
The Nephrotoxin Puromycin Aminonucleoside Induces Injury in Kidney Organoids Differentiated from Induced Pluripotent Stem Cells




Abstract
1. Introduction
2. Materials and Methods
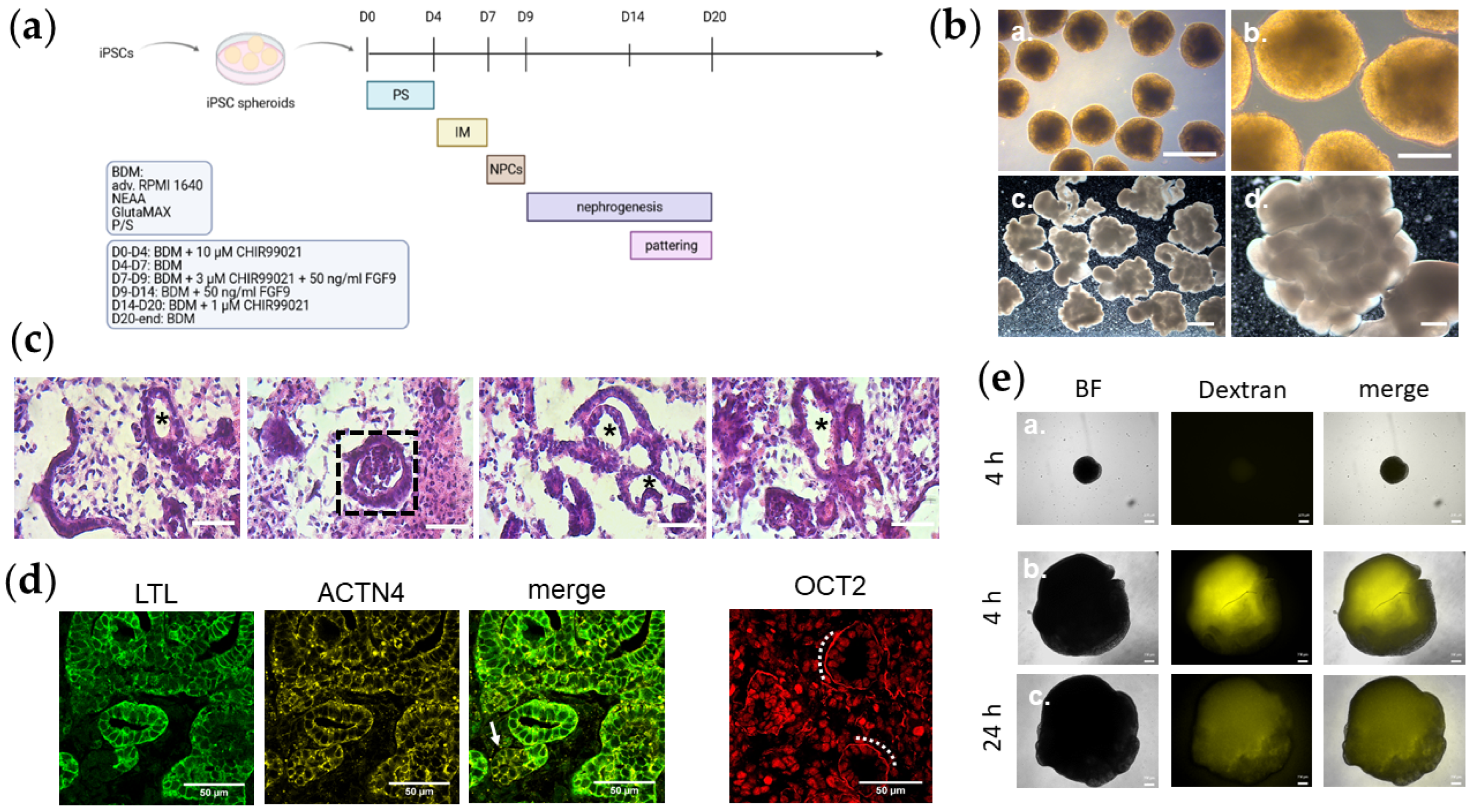
2.1. Cell Cultivation and Formation of Kidney Organoids
2.2. Cryosectioning
2.3. Western Blotting
2.4. Immunocytochemistry
2.5. Immunohistochemistry and Histology
2.6. Fluorometric Renin Assay
2.7. In Vitro Dextran Uptake Assay
2.8. qRT-PCR
2.9. Analysis of Gene Expression Data
2.10. Over-Representation Analysis of Pathways and Gene Ontologies (GOs)
2.11. Metascape Analysis
2.12. Kidney-Associated Cytokine Assay
2.13. Image and Data Analysis of the Kidney Cytokine Assay
3. Results
3.1. Kidney Organoids Possess Structured Lobes with Distinct Tubular and Basement Membrane Structures
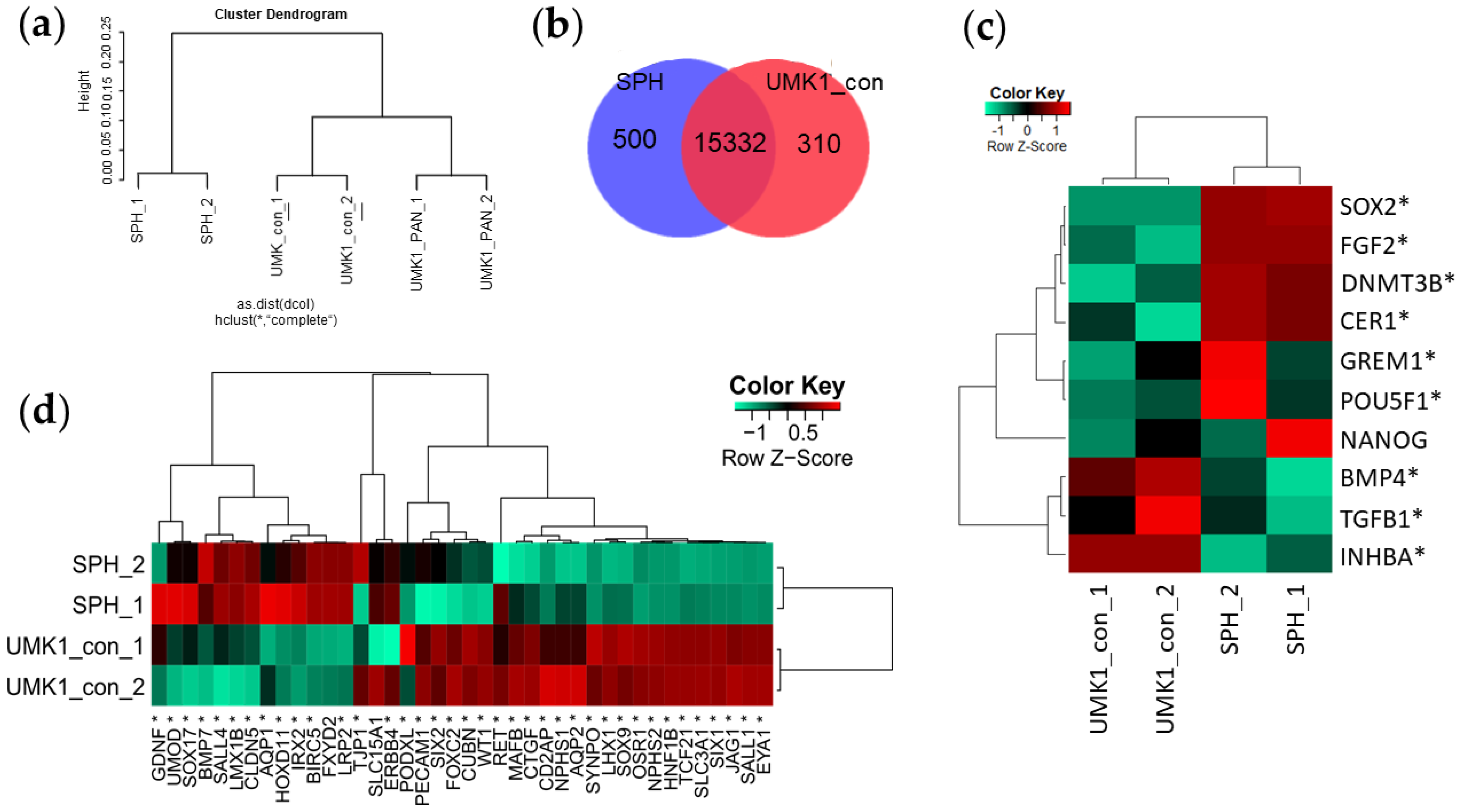
3.2. iPSC Spheroids Lose Pluripotent Gene Expression and Gain Kidney-Associated Genes during Differentiation
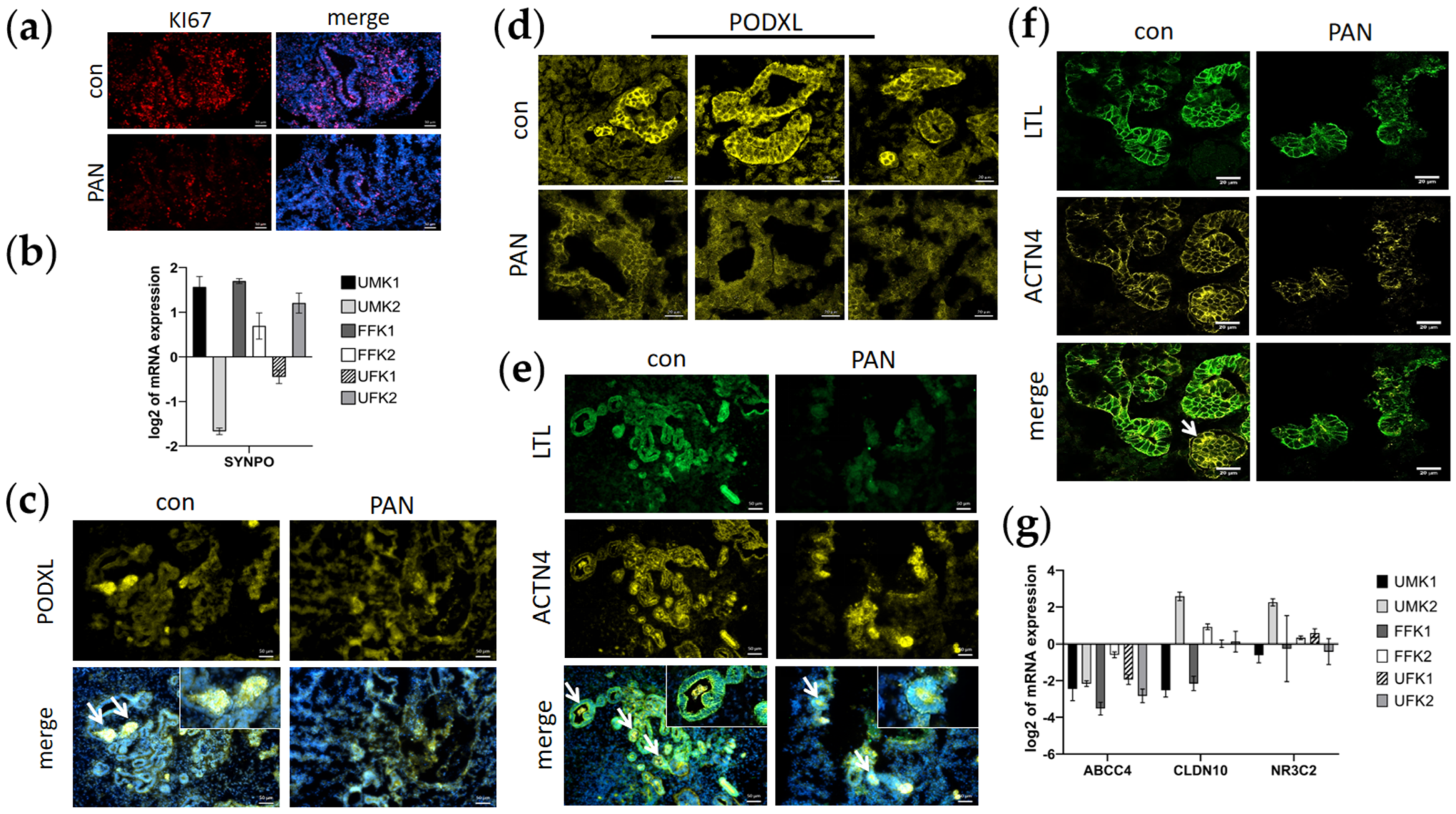
3.3. PAN Negatively Affects Podocytes and Partly Tubular Cells
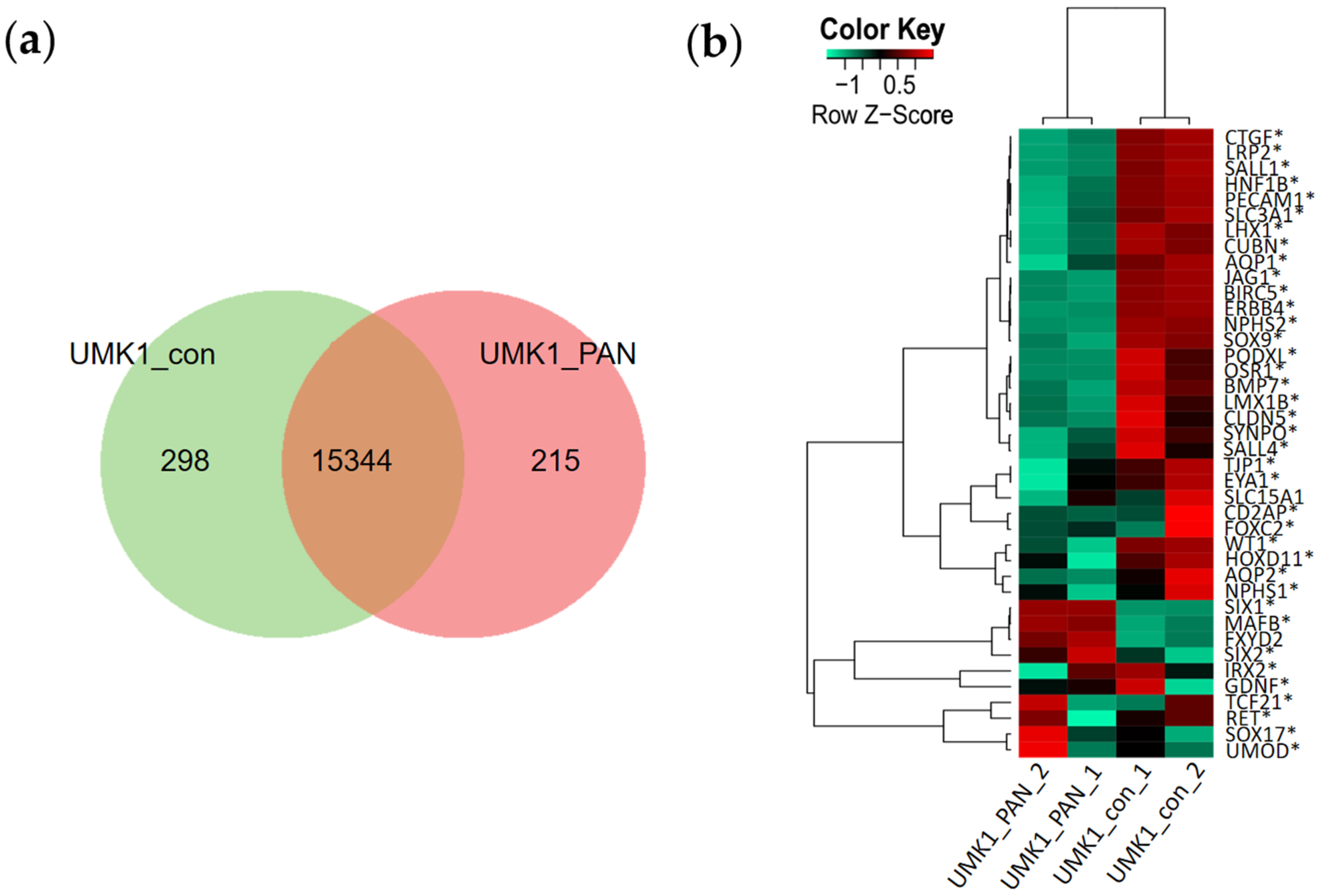
3.4. Transcriptome Analysis Reveal Kidney-Related GO Terms Are Expressed in Urine-Stem-Cell-Derived iPSC Spheroids and Kidney Organoids
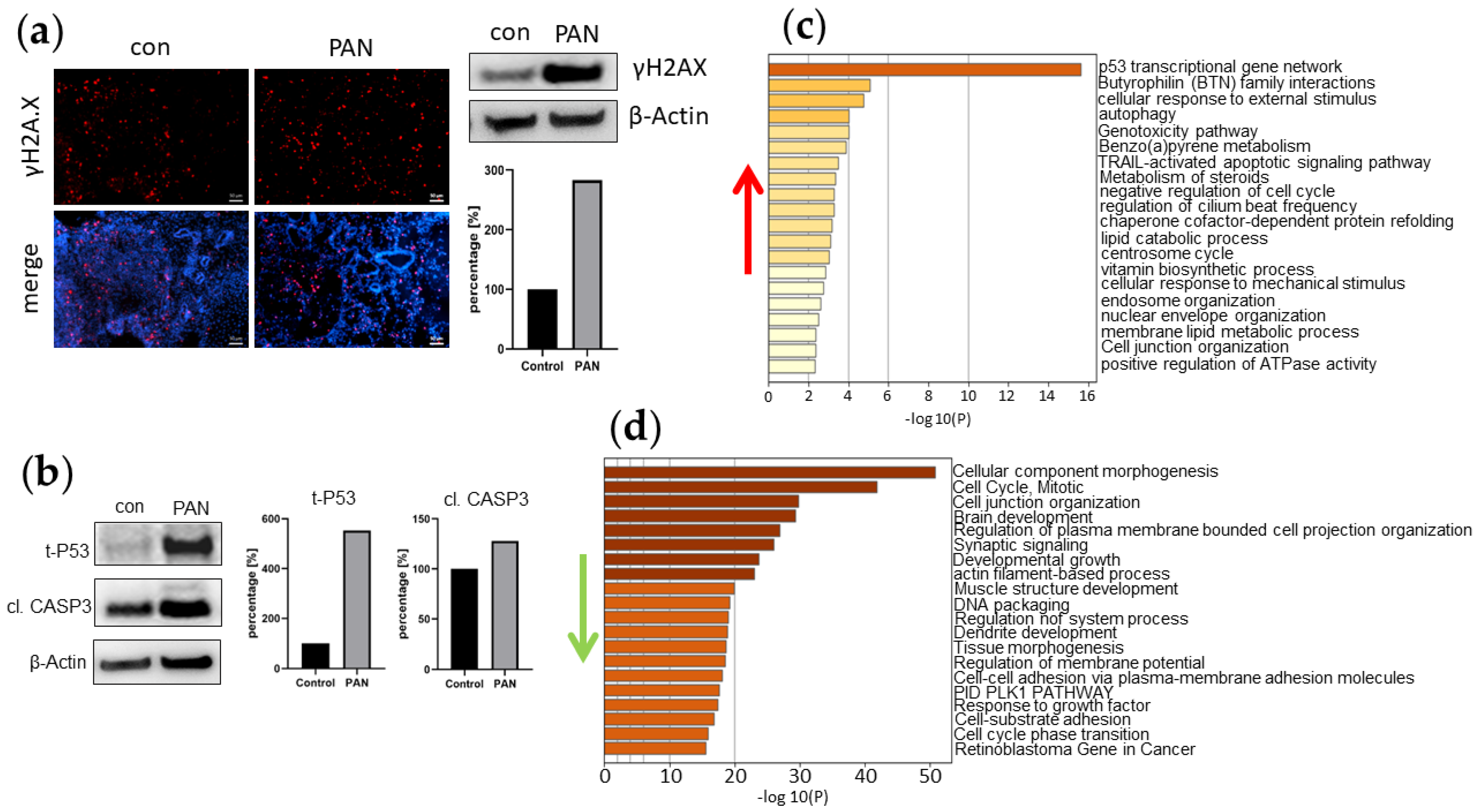
3.5. PAN Induces DNA Damage in Kidney Organoids
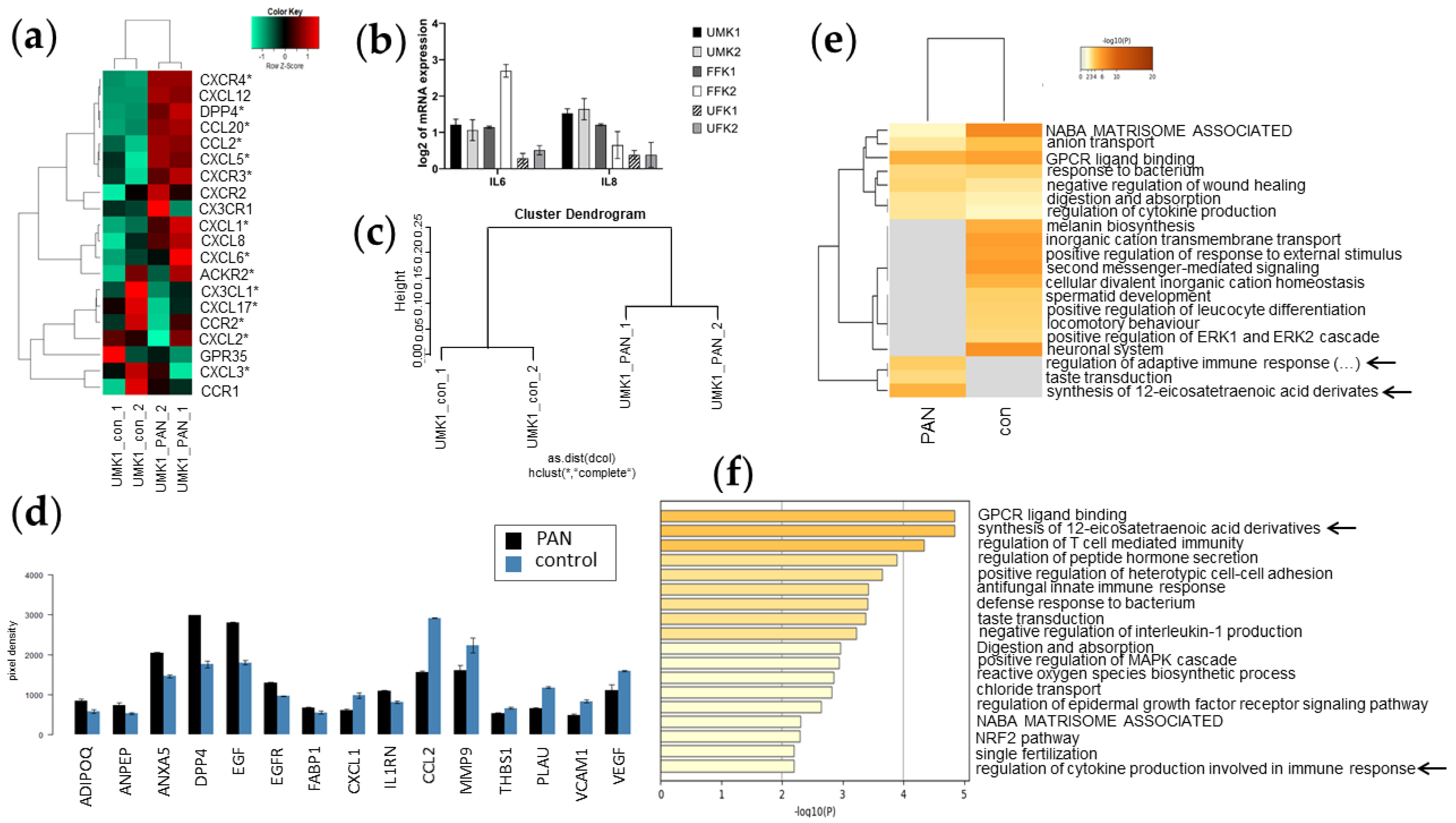
3.6. Inflammation-Associated Processes Are Elevated in PAN-Induced Kidney Organoids
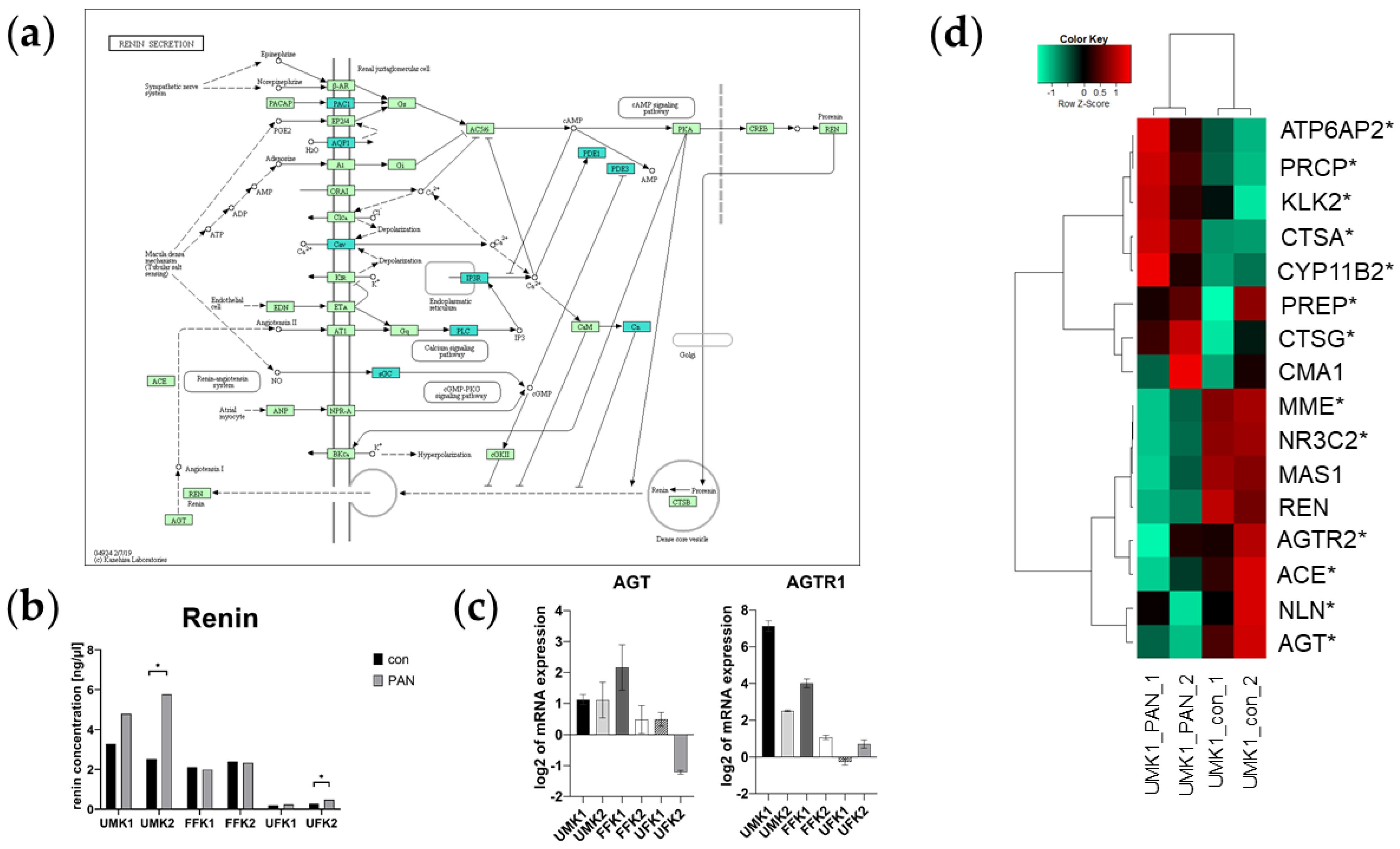
3.7. PAN Activates the Renin–Aldosterone–Angiotensin System (RAAS)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Gong, L.; Pan, Q.; Yang, N. Autophagy and Inflammation Regulation in Acute Kidney Injury. Front. Physiol. 2020, 11, 1170. [Google Scholar] [CrossRef]
- Webster, A.C.; Nagler, E.V.; Morton, R.L.; Masson, P. Chronic Kidney Disease. Lancet. 2017, 389, 1238–1252. [Google Scholar] [CrossRef]
- Mihai, S.; Codrici, E.; Popescu, I.D.; Enciu, A.M.; Albulescu, L.; Necula, L.G.; Mambet, C.; Anton, G.; Tanase, C. Inflammation-Related Mechanisms in Chronic Kidney Disease Prediction, Progression, and Outcome. J. Immunol. Res. 2018, 2018, 2180373. [Google Scholar] [CrossRef]
- Levey, A.S.; Levin, A.; Kellum, J.A. Definition and Classification of Kidney Diseases. Am. J. Kidney Dis. 2013, 61, 686–688. [Google Scholar] [CrossRef]
