
Betulinic Acid Protects from Ischemia-Reperfusion Injury in the Mouse Retina

 , , ,
, , ,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Application of Betulinic Acid and Induction of Ischemia-Reperfusion Injury
2.3. Retinal Wholemounts and Cell Counting
2.4. Optic Nerve Cross-Sections and Axon Counting
2.5. Measurements of Retinal Arteriole Reactivity
2.6. Assessment of ROS Levels
2.7. Quantitative PCR Analysis
2.8. Statistical Analysis
3. Results
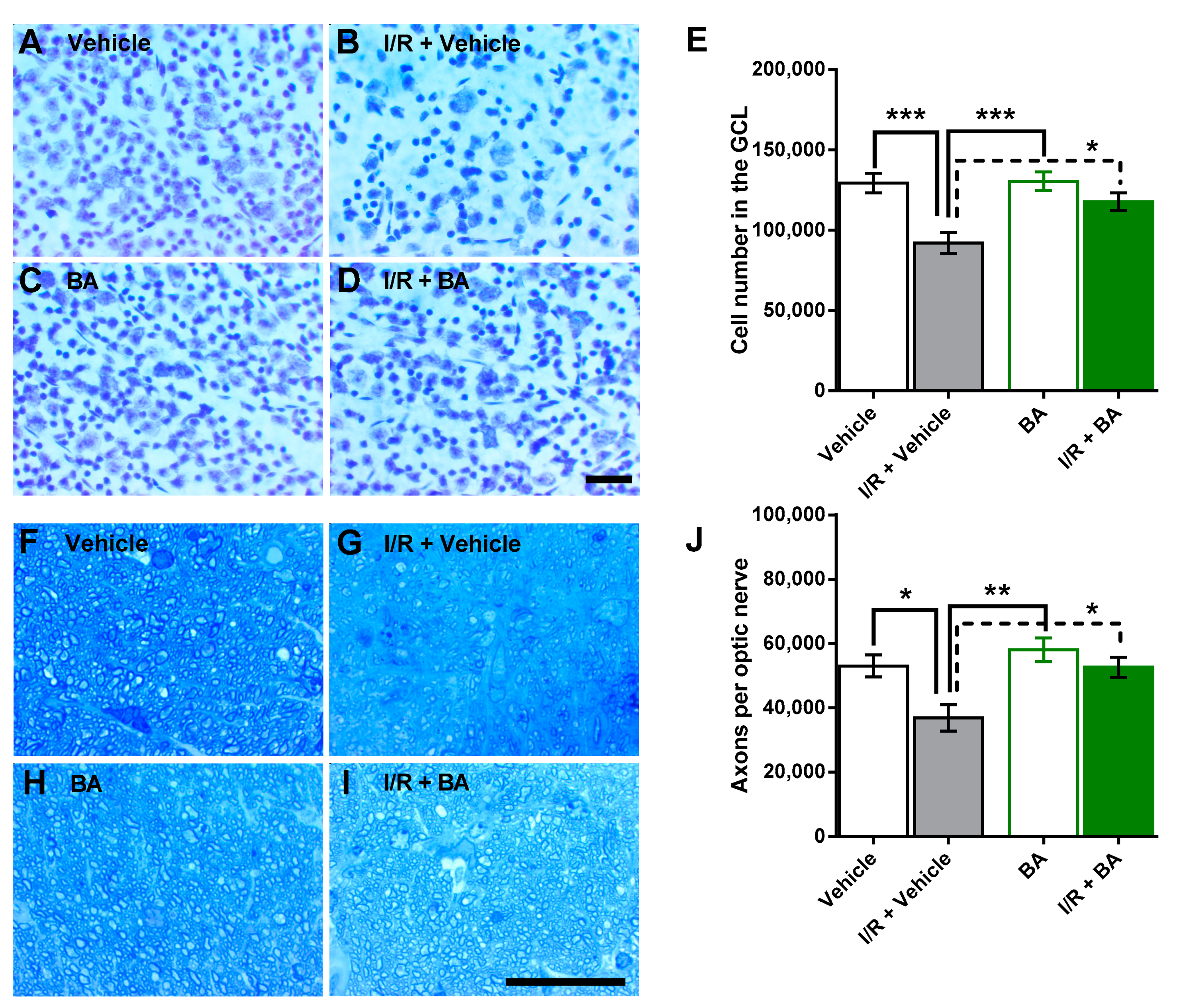
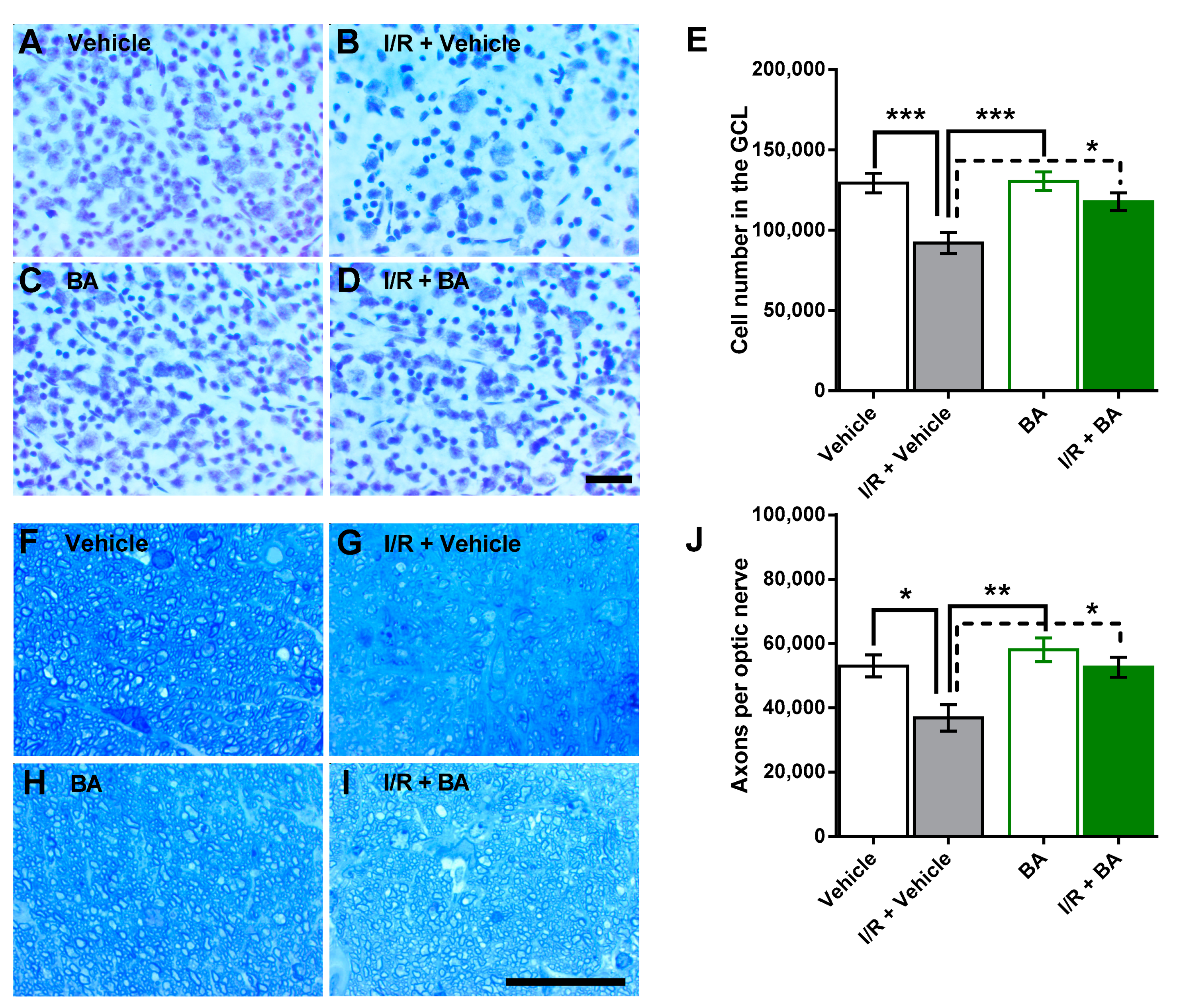
3.1. Number of Cells in the Retinal Ganglion Cell Layer and of Axons in the Optic Nerve
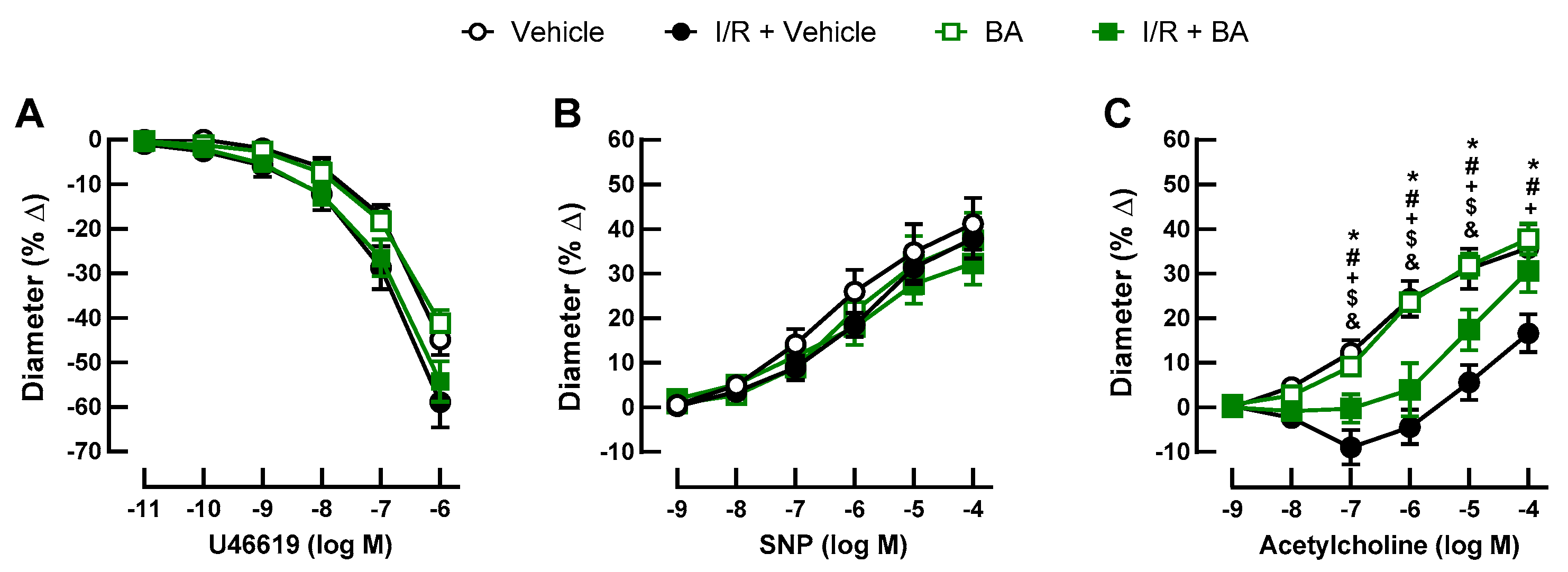
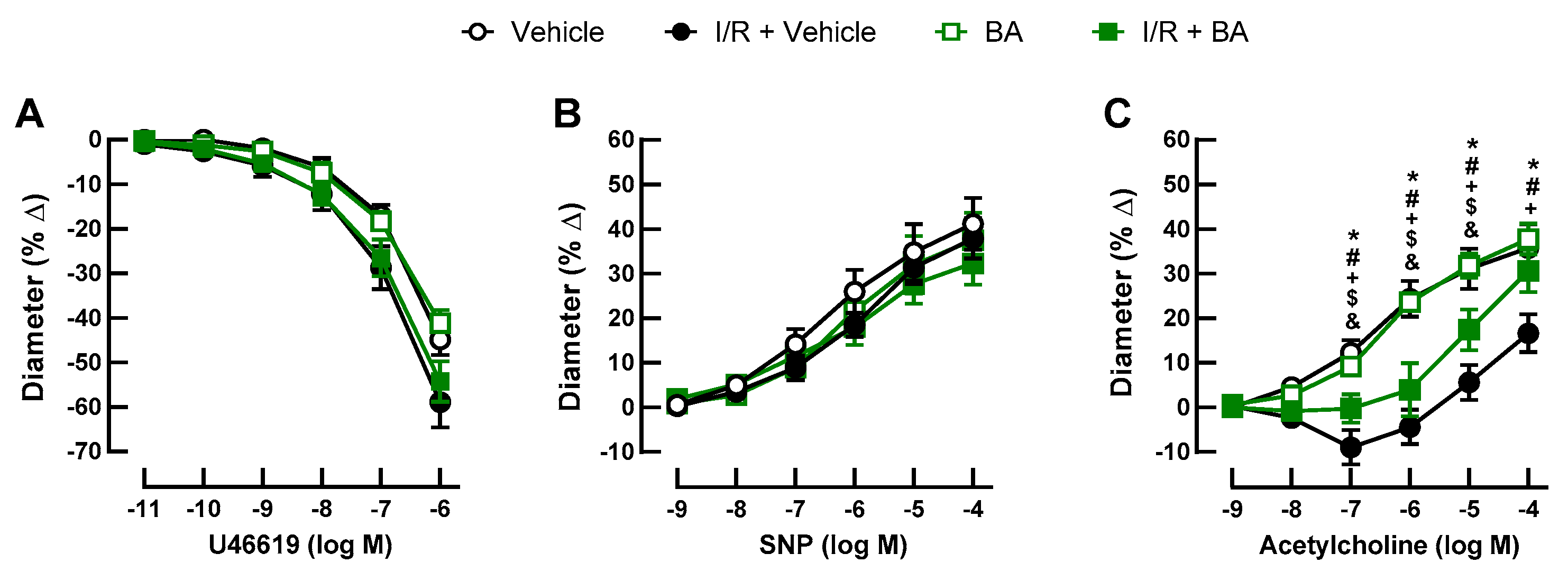
3.2. Retinal Arteriole Responses
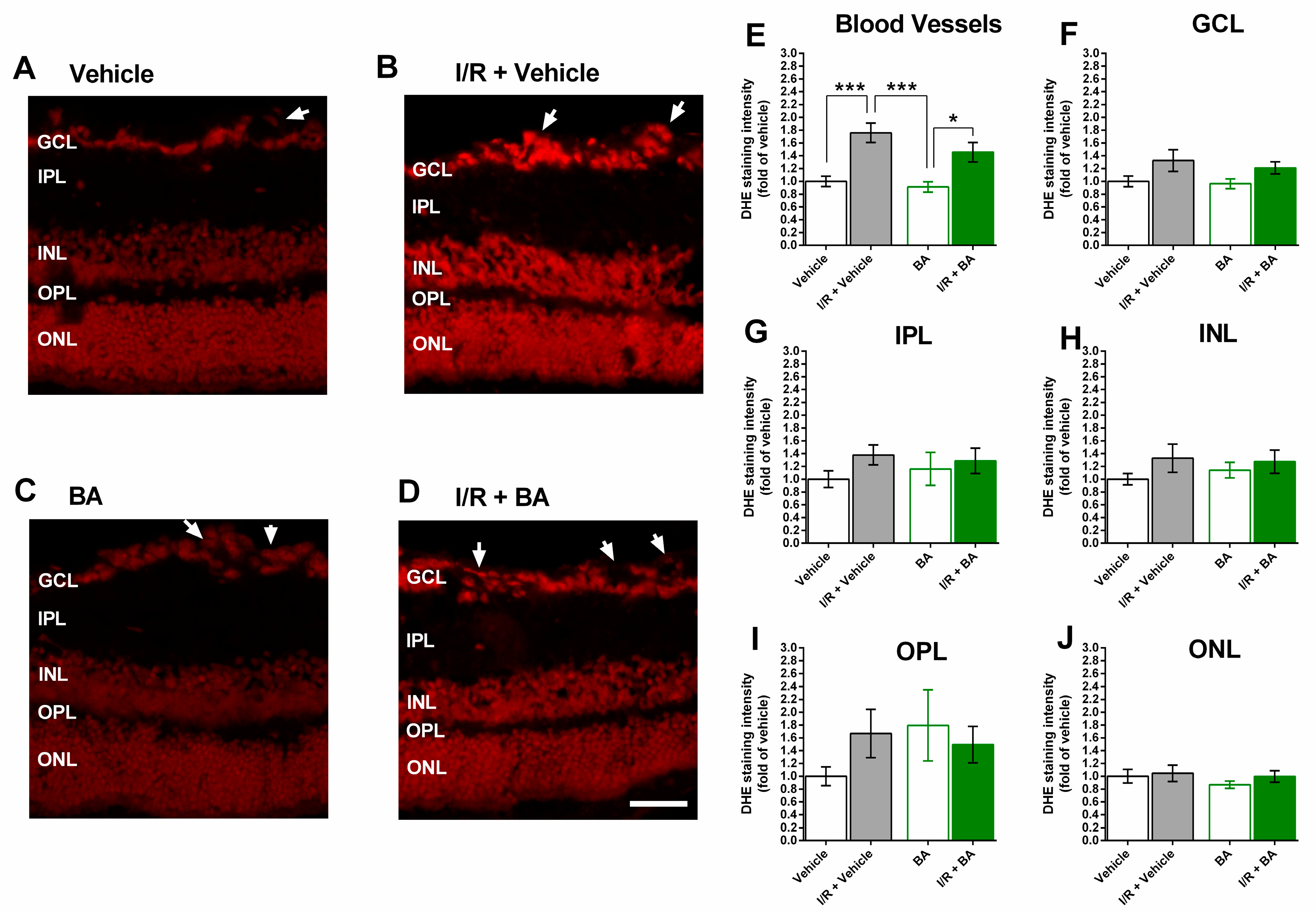
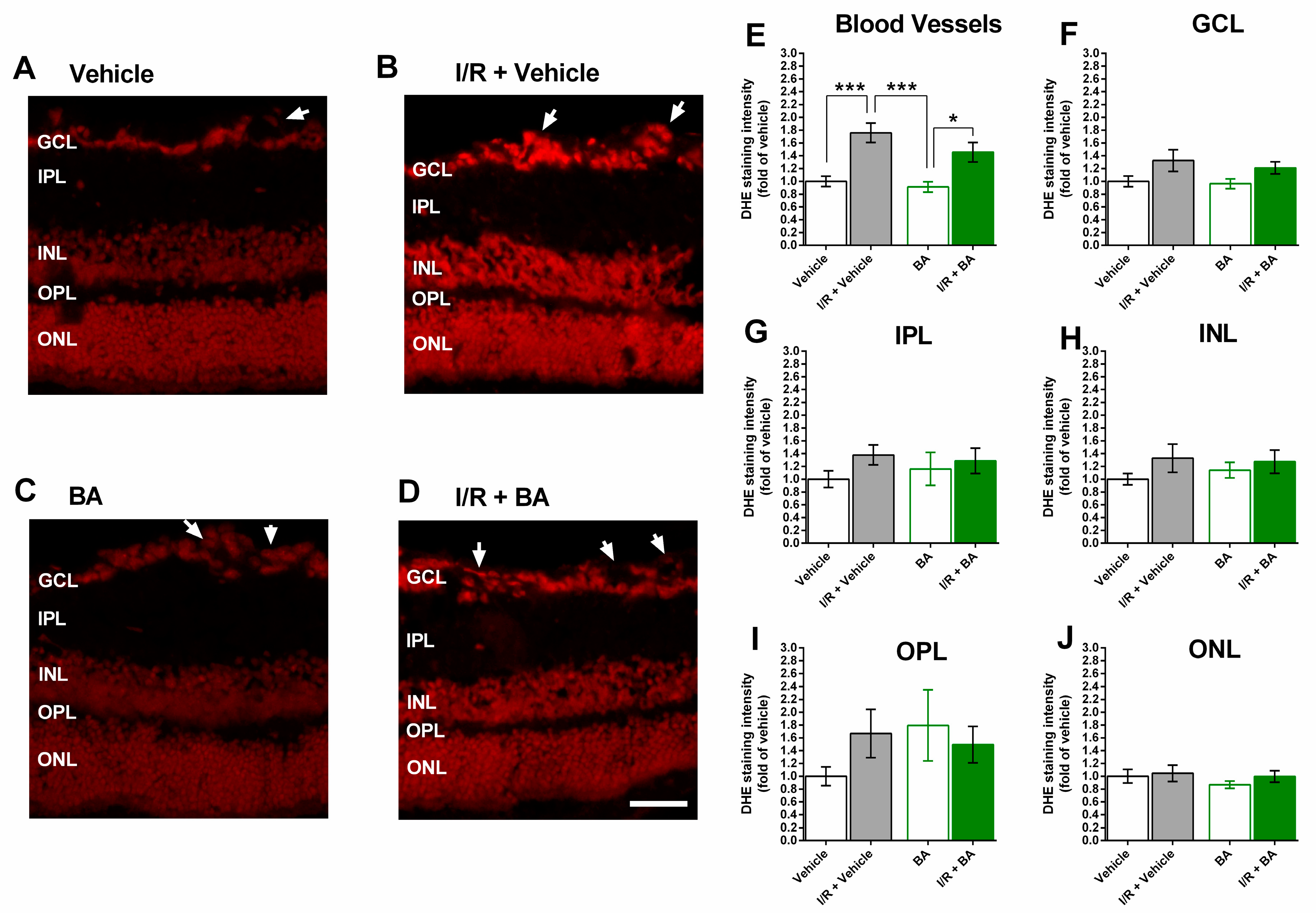
3.3. ROS Levels in the Retina
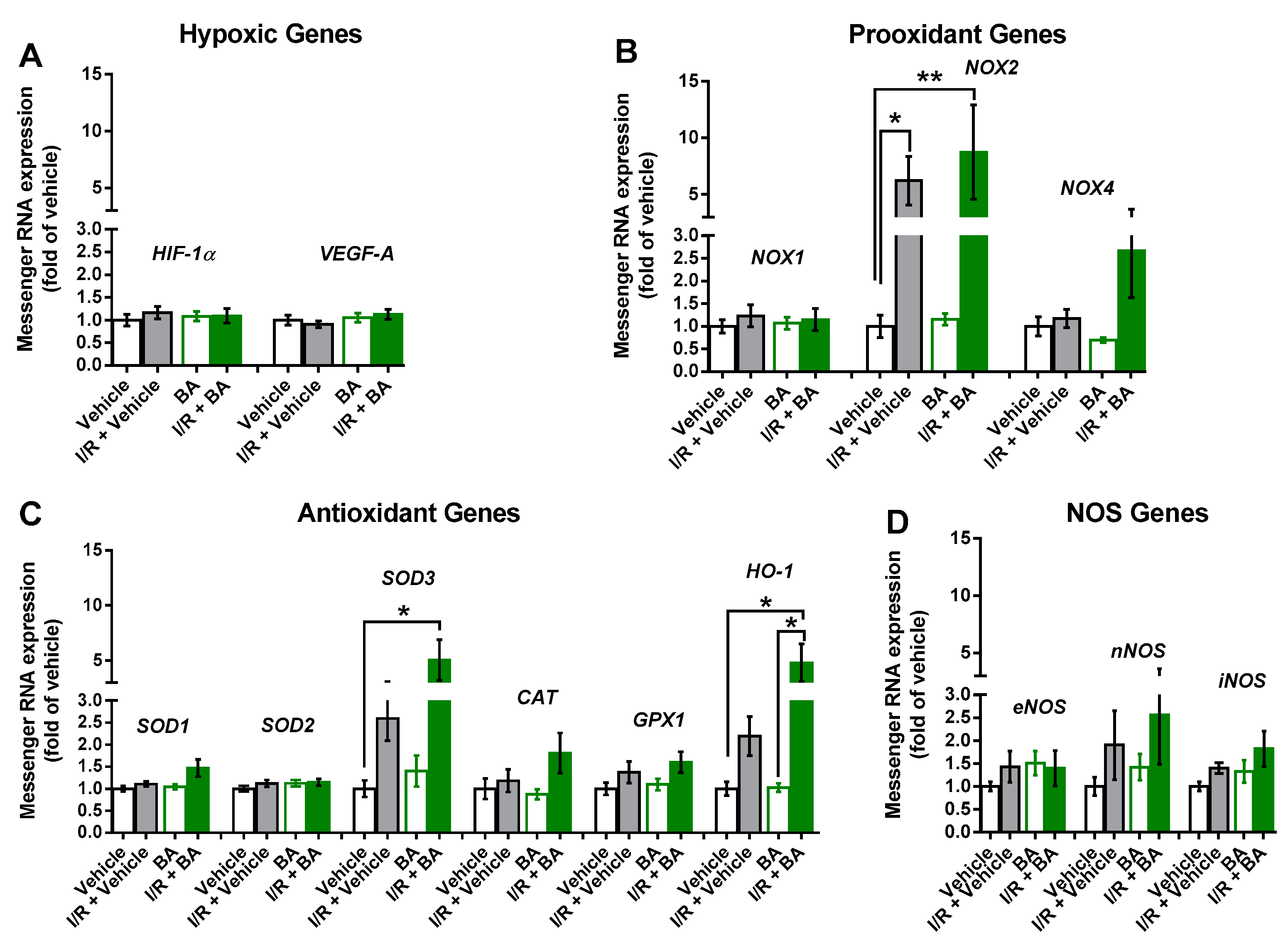
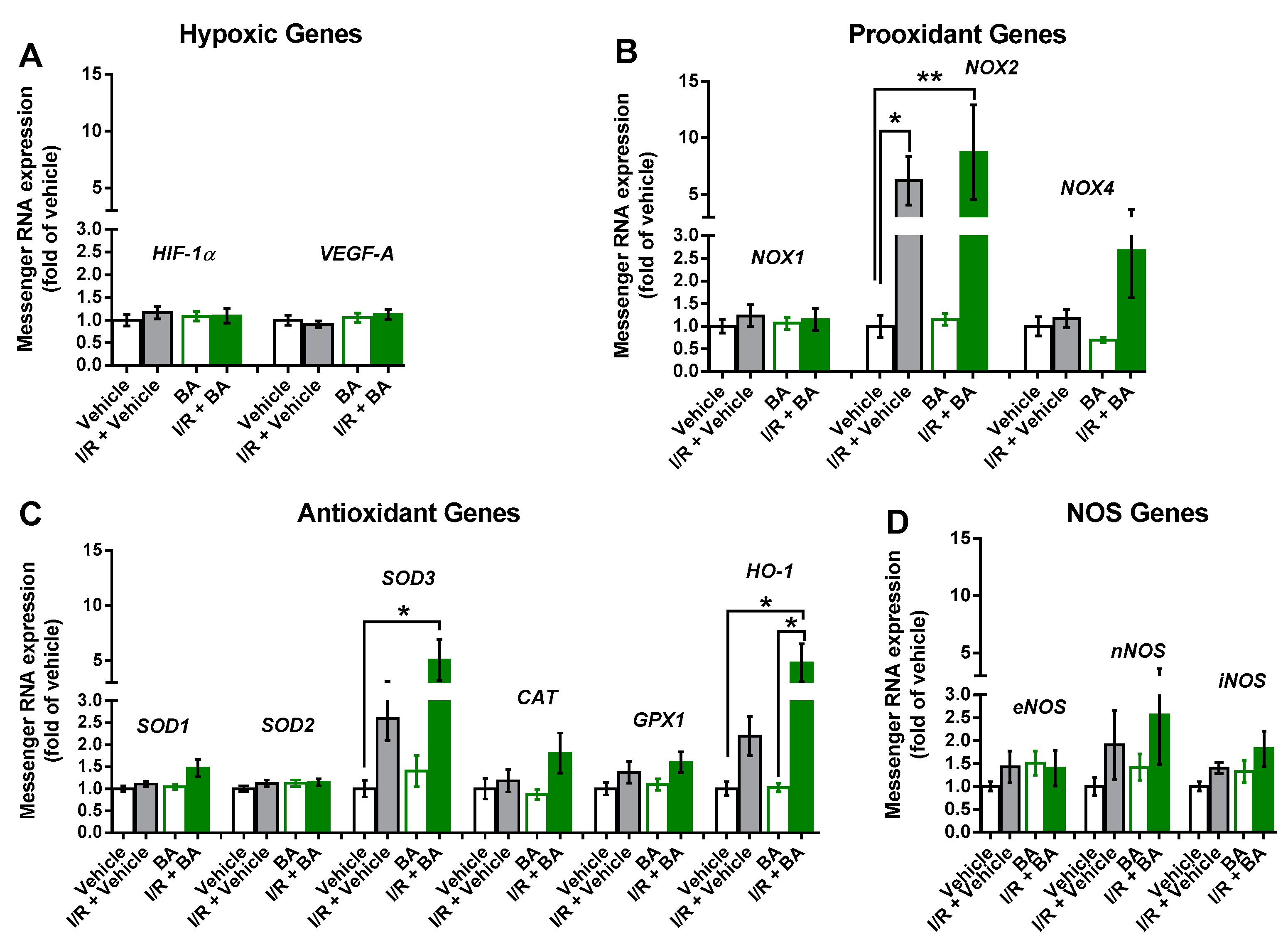
3.4. Messenger RNA Expression in the Retina
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ruan, Y.; Jiang, S.; Musayeva, A.; Gericke, A. Oxidative Stress and Vascular Dysfunction in the Retina: Therapeutic Strategies. Antioxidants 2020, 9, 761. [Google Scholar] [CrossRef] [PubMed]
- Rumelt, S.; Dorenboim, Y.; Rehany, U. Aggressive systematic treatment for central retinal artery occlusion. Am. J. Ophthalmol. 1999, 128, 733–738. [Google Scholar] [CrossRef]
