
Blood Cell In Vitro Cytokine Production in Response to Lipopolysaccharide Stimulation in a Healthy Population: Effects of Age, Sex, and Smoking

 ,
,  ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Design and Participants
2.2. Laboratory Visit
2.3. Blood Culture and LPS Stimulation
2.4. Cytokine Measurements
2.5. Statistical Analysis
3. Results
3.1. General Characteristics of Participants in the Study
3.2. Culture Cytokine Production
3.3. Correlations between Cytokine-Stimulated Production, Age, and BMI
3.4. Linear Regression Analysis for IL-6-, IL-10-, and TNF-α-Stimulated Production
3.5. Effects of the Menstrual Cycle on the Cytokine-Stimulated Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abbasi, A.; Fehrenbach, E.; Hauth, M.; Walter, M.; Hudemann, J.; Wank, V.; Niess, A.M.; Northoff, H. Changes in spontaneous and lps-induced ex vivo cytokine production and mrna expression in male and female athletes following prolonged exhaustive exercise. Exerc. Immunol. Rev. 2013, 19, 8–28. [Google Scholar] [PubMed]
- Beenakker, K.G.M.; Westendorp, R.G.J.; De Craen, A.J.M.; Chen, S.; Raz, Y.; Ballieux, B.E.P.B.; Nelissen, R.G.H.H.; Later, A.F.L.; Huizinga, T.W.; Slagboom, P.E.; et al. Men Have a Stronger Monocyte-Derived Cytokine Production Response upon Stimulation with the Gram-Negative Stimulus Lipopolysaccharide than Women: A Pooled Analysis Including 15 Study Populations. J. Innate Immun. 2020, 12, 142–153. [Google Scholar] [CrossRef]
- Damsgaard, C.T.; Lauritzen, L.; Calder, P.C.; Kjær, T.M.R.; Frøkiær, H. Whole-blood culture is a valid low-cost method to measure monocytic cytokines—A comparison of cytokine production in cultures of human whole-blood, mononuclear cells and monocytes. J. Immunol. Methods 2009, 340, 95–101. [Google Scholar] [CrossRef] [PubMed]
- De Craen, A.J.M.; Posthuma, D.; Remarque, E.J.; van den Biggelaar, A.H.J.; Westendorp, R.G.J.; Boomsma, D.I. Heritability estimates of innate immunity: An extended twin study. Genes Immun. 2005, 6, 167–170. [Google Scholar] [CrossRef]
- Gon, Y.; Hashimoto, S.; Hayashi, S.; Koura, T.; Matsumoto, K.; Horie, T. Lower serum concentrations of cytokines in elderly patients with pneumonia and the impaired production of cytokines by peripheral blood monocytes in the elderly. Clin. Exp. Immunol. 1996, 106, 120–126. [Google Scholar]
- Wijsman, C.A.; Maier, A.B.; De Craen, A.J.M.; Van Den Biggelaar, A.H.J.; Westendorp, R.G.J. An unopposed proinflammatory response is beneficial for survival in the oldest old. Results of the Leiden 85-plus study. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2011, 66 A, 393–399. [Google Scholar] [CrossRef] [Green Version]