- Zhou, Y.; Kim, C.; Lorenzo, J.; Pablo, B.; Zhang, F.; Jung, J.Y.; Xiao, L.; Bazua, S.; Emani, M.; Hopkins, C.; et al. Blocking the Rac1-TRPC5 Pathway Protects Human Podocytes. bioRxiv. 2021. [Google Scholar] [CrossRef]
- Dworkin, L.D.; Feiner, H.D.; Randazzo, J. Glomerular Hypertension and Injury in Desoxycorticosterone-Salt Rats on Antihypertensive Therapy. Kidney Int. 1987, 31, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Bikbov, B.; Perico, N.; Abbate, M.; Remuzzi, G. The Glomerulus: Mechanisms and Patterns of Injury☆. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar] [CrossRef]
- Kang, Y.L.; Saleem, M.A.; Chan, K.W.; Yung, B.Y.M.; Law, H.K.W. The Cytoprotective Role of Autophagy in Puromycin Aminonucleoside Treated Human Podocytes. Biochem. Biophys. Res. Commun. 2014, 443, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Bertram, J.F.; Messina, A.; Ryan, G.B. In Vitro Effects of Puromycin Aminonucleoside on the Ultrastructure of Rat Glomerular Podocytes. Cell Tissue Res. 1990, 260, 555–563. [Google Scholar] [CrossRef]
- Bao, Y.W.; Yuan, Y.; Chen, J.H.; Lin, W.Q. Kidney Disease Models: Tools to Identify Mechanisms and Potential Therapeutic Targets. Zool. Res. 2018, 39, 72–86. [Google Scholar] [CrossRef]
- Rabe, M.; Schaefer, F. Non-Transgenic Mouse Models of Kidney Disease. Nephron 2016, 133, 53–61. [Google Scholar] [CrossRef]
- Romero-Guevara, R.; Ioannides, A.; Xinaris, C. Kidney Organoids as Disease Models: Strengths, Weaknesses and Perspectives. Front. Physiol. 2020, 11, 1384. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.; Sun, G.; Liu, S.; Peng, E.; Wan, M.; Chen, L.; Jackson, J.; Atala, A. Three-Dimensional Renal Organoids from Whole Kidney Cells: Generation, Optimization, and Potential Application in Nephrotoxicology in Vitro. Cell Transplant. 2020, 29, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Schutgens, F.; Rookmaaker, M.B.; Margaritis, T.; Rios, A.; Ammerlaan, C.; Jansen, J.; Gijzen, L.; Vormann, M.; Vonk, A.; Viveen, M.; et al. Tubuloids Derived from Human Adult Kidney and Urine for Personalized Disease Modeling. Nat. Biotechnol. 2019, 37, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.; Spitzhorn, L.S.; Adjaye, J. Constructing an Isogenic 3D Human Nephrogenic Progenitor Cell Model Composed of Endothelial, Mesenchymal, and Six2-Positive Renal Progenitor Cells. Stem Cells Int. 2019, 2019, 3298432. [Google Scholar] [CrossRef]
