
Effect of Low Temperature on Changes in AGP Distribution during Development of Bellis perennis Ovules and Anthers


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material
2.2. Preparation of Material for Microscopic Studies
2.3. Immunocytochemical Reaction Procedure
2.4. Primary Antibodies
2.5. Imaging
3. Results
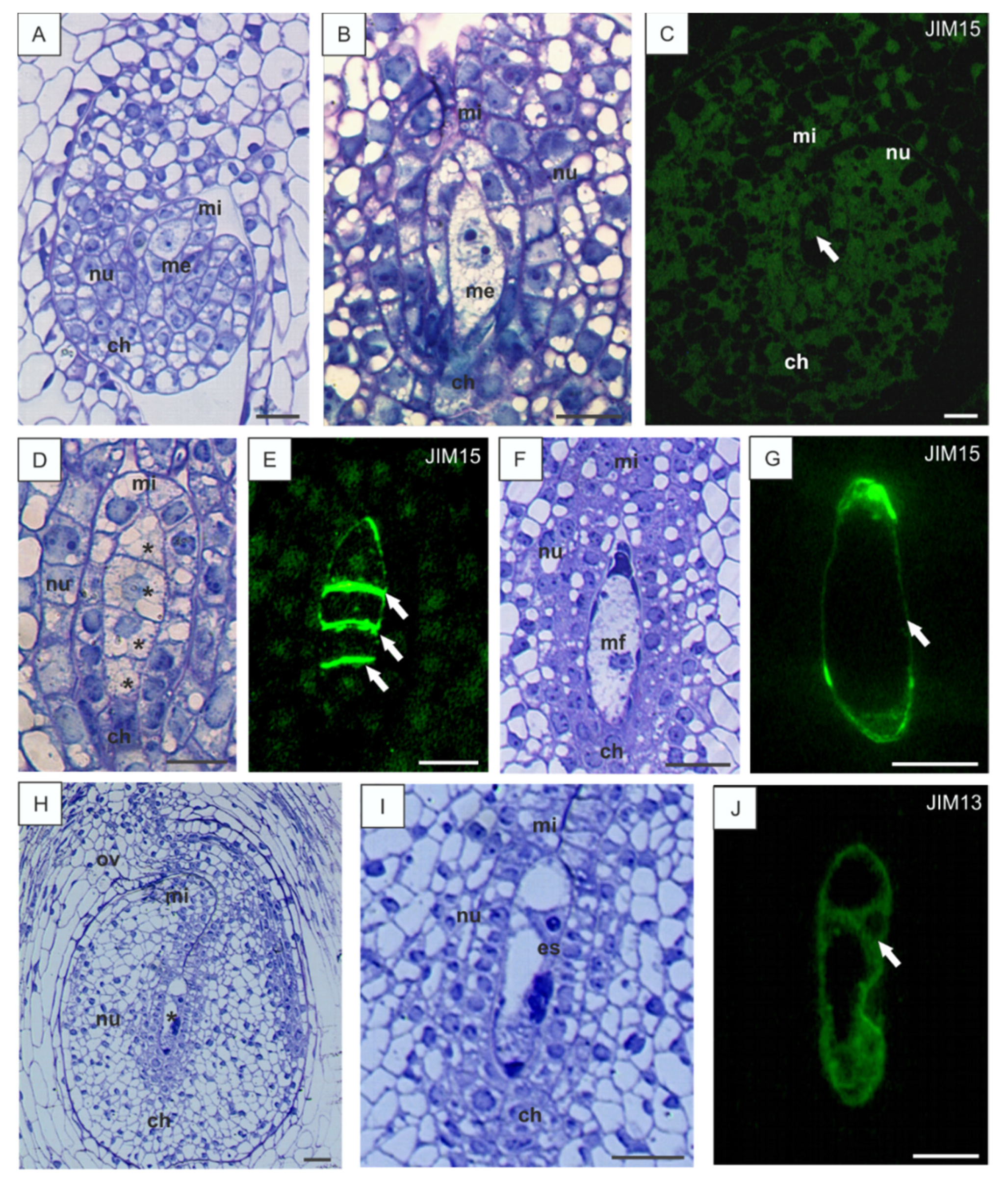
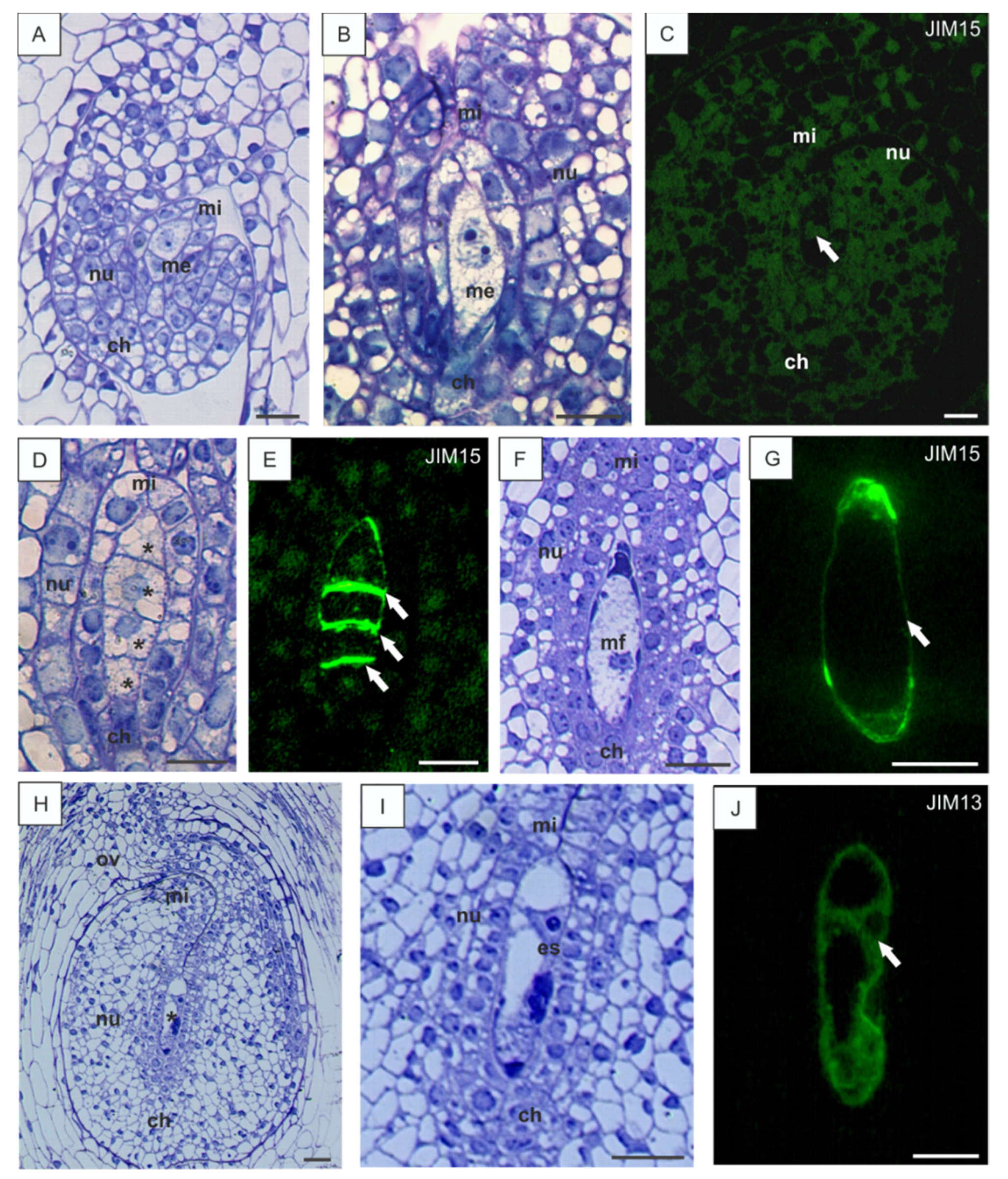
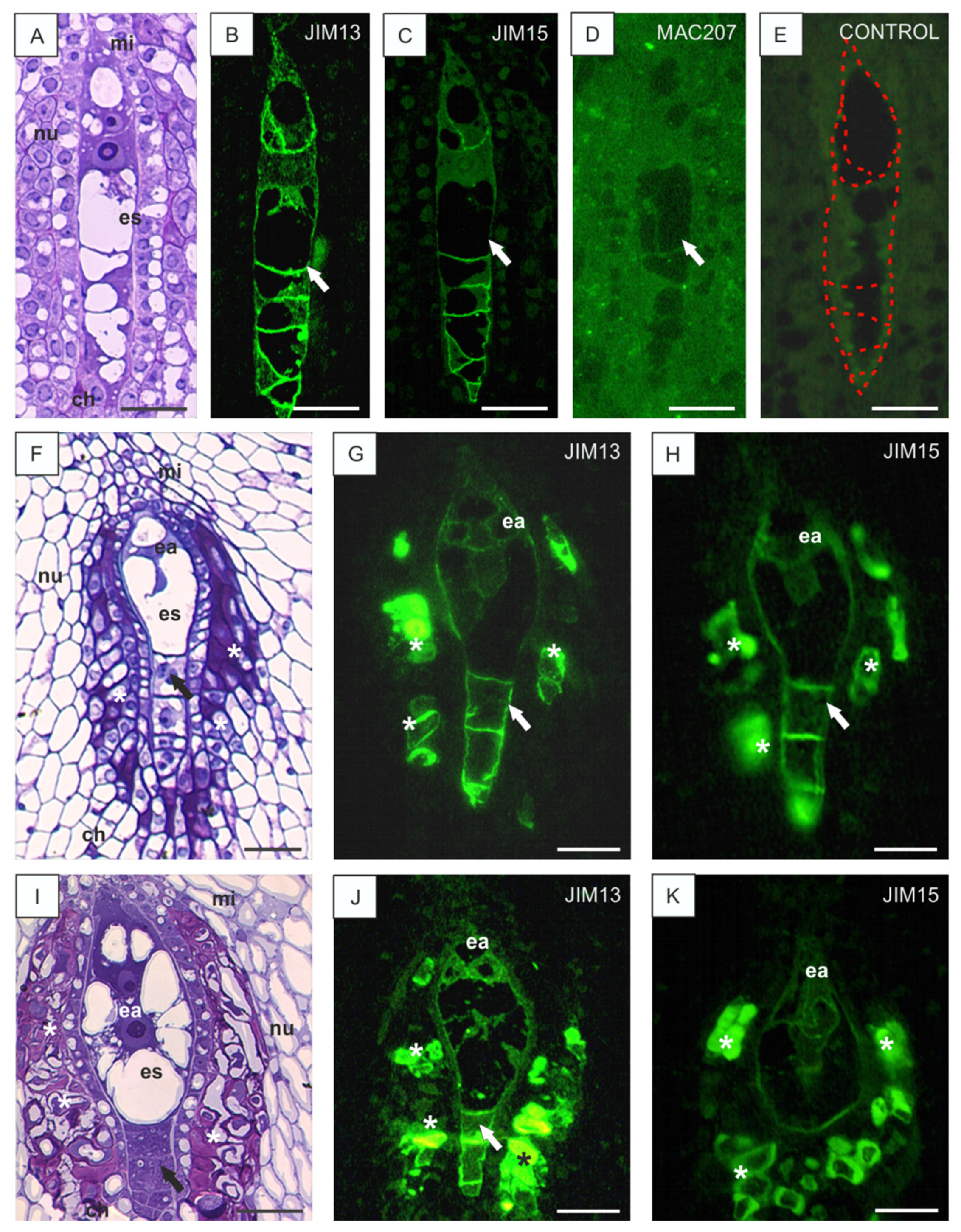
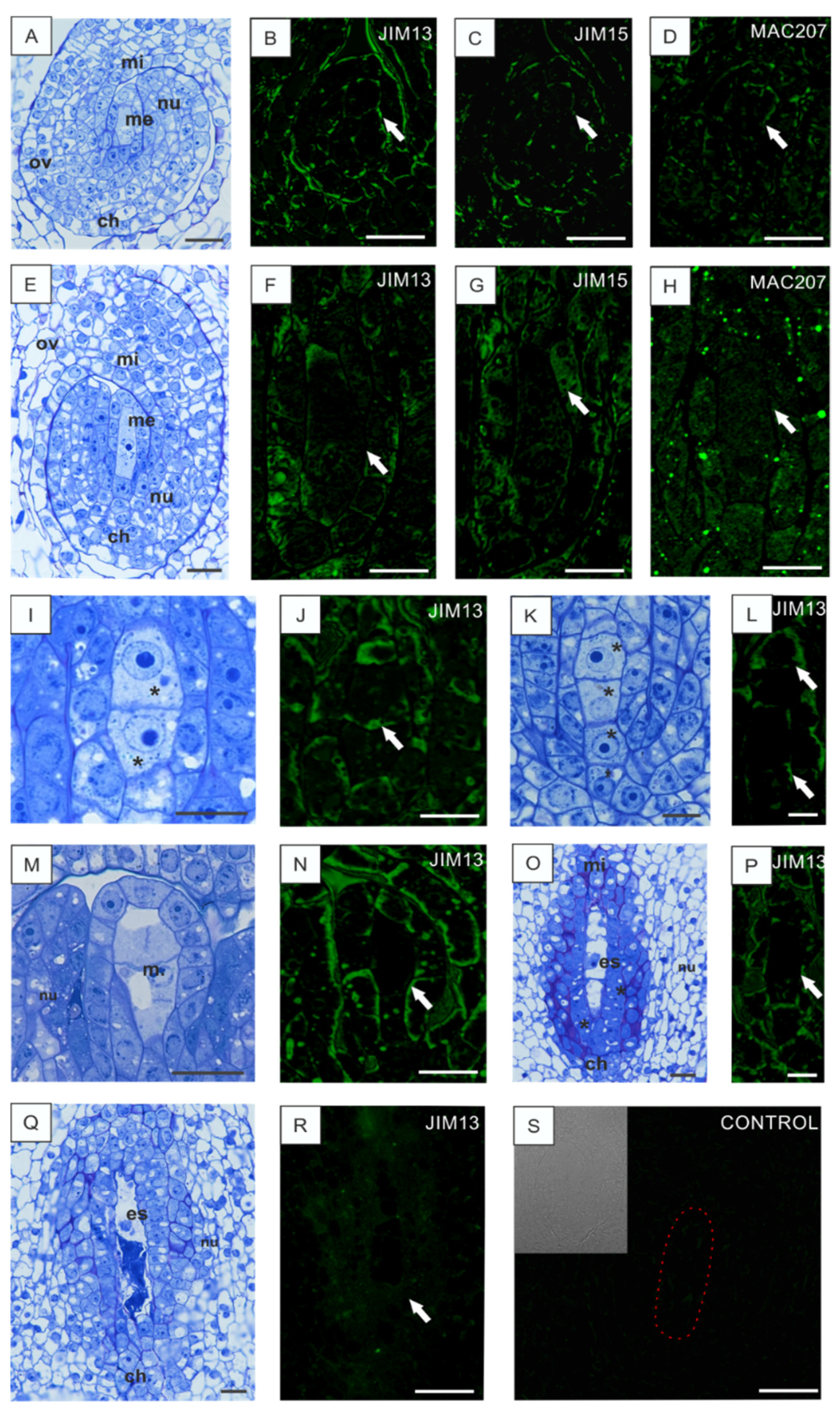
3.1. Localization of AGPs Epitopes in the Ovules of B. perennis during Megasporogenesis and Megagametogenesis
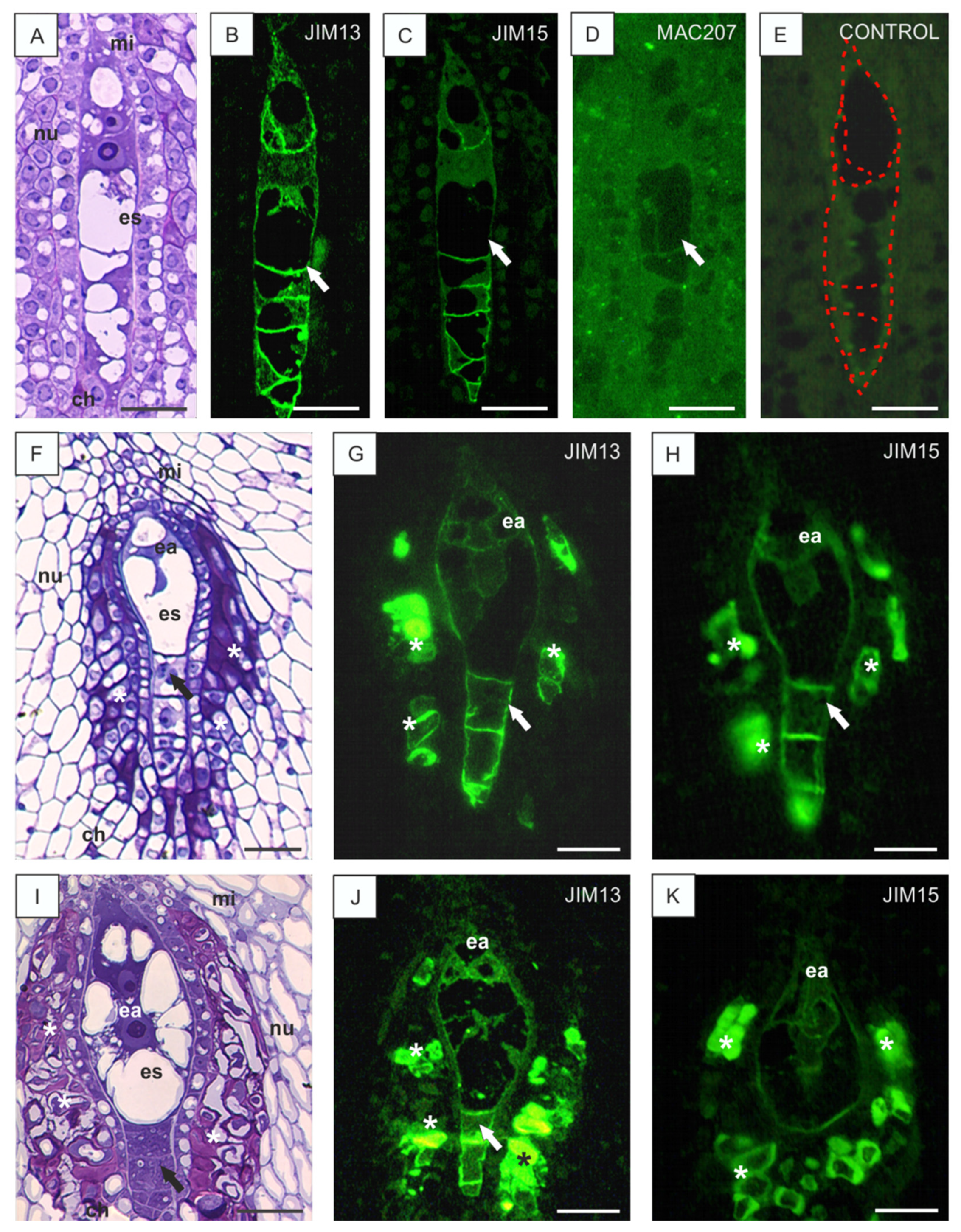
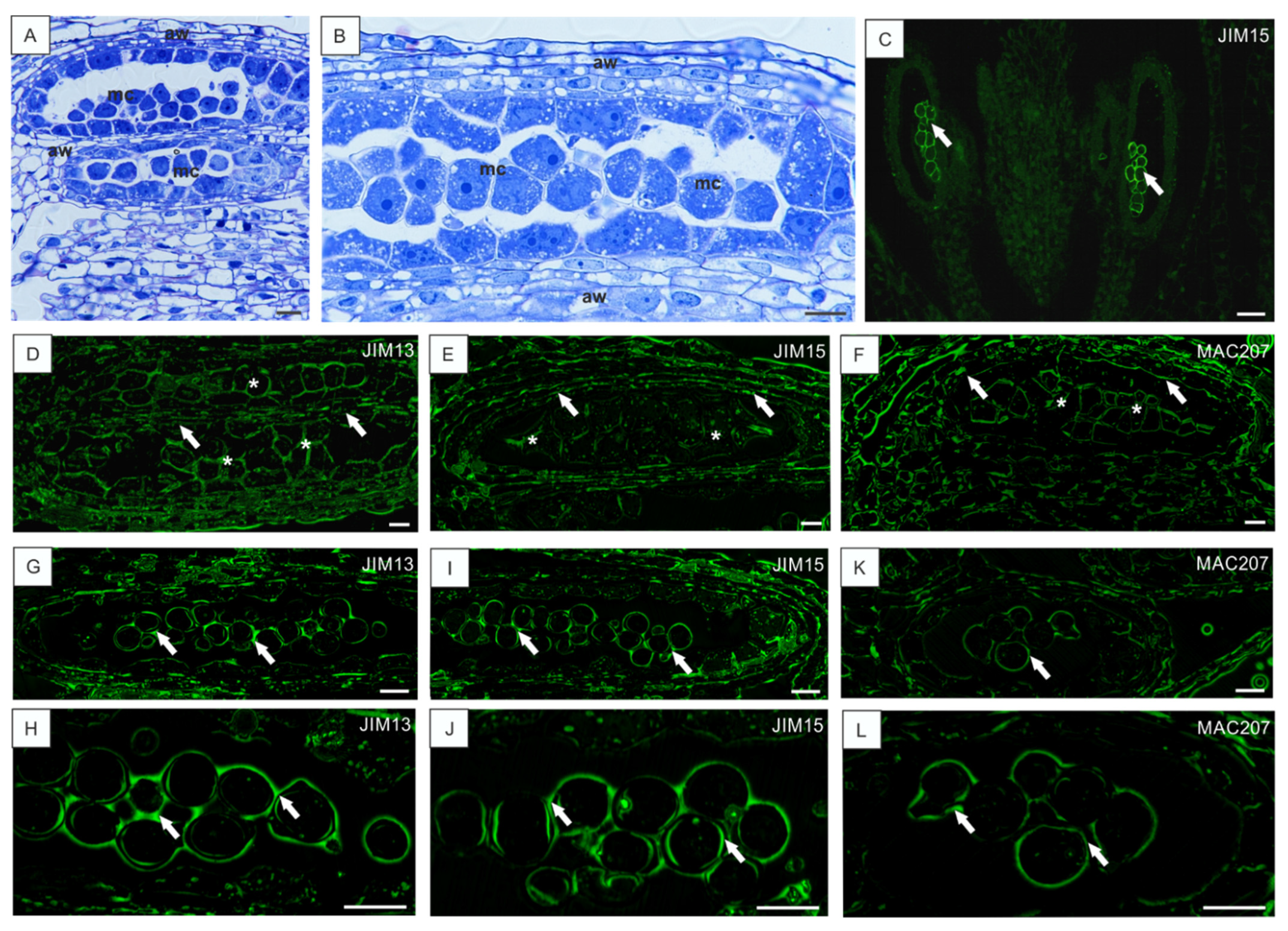
3.2. Disturbances in AGP Localization in Developing Ovules Caused by Low Temperature
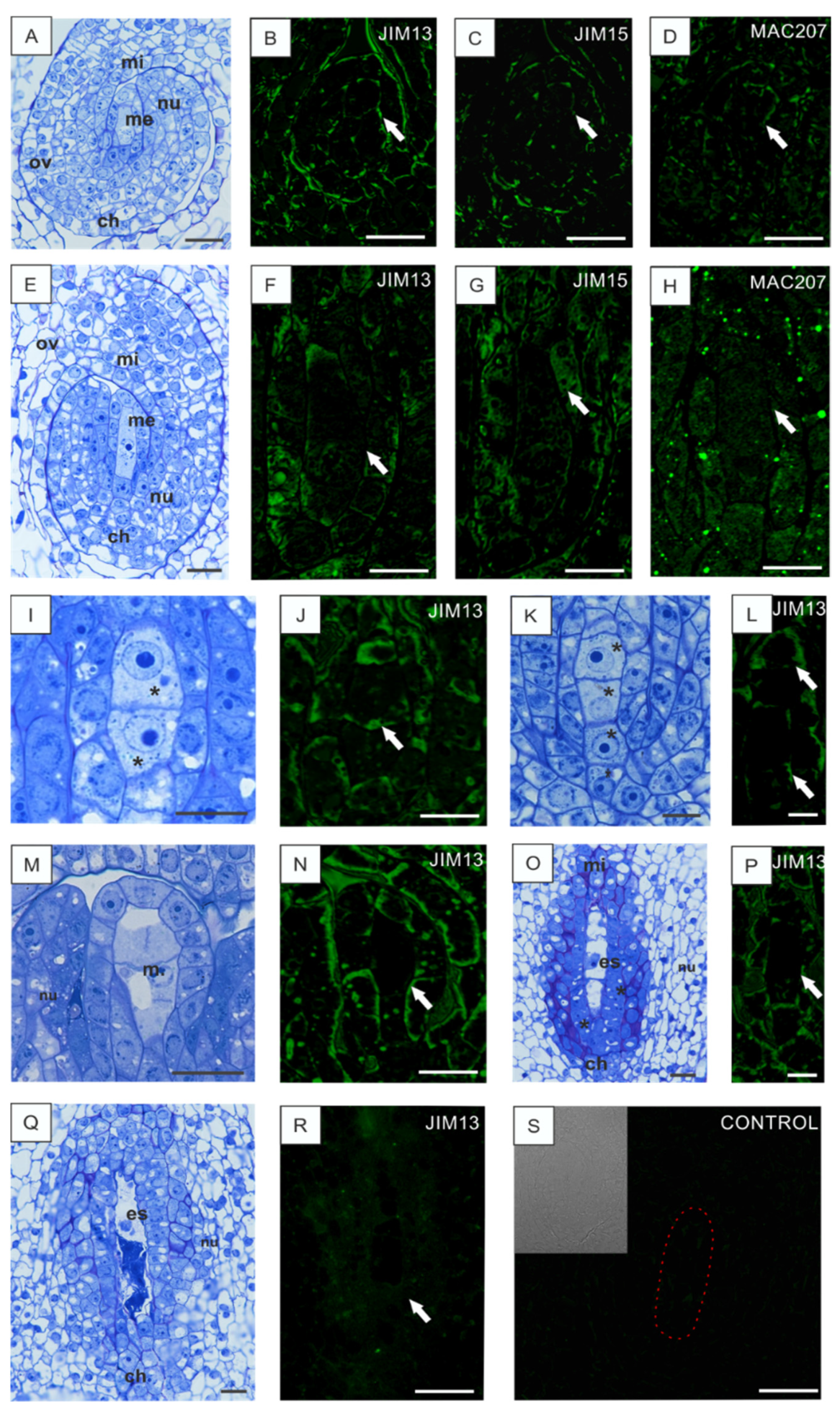
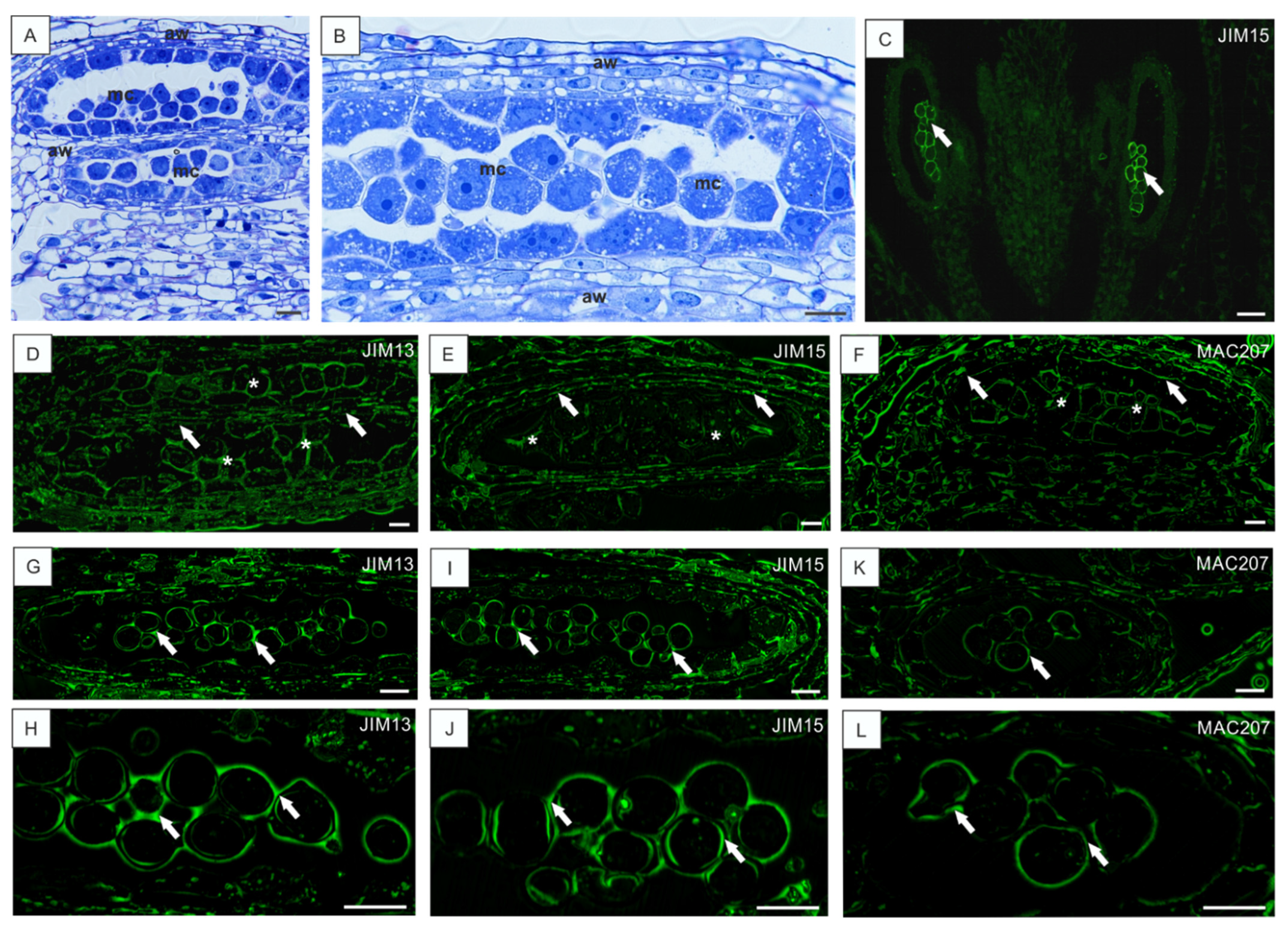
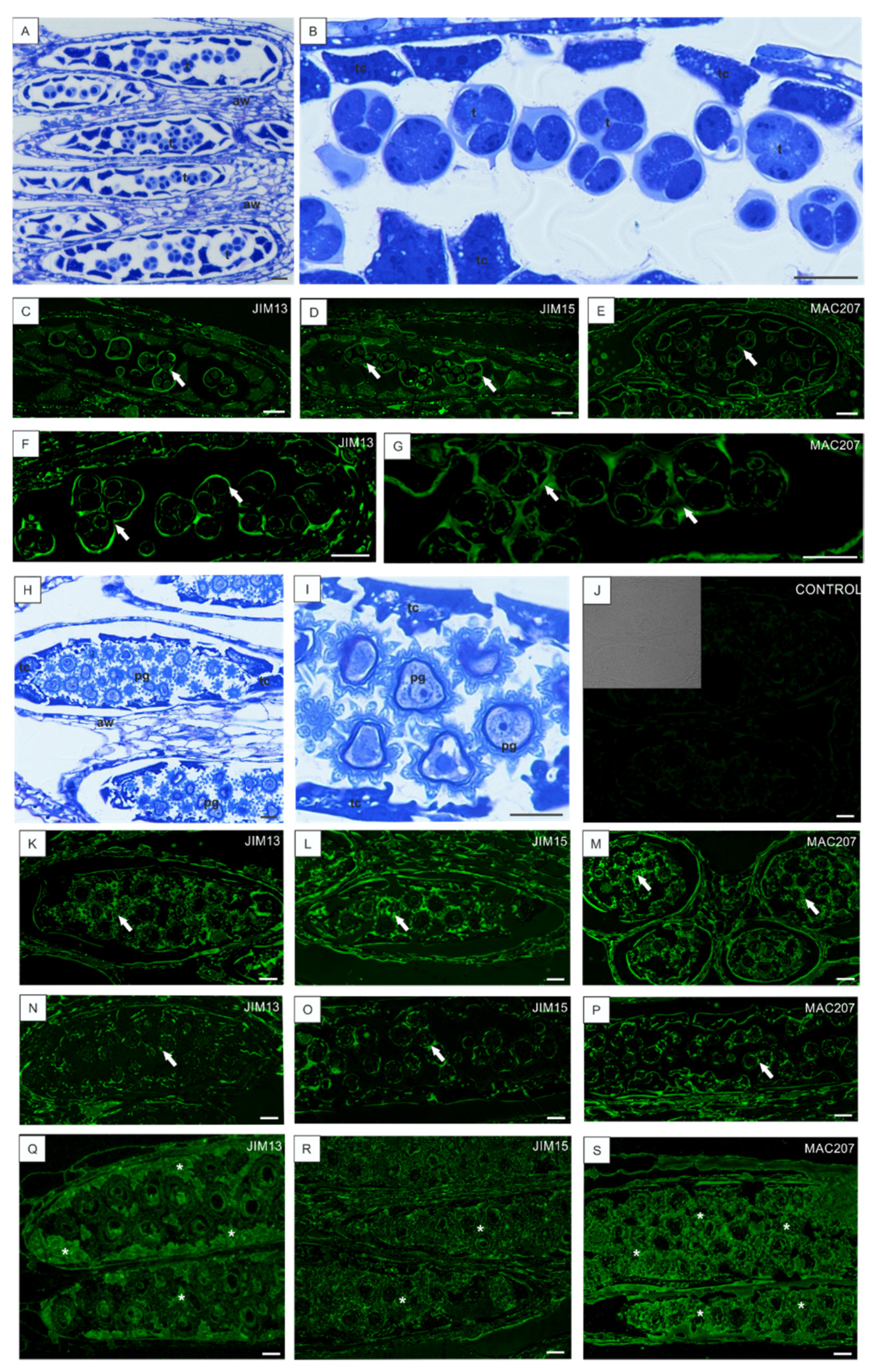
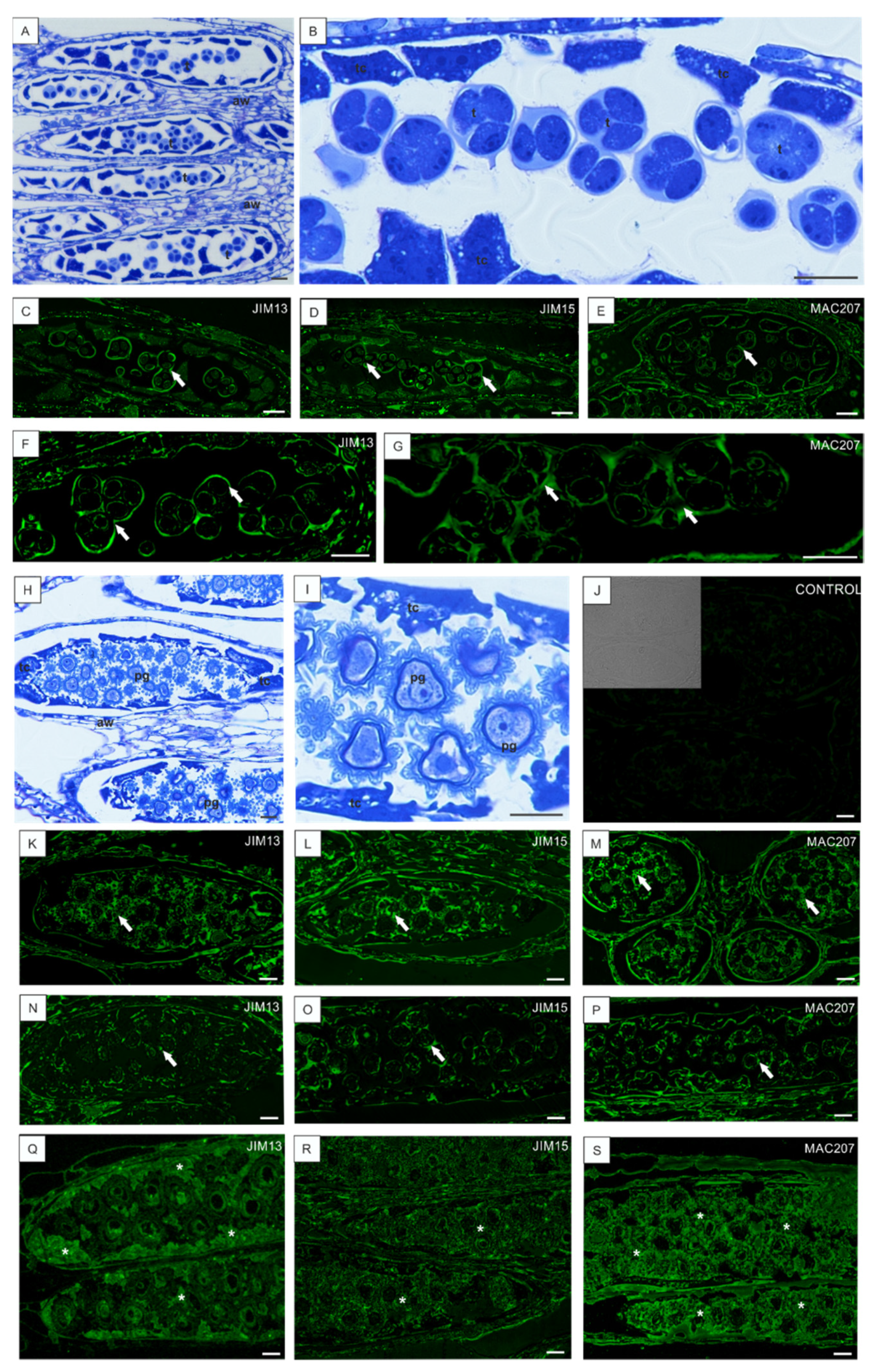
3.3. Low Temperature-Induced Disturbances in AGP Localization during Development of Anthers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ma, Y.; Yan, C.; Li, H.; Wu, W.; Liu, W.; Wang, Y.; Chen, Q.; Ma, H. Bioinformatic prediction and evolution analysis of arabinogalactan proteins in the plant kingdom. Front. Plant Sci. 2017, 8, 66. [Google Scholar] [CrossRef] [Green Version]
- Showalter, A.M. Arabinogalactan-proteins: Structure, expression and function. Cell. Mol. Life Sci. 2001, 58, 1399–1417. [Google Scholar] [CrossRef]
