
The Functional Interaction of EGFR with AT1R or TP in Primary Vascular Smooth Muscle Cells Triggers a Synergistic Regulation of Gene Expression

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Buffers, Chemicals, Antibodies and Primers
2.2. aVSMC Isolation from Mice
2.3. Cell Culture
2.4. RNA Sample Preparation
2.5. RNA Sequencing
2.6. Differential Expression Analysis and Functional Analysis
2.7. Quantitative PCR and Digital Droplet PCR
2.8. Western-Blot
2.9. Caspase-3 Activity
2.10. Senescence-Associated β-Galactosidase Measurement
2.11. ELISA for 5-Bromo-2′-deoxyuidine
2.12. Osteopontin Secretion Measurement
2.13. Statistical Analysis
3. Results
3.1. aVSMC Express Genes Encoding for AT1R, EGFR and TP but Respond to Exclusive EGF and U46619-Stimulation Only
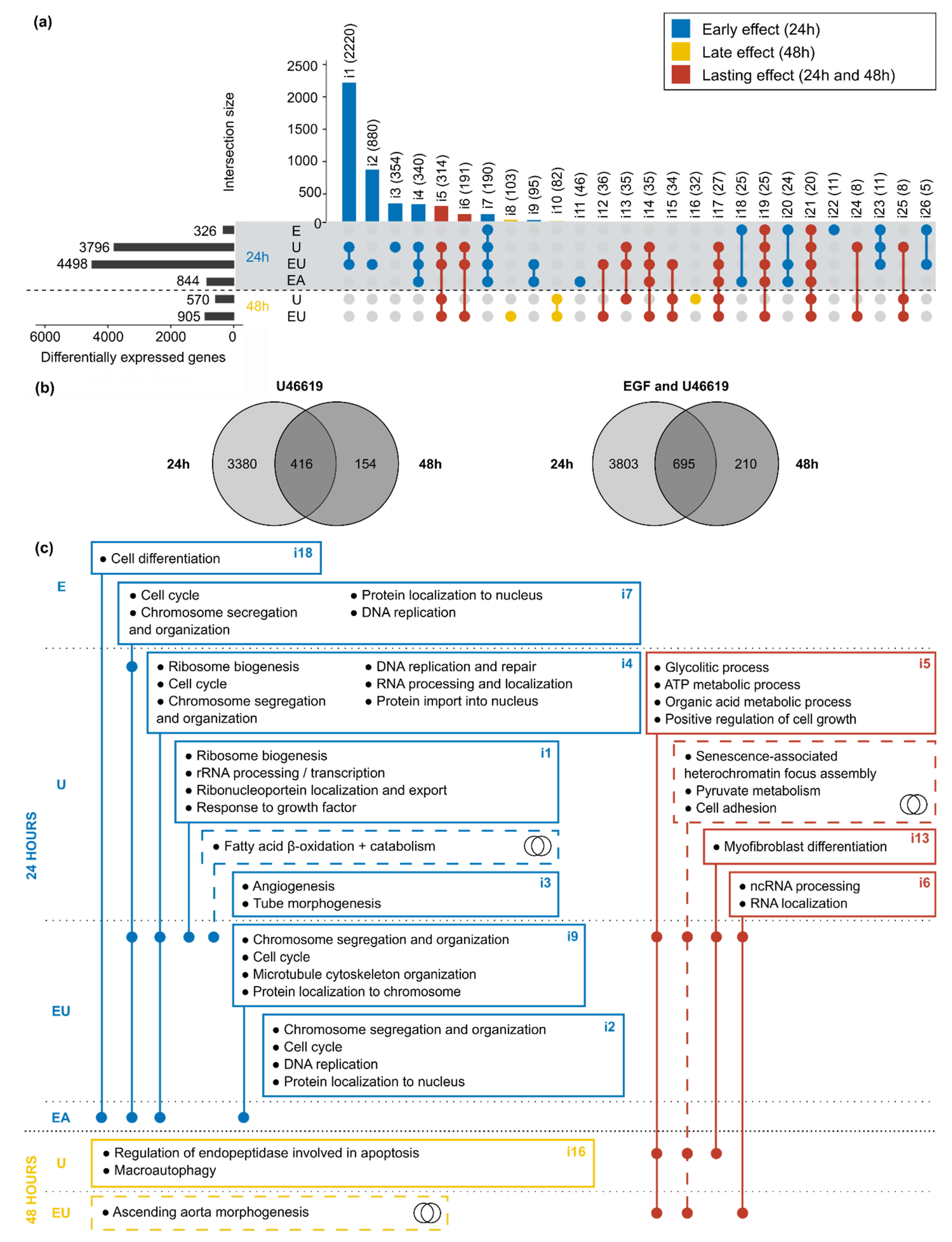
3.2. Exclusive EGFR and TP but Not AT1R-Activation Lead to Time-Dependent Gene Expression Regulation in aVSMC
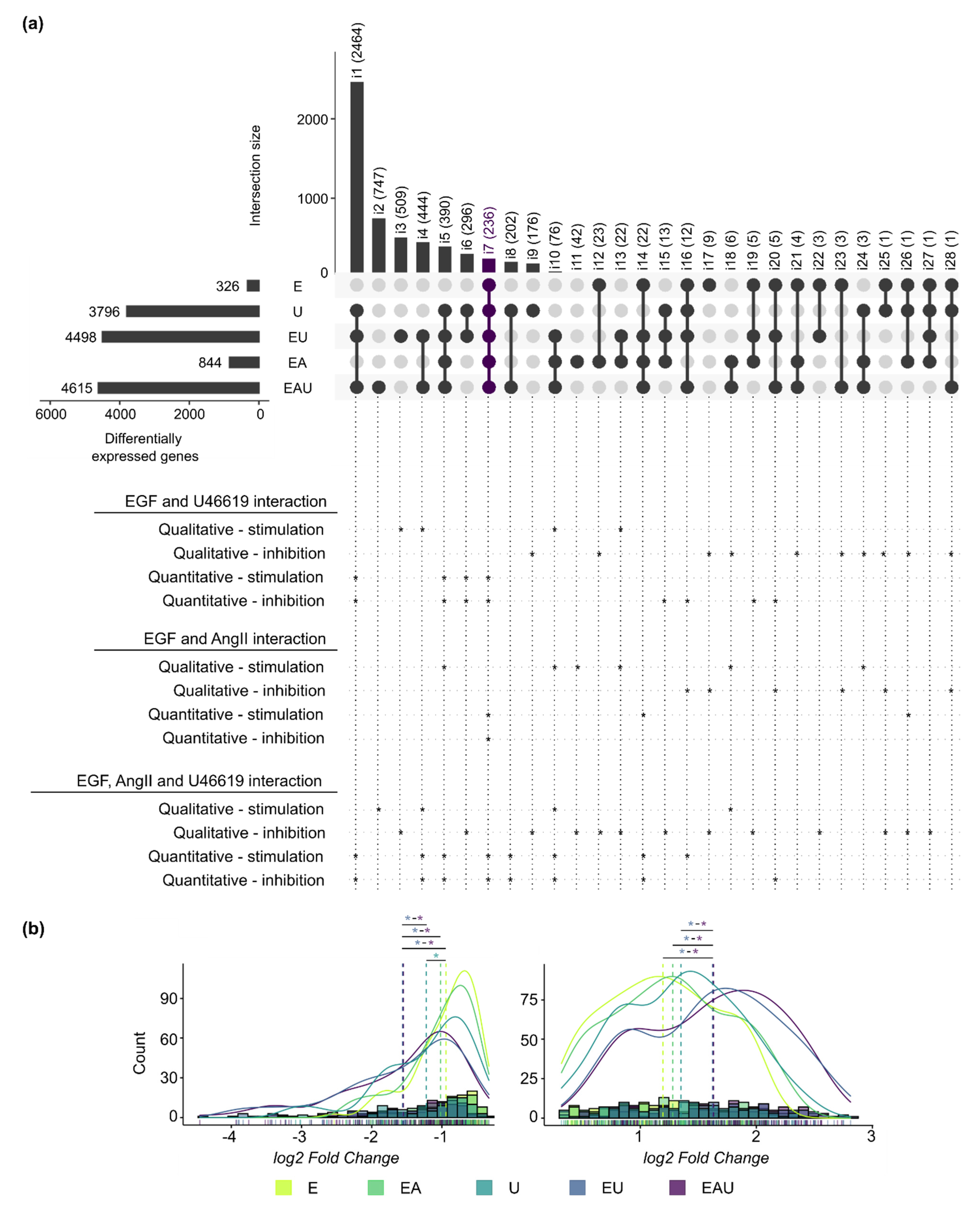
3.3. Simultaneous Receptor Activation Leads to Qualitative and Quantitative Synergistic Gene Expression Regulation
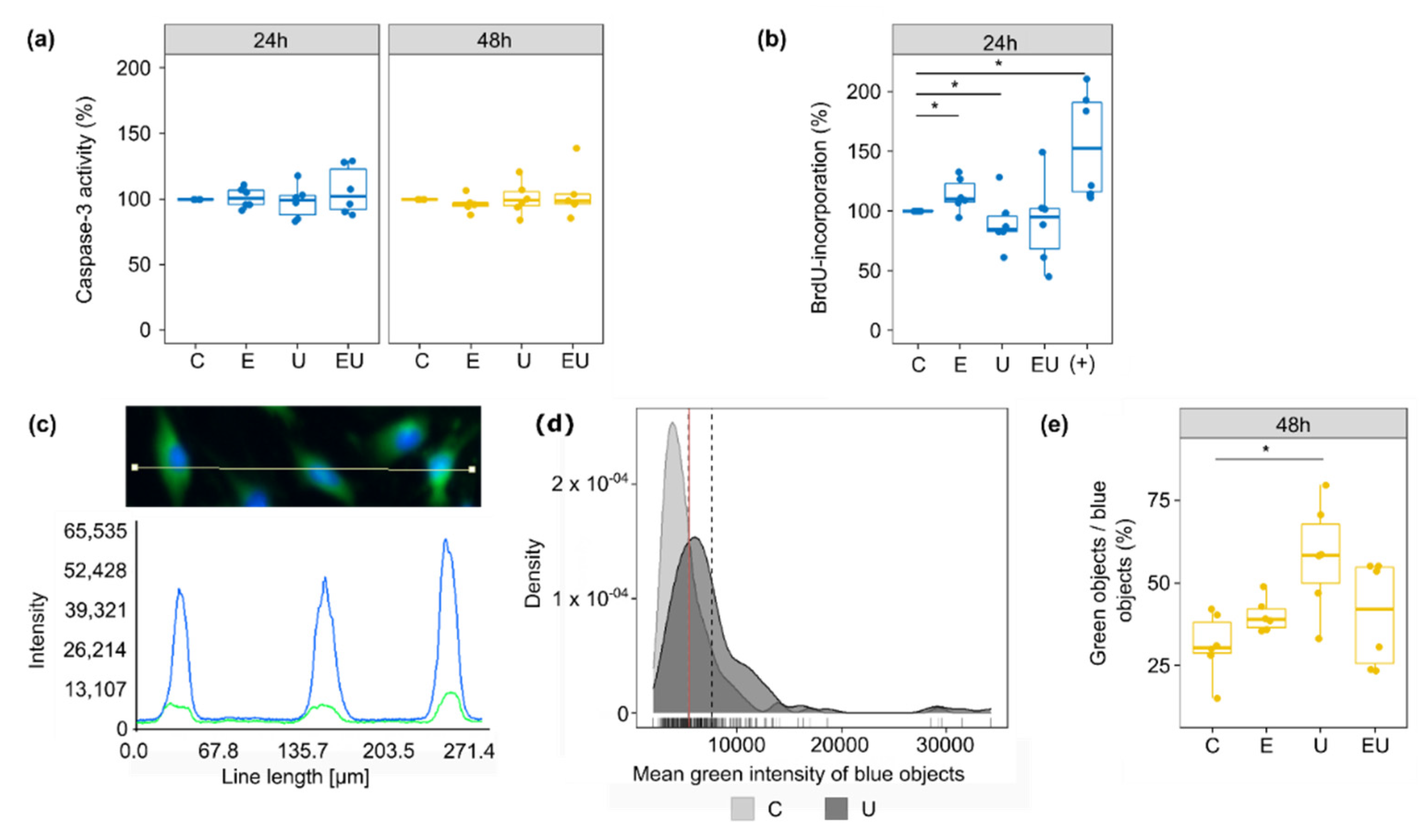
3.4. The Putative Regulators of the EGF and U46619-Induced Changes in Gene Expression Play a Role in Cell Proliferation and Cell Death
3.5. Genes Regulated by EGF and/or U46619 Are Enriched in Pathways Related to Cell Cycle, Cell Death and Metabolism