- Terelak-Borys, B.; Skonieczna, K.; Grabska-Liberek, I. Ocular ischemic syndrome—A systematic review. Med. Sci. Monit. 2012, 18, RA138–RA144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feltgen, N.; Neubauer, A.; Jurklies, B.; Schmoor, C.; Schmidt, D.; Wanke, J.; Maier-Lenz, H.; Schumacher, M. Multicenter study of the European Assessment Group for Lysis in the Eye (EAGLE) for the treatment of central retinal artery occlusion: Design issues and implications. EAGLE Study report no. 1: EAGLE Study report no. 1. Graefes Arch. Clin. Exp. Ophthalmol. 2006, 244, 950–956. [Google Scholar] [CrossRef]
- Schumacher, M.; Schmidt, D.; Jurklies, B.; Gall, C.; Wanke, I.; Schmoor, C.; Maier-Lenz, H.; Solymosi, L.; Brueckmann, H.; Neubauer, A.S.; et al. Central Retinal Artery Occlusion: Local Intra-arterial Fibrinolysis versus Conservative Treatment, a Multicenter Randomized Trial. Ophthalmology 2010, 117, 1367–1375.e1. [Google Scholar] [CrossRef] [PubMed]
- Zadeh, J.K.; Garcia-Bardon, A.; Hartmann, E.K.; Pfeiffer, N.; Omran, W.; Ludwig, M.; Patzak, A.; Xia, N.; Li, H.; Gericke, A. Short-Time Ocular Ischemia Induces Vascular Endothelial Dysfunction and Ganglion Cell Loss in the Pig Retina. Int. J. Mol. Sci. 2019, 20, 4685. [Google Scholar] [CrossRef] [Green Version]
- Zadeh, J.K.; Ruemmler, R.; Hartmann, E.K.; Ziebart, A.; Ludwig, M.; Patzak, A.; Xia, N.; Li, H.; Pfeiffer, N.; Gericke, A. Responses of retinal arterioles and ciliary arteries in pigs with acute respiratory distress syndrome (ARDS). Exp. Eye Res. 2019, 184, 152–161. [Google Scholar] [CrossRef]
- Yokota, H.; Narayanan, P.; Zhang, W.; Liu, H.; Rojas, M.; Xu, Z.; Lemtalsi, T.; Nagaoka, T.; Yoshida, A.; Brooks, S.E.; et al. Neuroprotection from Retinal Ischemia/Reperfusion Injury by NOX2 NADPH Oxidase Deletion. Investig. Opthalmol. Vis. Sci. 2011, 52, 8123–8131. [Google Scholar] [CrossRef]