- Silva, J.; Ferraz, R.; Dupree, P.; Showalter, A.M.; Coimbra, S. Three decades of advances in arabinogalactan-protein biosynthesis. Front. Plant Sci. 2020, 11, 610377. [Google Scholar] [CrossRef]
- Showalter, A.M.; Basu, D. Extensin and arabinogalactan-protein biosynthesis: Glycosyltransferases, research challenges, and biosensors. Front. Plant Sci. 2016, 7, 814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.; Eberhard, S.; Pattathil, S.; Warder, C.; Glushka, J.; Yuan, C.; Hao, Z.; Zhu, X.; Avci, U.; Miller, J.S.; et al. An Arabidopsis cell wall proteoglycan consists of pectin and arabinoxylan covalently linked to an arabinogalactan protein. Plant Cell 2013, 25, 270–287. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Persson, S.; Sánchez-Rodríguez, C. At the border: The plasma membrane-cell wall continuum. J. Exp. Bot. 2015, 66, 1553–1563. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.M.; Lopes, A.L.; Coimbra, S. Arabinogalactan proteins as interactors along the crosstalk between the pollen tube and the female tissues. Front. Plant Sci. 2016, 7, 1895. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.M.; Lopes, A.L.; Coimbra, S. JAGGER, an AGP essential for persistent synergid degeneration and polytubey block in Arabidopsis. Plant Signal. Behav. 