- Van Exel, E.; Gussekloo, J.; De Craen, A.J.M.; Frölich, M.; Van Der Wiel, A.B.; Westendorp, R.G.J. Low production capacity of interleukin-10 associates with the metabolic syndrome and type 2 diabetes: The Leiden 85-plus study. Diabetes 2002, 51, 1088–1092. [Google Scholar] [CrossRef] [Green Version]
- Rozing, M.P.; Westendorp, R.G.J.; Maier, A.B.; Wijsman, C.A.; Frölich, M.; De Craen, A.J.M.; Van Heemst, D. Serum triiodothyronine levels and inflammatory cytokine production capacity. Age 2012, 34, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Beenakker, K.G.M.; Westendorp, R.G.J.; De Craen, A.J.M.; Slagboom, P.E.; Van Heemst, D.; Maier, A.B. Pro-inflammatory capacity of classically activated monocytes relates positively to muscle mass and strength. Aging Cell 2013, 12, 682–689. [Google Scholar] [CrossRef] [PubMed]
- Lisi, L.; Camardese, G.; Treglia, M.; Tringali, G.; Carrozza, C.; Janiri, L.; Dello Russo, C.; Navarra, P. Monocytes from Depressed Patients Display an Altered Pattern of Response to Endotoxin Challenge. PLoS ONE 2013, 8, e52585. [Google Scholar]
- Bruunsgaard, H.; Pedersen, A.N.; Schroll, M.; Skinhoj, P.; Pedersen, B.K. Impaired production of proinflammatory cytokines in response to lipopolysaccharide (LPS) stimulation in elderly humans. Clin Exp. Immunol. 1999, 118, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Auger, G.; Corvec, S.; Roquilly, A.; Segain, J.P.; Lepelletier, D.; Reynaud, A.; Asehnoune, K. Escherichia coli-induced productions of pro-inflammatory cytokines are regulated by MAP kinases and G-protein but not by Akt: Relationship with phylogenetic groups and resistance patterns. Cytokine 2011, 56, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Heesen, M.; Bloemeke, B.; Heussen, N.; Kunz, D. Can the interleukin-6 response to endotoxin be predicted? Studies of the influence of a promoter polymorphism of the interleukin-6 gene, gender, the density of the endotoxin receptor CD14, and inflammatory cytokines. Crit. Care Med. 2002, 30, 664–669. [Google Scholar] [CrossRef]
- Lynch, E.A.; Dinarello, C.A.; Cannon, J.G. Gender differences in IL-1α, IL-1β, and IL-1 receptor antagonist secretion from mononuclear cells and urinary excretion. J. Immunol. 1994, 153, 300–306. [Google Scholar] [PubMed]
- Ter Horst, R.; Jaeger, M.; Smeekens, S.P.; Oosting, M.; Swertz, M.A.; Li, Y.; Kumar, V.; Diavatopoulos, D.A.; Jansen, A.F.M.; Lemmers, H.; et al. Host and Environmental Factors Influencing Individual Human Cytokine Responses. Cell 2016, 167, 1111–1124. [Google Scholar] [CrossRef] [Green Version]
