
β-Adrenergic Stimulation Synchronizes a Broad Spectrum of Action Potential Firing Rates of Cardiac Pacemaker Cells toward a Higher Population Average


Abstract
1. Introduction
2. Materials and Methods
2.1. Single Cell Preparation
- We accurately dissected the SA node to avoid (or minimize) contamination with atrial myocytes. The location of guinea pig SA nodes in relation to the whole heart during our isolation procedure is shown in Figure S1.
- We measured SA node cells that had a classical spindle-shaped morphology (Figure S2).
- We deliberately avoided cells with a typical atrial cell morphology (i.e., blocky and rectangular in shape).
- We described the electrophysiological properties of these cells in our recent study [12]. The cells used in the present study have similar properties because they were from the same isolations (from the same guinea pig hearts). The maximum diastolic potential was on average −58.5 mV. This is clearly different from SA node-residing atrial cells that have a stable resting membrane potential between −70 and −80 mV [21].
2.2. Two-Dimensional (2D) Ca Imaging of Single Cells
2.3. Detection and Analysis of the Whole Ensemble of AP-Induced Cytosolic Ca Transients and LCRs in 2D Ca Imaging
2.4. Evaluation of βAR Stimulation Effect
2.5. Statistics
3. Results
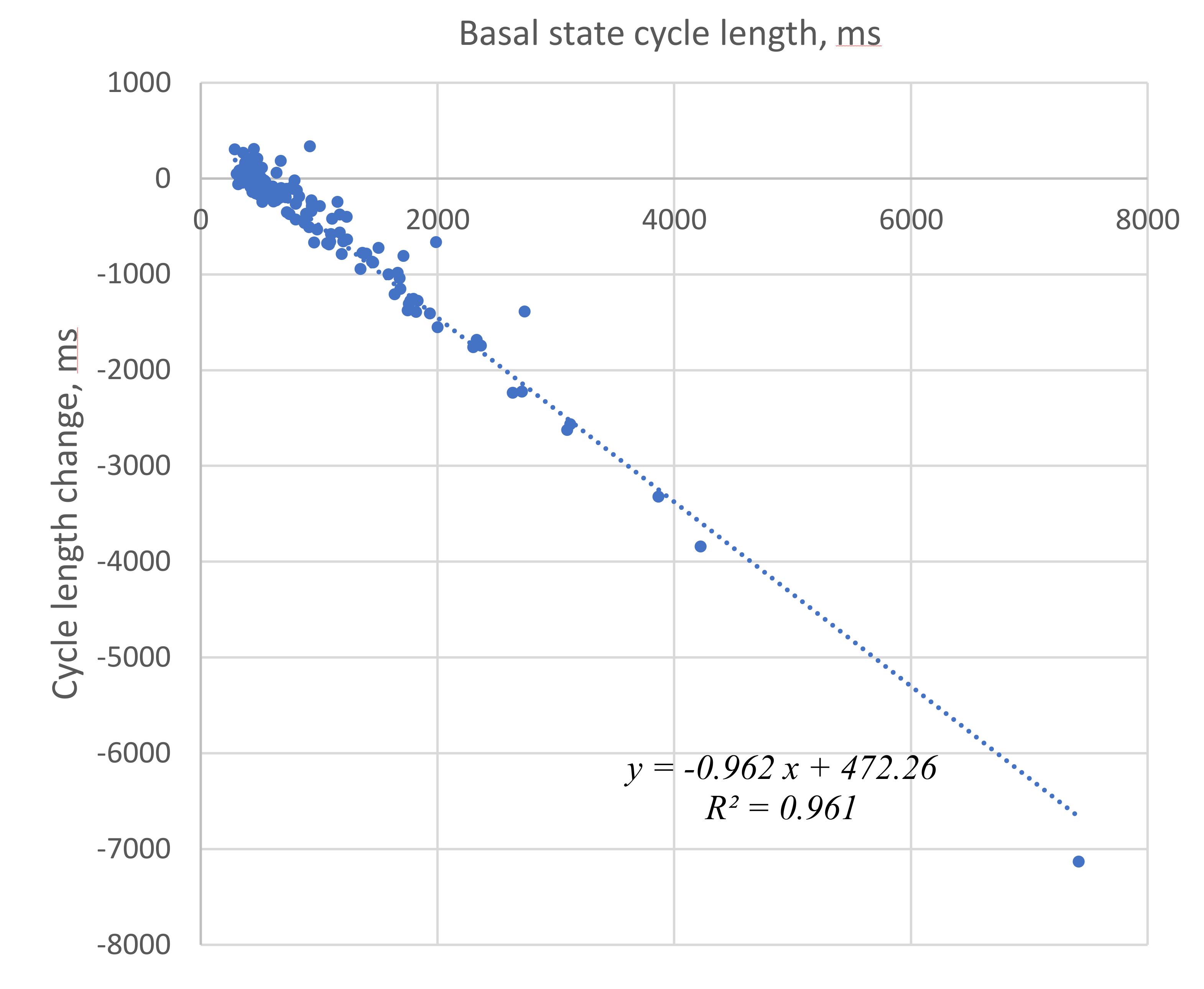
3.1. Cycle Length Changes
3.2. LCR Changes
4. Discussion
4.1. Result Summary
4.2. Possible Mechanisms Underlying Different Cell Sensitivity to βAR Stimulation
4.3. A Broader Interpretation of Our Results toward the Entire Range of Autonomic Modulation, Including Dormant Cells
4.4. A Broader Interpretation of Our Results toward Synchronization Mechanisms of Cell Signaling in Intact SAN: Cell Specialization and Local Control by Autonomic System
4.5. Importance of Beat-to-Beat Variability
4.6. Functional Benefits of Heterogeneity at Tissue Level
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Mangoni, M.E.; Nargeot, J. Genesis and regulation of the heart automaticity. Physiol. Rev. 2008, 88, 919–982. [Google Scholar] [CrossRef]
- Di Francesco, D. Pacemaker mechanisms in cardiac tissue. Annu. Rev. Physiol. 1993, 55, 455–472. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, T.; Kurachi, Y.; Noma, A.; Irisawa, H. Action potential and membrane currents of single pacemaker cells of the rabbit heart. Pflug. Arch. 1984, 402, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Sano, T.; Sawanobori, T.; Adaniya, H. Mechanism of rhythm determination among pacemaker cells of the mammalian sinus node. Am. J. Physiol. 1978, 235, H379–H384. [Google Scholar] [CrossRef] [PubMed]
- Bleeker, W.K.; Mackaay, A.J.; Masson-Pevet, M.; Bouman, L.N.; Becker, A.E. Functional and morphological organization of the rabbit sinus node. Circ. Res. 1980, 46, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Jalife, J. Mutual entrainment and electrical coupling as mechanisms for synchronous firing of rabbit sino-atrial pace-maker cells. J. Physiol. 1984, 356, 221–243. [Google Scholar] [CrossRef] [PubMed]
