
Extensive Phenotype of Human Inflammatory Monocyte-Derived Dendritic Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Isolation
2.2. Cell Preparation and Culture
2.3. Flow Cytometry
2.4. Statistical Analysis
3. Results
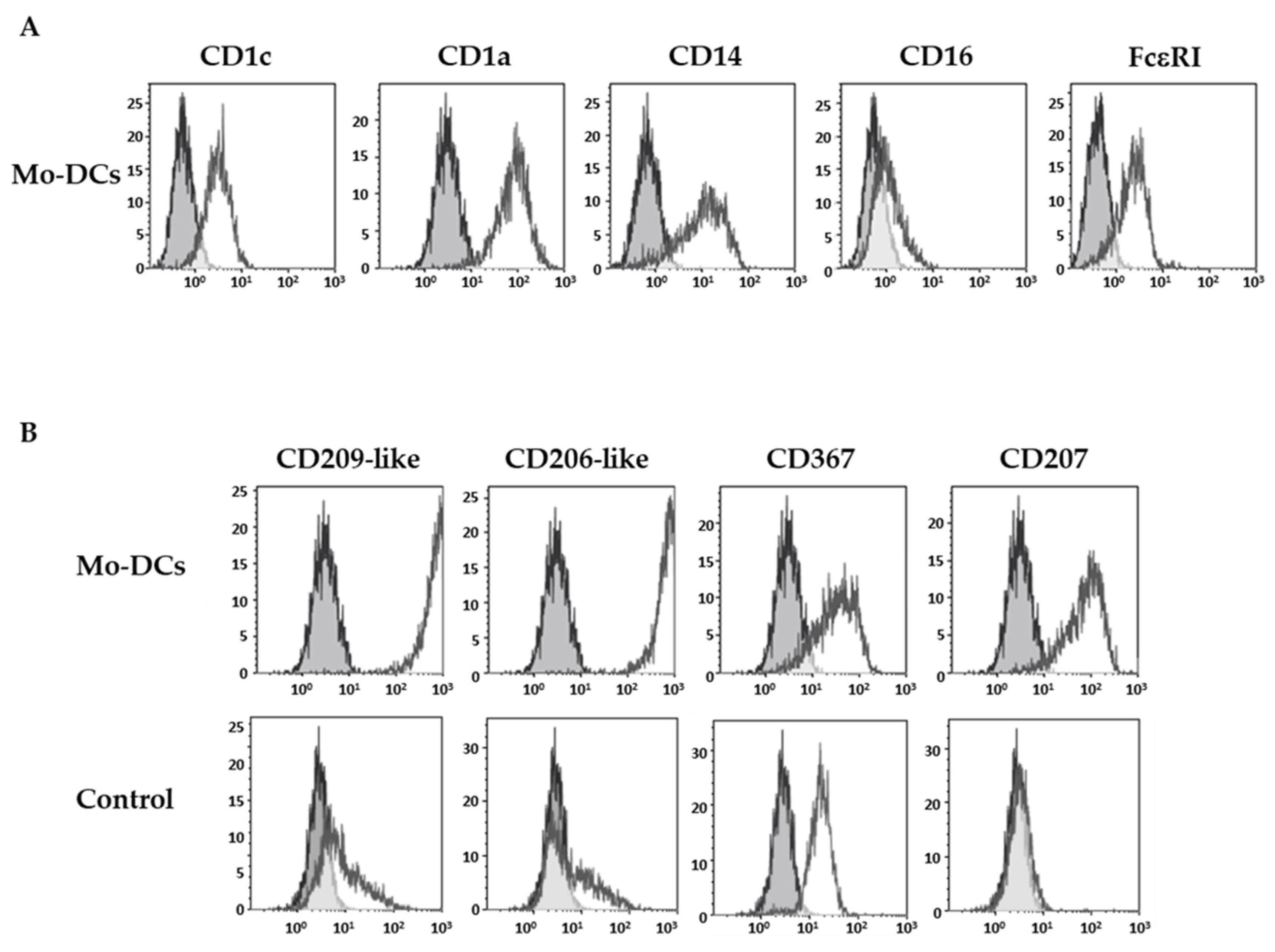
3.1. Inflammatory Mo-DCs Express a Large and Atypical Panel of C-Type Lectin Receptors
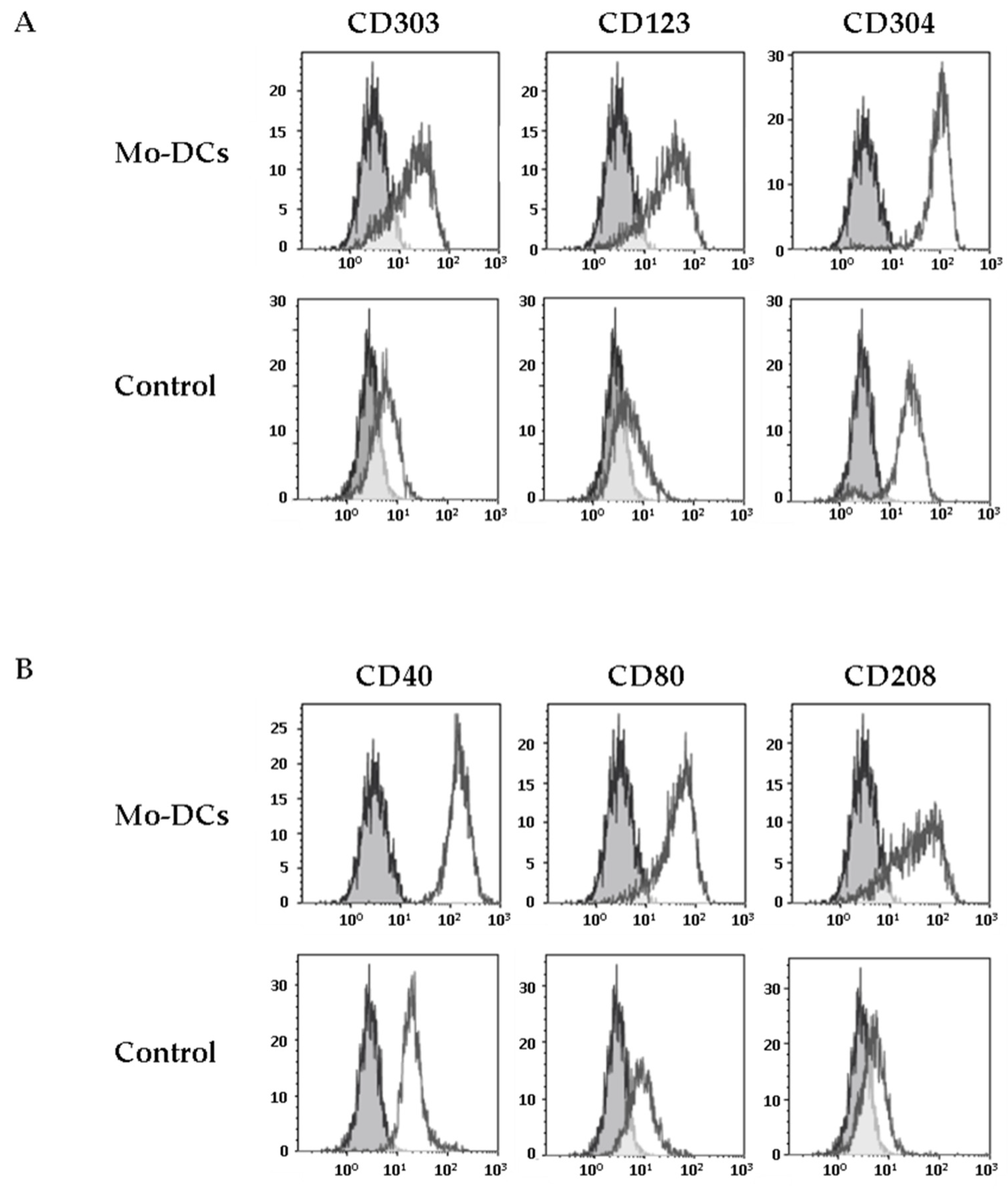
3.2. Inflammatory Mo-DCs Share Markers of Plasmacytoid Dendritic Cells
3.3. Inflammatory Mo-DCs Express the DC Maturation Marker CD208 at Their Surface
3.4. Ectopic Expression of Intracellular TLRs by Inflammatory DCs
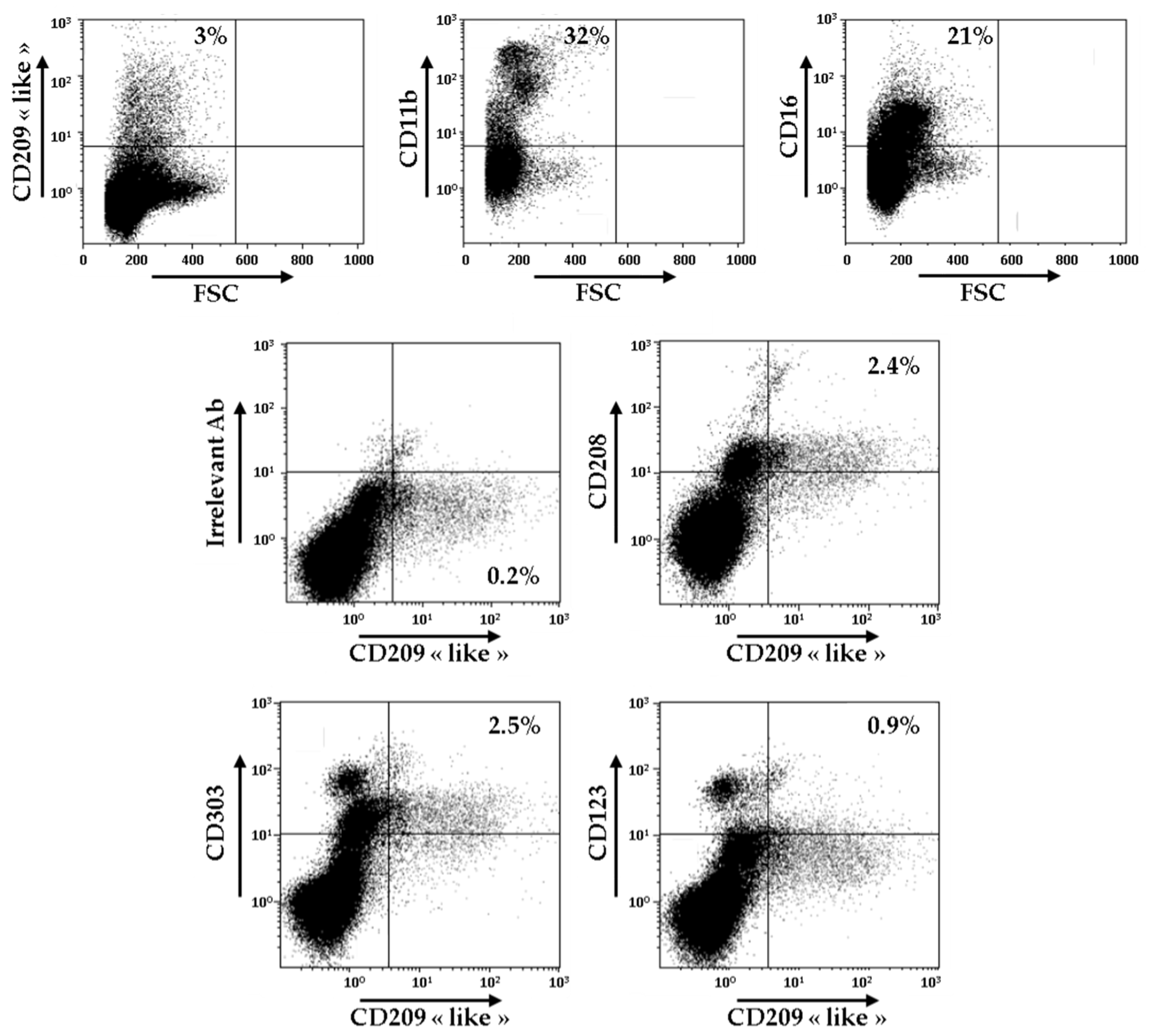
3.5. Identification of CD209-Like+ CD208+ Cells in the Synovial Fluid of Patients with RA
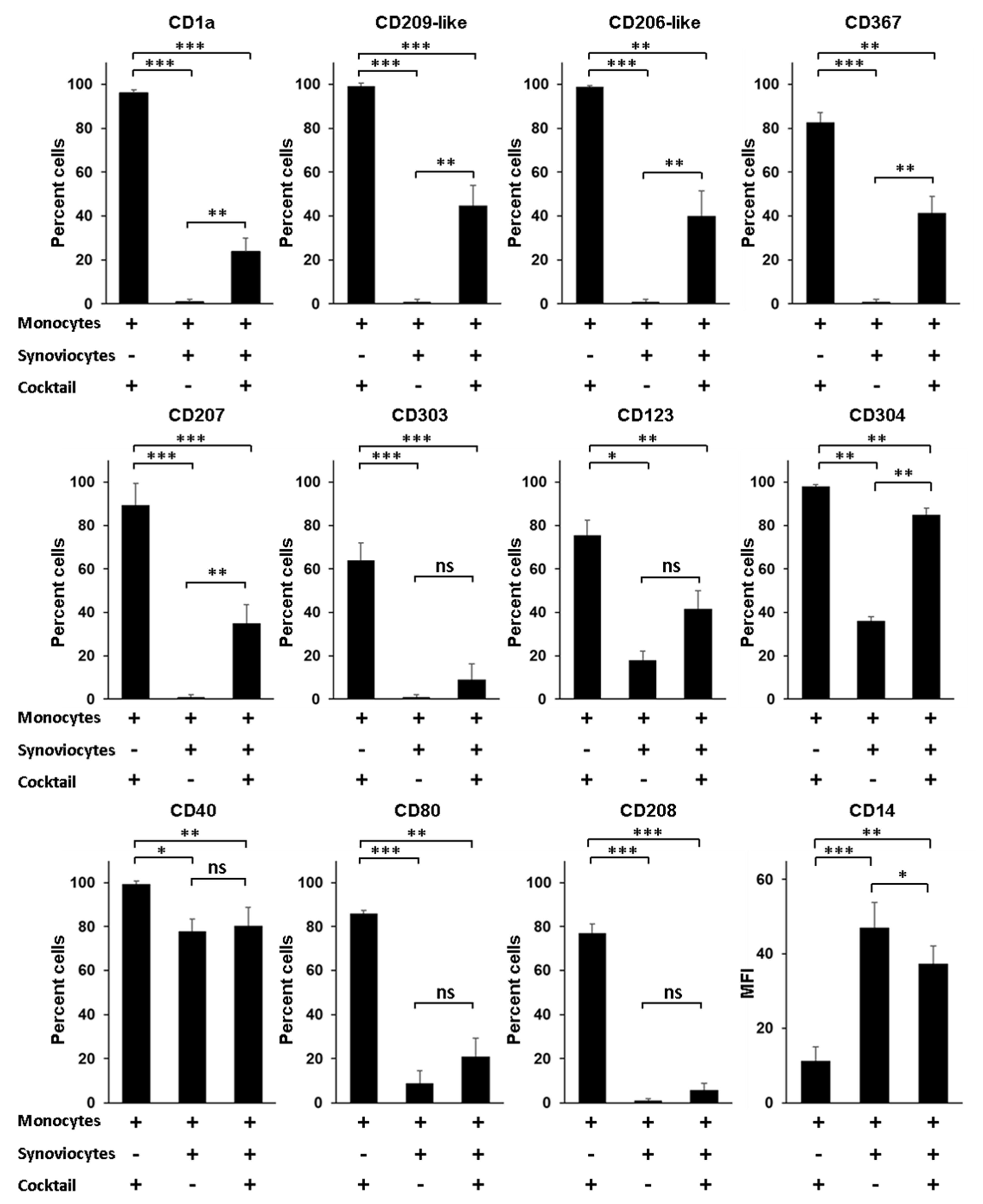
3.6. RA Synoviocytes Critically Affect the Differentiation of Monocytes into Mo-DCs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coutant, F.; Miossec, P. Altered Dendritic Cell Functions in Autoimmune Diseases: Distinct and Overlapping Profiles. Nat. Rev. Rheumatol. 2016, 12, 703–715. [Google Scholar] [CrossRef]
- Guilliams, M.; Ginhoux, F.; Jakubzick, C.; Naik, S.H.; Onai, N.; Schraml, B.U.; Segura, E.; Tussiwand, R.; Yona, S. Dendritic Cells, Monocytes and Macrophages: A Unified Nomenclature Based on Ontogeny. Nat. Rev. Immunol. 2014, 14, 571–578. [Google Scholar] [CrossRef]
- Macri, C.; Pang, E.S.; Patton, T.; O’Keeffe, M. Dendritic Cell Subsets. Semin. Cell Dev. Biol. 2018, 84, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Collin, M.; Bigley, V. Human Dendritic Cell Subsets: An Update. Immunology 2018, 154, 3–20. [Google Scholar] [CrossRef]