- Segre, E.; Fullerton, J.N. Stimulated whole blood cytokine release as a biomarker of immunosuppression in the critically ill: The need for a standardized methodology. Shock 2016, 45, 490–494. [Google Scholar] [CrossRef] [Green Version]
- Drewry, A.M.; Ablordeppey, E.A.; Murray, E.T.; Beiter, E.R.; Walton, A.H.; Hall, M.W.; Hotchkiss, R.S. Comparison of monocyte human leukocyte antigen-DR expression and stimulated tumor necrosis factor alpha production as outcome predictors in severe sepsis: A prospective observational study. Crit. Care 2016, 20, 334. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.W.; Knatz, N.L.; Vetterly, C.; Tomarello, S.; Wewers, M.D.; Volk, H.D.; Carcillo, J.A. Immunoparalysis and nosocomial infection in children with multiple organ dysfunction syndrome. Intensive Care Med. 2011, 37, 525–532. [Google Scholar] [CrossRef] [Green Version]
- Döcke, W.D.; Randow, F.; Syrbe, U.; Krausch, D.; Asadullah, K.; Reinke, P.; Volk, H.D.; Kox, W. Monocyte deactivation in septic patients: Restoration by IFN-γ treatment. Nat. Med. 1997, 3, 678–681. [Google Scholar] [CrossRef]
- Simon, A.K.; Hollander, G.A.; McMichael, A. Evolution of the immune system in humans from infancy to old age. Proc. R. Soc. B Biol. Sci. 2015, 282, 20143085. [Google Scholar] [CrossRef]
- Martínez de Toda, I.; Maté, I.; Vida, C.; Cruces, J.; De la Fuente, M. Immune function parameters as markers of biological age and predictors of longevity. Aging 2016, 8, 3110–3119. [Google Scholar] [CrossRef] [Green Version]
- Westendorp, R.G.J.; Van Dunne, F.M.; Kirkwood, T.B.L.; Helmerhorst, F.M.; Huizinga, T.W.J. Optimizing human fertility and survival. Nat. Med. 2001, 7, 873. [Google Scholar] [CrossRef]
- Gleeson, M.; Bishop, N.; Oliveira, M.; McCauley, T.; Tauler, P.; Muhamad, A.S. Respiratory infection risk in athletes: Association with antigen-stimulated IL-10 production and salivary IgA secretion. Scand. J. Med. Sci. Sports 2012, 22, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Smit, L.A.M.; Heederik, D.; Doekes, G.; Krop, E.J.M.; Rijkers, G.T.; Wouters, I.M. Ex vivo cytokine release reflects sensitivity to occupational endotoxin exposure. Eur. Respir. J. 2009, 34, 795–802. [Google Scholar] [CrossRef] [Green Version]
- Westendorp, R.G.J.; Langermans, J.A.M.; Huizinga, T.W.J.; Elouali, A.H.; Verweij, C.L.; Boomsma, D.I.; Vandenbrouke, J.P. Genetic influence on cytokine production and fatal meningococcal disease. Lancet 1997, 349, 170–173. [Google Scholar] [CrossRef]
- Van der Linden, M.W.; Westendorp, R.G.J.; Sturk, A.; Bergman, W.; Huizinga, T.W.J. High interleukin-10 production in first-degree relatives of patients with generalized but not cutaneous lupus erythematosus. J. Investig. Med. 2000, 48, 327–334. [Google Scholar] [PubMed]
