
Cryopreservation by Directional Freezing and Vitrification Focusing on Large Tissues and Organs


{kind=link}
Abstract
:1. Introduction
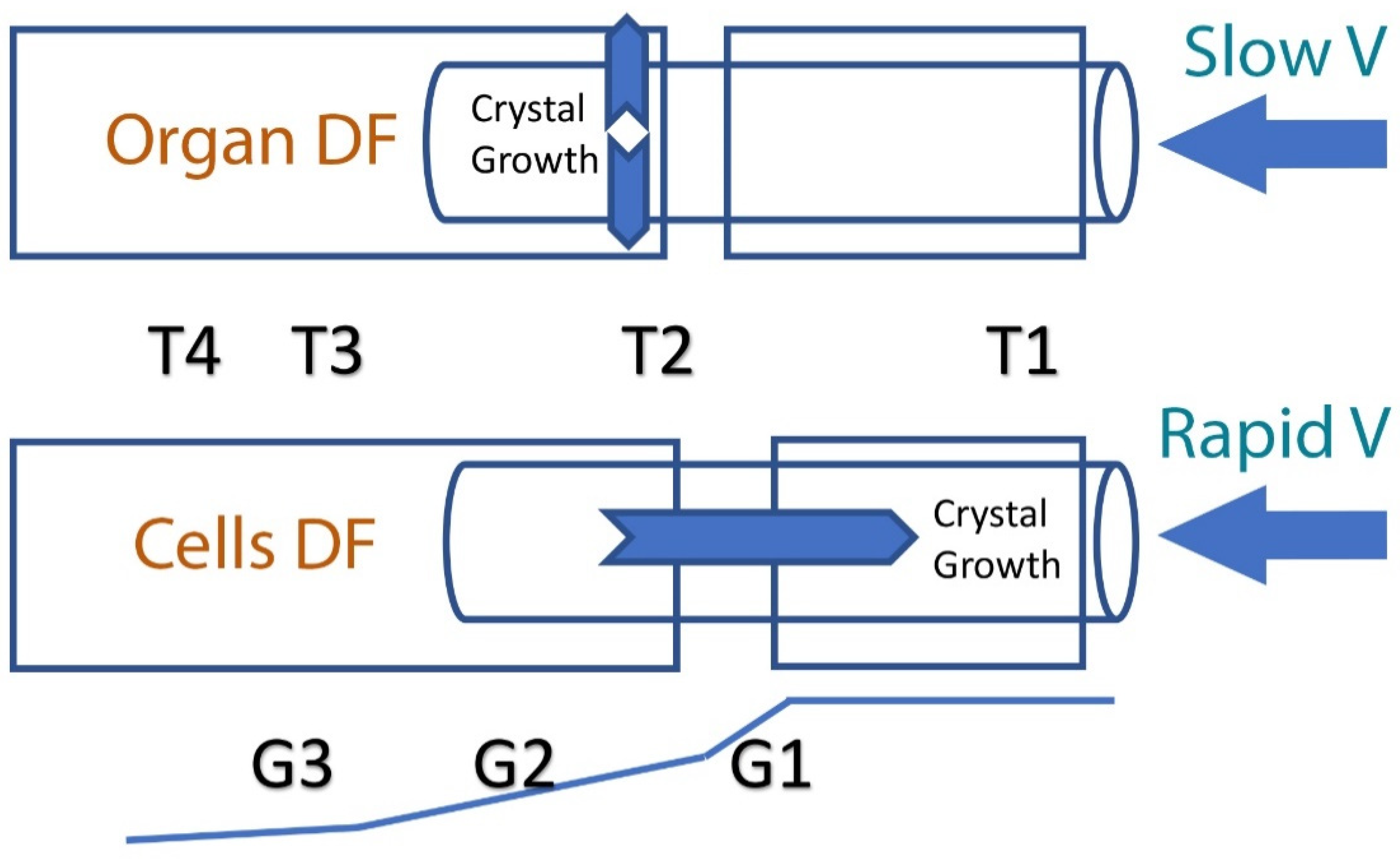
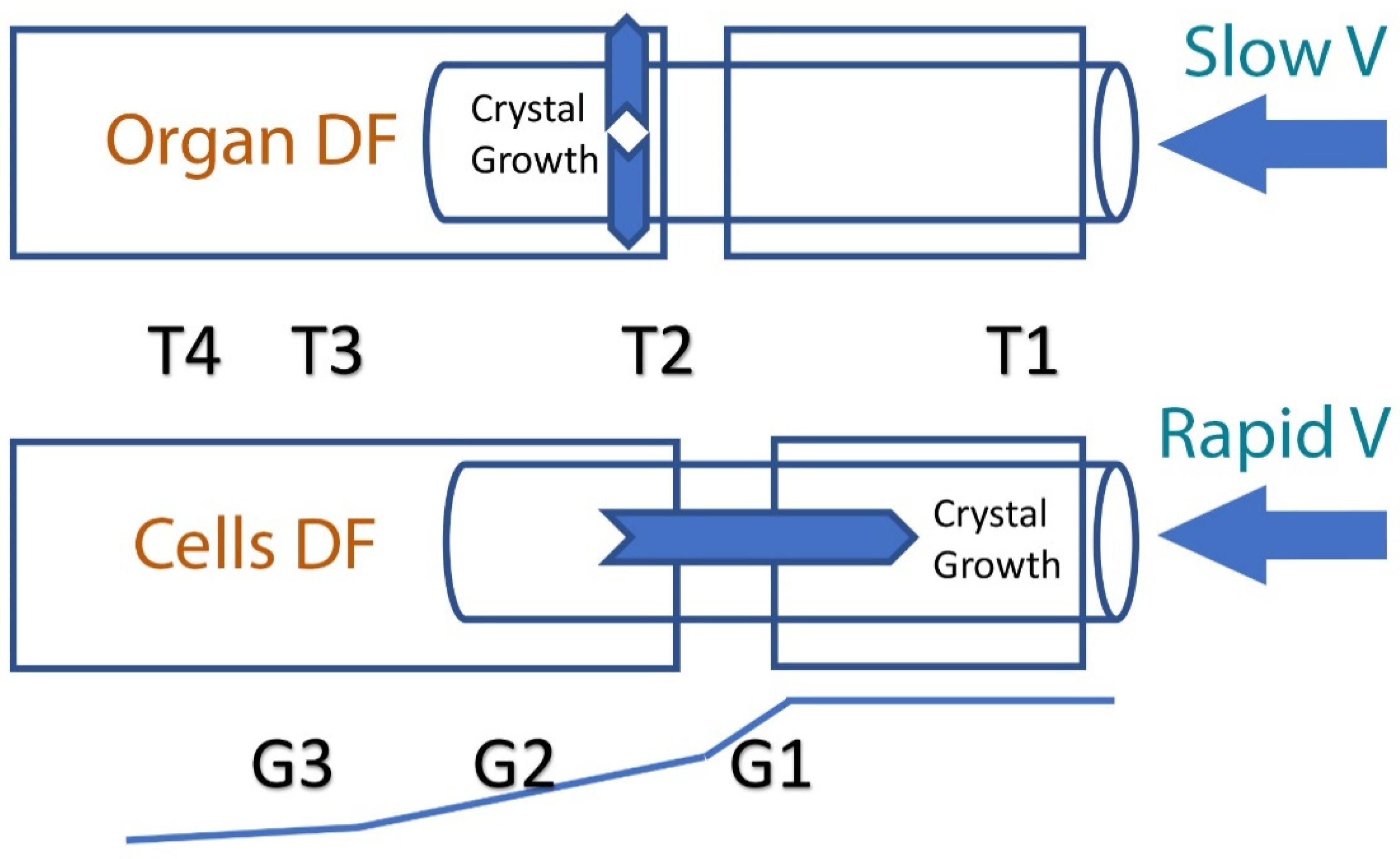
2. Principles of Directional Freezing (DF) and the Effect of Ice Crystal Propagation Velocity
3. The Evolution of Vitrification
Modern Vitrification
4. Factors Effecting Oocyte and Embryo Vitrification
5. Limitations and Challenges
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smith, A.U. Biological effects of freezing and supercooling. In The Effects of Glycerol and of Freezing on Mammalian Organs; Smith, A.U., Ed.; Edward Arnold: London, UK, 1961; pp. 247–269. [Google Scholar]
- Karlsson, J.O.; Toner, M. Long-term storage of tissues by cryopreservation: Critical issues. Biomaterials 1996, 17, 243–256. [Google Scholar] [CrossRef]
- Fahy, G.M.; Wowk, B. Cryopreservation and freeze-drying protocols. In Methods in Molecular Biology; Springer: London, UK, 2015; Volume 1257, pp. 21–82. [Google Scholar]
- Finger, E.B.; Bischof, J.C. Cryopreservation by vitrification: A promising approach for transplant organ banking. Curr. Opin. Organ Transplant. 2018, 23, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Arav, A. Device and Methods for Multigradient Directional Cooling and Warming of Biological Samples. U.S. Patent 5,873,254, 23 February 1999. [Google Scholar]
- Arav, A.; Revel, A.; Nathan, Y.; Bor, A.; Gacitua, H.; Yavin, S.; Gavish, Z.; Uri, M.; Elami, A. Oocyte recovery, embryo development and ovarian function after cryopreservation and transplantation of whole sheep ovary. Hum. Reprod. 2005, 20, 3554–3559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahari, L.; Bein, A.; Yashunsky, V.; Braslavsky, I. Directional freezing for the cryopreservation of adherent mammalian cells on a substrate. PLoS ONE 2018, 13, e0192265. [Google Scholar] [CrossRef]
- Reid, C.; Hermes, R.; Blottner, S.; Goeritz, F.; Wibbelt, G.; Walzer, C.; Bryant, R.; Portas, T.; Streich, W.; Hildebrandt, T. Split-sample comparison of directional and liquid nitrogen vapour freezing method on post-thaw semen quality in white rhinoceroses (Ceratotherium simum simum and Ceratotherium simum cottoni). Theriogenology 2009, 71, 275–291. [Google Scholar] [CrossRef]
- Robecka, T.; Gearhartb, S.; Steinmana, K.; Katsumatac, E.; Loureirod, J.; O’Brien, J. In vitro sperm characterization and development of a sperm cryopreservation method using directional solidification in the killer whale (Orcinus orca). Theriogenology 2011, 76, 267–279. [Google Scholar] [CrossRef]
- Fahy, G.M. Biological effects of vitrification and devitrification. In The Biophysics of Organ Cryopreservation; NATO ASI Series (Series A: Life Sciences); Pegg, D.E., Karow, A.M., Eds.; Springer: Boston, MA, USA, 1987; Volume 147. [Google Scholar]