- Bohndorf, M.; Ncube, A.; Spitzhorn, L.-S.; Enczmann, J.; Wruck, W.; Adjaye, J. Derivation and Characterization of Integration-Free IPSC Line ISRM-UM51 Derived from SIX2-Positive Renal Cells Isolated from Urine of an African Male Expressing the CYP2D6 *4/*17 Variant Which Confers Intermediate Drug Metabolizing Activity. Stem Cell Res. 2017, 25, 18–21. [Google Scholar] [CrossRef]
- Rahman, M.S.; Wruck, W.; Spitzhorn, L.S.; Nguyen, L.; Bohndorf, M.; Martins, S.; Asar, F.; Ncube, A.; Erichsen, L.; Graffmann, N.; et al. The FGF, TGFβ and WNT Axis Modulate Self-Renewal of Human SIX2+ Urine Derived Renal Progenitor Cells. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Wang, Y.; Adjaye, J. A Cyclic AMP Analog, 8-Br-CAMP, Enhances the Induction of Pluripotency in Human Fibroblast Cells. Stem Cell Rev. Rep. 2011, 7, 331–341. [Google Scholar] [CrossRef]
- Low, J.H.; Li, P.; Guo, E.G.Y.C.; Zhou, B.; Suzuki, K.; Zhang, T.; Lian, M.M.; Liu, M.; Aizawa, E.; Esteban, C.R.; et al. Generation of Human PSC-Derived Kidney Organoids with Patterned Nephron Segments and a De Novo Vascular Network Article Generation of Human PSC-Derived Kidney Organoids with Patterned Nephron Segments and a De Novo Vascular Network. Cell Stem Cell 2019, 25, 373–387. [Google Scholar] [CrossRef]
- Carvalho, B.S.; Irizarry, R.A. A Framework for Oligonucleotide Microarray Preprocessing. Bioinformatics 2010, 26, 2363–2367. [Google Scholar] [CrossRef]
- Chen, H.; Boutros, P.C. VennDiagram: A Package for the Generation of Highly-Customizable Venn and Euler Diagrams in R. BMC Bioinform. 2011, 12, 35. [Google Scholar] [CrossRef]
- Graffmann, N.; Ring, S.; Kawala, M.-A.; Wruck, W.; Ncube, A.; Trompeter, H.-I.; Adjaye, J. Modeling Nonalcoholic Fatty Liver Disease with Human Pluripotent Stem Cell-Derived Immature Hepatocyte-Like Cells Reveals Activation of PLIN2 and Confirms Regulatory Functions of Peroxisome Proliferator-Activated Receptor Alpha. Stem Cells Dev. 2016, 25, 1119–1133. [Google Scholar] [CrossRef] [PubMed]
- Warnes, G.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Mächler, M.; Magnusson, A.; Möller, S. Gplots: Various R Programming Tools for Plotting Data, v. 2.6.0; 2005. Available online: https://github.com/talgalili/gplots (accessed on 9 January 2021).