3.6. Simultaneous EGFR and TP-Activation Regulate Their Respective Effect on Cell Cycle and Cell Senescence
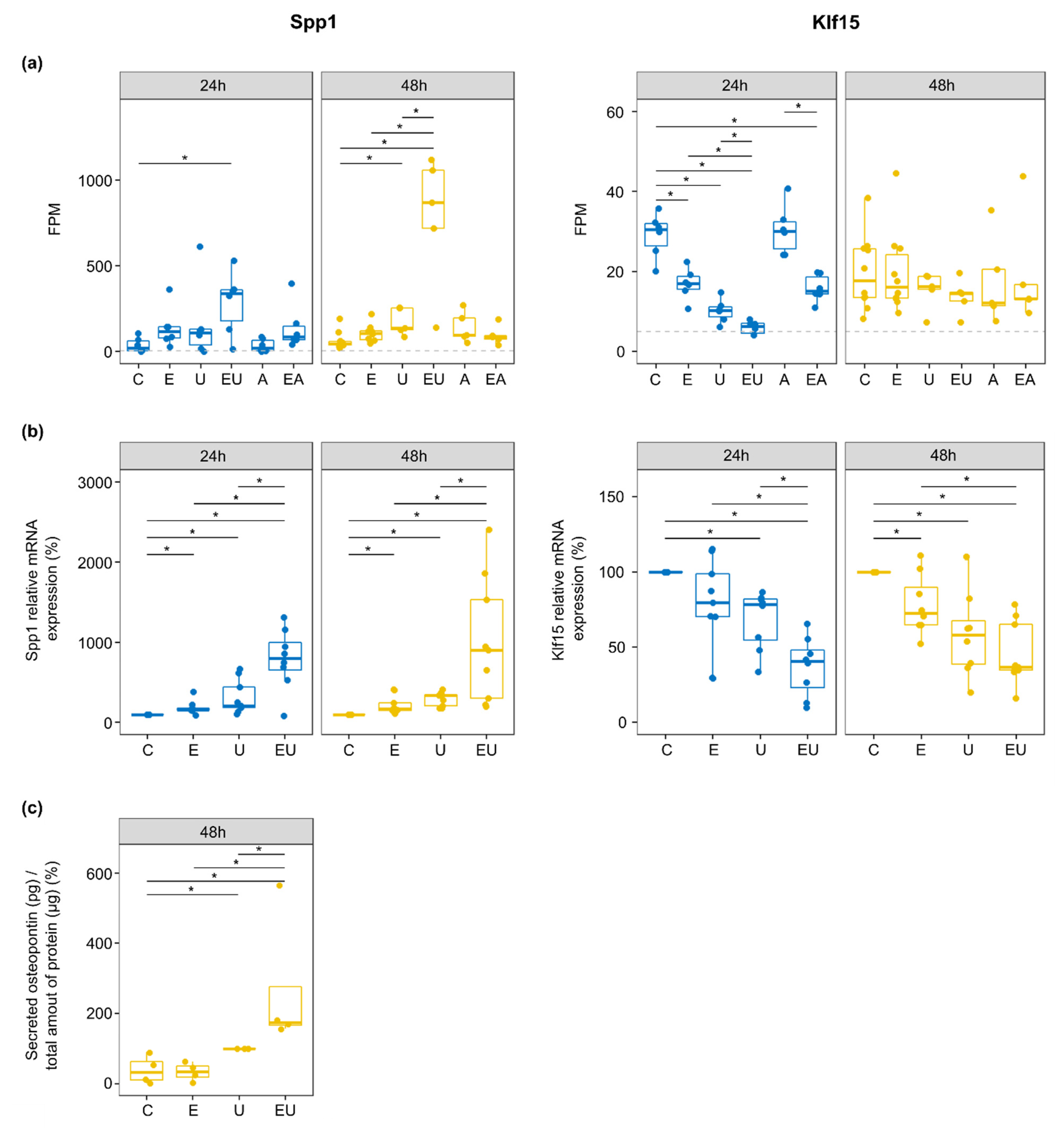
3.7. EGFR and TP-Activation Synergistically Regulate Genes Relevant in Cardiovascular Injuries and Diseases
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Global Status Report on Noncommunicable Diseases; World Health Organization: Geneva, Switzerland, 2010; Volume 53, pp. 1689–1699. [Google Scholar] [CrossRef]
- Nakahata, N. Thromboxane A2: Physiology/Pathophysiology, Cellular Signal Transduction and Pharmacology. Pharmacol. Ther. 2008, 118, 18–35. [Google Scholar] [CrossRef] [PubMed]
- Schreier, B.; Gekle, M.; Grossmann, C. Role of Epidermal Growth Factor Receptor in Vascular Structure and Function. Curr. Opin. Nephrol. Hypertens. 2014, 23, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Forrester, S.J.; Booz, G.W.; Sigmund, C.D.; Coffman, T.M.; Kawai, T.; Rizzo, V.; Scalia, R.; Eguchi, S. Angiotensin II Signal Transduction: An Update on Mechanisms of Physiology and Pathophysiology. Physiol. Rev. 2018, 98, 1627–1738. [Google Scholar] [CrossRef] [PubMed]
- Montezano, A.C.; Nguyen Dinh Cat, A.; Rios, F.J.; Touyz, R.M. Angiotensin II and Vascular Injury. Curr. Hypertens. Rep. 2014, 16, 431. [Google Scholar] [CrossRef]
- Smyth, E.M. Thromboxane and the Thromboxane Receptor in Cardiovascular Disease. Clin. Lipidol. 2010, 5, 209–219. [Google Scholar] [CrossRef]