2016, 11, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, A.M.; Nobre, M.S.; Pinto, S.C.; Lopes, A.L.; Costa, M.L.; Masiero, S.; Coimbra, S. ‘Love is strong, and you’re so sweet’: JAGGER is essential for persistent synergid degeneration and polytubey block in Arabidopsis thaliana. Mol. Plant 2016, 9, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Leszczuk, A.; Szczuka, E.; Zdunek, A. Arabinogalactan proteins: Distribution during the development of male and female gametophytes. Plant Physiol. Biochem. 2019, 135, 9–18. [Google Scholar] [CrossRef]
- Coimbra, S.; Almeida, J.; Junqueira, V.; Costa, M.L.; Pereira, L.G. Arabinogalactan proteins as molecular markers in Arabidopsis thaliana sexual reproduction. J. Exp. Bot. 2007, 58, 4027–4035. [Google Scholar] [CrossRef] [Green Version]
- Chudzik, B.; Szczuka, E.; Zarzyka, B.; Leszczuk, A. Distribution of arabinogalactan proteins during microsporogenesis in the anther of Bellis perennis L. (Asteraceae). Acta Biol. Crac. Ser. Bot. 2014, 56, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Leszczuk, A.; Szczuka, E. Arabinogalactan proteins: Immunolocalization in the developing ovary of a facultative apomict Fragaria x ananassa (Duch.). Plant Physiol. Biochem. 2018, 123, 24–33. [Google Scholar] [CrossRef]
- Qin, Y.; Zhao, J. Localization of arabinogalactan proteins in egg cells, zygotes, and two-celled proembryos and effects of β-D-glucosyl Yariv reagent on egg cell fertilization and zygote division in Nicotiana tabacum L. J. Exp. Bot. 2006, 57, 2061–2074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acosta-García, G.; Vielle-Calzada, J.P. A classical arabinogalactan protein is essential for the initiation