- Michaels, D.C.; Matyas, E.P.; Jalife, J. Mechanisms of sinoatrial pacemaker synchronization: A new hypothesis. Circ. Res. 1987, 61, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Bychkov, R.; Juhaszova, M.; Tsutsui, K.; Coletta, C.; Stern, M.D.; Maltsev, V.A.; Lakatta, E.G. Synchronized cardiac impulses emerge from multi-scale, heterogeneous local calcium signals within and among cells of heart pacemaker tissue. JACC Clin. Electrophysiol. 2020, 6, 907–931. [Google Scholar] [CrossRef]
- Fenske, S.; Hennis, K.; Rotzer, R.D.; Brox, V.F.; Becirovic, E.; Scharr, A.; Gruner, C.; Ziegler, T.; Mehlfeld, V.; Brennan, J.; et al. cAMP-dependent regulation of HCN4 controls the tonic entrainment process in sinoatrial node pacemaker cells. Nat. Commun. 2020, 11, 5555. [Google Scholar] [CrossRef]
- Kim, M.S.; Maltsev, A.V.; Monfredi, O.; Maltseva, L.A.; Wirth, A.; Florio, M.C.; Tsutsui, K.; Riordon, D.R.; Parsons, S.P.; Tagirova, S.; et al. Heterogeneity of calcium clock functions in dormant, dysrhythmically and rhythmically firing single pacemaker cells isolated from SA node. Cell Calcium 2018, 74, 168–179. [Google Scholar] [CrossRef]
- Tsutsui, K.; Monfredi, O.; Sirenko-Tagirova, S.G.; Maltseva, L.A.; Bychkov, R.; Kim, M.S.; Ziman, B.D.; Tarasov, K.V.; Tarasova, Y.S.; Zhang, J.; et al. A coupled-clock system drives the automaticity of human sinoatrial nodal pacemaker cells. Sci. Signal. 2018, 11, eaap7608. [Google Scholar] [CrossRef]
- Tsutsui, K.; Florio, M.C.; Yang, A.; Wirth, A.N.; Yang, D.; Kim, M.S.; Ziman, B.D.; Bychkov, R.; Monfredi, O.J.; Maltsev, V.A.; et al. cAMP-Dependent Signaling Restores AP Firing in Dormant SA Node Cells via Enhancement of Surface Membrane Currents and Calcium Coupling. Front. Physiol. 2021, 12, 596832. [Google Scholar] [CrossRef] [PubMed]
- Maltsev, V.A.; Lakatta, E.G. Synergism of coupled subsarcolemmal Ca2+ clocks and sarcolemmal voltage clocks confers robust and flexible pacemaker function in a novel pacemaker cell model. Am. J. Physiol. 2009, 296, H594–H615. [Google Scholar] [CrossRef] [PubMed]
- Lakatta, E.G.; Maltsev, V.A.; Vinogradova, T.M. A coupled SYSTEM of intracellular Ca2+ clocks and surface membrane voltage clocks controls the timekeeping mechanism of the heart’s pacemaker. Circ. Res. 2010, 106, 659–673. [Google Scholar] [CrossRef]
- Lyashkov, A.E.; Behar, J.; Lakatta, E.G.; Yaniv, Y.; Maltsev, V.A. Positive Feedback Mechanisms among Local Ca Releases, NCX, and ICaL Ignite Pacemaker Action Potentials. Biophys. J. 2018, 114, 1176–1189. [Google Scholar] [CrossRef] [PubMed]
