am. J. Physiol. Lung Cell. Mol. Physiol. 2010, 298, L558. [Google Scholar] [CrossRef]
- Du, X.; Yang, Y.; Xiao, G.; Yang, M.; Yuan, L.; Qin, L.; He, R.; Wang, L.; Wu, M.; Wu, S.Y.; et al. Respiratory syncytial virus infection-induced mucus secretion by down-regulation of miR-34b/c-5p expression in airway epithelial cells. J. Cell. Mol. Med. 2020, 24, 12694–12705. [Google Scholar] [CrossRef]
- Rajan, D.; O’Keefe, E.L.; Travers, C.; McCracken, C.; Geoghegan, S.; Caballero, M.T.; Acosta, P.L.; Polack, F.; Anderson, L.J. MUC5AC levels associated with respiratory syncytial virus disease severity. Clin. Infect. Dis. 2018, 67, 1441–1444. [Google Scholar] [CrossRef] [PubMed]
- Bonser, L.; Erle, D. Airway Mucus and Asthma: The Role of MUC5AC and MUC5B. J. Clin. Med. 2017, 6, 112. [Google Scholar] [CrossRef] [PubMed]
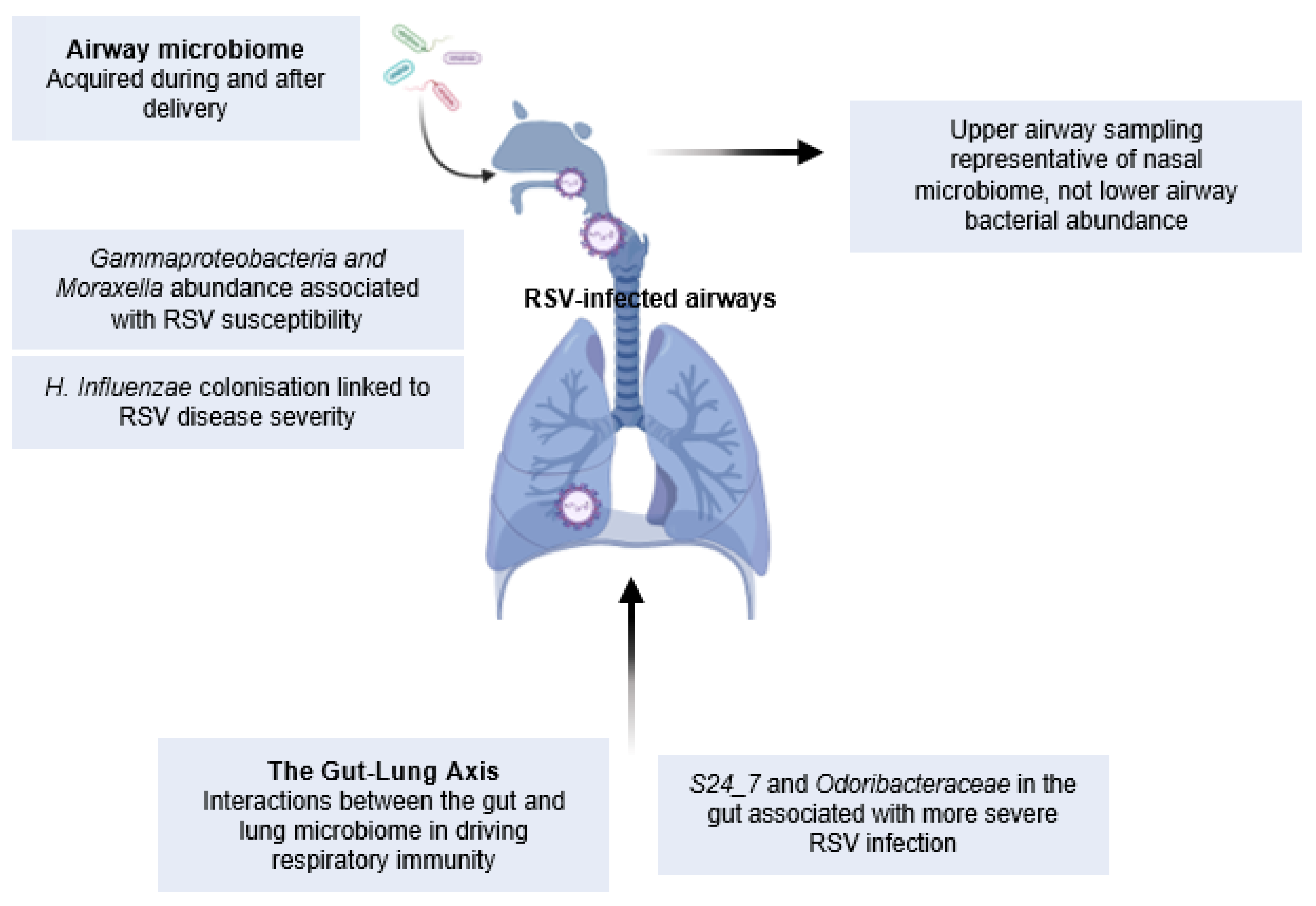
- Gallacher, D.J.; Kotecha, S. Respiratory microbiome of new-born infants. Front. Pediatr. 2016, 4, 10. [Google Scholar] [CrossRef]
- Biesbroek, G.; Bosch, A.A.T.M.; Wang, X.; Keijser, B.J.F.; Veenhoven, R.H.; Sanders, E.A.M.; Bogaert, D. The impact of breastfeeding on nasopharyngeal microbial communities in infants. Am. J. Respir. Crit. Care Med. 2014, 190, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Teo, S.M.; Mok, D.; Pham, K.; Kusel, M.; Serralha, M.; Troy, N.; Holt, B.J.; Hales, B.J.; Walker, M.L.; Hollams, E.; et al. The infant nasopharyngeal microbiome impacts severity of lower respiratory infection and risk of asthma development. Cell Host Microbe 2015, 17, 704–715. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, X.; Zhang, N.; Wang, X.; Sun, L.; Chen, N.; Zhao, S.; He, Q. Airway microbiome, host immune response and recurrent wheezing in infants with severe respiratory syncytial virus bronchiolitis. Pediatr. Allergy Immunol. 2020, 31, 281–289. [Google Scholar] [CrossRef]
- Sonawane, A.R.; Tian, L.; Chu, C.-Y.; Qiu, X.; Wang, L.; Holden-Wiltse, J.; Grier, A.; Gill, S.R.; caserta, M.; Falsey, A.R.; et al. Microbiome-transcriptome interactions Related to Severity of Respiratory Syncytial Virus infection. Sci. Rep. 2019, 9, 13824. [Google Scholar] [CrossRef]