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New Perspectives on Genomes, Pathways, Diseases and Drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef] [PubMed]
- Falcon, S.; Gentleman, R. Using GOstats to Test Gene Lists for GO Term Association. Bioinformatics 2007, 23, 257–258. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape Provides a Biologist-Oriented Resource for the Analysis of Systems-Level Datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S.; Ellis, B.; Gautier, L.; Ge, Y.; Gentry, J.; et al. Bioconductor: Open Software Development for Computational Biology and Bioinformatics. Genome Biol. 2004, 5, R80. [Google Scholar] [CrossRef]
- Du, P.; Kibbe, W.A.; Lin, S.M. Lumi: A Pipeline for Processing Illumina Microarray. Bioinformatics 2008, 24, 1547–1548. [Google Scholar] [CrossRef]
- Erichsen, L.; Bohndorf, M.; Rahman, M.S.; Wruck, W.; Adjaye, J. Angiotensin II Disrupts the Cytoskeletal Architecture of Human Urine-Derived Podocytes and Results in Activation of the Renin-Angiotensin System. bioRxiv 2021. [Google Scholar] [CrossRef]
- Takasato, M.; Er, P.X.; Chiu, H.S.; Maier, B.; Baillie, G.J.; Ferguson, C.; Parton, R.G.; Wolvetang, E.J.; Roost, M.S.; Chuva de Sousa Lopes, S.M.; et al. Kidney Organoids from Human IPS Cells Contain Multiple Lineages and Model Human Nephrogenesis. Nature 2015, 526, 564–568. [Google Scholar] [CrossRef]
- Refaeli, I.; Hughes, M.R.; Wong, A.K.W.; Bissonnette, M.L.Z.; Roskelley, C.D.; Wayne Vogl, A.; Barbour, S.J.; Freedman, B.S.; McNagny, K.M. Distinct Functional Requirements for Podocalyxin in Immature and Mature Podocytes Reveal Mechanisms of Human Kidney Disease. Sci. Rep. 2020, 10, 9419. [Google Scholar] [CrossRef]
- Dandapani, S.V.; Sugimoto, H.; Matthews, B.D.; Kolb, R.J.; Sinha, S.; Gerszten, R.E.; Zhou, J.; Ingber, D.E.; Kalluri, R.; Pollak, M.R. α-Actinin-4 Is Required for Normal Podocyte Adhesion. J. Biol. Chem. 2007, 282, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Luimula, P.; Sandström, N.; Novikov, D.; Holthöfer, H. Podocyte-Associated Molecules in Puromycin Aminonucleoside Nephrosis of the Rat. Lab. Investig. 2002, 82, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Kwon, T.; Kim, D.J.; Huh, W.; Kim, Y.G.; Oh, H.Y.; Kawachi, H. Ultrastructural Study on Nephrin Expression in Experimental Puromycin Aminonucleoside Nephrosis. Nephrol. Dial. Transplant. 2004, 19, 2981–2986. [Google Scholar] [CrossRef]