- Chen, J.; Zeng, F.; Forrester, S.J.; Eguchi, S.; Zhang, M.Z.; Harris, R.C. Expression and Function of the Epidermal Growth Factor Receptor in Physiology and Disease. Physiol. Rev. 2016, 96, 1025–1069. [Google Scholar] [CrossRef]
- Cattaneo, F.; Guerra, G.; Parisi, M.; De Marinis, M.; Tafuri, D.; Cinelli, M.; Ammendola, R. Cell-Surface Receptors Transactivation Mediated by G Protein-Coupled Receptors. Int. J. Mol. Sci. 2014, 15, 19700–19728. [Google Scholar] [CrossRef]
- Mifune, M.; Ohtsu, H.; Suzuki, H.; Nakashima, H.; Brailoiu, E.; Dun, N.J.; Frank, G.D.; Inagami, T.; Higashiyamii, S.; Thomas, W.G.; et al. G Protein Coupling and Second Messenger Generation Are Indispensable for Metalloprotease-Dependent, Heparin-Binding Epidermal Growth Factor Shedding through Angiotensin II Type-1 Receptor. J. Biol. Chem. 2005, 280, 26592–26599. [Google Scholar] [CrossRef]
- Touyz, R.M.; Wu, X.H.; He, G.; Salomon, S.; Schiffrin, E.L. Increased Angiotensin II-Mediated Src Signaling via Epidermal Growth Factor Receptor Transactivation Is Associated with Decreased C-Terminal Src Kinase Activity in Vascular Smooth Muscle Cells from Spontaneously Hypertensive Rats. Hypertension 2002, 39, 479–485. [Google Scholar] [CrossRef]
- Uchiyama, K.; Saito, M.; Sasaki, M.; Obara, Y.; Higashiyama, S.; Nakahata, N. Thromboxane A2 Receptor-Mediated Epidermal Growth Factor Receptor Transactivation: Involvement of PKC-δ and PKC-ε in the Shedding of Epidermal Growth Factor Receptor Ligands. Eur. J. Pharm. Sci. 2009, 38, 504–511. [Google Scholar] [CrossRef]