- Figdor, C.G.; van Kooyk, Y.; Adema, G.J. C-Type Lectin Receptors on Dendritic Cells and Langerhans Cells. Nat. Rev. Immunol. 2002, 2, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Del Fresno, C.; Iborra, S.; Saz-Leal, P.; Martínez-López, M.; Sancho, D. Flexible Signaling of Myeloid C-Type Lectin Receptors in Immunity and Inflammation. Front. Immunol. 2018, 9, 804. [Google Scholar] [CrossRef] [PubMed]
- Dzionek, A.; Sohma, Y.; Nagafune, J.; Cella, M.; Colonna, M.; Facchetti, F.; Günther, G.; Johnston, I.; Lanzavecchia, A.; Nagasaka, T.; et al. BDCA-2, a Novel Plasmacytoid Dendritic Cell-Specific Type II C-Type Lectin, Mediates Antigen Capture and Is a Potent Inhibitor of Interferon Alpha/Beta Induction. J. Exp. Med. 2001, 194, 1823–1834. [Google Scholar] [CrossRef]
- Valladeau, J.; Ravel, O.; Dezutter-Dambuyant, C.; Moore, K.; Kleijmeer, M.; Liu, Y.; Duvert-Frances, V.; Vincent, C.; Schmitt, D.; Davoust, J.; et al. Langerin, a Novel C-Type Lectin Specific to Langerhans Cells, Is an Endocytic Receptor That Induces the Formation of Birbeck Granules. Immunity 2000, 12, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Eguíluz-Gracia, I.; Bosco, A.; Dollner, R.; Melum, G.R.; Lexberg, M.H.; Jones, A.C.; Dheyauldeen, S.A.; Holt, P.G.; Bækkevold, E.S.; Jahnsen, F.L. Rapid Recruitment of CD14 (+) Monocytes in Experimentally Induced Allergic Rhinitis in Human Subjects. J. Allergy Clin. Immunol. 2016, 137, 1872–1881.e12. [Google Scholar] [CrossRef] [Green Version]