- Li, X.; Chuang, P.Y.; D’Agati, V.D.; Dai, Y.; Yacoub, R.; Fu, J.; Xu, J.; Taku, O.; Premsrirut, P.K.; Holzman, L.B.; et al. Nephrin Preserves Podocyte Viability and Glomerular Structure and Function in Adult Kidneys. J. Am. Soc. Nephrol. 2015, 26, 2361–2377. [Google Scholar] [CrossRef]
- Hagen, M.; Pfister, E.; Kosel, A.; Shankland, S.; Pippin, J.; Amann, K.; Daniel, C. Cell Cycle Re-Entry Sensitizes Podocytes to Injury Induced Death. Cell Cycle 2016, 15, 1929–1937. [Google Scholar] [CrossRef] [PubMed]
- Thomasova, D.; Anders, H.-J. Cell Cycle Control in the Kidney. Nephrol. Dial. Transplant. 2015, 30, 1622–1630. [Google Scholar] [CrossRef] [PubMed]
- Garreta, E.; Prado, P.; Tarantino, C.; Oria, R.; Fanlo, L.; Martí, E.; Zalvidea, D.; Trepat, X.; Roca-Cusachs, P.; Gavaldà-Navarro, A.; et al. Fine Tuning the Extracellular Environment Accelerates the Derivation of Kidney Organoids from Human Pluripotent Stem Cells. Nat. Mater. 2019, 18, 397–405. [Google Scholar] [CrossRef]
- Wu, H.; Uchimura, K.; Donnelly, E.; Kirita, Y.; Morris, S.A.; Humphreys, B.D. Comparative Analysis of Kidney Organoid and Adult Human Kidney Single Cell and Single Nucleus Transcriptomes. bioRxiv 2017, 232561. [Google Scholar] [CrossRef]
- Wever, K.E.; Wagener, F.A.D.T.G.; Frielink, C.; Boerman, O.C.; Scheffer, G.J.; Allison, A.; Masereeuw, R.; Rongen, G.A. Diannexin Protects against Renal Ischemia Reperfusion Injury and Targets Phosphatidylserines in Ischemic Tissue. PLoS ONE 2011, 6, e24276. [Google Scholar] [CrossRef]
- Zha, D.; Wu, X.; Gao, P. Adiponectin and Its Receptors in Diabetic Kidney Disease: Molecular Mechanisms and Clinical Potential. Endocrinology 2017, 158, 2022–2034. [Google Scholar] [CrossRef]
- Anders, H.J. Of Inflammasomes and Alarmins: IL-1β and IL-1α in Kidney Disease. J. Am. Soc. Nephrol. 2016, 27, 2564–2575. [Google Scholar] [CrossRef] [PubMed]
- Volarevic, V.; Al-Qahtani, A.; Arsenijevic, N.; Pajovic, S.; Lukic, M.L. Interleukin-1 Receptor Antagonist (IL-1Ra) and IL-1Ra Producing Mesenchymal Stem Cells as Modulators of Diabetogenesis. Autoimmunity 2009, 43, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Panter, S.S.; Faden, A.I. Biochemical Changes and Secondary Injury from Stroke and Trauma. In Principles and Practice of Restorative Neurology; Elsevier: Amsterdam, The Netherlands, 1992; pp. 32–53. [Google Scholar] [CrossRef]