of female gametogenesis in Arabidopsis. Plant Cell 2004, 16, 2614–2628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, T.; Dong, F.; Luan, D.; Hu, H.; Zhao, J. Gene expression and localization of arabinogalactan proteins during the development of anther, ovule, and embryo in rice. Protoplasma 2019, 256, 909–922. [Google Scholar] [CrossRef] [PubMed]
- Rafińska, K.; Bednarska, E. Localization pattern of homogalacturonan and arabinogalactan proteins in developing ovules of the gymnosperm plant Larix decidua Mill. Plant Reprod. 2011, 24, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Coimbra, S.; Salema, R. Immunolocalization of arabinogalactan proteins in Amaranthus hypochondriacus L. ovules. Protoplasma 1997, 199, 75–82. [Google Scholar] [CrossRef]
- Pereira, A.M.; Masiero, S.; Nobre, M.S.; Costa, M.L.; Solis, M.T.; Testillano, P.S.; Sprunck, S.; Coimbra, S. Differential expression patterns of arabinogalactan proteins in Arabidopsis thaliana reproductive tissues. J. Exp. Bot. 2014, 65, 5459–5471. [Google Scholar] [CrossRef] [Green Version]
- Leszczuk, A.; Kozioł, A.; Szczuka, E.; Zdunek, A. Analysis of AGP contribution to the dynamic assembly and mechanical properties of cell wall during pollen tube growth. Plant Sci. 2019, 281, 9–18. [Google Scholar] [CrossRef]
- Leszczuk, A.; Wydrych, J.; Szczuka, E. The occurrence of calcium oxalate crystals and distribution of arabinogalactan proteins (AGPs) in ovary cells during Fragaria x ananassa (Duch.) development. J. Plant Growth Regul. 2019, 38, 1028–1036. [Google Scholar] [CrossRef] [Green Version]