- Forrester, S.J.; Kawai, T.; O’Brien, S.; Thomas, W.; Harris, R.C.; Eguchi, S. Epidermal Growth Factor Receptor Transactivation: Mechanisms, Pathophysiology, and Potential Therapies in the Cardiovascular System. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 627–653. [Google Scholar] [CrossRef] [PubMed]
- Kagiyama, S.; Eguchi, S.; Frank, G.D.; Inagami, T.; Zhang, Y.C.; Phillips, M.I. Angiotensin II-Induced Cardiac Hypertrophy and Hypertension Are Attenuated by Epidermal Growth Factor Receptor Antisense. Circulation 2002, 106, 909–912. [Google Scholar] [CrossRef] [PubMed]
- Schreier, B.; Hünerberg, M.; Mildenberger, S.; Rabe, S.; Bethmann, D.; Wickenhauser, C.; Gekle, M. Deletion of the EGF Receptor in Vascular Smooth Muscle Cells Prevents Chronic Angiotensin II-Induced Arterial Wall Stiffening and Media Thickening. Acta Physiol. 2018, 222, 1–14. [Google Scholar] [CrossRef]
- Schreier, B.; Schwerdt, G.; Heise, C.; Bethmann, D.; Rabe, S.; Mildenberger, S.; Gekle, M. Substance-Specific Importance of EGFR for Vascular Smooth Muscle Cells Motility in Primary Culture. Biochim. Biophys. Acta—Mol. Cell Res. 2016, 1863, 1519–1533. [Google Scholar] [CrossRef] [PubMed]
- Lynn Ray, J.; Leach, R.; Herbert, J.M.; Benson, M. Isolation of Vascular Smooth Muscle Cells from a Single Murine Aorta. Methods Cell Sci. 2001, 23, 185–188. [Google Scholar] [CrossRef]