- Blanco, A.; Blanco, G. Biochemical Bases of Endocrinology (II) Hormones and Other Chemical Intermediates. In Medical Biochemistry; Elsevier: Amsterdam, The Netherlands, 2017; pp. 573–644. [Google Scholar] [CrossRef]
- Cecerska-Heryć, E.; Heryć, R.; Wiśniewska, M.; Serwin, N.; Grygorcewicz, B.; Dołęgowska, B. Effect of Renal Replacement Therapy on Selected Arachidonic Acid Derivatives Concentration. BMC Nephrol. 2020, 21, 394. [Google Scholar] [CrossRef] [PubMed]
- Montford, J.R.; Bauer, C.; Dobrinskikh, E.; Hopp, K.; Levi, M.; Weiser-Evans, M.; Nemenoff, R.; Furgeson, S.B. Inhibition of 5-Lipoxygenase Decreases Renal Fibrosis and Progression of Chronic Kidney Disease. Am. J. Physiol.-Ren. Physiol. 2019, 316, F732–F742. [Google Scholar] [CrossRef]
- Van der Hauwaert, C.; Savary, G.; Gnemmi, V.; Glowacki, F.; Pottier, N.; Bouillez, A.; Maboudou, P.; Zini, L.; Leroy, X.; Cauffiez, C.; et al. Isolation and Characterization of a Primary Proximal Tubular Epithelial Cell Model from Human Kidney by CD10/CD13 Double Labeling. PLoS ONE 2013, 8, e66750. [Google Scholar] [CrossRef]
- Greaves, P. Urinary Tract. In Histopathology of Preclinical Toxicity Studies; Elsevier: Amsterdam, The Netherlands, 2012; pp. 537–614. [Google Scholar] [CrossRef]
- Torigoe, K.; Muta, K.; Tsuji, K.; Yamashita, A.; Ota, Y.; Kitamura, M.; Mukae, H.; Nishino, T. Urinary Liver-Type Fatty Acid-Binding Protein Predicts Residual Renal Function Decline in Patients on Peritoneal Dialysis. Med. Sci. Monit. 2020, 26, e928236-1. [Google Scholar] [CrossRef]
- Gerber, C.; Harel, M.; Lynch, M.L.; Herbst, K.W.; Ferrer, F.A.; Shapiro, L.H. Proximal Tubule Proteins Are Significantly Elevated in Bladder Urine of Patients with Ureteropelvic Junction Obstruction and May Represent Novel Biomarkers: A Pilot Study. J. Pediatr. Urol. 2016, 12, e1–e120. [Google Scholar] [CrossRef]
- Yosypiv, I.V. Renin-Angiotensin System in Mammalian Kidney Development. Pediatric Nephrol. 2020, 36, 479–489. [Google Scholar] [CrossRef]
- Hoffmann, N.; Peters, J. Functions of the (pro)Renin Receptor (Atp6ap2) at Molecular and System Levels: Pathological Implications in Hypertension, Renal and Brain Development, Inflammation, and Fibrosis. Pharmacol. Res. 2021, 105922. [Google Scholar] [CrossRef]