- Le Gall, H.; Philippe, F.; Domon, J.M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell wall metabolism in response to abiotic stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef]
- Thakur, P.; Kumar, S.; Malik, J.A.; Berger, J.D.; Nayyar, H. Cold stress effects on reproductive development in grain crops: An overview. Environ. Exp. Bot. 2010, 67, 429–443. [Google Scholar] [CrossRef]
- Zinn, K.E.; Tunc-Ozdemir, M.; Harper, J.F. Temperature stress and plant sexual reproduction: Uncovering the weakest links. J. Exp. Bot. 2010, 61, 1959–1968. [Google Scholar] [CrossRef] [Green Version]
- Wilson, S.M.; Bacic, A. Preparation of plant cells for transmission electron microscopy to optimize immunogold labeling of carbohydrate and protein epitopes. Nat. Protoc. 2012, 7, 1716–1727. [Google Scholar] [CrossRef] [PubMed]
- Knox, J.P.; Linstead, P.J.; Peart, J.; Cooper, C.; Roberts, K. Developmentally regulated epitopes of cell surface arabinogalactan proteins and their relation to root tissue pattern formation. Plant J. 1991, 1, 317–326. [Google Scholar] [CrossRef]
- Yates, E.A.; Valdor, J.; Haslam, S.M.; Morris, H.R.; Dell, A.; Mackie, W.; Knox, J.P. Characterization of carbohydrate structural features recognized by anti-arabinogalactan-protein monoclonal antibodies. Glycobiology 1996, 6, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Pattathil, S.; Avci, U.; Baldwin, D.; Swennes, A.G.; McGill, J.A.; Popper, Z.; Bootten, T.; Albert, A.; Davis, R.H.; Chennareddy, C.; et al. A comprehensive toolkit of plant cell wall glycan-directed monoclonal antibodies. Plant Physiol. 