- Schreier, B.; Döhler, M.; Rabe, S.; Schneider, B.; Schwerdt, G.; Ruhs, S.; Sibilia, M.; Gotthardt, M.; Gekle, M.; Grossmann, C. Consequences of Epidermal Growth Factor Receptor (ErbB1) Loss for Vascular Smooth Muscle Cells from Mice with Targeted Deletion of ErbB1. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 1643–1652. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-Based Genome Alignment and Genotyping with HISAT2 and HISAT-Genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Durinck, S.; Spellman, P.T.; Birney, E.; Huber, W. Mapping Identifiers for the Integration of Genomic Datasets with the R/Bioconductor Package BiomaRt. Nat. Protoc. 2009, 4, 1184–1191. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Conway, J.R.; Lex, A.; Gehlenborg, N. UpSetR: An R Package for the Visualization of Intersecting Sets and Their Properties. Bioinformatics 2017, 33, 2938–2940. [Google Scholar] [CrossRef] [PubMed]
- Reimand, J.; Arak, T.; Adler, P.; Kolberg, L.; Reisberg, S.; Peterson, H.; Vilo, J. G:Profiler-a Web Server for Functional Interpretation of Gene Lists (2016 Update). Nucleic Acids Res. 2016, 44, W83–W89. [Google Scholar] [CrossRef] [PubMed]
- Sergi Sayols Rrvgo: A Bioconductor Package to Reduce and Visualize Gene Ontology Terms. Version 1.0.2. Available online: https://ssayols.github.io/rrvgo (accessed on 30 April 2021).