- Fahy, G.M.; Wowk, B.; Wu, J.; Phan, J.; Rasch, C.; Chang, A.; Zendejas, E. Cryopreservation of organs by vitrification: Perspectives and recent advances. Cryobiology 2004, 48, 157–178. [Google Scholar] [CrossRef]
- Saragusty, J.; Arav, A. Current progress in oocyte and embryo cryopreservation by slow freezing and vitrification. Reproduction 2011, 141, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Arav, A.; Yavin, S.; Zeron, Y.; Natan, D.; Dekel, I.; Gacitua, H. New trends in gamete’s cryopreservation. Mol. Cell. Endocrinol. 2002, 187, 77–81. [Google Scholar] [CrossRef]
- Maffei, F.; Pennarossa, G.; Brevini, T.; Arav, A.; Gandolfi, F. Beneficial effect of directional freezing on in vitro viability of cryopreserved sheep whole ovaries and ovarian cortical slices. Hum. Reprod. 2014, 29, 114–124. [Google Scholar] [CrossRef]
- Arav, A.; Gavish, Z.; Elami, A.; Natan, Y.; Revel, A.; Silber, S.; Gosden, R.G.; Patrizio, P. Ovarian function 6 years after cryopreservation and transplantation of whole sheep ovaries. Reprod. Biomed. Online 2010, 20, 48–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patrizio, P.; Arav, A. Whole human ovaries cryopreservation using a novel multi-gradient freezing device. Fertil. Steril. 2007, 88, S355. [Google Scholar] [CrossRef]
- Gavish, Z.; Ben-Haim, M.; Arav, A. Cryopreservation of whole murine and porcine livers. Rejuvenation Res. 2008, 11, 765–772. [Google Scholar] [CrossRef]
- Elami, A.; Gavish, Z.; Korach, A.; Houminer, E.; Schneider, A.; Schwalb, H.; Arav, A. Successful restoration of function of frozen and thawed isolated rat hearts. J. Thorac. Cardiovasc. Surg. 2008, 135, 666–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahy, G.M.; Wowk, B.; Pagotan, R.; Chang, A.; Phan, J.; Thomson, B.; Phan, L. Physical and biological aspects of renal vitrification. Organogenesis 2009, 5, 167–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courbiere, B.; Odagescu, V.; Baudot, A.; Massardier, J.; Mazoyer, C.; Salle, B.; Lornage, J. Cryopreservation of the ovary by vitrification as an alternative to slow-cooling protocols. Fertil. Steril. 2006, 86 (Suppl. 4), 1243–1251. [Google Scholar] [CrossRef]
- Arav, A.; Friedman, O.; Natan, Y.; Gur, E.; Shani, N. Rat hindlimb cryopreservation and transplantation: A step toward “organ banking”. Am. J. Transplant. 2017, 17, 2820–2828. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.C.; Khirabadi, B.S.; Lightfoot, F.; Brockbank, K.G.; Taylor, M.J. Vitreous cryopreservation maintains the function of vascular grafts. Nat. Biotechnol. 2000, 18, 296–299. [Google Scholar] [CrossRef]
