
Venlafaxine and L-Thyroxine Treatment Combination: Impact on Metabolic and Synaptic Plasticity Changes in an Animal Model of Coexisting Depression and Hypothyroidism


Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Chemicals and Drugs
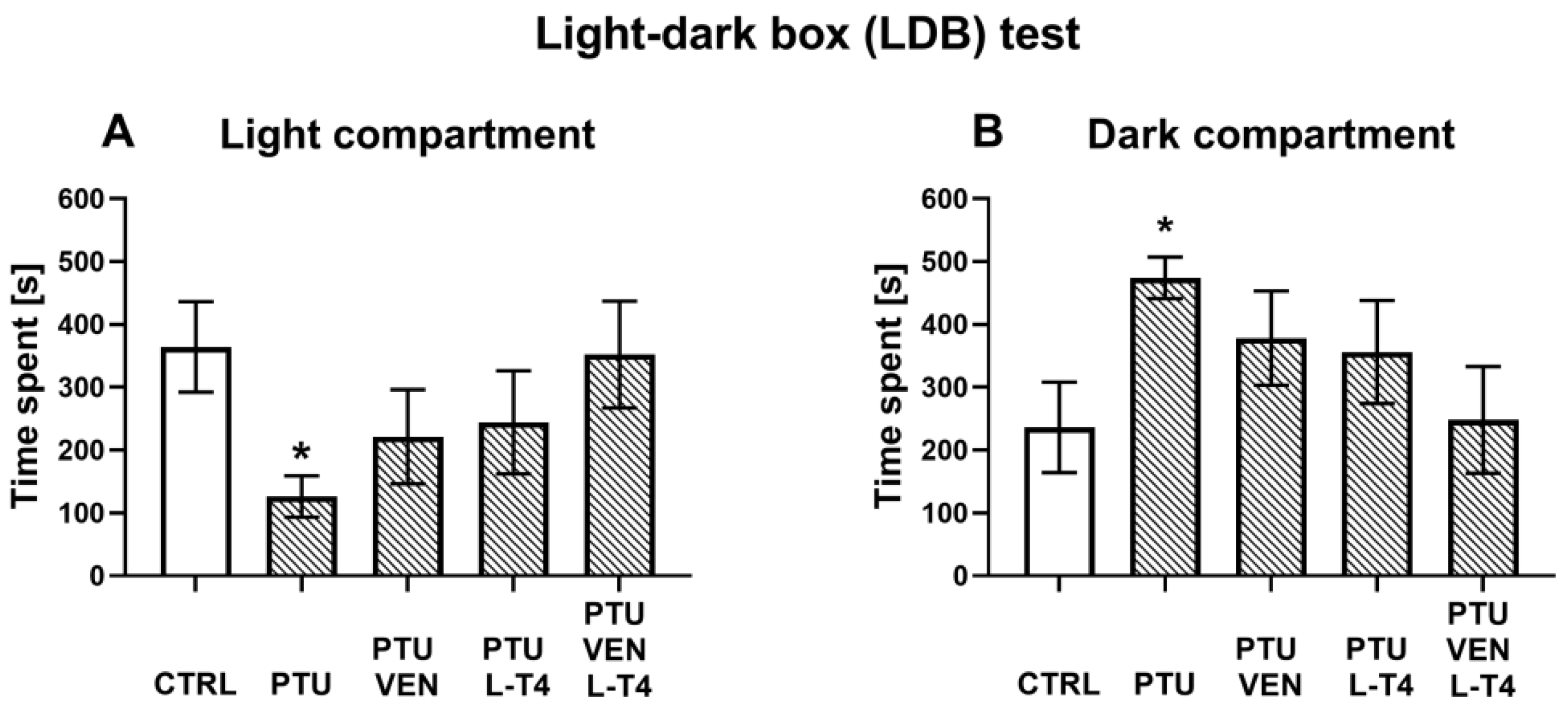
2.4. Behavioral Testing—Light-Dark Box (LDB) Test
2.5. Biochemical Analysis
2.5.1. Thyroid Hormone Measurement in Plasma
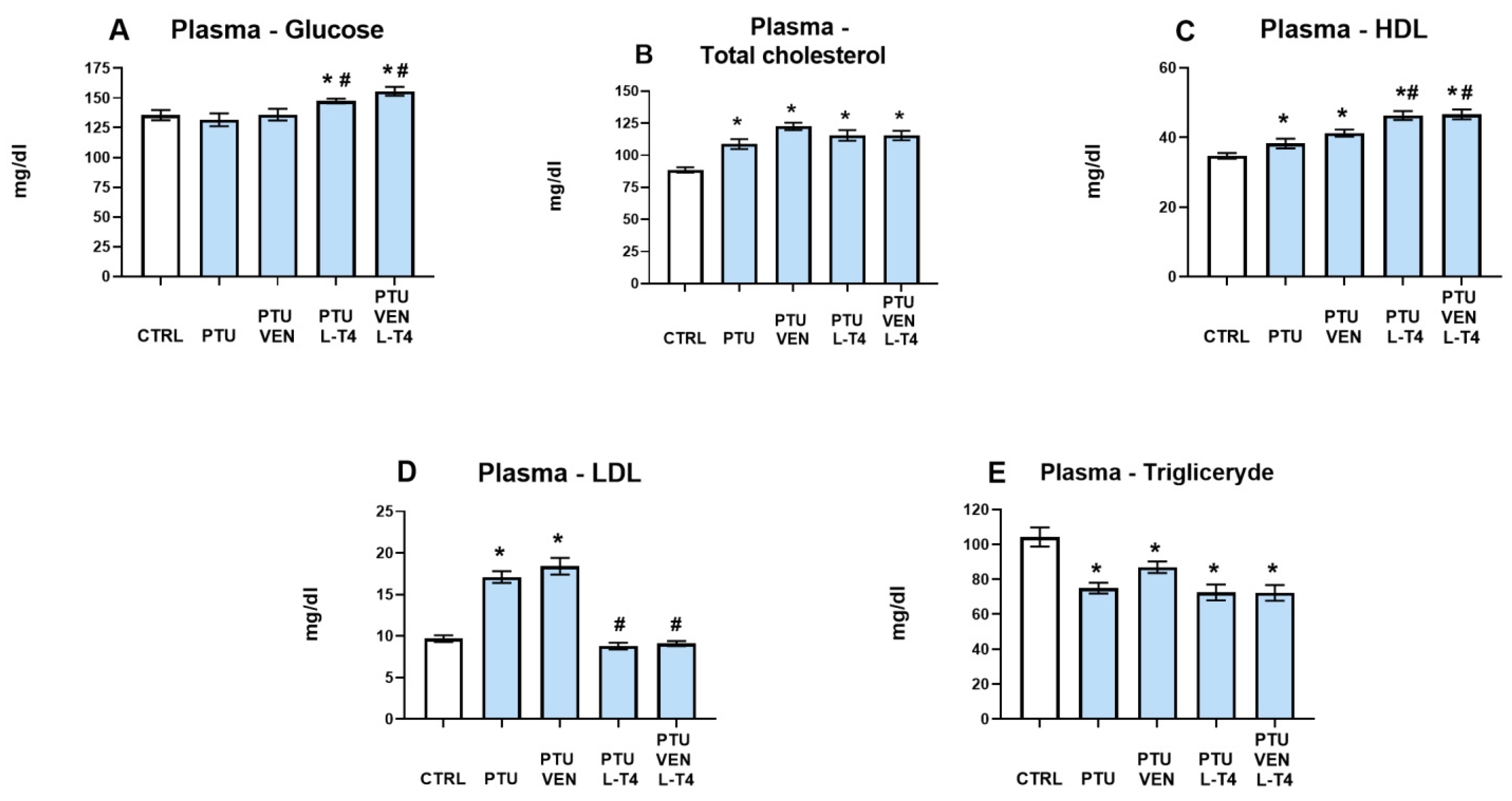
2.5.2. Glucose, Total Cholesterol, HDL, LDL, and Triglyceride Measurements in Plasma
2.5.3. Tissue Homogenate Preparation and Determination of Protein Concentration
2.5.4. Preparation of Mitochondria-Enriched and Cytosolic Fractions
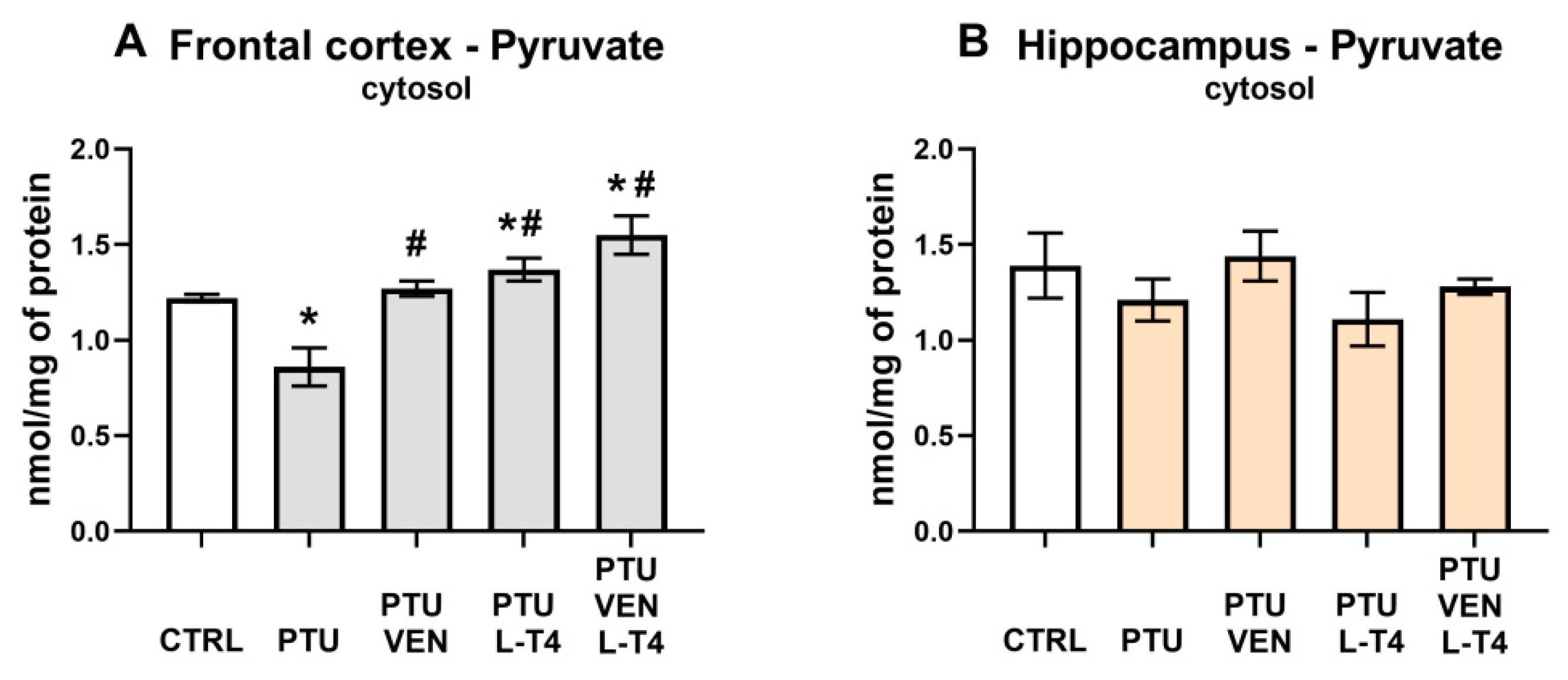
2.5.5. Pyruvate Measurement