- Harding, J.N.; Siefker, D.; Vu, L.; You, D.; Devincenzo, J.; Pierre, J.F.; Cormier, S.A. Altered gut microbiota in infants is associated with respiratory syncytial virus disease severity. BMC Microbiol. 2020, 20, 1–11. [Google Scholar] [CrossRef]
- De Steenhuijsen Piters, W.A.A.; Heinonen, S.; Hasrat, R.; Bunsow, E.; Smith, B.; Suarez-Arrabal, M.C.; Chaussabel, D.; Cohen, D.M.; Sanders, E.A.M.; Ramilo, O.; et al. Nasopharyngeal microbiota, host transcriptome, and disease severity in children with respiratory syncytial virus infection. Am. J. Respir. Crit. Care Med. 2016, 194, 1104–1115. [Google Scholar] [CrossRef]
- Brealey, J.C.; Young, P.R.; Sloots, T.P.; Ware, R.S.; Lambert, S.B.; Sly, P.D.; Grimwood, K.; Chappell, K.J. Bacterial colonization dynamics associated with respiratory syncytial virus during early childhood. Pediatr. Pulmonol. 2020, 55, 1237–1245. [Google Scholar] [CrossRef]
- Mansbach, J.M.; Luna, P.N.; Shaw, C.A.; Hasegawa, K.; Petrosino, J.F.; Piedra, P.A.; Sullivan, A.F.; Espinola, J.A.; Stewart, C.J.; Camargo, C.A. Increased Moraxella and Streptococcus species abundance after severe bronchiolitis is associated with recurrent wheezing. J. Allergy Clin. Immunol. 2020, 145, 518–527.e8. [Google Scholar] [CrossRef] [PubMed]
- Holster, A.; Nuolivirta, K.; Törmänen, S.; Lauhkonen, E.; Teräsjärvi, J.; Vuononvirta, J.; Koponen, P.; Helminen, M.; He, Q.; Korppi, M. Interleukin-10 gene polymorphism rs1800896 is associated with post-bronchiolitis asthma at 11–13 years of age. Acta Paediatr. Int. J. Paediatr. 2019, 108, 2064–2069. [Google Scholar] [CrossRef]
- Holster, A.; Teräsjärvi, J.; Vuononvirta, J.; Koponen, P.; Peltola, V.; Helminen, M.; He, Q.; Korppi, M.; Nuolivirta, K. Polymorphisms in the promoter region of IL10 gene are associated with virus etiology of infant bronchiolitis. World J. Pediatr. 2018, 14, 594–600. [Google Scholar] [CrossRef]
- Chambliss, J.M.; Ansar, M.; Kelley, J.P.; Spratt, H.; Garofalo, R.P.; Casola, A. A Polymorphism in the Catalase Gene Promoter Confers Protection against Severe RSV Bronchiolitis. Viruses 2020, 12, 57. [Google Scholar] [CrossRef] [PubMed]
- Leon-Icaza, S.A.; Zeng, M.; Rosas-Taraco, A.G. microRNAs in viral acute respiratory infections: Immune regulation, biomarkers, therapy, and vaccines. ExRNA 2019, 1, 1. [Google Scholar] [CrossRef] [PubMed]