- Krämer, A.; Green, J.; Pollard, J.; Tugendreich, S. Causal Analysis Approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Debacq-Chainiaux, F.; Erusalimsky, J.D.; Campisi, J.; Toussaint, O. Protocols to Detect Senescence-Associated Beta-Galactosidase (SA-Βgal) Activity, a Biomarker of Senescent Cells in Culture and in Vivo. Nat. Protoc. 2009, 4, 1798–1806. [Google Scholar] [CrossRef]
- Wetzker, R.; Böhmer, F.D. Transactivation Joins Multiple Tracks to the ERK/MAPK Cascade. Nat. Rev. Mol. Cell Biol. 2003, 4, 651–657. [Google Scholar] [CrossRef]
- Li, Y.; Liu, S.; Zhang, Y.; Gao, Q.; Sun, W.; Fu, L.; Cao, J. Histone Demethylase JARID1B Regulates Proliferation and Migration of Pulmonary Arterial Smooth Muscle Cells in Mice with Chronic Hypoxia-Induced Pulmonary Hypertension via Nuclear Factor-Kappa B (NFkB). Cardiovasc. Pathol. 2018, 37, 8–14. [Google Scholar] [CrossRef]
- Bao, L.; Odell, A.F.; Stephen, S.L.; Wheatcroft, S.B.; Walker, J.H.; Ponnambalam, S. The S100A6 Calcium-Binding Protein Regulates Endothelial Cell-Cycle Progression and Senescence. FEBS J. 2012, 279, 4576–4588. [Google Scholar] [CrossRef]
- Grandori, C.; Cowley, S.M.; James, L.P.; Eisenman, R.N. The myc/max/mad network and the transcriptional control of cell behavior. Annu. Rev. Cell. Dev. Biol. 2000, 16, 653–699. [Google Scholar] [CrossRef]
- Emanuele, M.J.; Ciccia, A.; Elia, A.E.H.; Elledge, S.J. Proliferating Cell Nuclear Antigen (PCNA)-Associated KIAA0101/PAF15 Protein Is a Cell Cycle-Regulated Anaphase-Promoting Complex/Cyclosome Substrate. Proc. Natl. Acad. Sci. USA 2011, 108, 9845–9850. [Google Scholar] [CrossRef]
- Amati, B.; Land, H. Myc-Max-Mad: A Transcription Factor Network Controlling Cell Cycle Progression, Differentiation and Death. Curr. Opin. Genet. Dev. 1994, 4, 102–108. [Google Scholar] [CrossRef]
- Napolitano, G.; Ballabio, A. TFEB at a glance. J. Cell Sci. 2016, 129, 2475–2481. [Google Scholar] [CrossRef] [PubMed]
- Nijwening, J.H.; Geutjes, E.J.; Bernards, R.; Beijersbergen, R.L. The Histone Demethylase Jarid1b (Kdm5b) Is a Novel Component of the Rb Pathway and Associates with E2f-Target Genes in MEFs during Senescence. PLoS ONE 2011, 6, e25235. [Google Scholar] [CrossRef] [PubMed]
- Ross, R.; Glomset, J.A. Atherosclerosis and the Arterial Smooth Muscle Cell: Proliferation of Smooth Muscle Is a Key Event in the Genesis of the Lesions of Atherosclerosis. Science 1973, 180, 1332–1339. [Google Scholar] [CrossRef] [PubMed]
- Clarke, M.; Bennett, M. The Emerging Role of Vascular Smooth Muscle Cell Apoptosis in Atherosclerosis and Plaque Stability. Am. J. Nephrol. 2007, 26, 531–535. [Google Scholar] [CrossRef]
- Katsuumi, G.; Shimizu, I.; Yoshida, Y.; Minamino, T. Vascular Senescence in Cardiovascular and Metabolic Diseases. Front. Cardiovasc. Med. 2018, 5, 1–13. [Google Scholar] [CrossRef]
- Lu, Y.; Haldar, S.; Croce, K.; Wang, Y.; Sakuma, M.; Morooka, T.; Wang, B.; Jeyaraj, D.; Gray, S.J.; Simon, D.I.; et al. Kruppel-like Factor 15 Regulates Smooth Muscle Response to Vascular Injury-Brief Report. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1550–1552. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, L.; Liao, X.; Sangwung, P.; Prosdocimo, D.; Zhou, G.; Votruba, A.; Brian, L.; Han, Y.J.; Gao, H.; et al. Kruppel-like Factor 15 Is Critical for Vascular Inflammation. J. Clin. Invest. 2013, 123, 4232–4241. [Google Scholar] [CrossRef]