2010, 153, 514–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pennell, R.I.; Roberts, K. Sexual development in the pea is presaged by altered expression of arabinogalactan protein. Nature 1990, 344, 547–549. [Google Scholar] [CrossRef]
- Coimbra, S.; Duarte, C. Arabinogalactan proteins may facilitate the movement of pollen tubes from the stigma to the ovules in Actinidia deliciosa and Amaranthus hypochondriacus. Euphytica 2003, 133, 171–178. [Google Scholar] [CrossRef]
- Suárez, C.; Zienkiewicz, A.; Castro, A.J.; Zienkiewicz, K.; Majewska-Sawka, A.; Rodríguez-García, M.I. Cellular localization and levels of pectins and arabinogalactan proteins in olive (Olea europaea L.) pistil tissues during development: Implications for pollen-pistil interaction. Planta 2013, 237, 305–319. [Google Scholar] [CrossRef]
- Płachno, B.J.; Kapusta, M.; Świątek, P.; Banaś, K.; Miranda, V.E.O.; Bogucka-Kocka, A. Spatio-temporal distribution of cell wall components in the placentas, ovules and female gametophytes of Utricularia during pollination. Int. J. Mol. Sci. 2021, 22, 5622. [Google Scholar] [CrossRef]
- Demesa-Arevalo, E.; Vielle-Calzada, J.P. The classical arabinogalactan protein AGP18 mediates megaspore selection in Arabidopsis. Plant Cell 2013, 25, 1274–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Y.; Zhao, J. Localization of arabinogalactan-proteins in different stages of embryos and their role in cotyledon formation of Nicotiana tabacum L. Sex. Plant Reprod. 2007, 20, 213–224. [Google Scholar] [CrossRef]
- Płachno, B.J.; Kapusta, M.; Świątek, P.; Stolarczyk, P.; Kocki, J. Immunodetection of pectic epitopes, arabinogalactan proteins, and extensis in mucilage cells from the ovules of Pilosella officinarum Vaill. and Taraxacum officinale Agg. (Asteraceae). Int. J. Mol. Sci. 2020, 21, 9642. [Google Scholar] [CrossRef] [PubMed]
- Corral-Martínez, P.; García-Fortea, E.; Bernard, S.; Driouich, A.; Seguí-Simarro, J.M. Ultrastructural immunolocalization of arabinogalactan protein, pectin and hemicellulose epitopes through anther development in Brassica napus. Plant Cell Physiol. 2016, 57, 2161–2174. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.L.; Sobral, R.; Costa, M.M.R.; Amorin, M.I.; Coimbra, S. Evaluation of the presence of arabinogalactan proteins and pectins Quercus suber male gametogenesis. Ann. Bot. 2015, 115, 81–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Tantawy, A.A.; Solis, M.T.; Costa, M.L.; Coimbra, S.; Risueño, M.C.; Testillano, P.S. Arabinogalactan protein profiles and distribution patterns during microspore embryogenesis and pollen development in Brassica napus. Plant Reprod. 