2.5.6. Pyruvate Dehydrogenase (PDH) Expression and Activity Estimation
2.5.7. Lactate Measurement
2.5.8. Protein Levels of the OXPHOS Complexes, Monocarboxylate Transporters (MCT2, MCT4), Hydroxycarboxylic Acid Receptor (HCAR1), Glucocorticoid Receptor (GR), FK506-Binding Protein 51 (FKBP51), Bax, Beclin 1, and Caspase-1—Western Blot Analysis
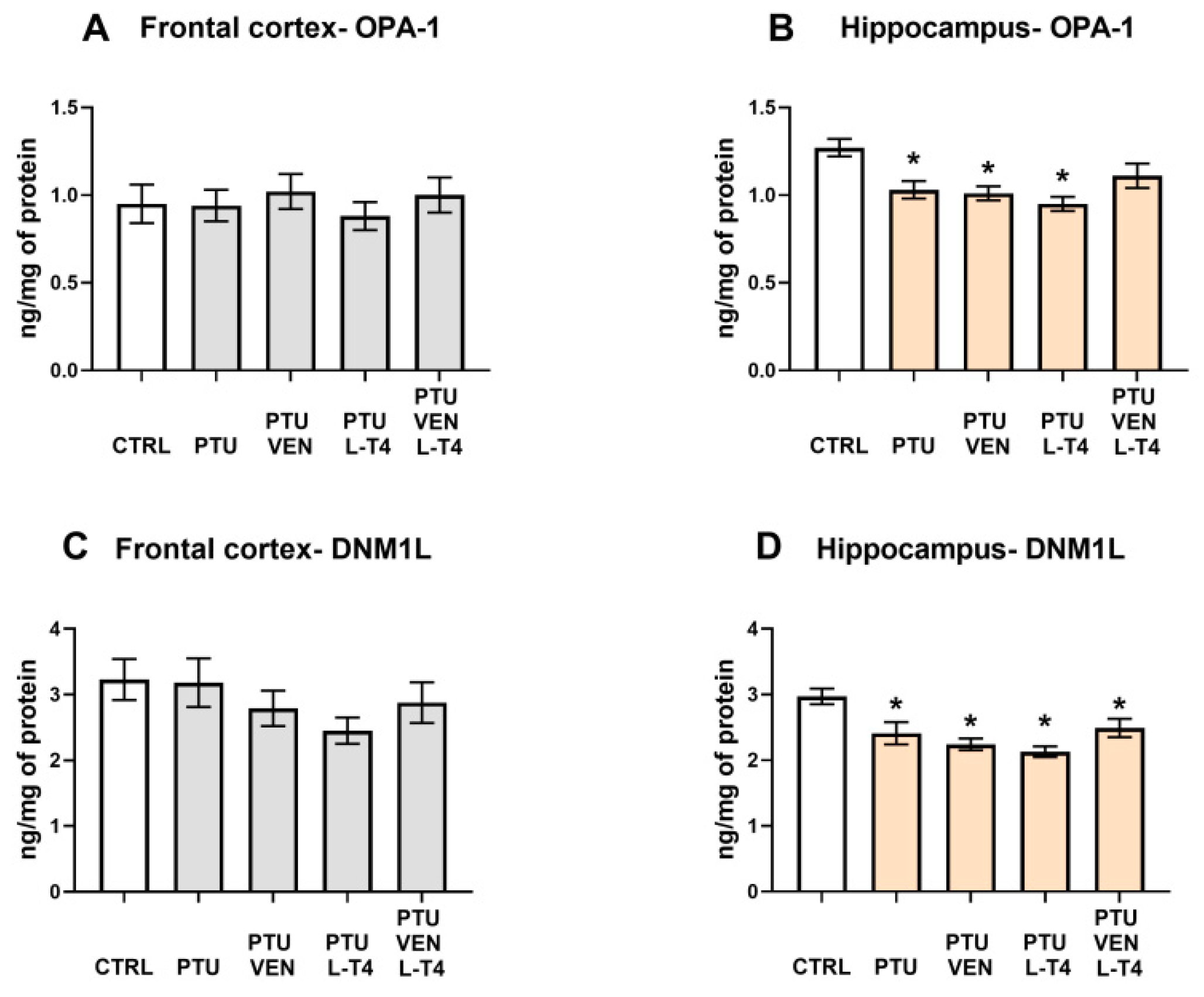
2.5.9. OPA1, DNM1L, GLT-1, GLAST Protein Levels
2.6. Statistical Analysis
3. Results
3.1. Light-Dark Box Test
3.2. Thyroid Hormone Plasma Concentrations
3.3. Glucose Level and Lipid Panel (Total Cholesterol, HDL, LDL, Triglyceride) in Plasma
3.4. Pyruvate Level in the Brain
3.5. Concentration and Activity of PDH in the Brain
3.6. Lactate Level in the Brain