- Stefanini, A.C.B.; Da Cunha, B.R.; Henrique, T.; Tajara, E.H. Involvement of Kallikrein-Related Peptidases in Normal and Pathologic Processes. Dis. Markers 2015, 2015, 946572. [Google Scholar] [CrossRef]
- Grobe, N.; Leiva, O.; Morris, M.; Elased, K.M. Loss of Prolyl Carboxypeptidase in Two-Kidney, One-Clip Goldblatt Hypertensive Mice. PLoS ONE 2015, 10, e0117899. [Google Scholar] [CrossRef] [PubMed]
- Cavasin, M.A.; Rhaleb, N.E.; Yang, X.P.; Carretero, O.A. Prolyl Oligopeptidase Is Involved in Release of the Antifibrotic Peptide Ac-SDKP. Hypertension 2004, 43, 1140–1145. [Google Scholar] [CrossRef] [PubMed]
- Kaltenecker, C.C.; Domenig, O.; Kopecky, C.; Antlanger, M.; Poglitsch, M.; Berlakovich, G.; Kain, R.; Stegbauer, J.; Rahman, M.; Hellinger, R.; et al. Critical Role of Neprilysin in Kidney Angiotensin Metabolism. Circ. Res. 2020, 127, 593–606. [Google Scholar] [CrossRef] [PubMed]
- Dilauro, M.; Burns, K.D. Angiotensin-(1-7) and Its Effects in the Kidney. Sci. World J. 2009, 9, 522–535. [Google Scholar] [CrossRef]
- Georgianos, P.I.; Agarwal, R. Mineralocorticoid Receptor Antagonism in Chronic Kidney Disease. Kidney Int. Rep. 2021, 6, 2281–2291. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, A.; Ferro, C.J.; Balafa, O.; Burnier, M.; Ekart, R.; Halimi, J.-M.; Kreutz, R.; Mark, P.B.; Persu, A.; Rossignol, P.; et al. Mineralocorticoid Receptor Antagonists for Nephroprotection and Cardioprotection in Patients with Diabetes Mellitus and Chronic Kidney Disease. Nephrol. Dial. Transplant. 2021, 1–16. [Google Scholar] [CrossRef]
- Belden, Z.; Deiuliis, J.A.; Dobre, M.; Rajagopalan, S. The Role of the Mineralocorticoid Receptor in Inflammation: Focus on Kidney and Vasculature. Am. J. Nephrol. 2017, 46, 298–314. [Google Scholar] [CrossRef]
- Vaidya, A.; Brown, J.M.; Williams, J.S. The Renin-Angiotensin-Aldosterone System and Calcium-Regulatory Hormones. J. Hum. Hypertens. 2015, 29, 515–521. [Google Scholar] [CrossRef]
- Siragy, H.M. The Angiotensin II Type 2 Receptor and the Kidney. JRAAS—J. Renin-Angiotensin-Aldosterone Syst. 2009, 11, 33–36. [Google Scholar] [CrossRef]
- Nishiyama, A.; Kobori, H. Independent Regulation of Renin–Angiotensin–Aldosterone System in the Kidney. Clin. Exp. Nephrol. 2018, 22, 1231–1239. [Google Scholar] [CrossRef]