- Segura, E.; Touzot, M.; Bohineust, A.; Cappuccio, A.; Chiocchia, G.; Hosmalin, A.; Dalod, M.; Soumelis, V.; Amigorena, S. Human Inflammatory Dendritic Cells Induce Th17 Cell Differentiation. Immunity 2013, 38, 336–348. [Google Scholar] [CrossRef] [Green Version]
- Zaba, L.C.; Fuentes-Duculan, J.; Eungdamrong, N.J.; Abello, M.V.; Novitskaya, I.; Pierson, K.C.; Gonzalez, J.; Krueger, J.G.; Lowes, M.A. Psoriasis Is Characterized by Accumulation of Immunostimulatory and Th1/Th17 Cell-Polarizing Myeloid Dendritic Cells. J. Investig. Derm. 2009, 129, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Tamoutounour, S.; Guilliams, M.; Montanana Sanchis, F.; Liu, H.; Terhorst, D.; Malosse, C.; Pollet, E.; Ardouin, L.; Luche, H.; Sanchez, C.; et al. Origins and Functional Specialization of Macrophages and of Conventional and Monocyte-Derived Dendritic Cells in Mouse Skin. Immunity 2013, 39, 925–938. [Google Scholar] [CrossRef] [Green Version]
- Richter, L.; Landsverk, O.J.B.; Atlasy, N.; Bujko, A.; Yaqub, S.; Horneland, R.; Øyen, O.; Aandahl, E.M.; Lundin, K.E.A.; Stunnenberg, H.G.; et al. Transcriptional Profiling Reveals Monocyte-Related Macrophages Phenotypically Resembling DC in Human Intestine. Mucosal Immunol. 2018, 11, 1512–1523. [Google Scholar] [CrossRef] [Green Version]
- Gu, F.-F.; Wu, J.-J.; Liu, Y.-Y.; Hu, Y.; Liang, J.-Y.; Zhang, K.; Li, M.; Wang, Y.; Zhang, Y.-A.; Liu, L. Human Inflammatory Dendritic Cells in Malignant Pleural Effusions Induce Th1 Cell Differentiation. Cancer Immunol. Immunother. 2020, 69, 779–788. [Google Scholar] [CrossRef] [Green Version]