- Qin, X.; Li, N.; Zhang, M.; Lin, S.; Zhu, J.; Xiao, D.; Cui, W.; Zhang, T.; Lin, Y.; Cai, X. Tetrahedral framework nucleic acids prevent retina ischemia-reperfusion injury from oxidative stress via activating the Akt/Nrf2 pathway. Nanoscale 2019, 11, 20667–20675. [Google Scholar] [CrossRef]
- Jäger, S.; Trojan, H.; Kopp, T.; Laszczyk, M.N.; Scheffler, A. Pentacyclic Triterpene Distribution in Various Plants—Rich Sources for a New Group of Multi-Potent Plant Extracts. Molecules 2009, 14, 2016–2031. [Google Scholar] [CrossRef] [Green Version]
- Trumbull, E.R.; Bianchi, E.; Eckert, D.J.; Wiedhopf, R.M.; Cole, J.R. Tumor Inhibitory Agents from Vauquelinia corymbosa (Rosaceae). J. Pharm. Sci. 1976, 65, 1407–1408. [Google Scholar] [CrossRef]
- Jiang, W.; Li, X.; Dong, S.; Zhou, W. Betulinic acid in the treatment of tumour diseases: Application and research progress. Biomed. Pharmacother. 2021, 142, 111990. [Google Scholar] [CrossRef]
- Yogeeswari, P. Betulinic Acid and Its Derivatives: A Review on their Biological Properties. Curr. Med. Chem. 2005, 12, 657–666. [Google Scholar] [CrossRef]
- Żwawiak, J.; Pawełczyk, A.; Olender, D.; Zaprutko, L. Structure and Activity of Pentacyclic Triterpenes Codrugs. A Review. Mini-Rev. Med. Chem. 2021, 21, 1–21. [Google Scholar] [CrossRef]
- Lu, Q.; Xia, N.; Xu, H.; Guo, L.; Wenzel, P.; Daiber, A.; Münzel, T.; Förstermann, U.; Li, H. Betulinic acid protects against cerebral ischemia–reperfusion injury in mice by reducing oxidative and nitrosative stress. Nitric Oxide 2011, 24, 132–138. [Google Scholar] [CrossRef]