- Zhu, Y.; Cui, H.; Lv, J.; Liang, H.; Zheng, Y.; Wang, S.; Wang, M.; Wang, H.; Ye, F. AT1 and AT2 Receptors Modulate Renal Tubular Cell Necroptosis in Angiotensin II-Infused Renal Injury Mice. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Gender and Ethnicity | Age | Abbreviation |
|---|---|---|---|
| iPSC spheroids derived from urine cell UM51 iPSCs | Male, African | 51 | iPSC spheroids (SPH) |
| Kidney organoids derived from urine cell UM51 iPSCs, biological replicate 1 | Male, African | 51 | UM51 kidney organoids_1 (UMK1) |
| Kidney organoids derived from urine cell UM51 iPSCs, biological replicate 2 | Male, African | 51 | UM51 kidney organoids_2 (UMK2) |
| Kidney organoids derived from foetal foreskin (FF) iPSCs, biological replicate 1 | Male, Caucasian | foetal | FF kidney organoids_1 (FFK1) |
| Kidney organoids derived from foetal foreskin (FF) iPSCs, biological replicate 2 | Male, Caucasian | foetal | FF kidney organoids_2 (FFK2) |
| Kidney organoids derived from urine cell UF21 iPSCs, biological replicate 1 | Female, Caucasian | 21 | UF21 kidney organoids_1 (UFK1) |
| Kidney organoids derived from urine cell UF21 iPSCs, biological replicate 2 | Female, Caucasian | 21 | UF21 kidney organoids_2 (UFK2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, L.; Wruck, W.; Erichsen, L.; Graffmann, N.; Adjaye, J. The Nephrotoxin Puromycin Aminonucleoside Induces Injury in Kidney Organoids Differentiated from Induced Pluripotent Stem Cells. Cells 2022, 11, 635. https://doi.org/10.3390/cells11040635
Nguyen L, Wruck W, Erichsen L, Graffmann N, Adjaye J. The Nephrotoxin Puromycin Aminonucleoside Induces Injury in Kidney Organoids Differentiated from Induced Pluripotent Stem Cells. Cells. 2022; 11(4):635. https://doi.org/10.3390/cells11040635
Chicago/Turabian StyleNguyen, Lisa, Wasco Wruck, Lars Erichsen, Nina Graffmann, and James Adjaye. 2022. "The Nephrotoxin Puromycin Aminonucleoside Induces Injury in Kidney Organoids Differentiated from Induced Pluripotent Stem Cells" Cells 11, no. 4: 635. https://doi.org/10.3390/cells11040635
APA StyleNguyen, L., Wruck, W., Erichsen, L., Graffmann, N., & Adjaye, J. (2022). The Nephrotoxin Puromycin Aminonucleoside Induces Injury in Kidney Organoids Differentiated from Induced Pluripotent Stem Cells. Cells, 11(4), 635. https://doi.org/10.3390/cells11040635