- Li, Y.; Memon, K.; Zheng, Y.; Cheng, Y.; Mbogba, M.; Wang, P.; Ouyang, X.; Zha, G. Microencapsulation facilitates low-cryoprotectant vitrification of human umbilical vein endothelial cells. ACS Biomater. Sci. Eng. 2019, 5, 5273–5283. [Google Scholar] [CrossRef]
- Armitage, J.; Hall, S.; Routledge, C. Recovery of Endothelial Function after Vitrification of Cornea at −110 °C. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2160–2164. [Google Scholar]
- Brockbank, K.G.M.; Chen, Z.; Greene, E.D.; Campbell, L.H. Vitrification of heart valve tissues. In Cryopreservation and Freeze—Drying Protocols, Methods in Molecular Biology, 4th ed.; Wolkers, W.F., Oldenhof, H., Eds.; Springer: London, UK, 2021; Chapter 31; pp. 593–605. [Google Scholar]
- Taylor, M.J.; Song, Y.C.; Brockbank, K.G. Vitrification in tissue preservation: New developments. In Life in the Frozen State; Fuller, B.J., Lane, N., Benson, E.E., Eds.; CRC Press: Boca Raton, FL, USA, 2004; pp. 603–641. [Google Scholar]
- Sebastian, G.; Lewis, J.; Alvarez, J.; Langer, R.; Roth, A.E.; Church, G.M.; Markmann, J.F.; Sachs, D.H.; Chandraker, A.; Wertheim, J.A.; et al. The promise of organ and tissue preservation to transform medicine. Nat. Biotechnol. 2017, 35, 530–542. [Google Scholar]
- Rubinsky, B.; Ikeda, M. A cryomicroscope using directional solidification for the controlled freezing of biological material. Cryobiology 1985, 22, 55–68. [Google Scholar] [CrossRef]
- Arav, A.; Zeron, Y.; Shturman, H.; Gacitua, H. Successful pregnancies in cows following double freezing of a large volume of semen. Reprod. Nutr. Dev. 2002, 4, 583–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wowk, B. Thermodynamic aspects of vitrification. Cryobiology 2010, 60, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Angell, C.A.; Sichina, W. Thermodynamics of the glass transition: Empirical aspects. Ann. N. Y. Acad. Sci. 1976, 279, 53–67. [Google Scholar] [CrossRef]
- Knight, C.A. The Freezing of Supercooled Liquids; D. Van Nostrand: Princeton, NJ, USA, 1967. [Google Scholar]
- Luyet, B.J.; Gehenio, P.M. Life and Death at Low Temperatures; Biodynamica: Normandy, MO, USA, 1940. [Google Scholar]
- Mousson, A. Einige Tatsachen betreffend das Schmelzen und Gefrieren des Wassers. Ann. Phys. 1858, 105, 161–174. [Google Scholar] [CrossRef] [Green Version]
- Luyet, B.; Rasmussen, D. Study by differential thermal analysis of the temperatures of instability of rapidly cooled solutions of glycerol, ethylene glycol, sucrose, and glucose. Biodynamica 1968, 10, 167–191. [Google Scholar]
- Rasmussen, D.; Luyet, B. Contribution to the establishment of the temperature concentration.curves of homogeneous nucleation in solutions of some cryoprotective agents. Biodynamica 1970, 11, 33–44. [Google Scholar]