- Xia, A.; Xue, Z.; Li, Y.; Wang, W.; Xia, J.; Wei, T.; Cao, J.; Zhou, W. Cardioprotective Effect of Betulinic Acid on Myocardial Ischemia Reperfusion Injury in Rats. Evid.-Based Complement. Altern. Med. 2014, 2014, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekşioğlu-Demiralp, E.; Kardaş, E.R.; Özgül, S.; Yağcı, T.; Bilgin, H.; Şehirli, Ö.; Ercan, F.; Şener, G. Betulinic acid protects against ischemia/reperfusion-induced renal damage and inhibits leukocyte apoptosis. Phytother. Res. 2010, 24, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Zhang, C.-C.; Zhang, X.-M.; Li, H.-G.; Luo, A.-L.; Tian, Y.-K.; Xu, H. Down-regulation of NOX4 by betulinic acid protects against cerebral ischemia-reperfusion in mice. Curr. Med. Sci. 2017, 37, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Shi, X.; Wang, J.; Mang, J.; Xu, Z. Betulinic Acid Ameliorates Cerebral Injury in Middle Cerebral Artery Occlusion Rats through Regulating Autophagy. ACS Chem. Neurosci. 2021, 12, 2829–2837. [Google Scholar] [CrossRef]
- Hartsock, M.J.; Cho, H.; Wu, L.; Chen, W.-J.; Gong, J.; Duh, E.J. A Mouse Model of Retinal Ischemia-Reperfusion Injury Through Elevation of Intraocular Pressure. J. Vis. Exp. 2016, e54065. [Google Scholar] [CrossRef] [PubMed]
- Hein, T.W.; Ren, Y.; Potts, L.B.; Yuan, Z.; Kuo, E.; Rosa, R.H.; Kuo, L. Acute Retinal Ischemia Inhibits Endothelium-Dependent Nitric Oxide–Mediated Dilation of Retinal Arterioles via Enhanced Superoxide Production. Investig. Opthalmol. Vis. Sci. 2012, 53, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Laspas, P.; Zhutdieva, M.B.; Brochhausen, C.; Musayeva, A.; Zadeh, J.K.; Pfeiffer, N.; Xia, N.; Li, H.; Wess, J.; Gericke, A. The M1 muscarinic acetylcholine receptor subtype is important for retinal neuron survival in aging mice. Sci. Rep. 2019, 9, 5222. [Google Scholar] [CrossRef] [PubMed]
- Gericke, A.; Sniatecki, J.J.; Goloborodko, E.; Steege, A.; Zavaritskaya, O.; Vetter, J.M.; Grus, F.H.; Patzak, A.; Wess, J.; Pfeiffer, N. Identification of the Muscarinic Acetylcholine Receptor Subtype Mediating Cholinergic Vasodilation in Murine Retinal Arterioles. Investig. Opthalmol. Vis. Sci. 2011, 52, 7479–7484. [Google Scholar] [CrossRef]
- Gericke, A.; Goloborodko, E.; Pfeiffer, N.; Manicam, C. Preparation Steps for Measurement of Reactivity in Mouse Retinal Arterioles Ex Vivo. J. Vis. Exp. 2018, e56199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gericke, A.; Mann, C.; Zadeh, J.K.; Musayeva, A.; Wolff, I.; Wang, M.; Pfeiffer, N.; Daiber, A.; Li, H.; Xia, N.; et al. Elevated Intraocular Pressure Causes Abnormal Reactivity of Mouse Retinal Arterioles. Oxidative Med. Cell. Longev. 2019, 2019, 9736047. [Google Scholar] [CrossRef] [Green Version]
- Zadeh, J.K.; Zhutdieva, M.B.; Laspas, P.; Yüksel, C.; Musayeva, A.; Pfeiffer, N.; Brochhausen, C.; Oelze, M.; Daiber, A.; Xia, N.; et al. Apolipoprotein E Deficiency Causes Endothelial Dysfunction in the Mouse Retina. Oxidative Med. Cell. Longev. 2019, 2019, 5181429. [Google Scholar] [CrossRef] [Green Version]