- Yaniv, Y.; Ganesan, A.; Yang, D.; Ziman, B.D.; Lyashkov, A.E.; Levchenko, A.; Zhang, J.; Lakatta, E.G. Real-time relationship between PKA biochemical signal network dynamics and increased action potential firing rate in heart pacemaker cells: Kinetics of PKA activation in heart pacemaker cells. J. Mol. Cell. Cardiol. 2015, 86, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Musa, H.; Lei, M.; Honjo, H.; Jones, S.A.; Dobrzynski, H.; Lancaster, M.K.; Takagishi, Y.; Henderson, Z.; Kodama, I.; Boyett, M.R. Heterogeneous expression of Ca2+ handling proteins in rabbit sinoatrial node. J. Histochem. Cytochem. 2002, 50, 311–324. [Google Scholar] [CrossRef]
- Honjo, H.; Boyett, M.R.; Kodama, I.; Toyama, J. Correlation between electrical activity and the size of rabbit sino-atrial node cells. J. Physiol. 1996, 496 (Pt 3), 795–808. [Google Scholar] [CrossRef]
- Monfredi, O.; Tsutsui, K.; Ziman, B.; Stern, M.D.; Lakatta, E.G.; Maltsev, V.A. Electrophysiological heterogeneity of pacemaker cells in the rabbit intercaval region, including the SA node: Insights from recording multiple ion currents in each cell. Am. J. Physiol. 2018, 314, H403–H414. [Google Scholar] [CrossRef]
- Vinogradova, T.M.; Zhou, Y.Y.; Maltsev, V.; Lyashkov, A.; Stern, M.; Lakatta, E.G. Rhythmic ryanodine receptor Ca2+ releases during diastolic depolarization of sinoatrial pacemaker cells do not require membrane depolarization. Circ. Res. 2004, 94, 802–809. [Google Scholar] [CrossRef]
- Verheijck, E.E.; Wessels, A.; van Ginneken, A.C.; Bourier, J.; Markman, M.W.; Vermeulen, J.L.; de Bakker, J.M.; Lamers, W.H.; Opthof, T.; Bouman, L.N. Distribution of atrial and nodal cells within the rabbit sinoatrial node: Models of sinoatrial transition. Circulation 1998, 97, 1623–1631. [Google Scholar] [CrossRef]
- Monfredi, O.; Maltseva, L.A.; Spurgeon, H.A.; Boyett, M.R.; Lakatta, E.G.; Maltsev, V.A. Beat-to-beat variation in periodicity of local calcium releases contributes to intrinsic variations of spontaneous cycle length in isolated single sinoatrial node cells. PLoS ONE 2013, 8, e67247. [Google Scholar] [CrossRef]
- Yaniv, Y.; Stern, M.D.; Lakatta, E.G.; Maltsev, V.A. Mechanisms of beat-to-beat regulation of cardiac pacemaker cell function by Ca2+ cycling dynamics. Biophys. J. 2013, 105, 1551–1561. [Google Scholar] [CrossRef] [PubMed]
- Maltsev, A.V.; Parsons, S.P.; Kim, M.S.; Tsutsui, K.; Stern, M.D.; Lakatta, E.G.; Maltsev, V.A.; Monfredi, O. Computer algorithms for automated detection and analysis of local Ca2+ releases in spontaneously beating cardiac pacemaker cells. PLoS ONE 2017, 12, e0179419. [Google Scholar] [CrossRef]
- Vinogradova, T.M.; Bogdanov, K.Y.; Lakatta, E.G. beta-Adrenergic stimulation modulates ryanodine receptor Ca2+ release during diastolic depolarization to accelerate pacemaker activity in rabbit sinoatrial nodal cells. Circ. Res. 2002, 90, 73–79. [Google Scholar] [CrossRef]
- Lyashkov, A.E.; Juhaszova, M.; Dobrzynski, H.; Vinogradova, T.M.; Maltsev, V.A.; Juhasz, O.; Spurgeon, H.A.; Sollott, S.J.; Lakatta, E.G. Calcium cycling protein density and functional importance to automaticity of isolated sinoatrial nodal cells are independent of cell size. Circ. Res. 2007, 100, 1723–1731. [Google Scholar] [CrossRef] [PubMed]