2013, 26, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Zhao, J. Genome-wide identification, classification, and expression analysis of the arabinogalactan protein gene family in rice (Oryza sativa L.). J. Exp. Bot. 2010, 61, 2647–2668. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Takác, T.; Li, X.; Chen, H.; Wang, Y.; Xu, E.; Xie, L.; Su, Z.; Šamaj, J.; Xu, C. Variable content and distribution of arabinogalactan proteins in banana (Musa spp.) under low temperature stress. Front. Plant Sci. 2015, 6, 353. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leszczuk, A.; Szczuka, E.; Lewtak, K.; Chudzik, B.; Zdunek, A. Effect of Low Temperature on Changes in AGP Distribution during Development of Bellis perennis Ovules and Anthers. Cells 2021, 10, 1880. https://doi.org/10.3390/cells10081880
Leszczuk A, Szczuka E, Lewtak K, Chudzik B, Zdunek A. Effect of Low Temperature on Changes in AGP Distribution during Development of Bellis perennis Ovules and Anthers. Cells. 2021; 10(8):1880. https://doi.org/10.3390/cells10081880
Chicago/Turabian StyleLeszczuk, Agata, Ewa Szczuka, Kinga Lewtak, Barbara Chudzik, and Artur Zdunek. 2021. "Effect of Low Temperature on Changes in AGP Distribution during Development of Bellis perennis Ovules and Anthers" Cells 10, no. 8: 1880. https://doi.org/10.3390/cells10081880
APA StyleLeszczuk, A., Szczuka, E., Lewtak, K., Chudzik, B., & Zdunek, A. (2021). Effect of Low Temperature on Changes in AGP Distribution during Development of Bellis perennis Ovules and Anthers. Cells, 10(8), 1880. https://doi.org/10.3390/cells10081880