- Musayeva, A.; Jiang, S.; Ruan, Y.; Zadeh, J.; Chronopoulos, P.; Pfeiffer, N.; Müller, W.; Ackermann, M.; Xia, N.; Li, H.; et al. Aged Mice Devoid of the M3 Muscarinic Acetylcholine Receptor Develop Mild Dry Eye Disease. Int. J. Mol. Sci. 2021, 22, 6133. [Google Scholar] [CrossRef] [PubMed]
- Dräger, U.C.; Olsen, J.F. Ganglion cell distribution in the retina of the mouse. Investig. Ophthalmol. Vis. Sci. 1981, 20, 285–293. [Google Scholar]
- Schlamp, C.L.; Montgomery, A.D.; Mac Nair, C.E.; Schuart, C.; Willmer, D.J.; Nickells, R.W. Evaluation of the percentage of ganglion cells in the ganglion cell layer of the rodent retina. Mol. Vis. 2013, 19, 1387–1396. [Google Scholar]
- Choi, J.; Kook, M.S. Systemic and Ocular Hemodynamic Risk Factors in Glaucoma. BioMed. Res. Int. 2015, 2015, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Flammer, J.; Mozaffarieh, M. What is the present pathogenetic concept of glaucomatous optic neuropathy? Surv. Ophthalmol. 2007, 52 (Suppl. 2), S162–S173. [Google Scholar] [CrossRef]
- Resch, H.; Garhofer, G.; Fuchsjäger-Mayrl, G.; Hommer, A.; Schmetterer, L. Endothelial dysfunction in glaucoma. Acta Ophthalmol. 2009, 87, 4–12. [Google Scholar] [CrossRef]
- Bai, Y.-Y.; Yan, D.; Zhou, H.-Y.; Li, W.-X.; Lou, Y.-Y.; Zhou, X.-R.; Qian, L.-B.; Xiao, C. Betulinic acid attenuates lipopolysaccharide-induced vascular hyporeactivity in the rat aorta by modulating Nrf2 antioxidative function. Inflammopharmacology 2020, 28, 165–174. [Google Scholar] [CrossRef]