3.7. Hydroxycarboxylic Acid Receptor 1 (HCAR1) Level in the Brain
3.8. Monocarboxylate Transporters MCT2 and MCT4 Levels in the Brain
3.9. Levels of Oxidative Phosphorylation (OXPHOS) Complexes in the Mitochondria-Enriched Fraction of the Brain
3.10. Levels of Mitochondrial Fusion Factor, OPA-1, and Fission Factor, DNM1L
3.11. Levels of Glucocorticoid Receptor (GR) and FKBP51-Factor Regulating GR-Function in the Brain
3.12. Levels of the Cell Damage Markers Caspase-1 p20, Bax, and Beclin1 in the Brain
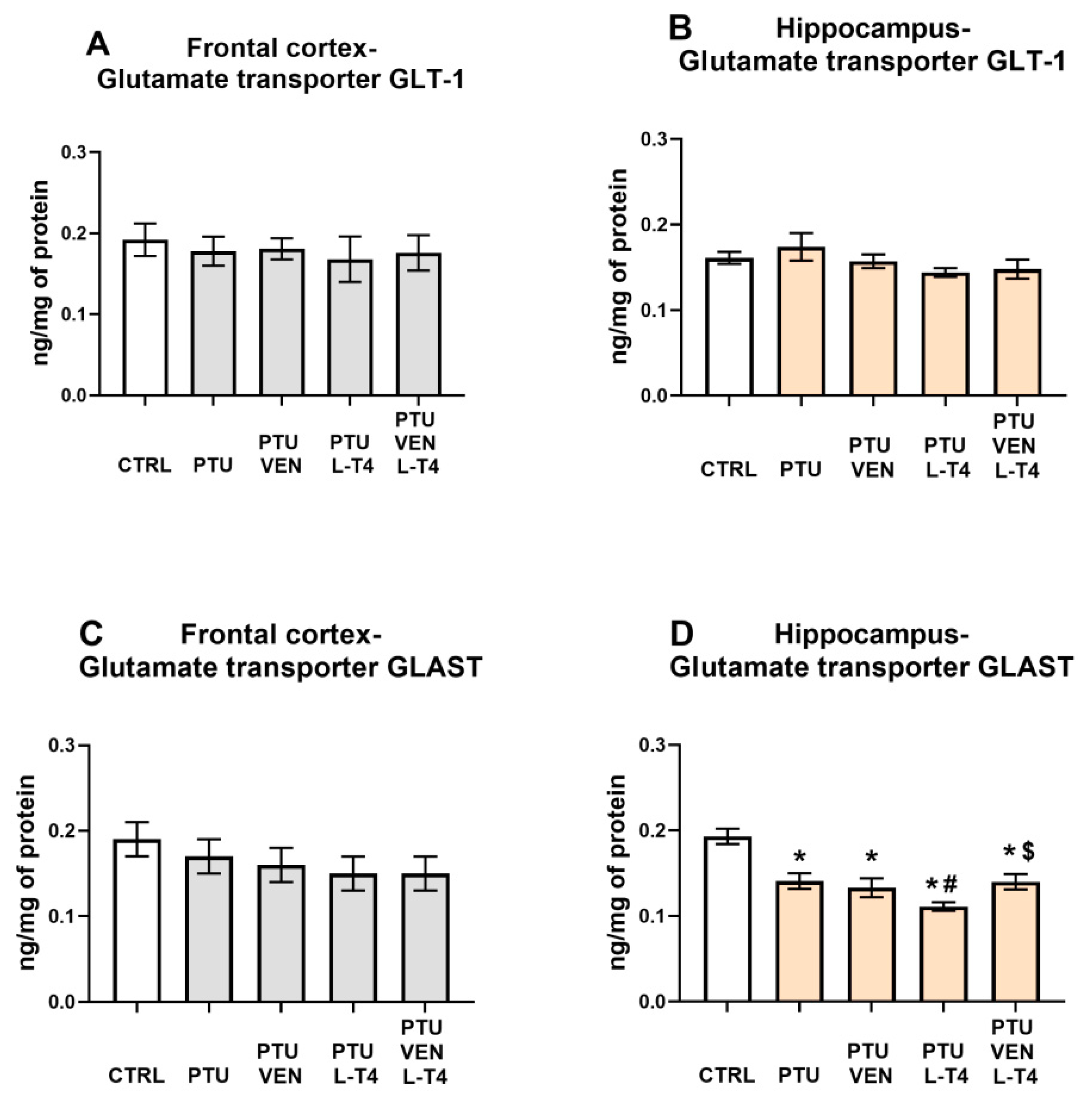
3.13. Concentrations of the Glutamate Transporters GLT-1 and GLAST in the Brain
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Szymańska, M.; Budziszewska, B.; Jaworska-Feil, L.; Basta-Kaim, A.; Kubera, M.; Leśkiewicz, M.; Regulska, M.; Lasoń, W. The