- Arav, A. Cryopreservation of oocytes and embryos. Theriogenology 2014, 81, 96–102. [Google Scholar] [CrossRef]
- Rapatz, G.; Luyet, B. Electron microscope study of erythrocytes in rapidly cooled suspensions containing various concentrations of glycerol. Biodynamica 1968, 10, 193–210. [Google Scholar]
- Rall, W.F.; Fahy, G.M. Ice-free cryopreservation of mouse embryos at −196 degrees C by vitrification. Nature 1985, 313, 573–575. [Google Scholar] [CrossRef] [PubMed]
- Fahy, G.M.; MacFarlane, D.R.; Angell, C.A.; Meryman, H.T. Vitrification as an approach to cryopreservation. Cryobiology 1984, 4, 407–426. [Google Scholar] [CrossRef]
- Fahy, G.M.; Rall, W.F. Vitrification: An overview. In Vitrification in Assisted Reproduction: A User’s Manual and Troubleshooting Guide; Liebermann, J., Tucker, M.J., Eds.; Informa Healthcare: London, UK, 2007; pp. 1–20. [Google Scholar]
- Fahy, G.M. Principles of vitrification as a method of cryopreservation in reproductive biology and medicine. In Fertility Preservation: Principles & Practice, 2nd ed.; Donnez, J., Kim, S.S., Eds.; Cambridge University Press: Cambridge, UK, 2019. [Google Scholar]
- Mullen, S.F.; Fahy, G.M. Fundamental aspects of vitrification as a method of reproductive cell, tissue, and organ cryopreservation. In Principles & Practice of Fertility Preservation; Donnez, J., Kim, S., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 145–163. [Google Scholar]
- Taylor, J.; Song, Y.; Kheirabadi, B.; Lightfoot, F.; Brockbank, K. Vitrification fulfills its promise as an approach to reducing freeze-induced injury in a multicellular tissue. Adv. Heat Mass Transf. Biotechnol. 1999, 363, 93–102. [Google Scholar]
- Arav, A.; Gianaroli, L.; Bafaro, G.; Diotalevi, L. A new vitrification technique for 8-cell stage mouse embryos. In Proceeding of the IVF Meeting, Barcelona, Spain; 1987; Volume 373, p. 118. [Google Scholar]
- Arav, A.; Patrizio, P. Automation in oocyte and ovarian tissue vitrification. In Female and Male Fertility Preservation; Grynberg, M., Patrizio, P., Eds.; Springer: Cham, Switzerland, 2022. [Google Scholar] [CrossRef]
- Wowk, B.; Fahy, G.; Ahmedyar, S.; Taylor, M.; Rabin, Y. Vitrification tendency and stability of DP6-based vitrification solutions for complex tissue cryopreservation. Cryobiology 2018, 82, 70–77. [Google Scholar] [CrossRef] [PubMed]
- MacFarlane, D.R.; Forsyth, M.; Barton, C.A. Vitrification and devitrification in cryopreservation. In Advances in Low-Temperature Biology; Elsevier: Amsterdam, The Netherlands, 1991; Volume 1, pp. 221–277. [Google Scholar]