- Maltsev, V.A.; Lakatta, E.G. Numerical models based on a minimal set of sarcolemmal electrogenic proteins and an intracellular Ca clock generate robust, flexible, and energy-efficient cardiac pacemaking. J. Mol. Cell. Cardiol. 2013, 59, 181–195. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nivala, M.; Ko, C.Y.; Nivala, M.; Weiss, J.N.; Qu, Z. Criticality in intracellular calcium signaling in cardiac myocytes. Biophys. J. 2012, 102, 2433–2442. [Google Scholar] [CrossRef] [PubMed]
- Maltsev, A.V.; Maltsev, V.A.; Mikheev, M.; Maltseva, L.A.; Sirenko, S.G.; Lakatta, E.G.; Stern, M.D. Synchronization of stochastic Ca2+ release units creates a rhythmic Ca2+ clock in cardiac pacemaker cells. Biophys. J. 2011, 100, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.N.; Qu, Z. The Sinus Node: Still Mysterious After All These Years. JACC Clin. Electrophysiol. 2020, 6, 1841–1843. [Google Scholar] [CrossRef]
- Bak, P. How Nature Works: The Science of Self-Organized Criticality; Springer: New York, NY, USA, 1999. [Google Scholar]
- Yang, D.; Morrell, C.H.; Lyashkov, A.E.; Tagirova, S.; Zahanich, I.; Yaniv, Y.; Vinogradova, T.; Ziman, B.; Maltsev, V.A.; Lakatta, E.G. Ca2+ and Membrane Potential Transitions During Action Potentials are Self-Similar to Each Other and to Variability of AP Firing Intervals Across the Broad Physiologic Range of AP Intervals During Autonomic Receptor Stimulation. Front. Physiol. 2021, in press. [Google Scholar] [CrossRef]
- Tagirova Sirenko, S.; Tsutsui, K.; Tarasov, K.V.; Yang, D.; Wirth, A.N.; Maltsev, V.A.; Ziman, B.D.; Yaniv, Y.; Lakatta, E.G. Self-Similar Synchronization of Calcium and Membrane Potential Transitions During Action Potential Cycles Predict Heart Rate Across Species. JACC Clin. Electrophysiol. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Lyashkov, A.E.; Vinogradova, T.M.; Zahanich, I.; Li, Y.; Younes, A.; Nuss, H.B.; Spurgeon, H.A.; Maltsev, V.A.; Lakatta, E.G. Cholinergic receptor signaling modulates spontaneous firing of sinoatrial nodal cells via integrated effects on PKA-dependent Ca2+ cycling and IKACh. Am. J. Physiol. 2009, 297, H949–H959. [Google Scholar] [CrossRef] [PubMed]
- Lang, D.; Glukhov, A.V. Functional Microdomains in Heart’s Pacemaker: A Step Beyond Classical Electrophysiology and Remodeling. Front. Physiol. 2018, 9, 1686. [Google Scholar] [CrossRef] [PubMed]
- Younes, A.; Lyashkov, A.E.; Graham, D.; Sheydina, A.; Volkova, M.V.; Mitsak, M.; Vinogradova, T.M.; Lukyanenko, Y.O.; Li, Y.; Ruknudin, A.M.; et al. Ca2+-stimulated basal adenylyl cyclase activity localization in membrane lipid microdomains of cardiac sinoatrial nodal pacemaker cells. J. Biol. Chem. 2008, 283, 14461–14468. [Google Scholar] [CrossRef]