- León, B.; López-Bravo, M.; Ardavín, C. Monocyte-Derived Dendritic Cells Formed at the Infection Site Control the Induction of Protective T Helper 1 Responses against Leishmania. Immunity 2007, 26, 519–531. [Google Scholar] [CrossRef] [Green Version]
- Nakano, H.; Lin, K.L.; Yanagita, M.; Charbonneau, C.; Cook, D.N.; Kakiuchi, T.; Gunn, M.D. Blood-Derived Inflammatory Dendritic Cells in Lymph Nodes Stimulate Acute T Helper Type 1 Immune Responses. Nat. Immunol. 2009, 10, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Iijima, N.; Mattei, L.M.; Iwasaki, A. Recruited Inflammatory Monocytes Stimulate Antiviral Th1 Immunity in Infected Tissue. Proc. Natl. Acad. Sci. USA 2011, 108, 284–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammad, H.; Plantinga, M.; Deswarte, K.; Pouliot, P.; Willart, M.A.M.; Kool, M.; Muskens, F.; Lambrecht, B.N. Inflammatory Dendritic Cells—Not Basophils—Are Necessary and Sufficient for Induction of Th2 Immunity to Inhaled House Dust Mite Allergen. J. Exp. Med. 2010, 207, 2097–2111. [Google Scholar] [CrossRef] [PubMed]
- Plantinga, M.; Guilliams, M.; Vanheerswynghels, M.; Deswarte, K.; Branco-Madeira, F.; Toussaint, W.; Vanhoutte, L.; Neyt, K.; Killeen, N.; Malissen, B.; et al. Conventional and Monocyte-Derived CD11b (+) Dendritic Cells Initiate and Maintain T Helper 2 Cell-Mediated Immunity to House Dust Mite Allergen. Immunity 2013, 38, 322–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segura, E.; Amigorena, S. Inflammatory Dendritic Cells in Mice and Humans. Trends Immunol. 2013, 34, 440–445. [Google Scholar] [CrossRef] [Green Version]