- Pegg, D.E.; Wang, L.; Vaughan, D. Cryopreservation of articular cartilage. Part 3: The liquidus tracking method. Cryobiology 2006, 52, 360–368. [Google Scholar] [CrossRef]
- Zeron, M.; Tomczak, M.; Crowe, J.; Arav, A. The effect of liposomes on thermotropic membrane phase transitions of bovine spermatozoa and oocytes: Implications for reducing chilling sensitivity. Cryobiology 2002, 45, 143–152. [Google Scholar] [CrossRef]
- Ghetler, Y.; Yavin, S.; Shalgi, R.; Arav, A. The effect of chilling on membrane lipid phase transition in human oocytes and zygotes. Hum. Reprod. 2005, 20, 3385–3389. [Google Scholar] [CrossRef] [Green Version]
- Kemp, E.; Clark, P.B.; Anderson, C.K.; Parsons, F.M. Long-term preservation of the kidney. Proc. Eur. Dial. Transplant. Assoc. 1966, 3, 236–240. [Google Scholar]
- Rapatz, G. Resumption of activity in frog hearts after exposure to very low temperatures. Cryobiology 1970, 6, 588. [Google Scholar]
- Rapatz, G. Recovery of activity of frog hearts after exposure to −78 °C. Cryobiology 1972, 9, 322. [Google Scholar] [CrossRef]
- Fahy, G.M. Prospects for vitrification of whole organs. Cryobiology 1981, 18, 617. [Google Scholar] [CrossRef]
- Fahy, G.M.; MacFarlane, D.R.; Angell, C.A. Recent progress toward vitrification of kidneys. Cryobiology 1982, 19, 668–669. [Google Scholar] [CrossRef]
- Luyet, B.; Kroener, C. The temperature of the “glass transition” in aqueous solutions of glycerol and ethylene glycol. Biodynamica 1966, 10, 33–40. [Google Scholar] [PubMed]
- Kroener, C.; Luyet, B. Formation of cracks during the vitrification of glycerol solutions and disappearance of the cracks during rewarming. Biodynamica 1966, 10, 47–52. [Google Scholar]
- Rabin, Y.; Steif, P.S.; Hess, J.L.; Jimenez-Rios, J.L.; Palastro, M.C. Fracture formation in vitrified thin films of cryoprotectants. Cryobiology 2006, 53, 75–95. [Google Scholar] [CrossRef] [Green Version]
- Pegg, D.E.; Wusteman, M.C.; Boylan, S. Fractures in cryopreserved elastic arteries. Cryobiology 1997, 34, 183–192. [Google Scholar] [CrossRef]
- Steif, P.S.; Palastro, M.C.; Rabin, Y. The effect of temperature gradients on stress development during cryopreservation via vitrification. Cell Preserv. Technol. 2007, 5, 104–115. [Google Scholar] [CrossRef] [Green Version]
- Rall, W.F.; Reid, D.S.; Farrant, J. Innocuous biological freezing during warming. Nature 1980, 286, 511–514. [Google Scholar] [CrossRef]
- Fahy, G.M.; Saur, J.; Williams, R.J. Physical problems with the vitrification of large biological systems. Cryobiology 1990, 27, 492–510. [Google Scholar] [CrossRef]
- Rall, W.F.; Meyer, T.K. Zona fracture damage and its avoidance during the cryopreservation of mammalian embryos. Theriogenology 1989, 31, 683–692. [Google Scholar] [CrossRef]