- De Jong, B.A.; Huizinga, T.W.J.; Bollen, E.L.E.M.; Uitdehaag, B.M.J.; Bosma, G.P.T.; Van Buchem, M.A.; Remarque, E.J.; Burgmans, A.C.S.; Kalkers, N.F.; Polman, C.H.; et al. Production of IL-1β and IL-1Ra as risk factors for susceptibility and progression of relapse-onset multiple sclerosis. J. Neuroimmunol. 2002, 126, 172–179. [Google Scholar] [CrossRef]
- Von Aulock, S.; Deininger, S.; Draing, C.; Gueinzius, K.; Dehus, O.; Hermann, C. Gender difference in cytokine secretion on immune stimulation with LPS and LTA. J. Interf. Cytokine Res. 2006, 26, 887–892. [Google Scholar] [CrossRef]
- Dombrovskiy, V.Y.; Martin, A.A.; Sunderram, J.; Paz, H.L. Rapid increase in hospitalization and mortality rates for severe sepsis in the United States: A trend analysis from 1993 to 2003. Crit. Care Med. 2007, 35, 1244–1250. [Google Scholar] [CrossRef]
- Sopori, M. Effects of cigarette smoke on the immune system. Nat. Rev. Immunol. 2002, 2, 372–377. [Google Scholar] [CrossRef]
- Chen, H.; Cowan, M.J.; Hasday, J.D.; Vogel, S.N.; Medvedev, A.E. Tobacco Smoking Inhibits Expression of Proinflammatory Cytokines and Activation of IL-1R-Associated Kinase, p38, and NF-κB in Alveolar Macrophages Stimulated with TLR2 and TLR4 Agonists. J. Immunol. 2007, 179, 6097–6106. [Google Scholar] [CrossRef] [PubMed]
- Van Zyl-Smit, R.N.; Binder, A.; Meldau, R.; Semple, P.L.; Evans, A.; Smith, P.; Bateman, E.D.; Dheda, K. Cigarette smoke impairs cytokine responses and BCG containment in alveolar macrophages. Thorax 2014, 69, 363–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsunaga, K.; Klein, T.W.; Friedman, H.; Yamamoto, Y. Involvement of Nicotinic Acetylcholine Receptors in Suppression of Antimicrobial Activity and Cytokine Responses of Alveolar Macrophages to Legionella pneumophila Infection by Nicotine. J. Immunol. 2001, 167, 6518–6524. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.G.; Danis, V.A.; Brooks, P.M. Effect of gonadal steroids on the production of IL-1 and IL-6 blood mononuclear cells in vitro. Clin. Exp. Rheumatol. 1993, 11, 157–162. [Google Scholar] [PubMed]
- Pacifici, R.; Brown, C.; Puscheck, E.; Friedrich, E.; Slatopolsky, E.; Maggio, D.; Mccracken, R.; Avioli, L.V. Effect of surgical menopause and estrogen replacement on cytokine release from human blood mononuclear cells. Proc. Natl. Acad. Sci. USA 1991, 88, 5134–5138. [Google Scholar] [CrossRef] [Green Version]
- Allen, A.M.; McRae-Clark, A.L.; Carlson, S.; Saladin, M.E.; Gray, K.M.; Wetherington, C.L.; McKee, S.A.; Allen, S.S. Determining menstrual phase in human biobehavioral research: A review with recommendations. Exp. Clin. Psychopharmacol. 2016, 24, 1–11. [Google Scholar] [CrossRef]