- Goudot, C.; Coillard, A.; Villani, A.-C.; Gueguen, P.; Cros, A.; Sarkizova, S.; Tang-Huau, T.-L.; Bohec, M.; Baulande, S.; Hacohen, N.; et al. Aryl Hydrocarbon Receptor Controls Monocyte Differentiation into Dendritic Cells versus Macrophages. Immunity 2017, 47, 582–596.e6. [Google Scholar] [CrossRef] [Green Version]
- Zrioual, S.; Ecochard, R.; Tournadre, A.; Lenief, V.; Cazalis, M.-A.; Miossec, P. Genome-Wide Comparison between IL-17A- and IL-17F-Induced Effects in Human Rheumatoid Arthritis Synoviocytes. J. Immunol. 2009, 182, 3112–3120. [Google Scholar] [CrossRef] [PubMed]
- Clark, G.J.; Kupresanin, F.; Fromm, P.D.; Ju, X.; Muusers, L.; Silveira, P.A.; Elgundi, Z.; Gasiorowski, R.E.; Papadimitrious, M.S.; Bryant, C.; et al. New Insights into the Phenotype of Human Dendritic Cell Populations. Clin. Transl. Immunol. 2016, 5, e61. [Google Scholar] [CrossRef] [Green Version]
- Autenrieth, S.E.; Grimm, S.; Rittig, S.M.; Grünebach, F.; Gouttefangeas, C.; Bühring, H.-J. Profiling of Primary Peripheral Blood- and Monocyte-Derived Dendritic Cells Using Monoclonal Antibodies from the HLDA10 Workshop in Wollongong, Australia. Clin. Transl. Immunol. 2015, 4, e50. [Google Scholar] [CrossRef]
- Tordjman, R.; Lepelletier, Y.; Lemarchandel, V.; Cambot, M.; Gaulard, P.; Hermine, O.; Roméo, P.-H. A Neuronal Receptor, Neuropilin-1, Is Essential for the Initiation of the Primary Immune Response. Nat. Immunol. 2002, 3, 477–482. [Google Scholar] [CrossRef]
- de Saint-Vis, B.; Vincent, J.; Vandenabeele, S.; Vanbervliet, B.; Pin, J.J.; Aït-Yahia, S.; Patel, S.; Mattei, M.G.; Banchereau, J.; Zurawski, S.; et al. A Novel Lysosome-Associated Membrane Glycoprotein, DC-LAMP, Induced upon DC Maturation, Is Transiently Expressed in MHC Class II Compartment. Immunity 1998, 9, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Salaun, B.; de Saint-Vis, B.; Pacheco, N.; Pacheco, Y.; Riesler, A.; Isaac, S.; Leroux, C.; Clair-Moninot, V.; Pin, J.