- Dokos, S.; Celler, B.G.; Lovell, N.H. Vagal control of sinoatrial rhythm: A mathematical model. J. Theor. Biol. 1996, 182, 21–44. [Google Scholar] [CrossRef]
- Demir, S.S.; Clark, J.W.; Giles, W.R. Parasympathetic modulation of sinoatrial node pacemaker activity in rabbit heart: A unifying model. Am. J. Physiol. 1999, 276, H2221–H2244. [Google Scholar] [CrossRef]
- Zhang, H.; Holden, A.V.; Noble, D.; Boyett, M.R. Analysis of the chronotropic effect of acetylcholine on sinoatrial node cells. J. Cardiovasc. Electrophysiol. 2002, 13, 465–474. [Google Scholar] [CrossRef]
- Himeno, Y.; Sarai, N.; Matsuoka, S.; Noma, A. Ionic mechanisms underlying the positive chronotropy induced by beta1-adrenergic stimulation in guinea pig sinoatrial node cells: A simulation study. J. Physiol. Sci. 2008, 58, 53–65. [Google Scholar] [CrossRef]
- Maltsev, V.A.; Lakatta, E.G. A novel quantitative explanation for autonomic modulation of cardiac pacemaker cell automaticity via a dynamic system of sarcolemmal and intracellular proteins. Am. J. Physiol. 2010, 298, H2010–H2023. [Google Scholar] [CrossRef]
- Yuan, X.; Ratajczyk, L.N.; Alvarado, F.; Valdivia, H.H.; Glukhov, A.V.; Lang, D. Hierarchical Pacemaker Clustering within the Rabbit Sinoatrial Node is Driven by Dynamic Interaction between the Components of the Coupled-Clock System. Biophys. J. 2020, 118, 345A. [Google Scholar] [CrossRef]
- Brennan, J.A.; Chen, Q.; Gams, A.; Dyavanapalli, J.; Mendelowitz, D.; Peng, W.; Efimov, I.R. Evidence of Superior and Inferior Sinoatrial Nodes in the Mammalian Heart. JACC Clin. Electrophysiol. 2020, 6, 1827–1840. [Google Scholar] [CrossRef]
- Opthof, T. Embryological development of pacemaker hierarchy and membrane currents related to the function of the adult sinus node: Implications for autonomic modulation of biopacemakers. Med. Biol. Eng. Comput. 2007, 45, 119–132. [Google Scholar] [CrossRef]
- Opthof, T.; Van Ginneken, A.C.; Bouman, L.N.; Jongsma, H.J. The intrinsic cycle length in small pieces isolated from the rabbit sinoatrial node. J. Mol. Cell. Cardiol. 1987, 19, 923–934. [Google Scholar] [CrossRef]
- Clancy, C.E.; Santana, L.F. Evolving Discovery of the Origin of the Heartbeat: A New Perspective on Sinus Rhythm. JACC Clin. Electrophysiol. 2020, 6, 932–934. [Google Scholar] [CrossRef]
- Boyett, M.R.; Honjo, H.; Kodama, I. The sinoatrial node, a heterogeneous pacemaker structure. Cardiovasc. Res. 2000, 47, 658–687. [Google Scholar] [CrossRef]
- Feldman, J.L.; Kam, K. Facing the challenge of mammalian neural microcircuits: Taking a few breaths may help. J. Physiol. 2015, 593, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Holden, A.V.; Kodama, I.; Honjo, H.; Lei, M.; Varghese, T.; Boyett, M.R. Mathematical models of action potentials in the periphery and center of the rabbit sinoatrial node. Am. J. Physiol. 2000, 279, H397–H421. [Google Scholar] [CrossRef]