- Williams, R.J.; Carnahan, D.L. Fracture faces and other interfaces as ice nucleation sites. Cryobiology 1990, 27, 479–482. [Google Scholar] [CrossRef]
- Arav, A.; Natan, Y. Vitrification of oocytes: From basic science to clinical application. In Oocyte Biology in Fertility Preservation; Advances in Experimental Medicine and Biology; Kim, S., Ed.; Springer: New York, NY, USA, 2013; Volume 761. [Google Scholar] [CrossRef]
- Fahy, G.M. Analysis of “solution effects” injury: Cooling rate dependence of the functional and morphological sequalae of freezing in rabbit renal cortex protected with dimethyl sulfoxide. Cryobiology 1981, 18, 550–570. [Google Scholar] [CrossRef]
- Fahy, G.; Wowk, B.; Wu, J.; Paynter, S. Improved vitrification solutions based on the predictability of vitrification solution toxicity. Cryobiology 2004, 48, 365. [Google Scholar] [CrossRef]
- Arav, A.; Shehu, D.; Mattioli, M. Osmotic and cytotoxic study of vitrification of immature bovine oocytes. J. Reprod. Fertil. 1993, 99, 353–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahy, G.M. Cryoprotectant toxicity: Biochemical or osmotic? Cryo-Letters 1984, 5, 79–90. [Google Scholar]
- Fahy, G.M. Prevention of toxicity from high concentrations of cryoprotective agents. In Organ Preservation, Basic and Applied Aspects; Pegg, D.E., Jacobsen, I.A., Halasz, N.A., Eds.; MTP Press: Lancaster, UK, 1982; pp. 367–369. [Google Scholar]
- Rubinsky, B.; Lee, H. Freezing of Mammalian Livers with Glycerol and Antifreeze Proteins. Biochem. Biophys. Res. Commun. 1994, 200, 732–741. [Google Scholar] [CrossRef]
- Wowk, B.; Leitl, E.; Rasch, C.M.; Mesbah-Karimi, N.; Harris, S.B.; Fahy, G.M. Vitrification enhancement by synthetic ice blocking agents. Cryobiology 2000, 40, 228–236. [Google Scholar] [CrossRef]
- Wowk, B.; Fahy, G.M. Inhibition of bacterial ice nucleation by polyglycerol polymers. Cryobiology 2002, 44, 14–23. [Google Scholar] [CrossRef]
- Arav, A.; Zeron, Y.; Ocheretny, A. A new device and method for vitrification increases the cooling rate and allows successful cryopreservation of bovine oocytes. Theriogenology 2000, 53, 248. [Google Scholar]
- Steponkus, P.; Myers, S.; Lynch, D.; Gardner, L.; Bronshteyn, V.; Leibo, S.; Rall, W.; Pitt, R.; Lint, T.; MacIntyre, R. Cryopreservation of Drosophila melanogaster embryos. Nature 1990, 345, 170–172. [Google Scholar] [CrossRef]
- Mazur, P.; Cole, K.W.; Hall, J.W.; Schreuders, P.D.; Mahowald, A.P. Cryobiological preservation of drosophila embryos. Science 1992, 258, 1896–1897. [Google Scholar] [CrossRef] [PubMed]