-J.; Griffith, J.; et al. CD208/Dendritic Cell-Lysosomal Associated Membrane Protein Is a Marker of Normal and Transformed Type II Pneumocytes. Am. J. Pathol. 2004, 164, 861–871. [Google Scholar] [CrossRef] [Green Version]
- Geijtenbeek, T.B.; Krooshoop, D.J.; Bleijs, D.A.; van Vliet, S.J.; van Duijnhoven, G.C.; Grabovsky, V.; Alon, R.; Figdor, C.G.; van Kooyk, Y. DC-SIGN-ICAM-2 Interaction Mediates Dendritic Cell Trafficking. Nat. Immunol. 2000, 1, 353–357. [Google Scholar] [CrossRef] [Green Version]
- Geijtenbeek, T.B.; Torensma, R.; van Vliet, S.J.; van Duijnhoven, G.C.; Adema, G.J.; van Kooyk, Y.; Figdor, C.G. Identification of DC-SIGN, A Novel Dendritic Cell-Specific ICAM-3 Receptor That Supports Primary Immune Responses. Cell 2000, 100, 575–585. [Google Scholar] [CrossRef] [Green Version]
- Anthony, R.M.; Kobayashi, T.; Wermeling, F.; Ravetch, J.V. Intravenous Gammaglobulin Suppresses Inflammation through a Novel T(H)2 Pathway. Nature 2011, 475, 110–113. [Google Scholar] [CrossRef]
- Caparrós, E.; Munoz, P.; Sierra-Filardi, E.; Serrano-Gómez, D.; Puig-Kröger, A.; Rodríguez-Fernández, J.L.; Mellado, M.; Sancho, J.; Zubiaur, M.; Corbí, A.L. DC-SIGN Ligation on Dendritic Cells Results in ERK and PI3K Activation and Modulates Cytokine Production. Blood 2006, 107, 3950–3958. [Google Scholar] [CrossRef] [Green Version]