- Xia, M.-L.; Qian, L.-B.; Fu, J.-Y.; Cai, X. Betulinic acid inhibits superoxide anion-mediated impairment of endothelium-dependent relaxation in rat aortas. Indian J. Pharmacol. 2012, 44, 588–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.-Y.; Qian, L.-B.; Zhu, L.-G.; Liang, H.-T.; Tan, Y.-N.; Lu, H.-T.; Lu, J.-F.; Wang, H.-P.; Xia, Q. Betulinic acid ameliorates endothelium-dependent relaxation in l-NAME-induced hypertensive rats by reducing oxidative stress. Eur. J. Pharm. Sci. 2011, 44, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Fan, N.; Silverman, S.M.; Liu, Y.; Wang, X.; Kim, B.-J.; Tang, L.; Clark, A.F.; Liu, X.; Pang, I.-H. Rapid repeatable in vivo detection of retinal reactive oxygen species. Exp. Eye Res. 2017, 161, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Wert, K.; Velez, G.; Cross, M.R.; Wagner, B.A.; Teoh-Fitzgerald, M.L.; Buettner, G.R.; McAnany, J.J.; Olivier, A.; Tsang, S.H.; Harper, M.; et al. Extracellular superoxide dismutase (SOD3) regulates oxidative stress at the vitreoretinal interface. Free. Radic. Biol. Med. 2018, 124, 408–419. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y. Therapeutic Potential of Heme Oxygenase-1/carbon Monoxide System Against Ischemia-Reperfusion Injury. Curr. Pharm. Des. 2017, 23, 3884–3898. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| NOX1 | GGAGGAATTAGGCAAAATGGATT | GCTGCATGACCAGCAATGTT |
| NOX2 | CCAACTGGGATAACGAGTTCA | GAGAGTTTCAGCCAAGGCTTC |
| NOX4 | TGTAACAGAGGGAAAACAGTTGGA | GTTCCGGTTACTCAAACTATGAAGAGT |
| eNOS | CCTTCCGCTACCAGCCAGA | CAGAGATCTTCACTGCATTGGCTA |
| iNOS | CAGCTGGGCTGTACAAACCTT | CATTGGAAGTGAAGCGTTTCG |
| nNOS | TCCACCTGCCTCGAAACC | TTGTCGCTGTTGCCAAAAAC |
| CAT | CAAGTACAACGCTGAGAAGCCTAAG | CCCTTCGCAGCCATGTG |
| GPX1 | CCCGTGCGCAGGTACAG | GGGACAGCAGGGTTTCTATGTC |