- Cloherty, S.L.; Dokos, S.; Lovell, N.H. A comparison of 1-D models of cardiac pacemaker heterogeneity. IEEE Trans. Biomed. Eng. 2006, 53, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Oren, R.V.; Clancy, C.E. Determinants of heterogeneity, excitation and conduction in the sinoatrial node: A model study. PLoS Comput. Biol. 2010, 6, e1001041. [Google Scholar] [CrossRef]
- Huang, X.; Mi, Y.; Qian, Y.; Hu, G. Phase-locking behaviors in an ionic model of sinoatrial node cell and tissue. Phys. Rev. E Stat. Nonlin. Soft Matter Phys. 2011, 83, 61917. [Google Scholar] [CrossRef] [PubMed]
- Glynn, P.; Onal, B.; Hund, T.J. Cycle length restitution in sinoatrial node cells: A theory for understanding spontaneous action potential dynamics. PLoS ONE 2014, 9, e89049. [Google Scholar] [CrossRef]
- Li, K.; Chu, Z.; Huang, X. Annihilation of the pacemaking activity in the sinoatrial node cell and tissue. AIP Adv. 2018, 8, 125319. [Google Scholar] [CrossRef]
- Gratz, D.; Onal, B.; Dalic, A.; Hund, T.J. Synchronization of pacemaking in the sinoatrial node: A mathematical modeling study. Front. Phys. 2018, 6, 63. [Google Scholar] [CrossRef]
- Mata, A.N.; Alonso, G.R.; Garza, G.L.; Fernández, J.R.G.; García, M.A.C.; Ábrego, N.P.C. Parallel simulation of the synchronization of heterogeneous cells in the sinoatrial node. Concurr. Comput. Pract. Exp. 2019, e5317. [Google Scholar] [CrossRef]
- Campana, C.A. 2-Dimensional Computational Model to Analyze the Effects of Cellular Heterogeinity on Cardiac Pacemaking. Ph.D. Thesis, Universita di Bologna, Corso di Studio in Ingegneria Biomedical, Bologna, Italy, 2015. [Google Scholar]
- Munoz, M.A.; Kaur, J.; Vigmond, E.J. Onset of atrial arrhythmias elicited by autonomic modulation of rabbit sinoatrial node activity: A modeling study. Am. J. Physiol. 2011, 301, H1974–H1983. [Google Scholar] [CrossRef]
- Monfredi, O.; Lyashkov, A.E.; Johnsen, A.B.; Inada, S.; Schneider, H.; Wang, R.; Nirmalan, M.; Wisloff, U.; Maltsev, V.A.; Lakatta, E.G.; et al. Biophysical characterization of the underappreciated and important relationship between heart rate variability and heart rate. Hypertension 2014, 64, 1334–1343. [Google Scholar] [CrossRef]
- Kori, H.; Kawamura, Y.; Masuda, N. Structure of cell networks critically determines oscillation regularity. J. Theor. Biol. 2012, 297, 61–72. [Google Scholar] [CrossRef][Green Version]
- Zhang, Y.; Ocampo-Espindola, J.L.; Kiss, I.Z.; Motter, A.E. Random heterogeneity outperforms design in network synchronization. Proc. Natl. Acad. Sci. USA 2021, 118, 2024299118. [Google Scholar] [CrossRef]
- Yaniv, Y.; Juhaszova, M.; Lyashkov, A.E.; Spurgeon, H.A.; Sollott, S.J.; Lakatta, E.G. Ca2+-regulated-cAMP/PKA signaling in cardiac pacemaker cells links ATP supply to demand. J. Mol. Cell. Cardiol. 2011, 51, 740–748. [Google Scholar] [CrossRef]