- Gringhuis, S.I.; Kaptein, T.M.; Wevers, B.A.; Mesman, A.W.; Geijtenbeek, T.B.H. Fucose-Specific DC-SIGN Signalling Directs T Helper Cell Type-2 Responses via IKKε- and CYLD-Dependent Bcl3 Activation. Nat. Commun. 2014, 5, 3898. [Google Scholar] [CrossRef] [Green Version]
- Ponichtera, H.E.; Shainheit, M.G.; Liu, B.C.; Raychowdhury, R.; Larkin, B.M.; Russo, J.M.; Salantes, D.B.; Lai, C.-Q.; Parnell, L.D.; Yun, T.J.; et al. CD209a Expression on Dendritic Cells Is Critical for the Development of Pathogenic Th17 Cell Responses in Murine Schistosomiasis. J. Immunol. 2014, 192, 4655–4665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, G.; Miossec, P. Paired Synovium and Lymph Nodes from Rheumatoid Arthritis Patients Differ in Dendritic Cell and Chemokine Expression. J. Pathol. 2004, 204, 28–38. [Google Scholar] [CrossRef]
- Coutant, F.; Miossec, P. Evolving Concepts of the Pathogenesis of Rheumatoid Arthritis with Focus on the Early and Late Stages. Curr. Opin. Rheumatol. 2020, 32, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Svensson, M.; Kaye, P.M. Stromal-Cell Regulation of Dendritic-Cell Differentiation and Function. Trends Immunol. 2006, 27, 580–587. [Google Scholar] [CrossRef]
- Schirmer, C.; Klein, C.; von Bergen, M.; Simon, J.C.; Saalbach, A. Human Fibroblasts Support the Expansion of IL-17-Producing T Cells via up-Regulation of IL-23 Production by Dendritic Cells. Blood 2010, 116, 1715–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chomarat, P.; Banchereau, J.; Davoust, J.; Palucka, A.K. IL-6 Switches the Differentiation of Monocytes from Dendritic Cells to Macrophages. Nat. Immunol. 2000, 1, 510–514. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Clone | MFI (SEM) 1 |