- Straub, R.H. The complex role of estrogens in inflammation. Endocr. Rev. 2007, 28, 521–574. [Google Scholar] [CrossRef] [Green Version]
- Angstwurm, M.W.A.; Gärtner, R.; Ziegler-Heitbrock, H.W.L. Cyclic plasma IL-6 levels during normal menstrual cycle. Cytokine 1997, 9, 370–374. [Google Scholar] [CrossRef]
- Janský, L.; Reymanová, P.; Kopecký, J. Dynamics of Cytokine Production in Human Peripheral Blood Mononuclear Cells Stimulated by LPS or Infected by Borrelia. Physiol. Res. 2003, 52, 593–598. [Google Scholar]
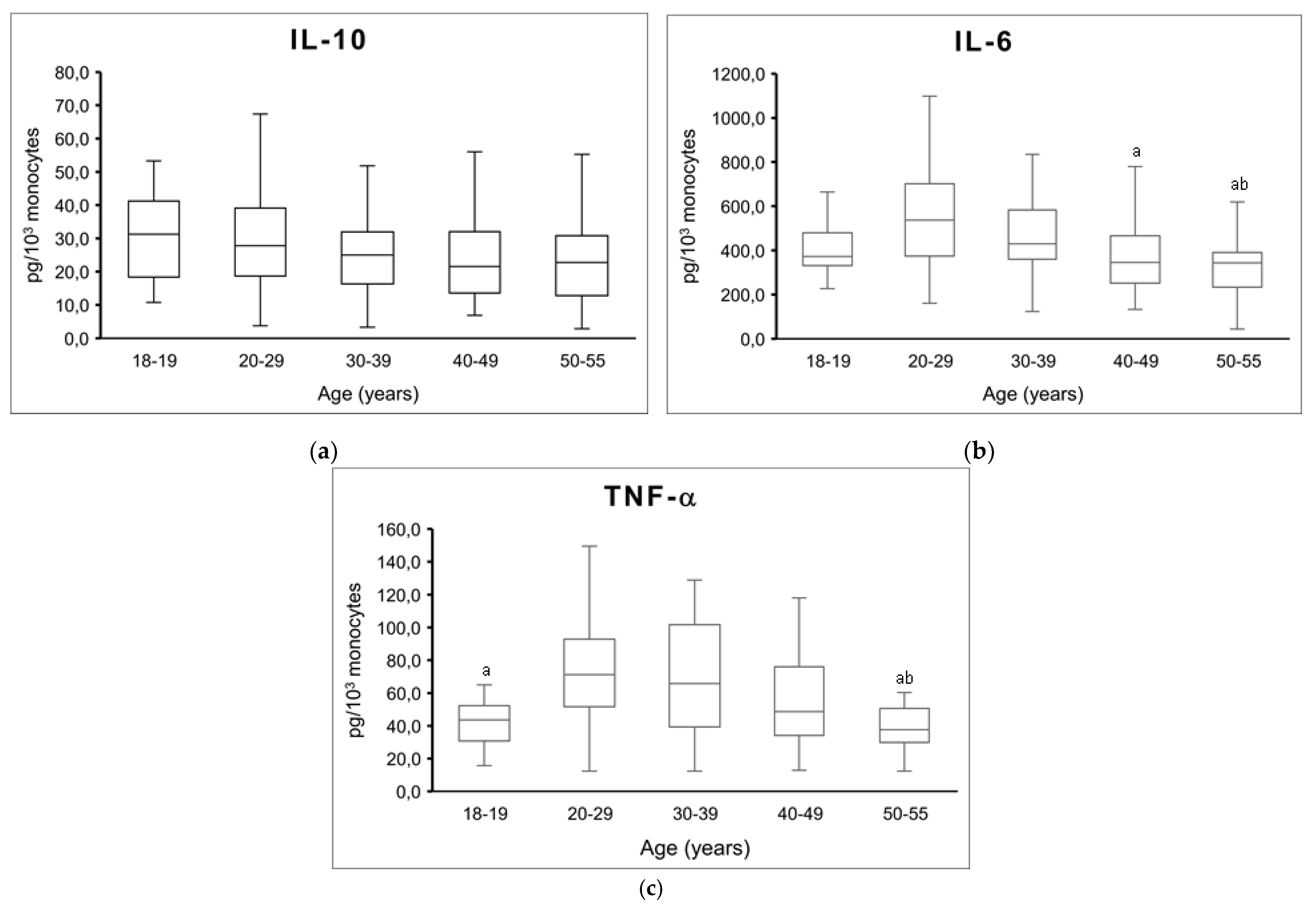

{kind=link}
{kind=link}
| Parameter | All (n = 253) | Men (n = 120) | Women (n = 133) | p Value |
|---|---|---|---|---|
| Age (years) | 33.0 ± 10.9 | 33.4 ± 11.0 | 32.6 ± 10.9 | 0.534 |
| Body mass (kg) | 67.0 ± 13.5 | 76.3 ± 11.8 | 58.6 ± 8.6 | <0.001 * |
| Stature (cm) | 170.0 ± 9.6 | 177.0 ± 7.1 | 163.6 ± 6.4 | <0.001 * |
| BMI (kg·m−2) | 23.1 ± 3.4 | 24.3 ± 3.3 | 22.0 ± 3.2 | <0.001 * |
| Leukocytes (103·μL−1) | 5.84 ± 1.38 | 5.80 ± 1.18 | 5.87 ± 1.55 | 0.681 |
| Neutrophils (103·μL−1) | 3.13 ± 0.98 | 3.07 ± 0.84 | 3.18 ± 1.08 | 0.363 |
| Lymphocytes (103·μL−1) | 1.98 ± 0.60 | 1.96 ± 0.59 | 1.99 ± 0.60 | 0.674 |
| Monocytes (103·μL−1) | 0.49 ± 0.41 | 0.52 ± 0.37 | 0.47 ± 0.44 | 0.407 |
| Cytokine | Men (n = 120) | Women (n = 133) | p Value |
|---|---|---|---|
| IL-10 (pg·10−3 cells) | 20.6 (13.5, 28.4) | 29.1 (17.4, 42.1) | <0.001 * |
| IL-6 (pg·10−3 cells) | 372.8 (262.7, 536.6) | 428.8 (331.7, 586.4) | 0.068 |
| TNF-α (pg·10−3 cells) | 56.0 (33.9, 85.3) | 49.4 (30.8, 69.1) | 0.039 * |
| Cytokine | Non-smokers (n = 221) | Smokers (n = 32) | p Value |
|---|---|---|---|
| IL-10 (pg·10−3 cells) | 25.0 (16.3, 35.7) | 14.3 (10.5, 22.6) | <0.001 * |
| IL-6 (pg·10−3 cells) | 397.8 (299.0, 562.0) | 288.3 (223.2, 445.4) | 0.003 * |
| TNF-α (pg·10−3 cells) | 53.0 (34.1, 81.5) | 42.1 (26.8, 60.0) | 0.033 * |
| B | β | 95%CI | t | p Value | R2 | Adjusted R2 | p Value (ANOVA) | R2 Change | |
|---|---|---|---|---|---|---|---|---|---|
| Model 1 | 0.064 | 0.060 | <0.001 * | 0.064 | |||||
| Smoking | −0.196 | −0.253 | (−0.289, −0.103) | −4.148 | <0.001 * | ||||
| Model 2 | 0.134 | 0.127 | <0.001 * | 0.070 | |||||
| Smoking | −0.216 | −0.280 | (−0.306, −0.126) | −4.731 | <0.001 * | ||||
| Sex | 0.137 | 0.266 | (0.077, 0.197) | 4.504 | <0.001 * | ||||
| Model 3 | 0.155 | 0.145 | <0.001 * | 0.021 | |||||
| Smoking | −0.220 | −0.285 | (−0.309, −0.131) | −4.859 | <0.001 * | ||||
| Sex | 0.134 | 0.261 | (0.075, 0.194) | 4.459 | <0.001 * | ||||
| Age | −0.003 | −0.145 | (−0.006, −0.001) | −2.490 | 0.013 * |
| B | β | 95%CI | t | p Value | R2 | Adjusted R2 | p Value (ANOVA) | R2 Change | |