- Lok, Z.S.Y.; Lyle, A.N. Osteopontin in Vascular Disease. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 613–622. [Google Scholar] [CrossRef]
- Bagby, S.P.; Kirk, E.A.; Mitchell, L.H.; O’Reilly, M.M.; Holden, W.E.; Stenberg, P.E.; Bakke, A.C. Proliferative Synergy of ANG II and EGF in Porcine Aortic Vascular Smooth Muscle Cells. Am. J. Physiol.—Ren. Fluid Electrolyte Physiol. 1993, 265, F239–F249. [Google Scholar] [CrossRef]
- Kong, K.C.; Billington, C.K.; Gandhi, U.; Panettieri, R.A.; Penn, R.B.; Kong, K.C.; Billington, C.K.; Gandhi, U.; Panettieri, R.A.; Penn, R.B. Cooperative Mitogenic Signaling by G Protein-coupled Receptors and Growth Factors Is Dependent on G q/11. FASEB J. 2006, 20, 1558–1560. [Google Scholar] [CrossRef] [PubMed]
- Ko, Y.; Stiebler, H.; Nickenig, G.; Wieczorek, A.J.; Vetter, H.; Sachinidis, A. Synergistic Action of Angiotensin II, Insulin-like Growth Factor-I, and Transforming Growth Factor-b on Platelet-Derived Growth Factor-BB, Basic Fibroblastic Growth Factor, and Epidermal Growth Factor-Induced DNA Synthesis in Vascular Smooth Muscle Cells. Am. J. Hypertens. 1993, 6, 496–499. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.L.; Tung, W.H.; Wu, C.Y.; Wang, H.H.; Lin, C.C.; Wang, T.S.; Yang, C.M. Thrombin Induces EGF Receptor Expression and Cell Proliferation via a PKC(δ)/c-Src-Dependent Pathway in Vascular Smooth Muscle Cells. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1594–1601. [Google Scholar] [CrossRef] [PubMed]
- Capra, V.; Habib, A.; Accomazzo, M.R.; Ravasi, S.; Citro, S.; Levy-Toledano, S.; Nicosia, S.; Rovati, G.E. Thromboxane Prostanoid Receptor in Human Airway Smooth Muscle Cells: A Relevant Role in Proliferation. Eur. J. Pharmacol. 2003, 474, 149–159. [Google Scholar] [CrossRef]
- Sachinidis, A.; Flesch, M.; Ko, Y.; Schrör, K.; Böhm, M.; Düsing, R.; Vetter, H. Thromboxane A2 and Vascular Smooth Muscle Cell Proliferation. Hypertension 1995, 26, 771–780. [Google Scholar] [CrossRef]
- Grosser, T.; Zucker, T.P.; Weber, A.A.; Schulte, K.; Sachinidis, A.; Vetter, H.; Schrör, K. Thromboxane A2 Induces Cell Signaling but Requires Platelet-Derived Growth Factor to Act as a Mitogen. Eur. J. Pharmacol. 1997, 319, 327–332. [Google Scholar] [CrossRef]
- Postolow, F.; Fediuk, J.; Nolette, N.; Hinton, M.; Dakshinamurti, S. Thromboxane Promotes Smooth Muscle Phenotype Commitment but Not Remodeling of Hypoxic Neonatal Pulmonary Artery. Fibrogenes. Tissue Repair 2015, 8, 20. [Google Scholar] [CrossRef]
- González-Gualda, E.; Baker, A.G.; Fruk, L.; Muñoz-Espín, D. A Guide to Assessing Cellular Senescence in Vitro and in Vivo. FEBS J. 2021, 288, 56–80. [Google Scholar] [CrossRef]
- Childs, B.G.; Li, H.; Van Deursen, J.M. Senescent Cells: A Therapeutic Target for Cardiovascular Disease. J. Clin. Investig. 2018, 128, 1217–1228. [Google Scholar] [CrossRef]
- Frismantiene, A.; Philippova, M.; Erne, P.; Resink, T.J. Smooth Muscle Cell-Driven Vascular Diseases and Molecular Mechanisms of VSMC Plasticity. Cell. Signal. 2018, 52, 48–64. [Google Scholar] [CrossRef]