- Opthof, T.; de Jonge, B.; Masson-Pevet, M.; Jongsma, H.J.; Bouman, L.N. Functional and morphological organization of the cat sinoatrial node. J. Mol. Cell. Cardiol. 1986, 18, 1015–1031. [Google Scholar] [CrossRef]
- Pauza, D.H.; Rysevaite, K.; Inokaitis, H.; Jokubauskas, M.; Pauza, A.G.; Brack, K.E.; Pauziene, N. Innervation of sinoatrial nodal cardiomyocytes in mouse. A combined approach using immunofluorescent and electron microscopy. J. Mol. Cell. Cardiol. 2014, 75, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Inokaitis, H.; Pauziene, N.; Rysevaite-Kyguoliene, K.; Pauza, D.H. Innervation of sinoatrial nodal cells in the rabbit. Ann. Anat. 2016, 205, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.H. The disposition, morphology and innervation of cardiac specialized tissue in the guinea-pig. J. Anat. 1972, 111, 453–468. [Google Scholar] [PubMed]
- Armour, J.A. Potential clinical relevance of the ‘little brain’ on the mammalian heart. Exp. Physiol. 2008, 93, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Mitrofanova, L.B.; Gorshkov, A.N.; Konovalov, P.V.; Krylova, J.S. Telocytes in the human sinoatrial node. J. Cell. Mol. Med. 2018, 22, 521–532. [Google Scholar] [CrossRef]
- Kondo, A.; Kaestner, K.H. Emerging diverse roles of telocytes. Development 2019, 146. [Google Scholar] [CrossRef]
- Camelliti, P.; Green, C.R.; LeGrice, I.; Kohl, P. Fibroblast network in rabbit sinoatrial node: Structural and functional identification of homogeneous and heterogeneous cell coupling. Circ. Res. 2004, 94, 828–835. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Before Stimulation | βAR Stimulation |
|---|---|---|
| Mean cycle length, ms | 921.8 1 | 507.3 1 |
| SEM, ms | 66.5 | 13.1 |
| SD, ms | 856.4 | 169.1 |
| CV% | 92.9 | 33.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.S.; Monfredi, O.; Maltseva, L.A.; Lakatta, E.G.; Maltsev, V.A. β-Adrenergic Stimulation Synchronizes a Broad Spectrum of Action Potential Firing Rates of Cardiac Pacemaker Cells toward a Higher Population Average. Cells 2021, 10, 2124. https://doi.org/10.3390/cells10082124
Kim MS, Monfredi O, Maltseva LA, Lakatta EG, Maltsev VA. β-Adrenergic Stimulation Synchronizes a Broad Spectrum of Action Potential Firing Rates of Cardiac Pacemaker Cells toward a Higher Population Average. Cells. 2021; 10(8):2124. https://doi.org/10.3390/cells10082124
Chicago/Turabian StyleKim, Mary S., Oliver Monfredi, Larissa A. Maltseva, Edward G. Lakatta, and Victor A. Maltsev. 2021. "β-Adrenergic Stimulation Synchronizes a Broad Spectrum of Action Potential Firing Rates of Cardiac Pacemaker Cells toward a Higher Population Average" Cells 10, no. 8: 2124. https://doi.org/10.3390/cells10082124
APA StyleKim, M. S., Monfredi, O., Maltseva, L. A., Lakatta, E. G., & Maltsev, V. A. (2021). β-Adrenergic Stimulation Synchronizes a Broad Spectrum of Action Potential Firing Rates of Cardiac Pacemaker Cells toward a Higher Population Average. Cells, 10(8), 2124. https://doi.org/10.3390/cells10082124