|---|---|---|---|---|---|---|---|---|---|
| IL-6 | |||||||||
| Model 1 | 0.089 | 0.085 | <0.001 * | 0.089 | |||||
| Age | −0.006 | −0.298 | (−0.009, −0.004) | −4.744 | <0.001 * | ||||
| Model 2 | 0.126 | 0.118 | <0.001 * | 0.037 | |||||
| Age | −0.006 | −0.311 | (−0.009, −0.004) | −5.032 | <0.001 * | ||||
| Smoking | −0.123 | −0.193 | (−0.201, −0.045) | −3.119 | 0.002 * | ||||
| TNF-α | |||||||||
| Model 1 | 0.029 | 0.025 | 0.009 * | 0.029 | |||||
| Age | −0.005 | −0.170 | (−0.009, −0.001) | −2.623 | 0.009 * | ||||
| Model 2 | 0.051 | 0.042 | 0.002 * | 0.022 | |||||
| Age | −0.005 | −0.180 | (−0.009, −0.002) | −2.796 | 0.006 * | ||||
| Smoking | −0.137 | −0.148 | (−0.254, −0.020) | −2.309 | 0.022 * | ||||
| Model 3 | 0.067 | 0.055 | 0.001 * | 0.016 | |||||
| Age | −0.005 | −0.181 | (−0.009, −0.002) | −2.836 | 0.005 * | ||||
| Smoking | −0.137 | −0.132 | (−0.239, −0.005) | −2.309 | 0.041 * | ||||
| Sex | −0.081 | −0.129 | (−0.160, −0.002) | −3.063 | 0.045 * |
| Cytokine | Week 1 (n = 37) | Week 2 (n = 31) | Week 3 (n = 28) | Week 4 (n = 37) | p Value |
|---|---|---|---|---|---|
| IL-10 (pg·10−3 cells) | 31.3 (20.8; 44.6) | 42.3 (29.2; 51.4) a | 25.9 (19.0; 38.1) | 31.3 (14.7; 41.4) | 0.035 * |
| IL-6 (pg·10−3 cells) | 462.8 (346.3; 610.6) | 414.7 (349.1; 569.5) | 470.8 (354.9; 709.4) | 448.8 (342.1; 618.9) | 0.681 |
| TNF-α (pg·10−3 cells) | 52.1 (29.3; 85.7) | 43.4 (31.8; 58.4) | 57.7 (39.6; 80.7) | 51.3 (32.3; 78.7) | 0.383 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodas, L.; Martínez, S.; Riera-Sampol, A.; Moir, H.J.; Tauler, P. Blood Cell In Vitro Cytokine Production in Response to Lipopolysaccharide Stimulation in a Healthy Population: Effects of Age, Sex, and Smoking. Cells 2022, 11, 103. https://doi.org/10.3390/cells11010103
Rodas L, Martínez S, Riera-Sampol A, Moir HJ, Tauler P. Blood Cell In Vitro Cytokine Production in Response to Lipopolysaccharide Stimulation in a Healthy Population: Effects of Age, Sex, and Smoking. Cells. 2022; 11(1):103. https://doi.org/10.3390/cells11010103
Chicago/Turabian StyleRodas, Lluis, Sonia Martínez, Aina Riera-Sampol, Hannah J. Moir, and Pedro Tauler. 2022. "Blood Cell In Vitro Cytokine Production in Response to Lipopolysaccharide Stimulation in a Healthy Population: Effects of Age, Sex, and Smoking" Cells 11, no. 1: 103. https://doi.org/10.3390/cells11010103
APA StyleRodas, L., Martínez, S., Riera-Sampol, A., Moir, H. J., & Tauler, P. (2022). Blood Cell In Vitro Cytokine Production in Response to Lipopolysaccharide Stimulation in a Healthy Population: Effects of Age, Sex, and Smoking. Cells, 11(1), 103. https://doi.org/10.3390/cells11010103