- Lee, S.J.; Baek, S.E.; Jang, M.A.; Kim, C.D. Osteopontin Plays a Key Role in Vascular Smooth Muscle Cell Proliferation via EGFR-Mediated Activation of AP-1 and C/EBPβ Pathways. Pharmacol. Res. 2016, 108, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Berezin, A.E. The Controversial Role of Osteopontin in Vascular Calcification: From Bench to Bedside. MOJ Proteom. Bioinform. 2017, 5, 129–130. [Google Scholar] [CrossRef][Green Version]
- Wolak, T. Osteopontin—A Multi-Modal Marker and Mediator in Atherosclerotic Vascular Disease. Atherosclerosis 2014, 236, 327–337. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EGF | U46619 | EGF and U46619 | |||||
|---|---|---|---|---|---|---|---|
| Z-Score | −log(adj-p) | Z-Score | −log(adj-p) | Z-Score | −log(adj-p) | ||
| EGF-specific | IL2 | 2.12 | 3.20 | 1.32 | 1.32 | 0.92 | 1.14 |
| NLRP3 | −3.00 | 3.64 | −0.93 | 1.18 | −1.73 | 0.97 | |
| U46619-specific | EIF2AK3 | N/A | 0.00 | 3.08 | 4.21 | 1.86 | 2.18 |
| PNPT1 | N/A | 0.00 | 3.34 | 3.80 | N/A | 0.00 | |
| EGF and U46619-specific | IKBKG | N/A | 0.00 | −1.79 | 2.45 | −2.18 | 3.92 |
| KDM5B | −1.97 | 1.78 | −1.75 | 0.83 | −2.64 | 4.13 | |
| MAX | N/A | 1.21 | N/A | 1.82 | 2.00 | 4.04 | |
| PCLAF | N/A | 0.00 | N/A | 0.00 | 3.16 | 3.37 | |
| PTGER2 | N/A | 0.00 | N/A | 0.00 | 4.70 | 6.61 | |
| S100A6 | 1.07 | 2.07 | 1.71 | 0.74 | 2.35 | 3.02 | |
| TFEB | N/A | 0.00 | −1.92 | 2.30 | −3.92 | 3.81 | |
| Gene | Ensemble Identifier | Intersection | Substances | Regulation |
|---|---|---|---|---|
| Ctdsp1 | ENSMUSG00000026176 | i1 | EGF, U46619 | Down |
| Fam110b | ENSMUSG00000049119 | i5 | EGF, U46619 | Down |
| Gm6665 | ENSMUSG00000091561 | i17 | EGF, U46619 | Down |
| Gstm2 | ENSMUSG00000040562 | i17 | EGF, U46619 | Down |
| Hadhb-ps | ENSMUSG00000063684 | i1 | EGF, U46619 | Down |
| Inpp4a | ENSMUSG00000026113 | i1 | EGF, U46619 | Down |
| Klf15 | ENSMUSG00000030087 | i7 | EGF, U46619 | Down |
| Mtap | ENSMUSG00000062937 | i1 | EGF, U46619 | Up |
| Pcif1 | ENSMUSG00000039849 | i1 | EGF, U46619 | Down |
| Spp1 | ENSMUSG00000029304 | i17 | EGF, U46619 | Up |
| Srr | ENSMUSG00000001323 | i6 | EGF, U46619 | Down |
| Tpst1 | ENSMUSG00000034118 | i1 | EGF, U46619 | Down |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dubourg, V.; Schreier, B.; Schwerdt, G.; Rabe, S.; Benndorf, R.A.; Gekle, M. The Functional Interaction of EGFR with AT1R or TP in Primary Vascular Smooth Muscle Cells Triggers a Synergistic Regulation of Gene Expression. Cells 2022, 11, 1936. https://doi.org/10.3390/cells11121936
Dubourg V, Schreier B, Schwerdt G, Rabe S, Benndorf RA, Gekle M. The Functional Interaction of EGFR with AT1R or TP in Primary Vascular Smooth Muscle Cells Triggers a Synergistic Regulation of Gene Expression. Cells. 2022; 11(12):1936. https://doi.org/10.3390/cells11121936
Chicago/Turabian StyleDubourg, Virginie, Barbara Schreier, Gerald Schwerdt, Sindy Rabe, Ralf A. Benndorf, and Michael Gekle. 2022. "The Functional Interaction of EGFR with AT1R or TP in Primary Vascular Smooth Muscle Cells Triggers a Synergistic Regulation of Gene Expression" Cells 11, no. 12: 1936. https://doi.org/10.3390/cells11121936
APA StyleDubourg, V., Schreier, B., Schwerdt, G., Rabe, S., Benndorf, R. A., & Gekle, M. (2022). The Functional Interaction of EGFR with AT1R or TP in Primary Vascular Smooth Muscle Cells Triggers a Synergistic Regulation of Gene Expression. Cells, 11(12), 1936. https://doi.org/10.3390/cells11121936