|---|---|---|
| C-type lectin receptors | ||
| CD209 isoform (DC-SIGN like) | 118A8 | 413 (59) |
| CD206 (Mannose receptor) | 122D2 | 406 (61) |
| CD367 (DCIR) | 111F8 | 17 (4) |
| CD207 (Langerin) | 808E10 | 13 (3) |
| CD303 | 104C12 | 7 (1) |
| CD304 (Neuropilin-1) | 211H6 | 41 (6) |
| Toll-like receptors | ||
| TLR2 | 1308F10 | 0 |
| TLR3 | 1213F10 | 0 |
| TLR7 | 66H3 | 8 (1) |
| TLR8 | 112H7 | 0 |
| Antigen-presenting molecules | ||
| CD1a | 214A9 | 10 (2) |
| MHC-II | 216H1 | 132 (23) |
| Costimulatory molecules/Maturation markers | ||
| CD40 | G28–5 | 139 (18) |
| CD80 | Mab104 | 16 (3) |
| CD208 (DC-LAMP) | 109G3 | 15 (4) |
| Interleukin receptor | ||
| CD123 | 107D2 | 5 (2) |
| ITIM-bearing receptor | ||
| FDFO3 | 36H2 | 28 (2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coutant, F.; Pin, J.-J.; Miossec, P. Extensive Phenotype of Human Inflammatory Monocyte-Derived Dendritic Cells. Cells 2021, 10, 1663. https://doi.org/10.3390/cells10071663
Coutant F, Pin J-J, Miossec P. Extensive Phenotype of Human Inflammatory Monocyte-Derived Dendritic Cells. Cells. 2021; 10(7):1663. https://doi.org/10.3390/cells10071663
Chicago/Turabian StyleCoutant, Frédéric, Jean-Jacques Pin, and Pierre Miossec. 2021. "Extensive Phenotype of Human Inflammatory Monocyte-Derived Dendritic Cells" Cells 10, no. 7: 1663. https://doi.org/10.3390/cells10071663
APA StyleCoutant, F., Pin, J.-J., & Miossec, P. (2021). Extensive Phenotype of Human Inflammatory Monocyte-Derived Dendritic Cells. Cells, 10(7), 1663. https://doi.org/10.3390/cells10071663