- Arav, A.; Natan, D. Freeze Drying of Red Blood Cells: The Use of Directional Freezing and a New Radio Frequency Lyophilization Device. Biopreserv. Biobank. 2012, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Etheridge, M.; Xu, Y.; Rott, L.; Choi, J.; Glasmacher, B.; Bischof, J. RF heating of magnetic nanoparticles improves the thawing of cryopreserved biomaterials. Technology 2014, 2, 229–242. [Google Scholar] [CrossRef]
- Chiu-Lam, A.; Staples, E.; Pepine, C.; Rinaldi, C. Perfusion, cryopreservation, and nano-warming of whole hearts using colloidally stable magnetic cryopreservation agent solutions. Sci. Adv. 2021, 7, eabe3005. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, D.; Bischof, J.; Rabin, Y. Thermomechanical stress in cryopreservation via vitrification with nanoparticle heating as a stress-moderating effect. J. Biomech. Eng. 2015, 138. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, G.; Zhang, Z.; Xu, X.; He, X. Magnetic induction heating of superparamagnetic nanoparticles during rewarming augments the recovery of hUCM-MSCs cryopreserved by vitrification. Acta Biomater. 2016, 33, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Ring, H.; Sharma, A.; Namsrai, B.; Tran, N.; Garwood, M.; Haynes, C.; Bischof, J. Preparation of scalable silica-coated iron oxide nanoparticles for nanowarming. Adv. Sci. 2020, 7, 1901624. [Google Scholar] [CrossRef] [Green Version]
- Ring, H.; Gao, Z.; Sharma, A.; Han, Z.; Lee, C.; Brockbank, K.; Greene, E.; Helke, K.; Chen, L.; Campbell, B.; et al. Imaging the distribution of iron oxide nanoparticle in hypothermic perfused tissues. Magn. Reson. Med. 2020, 83, 1750–1759. [Google Scholar] [CrossRef]
- Arav, A.; Zeron, Y. Vitrification of bovine oocytes using modified minimum drop size technique (MDS) is affected by the composition and the concentration of the vitrification solution and by the cooling conditions. Theriogenology 1997, 47, 341. [Google Scholar] [CrossRef]
- Arav, A. Vitrification of oocyte and embryos. In New Trends in Embryo Transfer; Lauria, A., Gandofi, F., Eds.; Portland Press: Cambridge, UK, 1992; pp. 255–264. [Google Scholar]
- Arav, A. Vitrification of Oocytes and Embryos. DVM Thesis, Bologna University, Bologna, Italy, 1989. [Google Scholar]
- Kuwayama, M.; Vajta, G.; Kato, O.; Leibo, S.P. Highly efficient vitrification method for cryopreservation of human oocytes. Reprod. Biomed. Online 2005, 11, 300–308. [Google Scholar] [CrossRef]
- Vajta, G.; Kuwayama, M.; Vanderzwalmen, P. Disadvantages and benefits of vitrification. In Vitrification in Assisted Reproduction, A User’s Manual and Trouble-Shooting Guide; Tucker, M.J., Liebermann, J., Eds.; Informa: London, UK, 2007; pp. 33–44. [Google Scholar]