| HO-1 | GGTGATGCTGACAGAGGAACAC | TAGCAGGCCTCTGACGAAGTG |
| SOD1 | CCAGTGCAGGACCTCATTTTAAT | TCTCCAACATGCCTCTCTTCATC |
| SOD2 | CCTGCTCTAATCAGGACCCATT | CGTGCTCCCACACGTCAAT |
| SOD3 | TTCTTGTTCTACGGCTTGCTACTG | AGCTGGACTCCCCTGGATTT |
| TBP | CTTCGTGCAAGAAATGCTGAAT | CAGTTGTCCGTGGCTCTCTTATT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Musayeva, A.; Unkrig, J.C.; Zhutdieva, M.B.; Manicam, C.; Ruan, Y.; Laspas, P.; Chronopoulos, P.; Göbel, M.L.; Pfeiffer, N.; Brochhausen, C.; et al. Betulinic Acid Protects from Ischemia-Reperfusion Injury in the Mouse Retina. Cells 2021, 10, 2440. https://doi.org/10.3390/cells10092440
Musayeva A, Unkrig JC, Zhutdieva MB, Manicam C, Ruan Y, Laspas P, Chronopoulos P, Göbel ML, Pfeiffer N, Brochhausen C, et al. Betulinic Acid Protects from Ischemia-Reperfusion Injury in the Mouse Retina. Cells. 2021; 10(9):2440. https://doi.org/10.3390/cells10092440
Chicago/Turabian StyleMusayeva, Aytan, Johanna C. Unkrig, Mayagozel B. Zhutdieva, Caroline Manicam, Yue Ruan, Panagiotis Laspas, Panagiotis Chronopoulos, Marie L. Göbel, Norbert Pfeiffer, Christoph Brochhausen, and et al. 2021. "Betulinic Acid Protects from Ischemia-Reperfusion Injury in the Mouse Retina" Cells 10, no. 9: 2440. https://doi.org/10.3390/cells10092440
APA StyleMusayeva, A., Unkrig, J. C., Zhutdieva, M. B., Manicam, C., Ruan, Y., Laspas, P., Chronopoulos, P., Göbel, M. L., Pfeiffer, N., Brochhausen, C., Daiber, A., Oelze, M., Li, H., Xia, N., & Gericke, A. (2021). Betulinic Acid Protects from Ischemia-Reperfusion Injury in the Mouse Retina. Cells, 10(9), 2440. https://doi.org/10.3390/cells10092440