- Bakre, A.; Mitchell, P.; Coleman, J.K.; Jones, L.P.; Saavedra, G.; Teng, M.; Tompkins Mark, S.; Tripp, R.A. Respiratory syncytial virus modifies microRNAs regulating host genes that affect virus replication. J. Gen. Virol. 2012, 93, 2346–2356. [Google Scholar] [CrossRef] [PubMed]
- Inchley, C.S.; Sonerud, T.; Fjærli, H.O.; Nakstad, B. Nasal mucosal microRNA expression in children with respiratory syncytial virus infection. BMC Infect. Dis. 2015, 15, 150. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, F.; Yang, D.; Peng, T.; Lu, G. Identification of miRNA-mRNA Crosstalk in Respiratory Syncytial Virus-(RSV-) Associated Pediatric Pneumonia through Integrated miRNAome and Transcriptome Analysis. Mediators Inflamm. 2020, 2020, 8919534. [Google Scholar] [CrossRef]
- Graff, J.W.; Dickson, A.M.; Clay, G.; McCaffrey, A.P.; Wilson, M.E. Identifying functional microRNAs in macrophages with polarized phenotypes. J. Biol. Chem. 2012, 287, 21816–21825. [Google Scholar] [CrossRef]
- Arroyo, M.; Salka, K.; Chorvinsky, E.; Xuchen, X.; Abutaleb, K.; Perez, G.F.; Weinstock, J.; Gaviria, S.; Gutierrez, M.J.; Nino, G. Airway mir-155 responses are associated with TH1 cytokine polarization in young children with viral respiratory infections. PLoS ONE 2020, 15, e0233352. [Google Scholar] [CrossRef]
- Fonseca, W.; Lukacs, N.W.; Ptaschinski, C. Factors affecting the immunity to respiratory syncytial virus: From epigenetics to microbiome. Front. Immunol. 2018, 9, 226. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barnes, M.V.C.; Openshaw, P.J.M.; Thwaites, R.S. Mucosal Immune Responses to Respiratory Syncytial Virus. Cells 2022, 11, 1153. https://doi.org/10.3390/cells11071153
Barnes MVC, Openshaw PJM, Thwaites RS. Mucosal Immune Responses to Respiratory Syncytial Virus. Cells. 2022; 11(7):1153. https://doi.org/10.3390/cells11071153
Chicago/Turabian StyleBarnes, Megan V. C., Peter J. M. Openshaw, and Ryan S. Thwaites. 2022. "Mucosal Immune Responses to Respiratory Syncytial Virus" Cells 11, no. 7: 1153. https://doi.org/10.3390/cells11071153
APA StyleBarnes, M. V. C., Openshaw, P. J. M., & Thwaites, R. S. (2022). Mucosal Immune Responses to Respiratory Syncytial Virus. Cells, 11(7), 1153. https://doi.org/10.3390/cells11071153