- Bielanski, A.; Nadin-Davis, S.; Sapp, T.; Lutze Wallace, C. Viral contamination of embryos cryopreserved in liquid nitrogen. Cryobiology 2000, 40, 110–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bielanski, A. A review of the risk of contamination of semen and embryos during cryopreservation and measures to limit cross-contamination during banking to prevent disease transmission in ET practices. Theriogenology 2012, 77, 467–482. [Google Scholar] [CrossRef] [PubMed]
- Arav, A.; Natan, Y.; Levi-Setti, P.E.; Menduni, F.; Patrizio, P. New methods for cooling and storing oocytes and embryos in a clean environment of −196 °C. Reprod. Biomed. Online 2016, 33, 71–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobo, A.; Domingo, J.; Pérez, S.; Crespo, J.; Remohí, J.; Pellicer, A. Vitrification: An effective new approach to oocyte banking and preserving fertility in cancer patients. Clin. Transl. Oncol. 2008, 10, 268–273. [Google Scholar] [CrossRef]
- Karlsson, J.O.; Cravalho, E.G.; Borel Rinkes, I.H.; Tompkins, R.G.; Yarmush, M.L.; Toner, M. Nucleation and growth of ice crystals inside cultured hepatocytes during freezing in the presence of dimethyl sulfoxide. Biophys. J. 1993, 65, 2524–2536. [Google Scholar] [CrossRef] [Green Version]
- Adrian, M.; Dubochet, J.; Lepault, J.; McDowall, A.W. Cryo-electron microscopy of viruses. Nature 1984, 308, 32–36. [Google Scholar] [CrossRef]
- Coger, R.; Rubinsky, B.; Pegg, D.E. Dependence of probability of vitrification on time and volume. Cryo-Letters 1990, 11, 359–372. [Google Scholar]
- Turnbull, D. On the relation between crystallization rate and liquid structure. J. Phys. Chem. 1962, 66, 609–613. [Google Scholar] [CrossRef]
- Turnbull, D. Under what conditions can a glass be formed? Contemp Phys. 1969, 10, 473–488. [Google Scholar] [CrossRef]
- Mazur, P. Equilibrium, quasi-equilibrium, and nonequilibrium freezing of mammalian embryos. Cell Biophys. 1990, 17, 53–92. [Google Scholar] [CrossRef]
- Karlsson, J.M. Effects of solution composition on the theoretical prediction of ice nucleation kinetics and thermodynamics. Cryobiology 2010, 60, 43–51. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arav, A. Cryopreservation by Directional Freezing and Vitrification Focusing on Large Tissues and Organs. Cells 2022, 11, 1072. https://doi.org/10.3390/cells11071072
Arav A. Cryopreservation by Directional Freezing and Vitrification Focusing on Large Tissues and Organs. Cells. 2022; 11(7):1072. https://doi.org/10.3390/cells11071072
Chicago/Turabian StyleArav, Amir. 2022. "Cryopreservation by Directional Freezing and Vitrification Focusing on Large Tissues and Organs" Cells 11, no. 7: 1072. https://doi.org/10.3390/cells11071072
APA StyleArav, A. (2022). Cryopreservation by Directional Freezing and Vitrification Focusing on Large Tissues and Organs. Cells, 11(7), 1072. https://doi.org/10.3390/cells11071072