effect of antidepressant drugs on the HPA axis activity, glucocorticoid receptor level and FKBP51 concentration in prenatally stressed rats. Psychoneuroendocrinology 2009, 34, 822–832. [Google Scholar] [CrossRef]
- Maes, M. Evidence for an immune response in major depression: A review and hypothesis. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1995, 19, 11–38. [Google Scholar] [CrossRef]
- Streck, E.L.; Gonçalves, C.L.; Furlanetto, C.B.; Scaini, G.; Dal-Pizzol, F.; Quevedo, J. Mitochondria and the central nervous system: Searching for a pathophysiological basis of psychiatric disorders. Rev. Bras. Psiquiatr. 2014, 36, 156–167. [Google Scholar] [CrossRef]
- Horowitz, M.A.; Zunszain, P.A. Neuroimmune and neuroendocrine abnormalities in depression: Two sides of the same coin. Ann. N. Y. Acad. Sci. 2015, 1351, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Munoz, F.; Alamo, C. Monoaminergic Neurotransmission: The History of the Discovery of Antidepressants from 1950s Until Today. Curr. Pharm. Des. 2009, 15, 1563–1586. [Google Scholar] [CrossRef] [PubMed]
- Enache, D.; Pariante, C.M.; Mondelli, V. Markers of central inflammation in major depressive disorder: A systematic review and meta-analysis of studies examining cerebrospinal fluid, positron emission tomography and post-mortem brain tissue. Brain Behav. Immun. 2019, 81, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Gao, Z.; Hu, H. Microglia in depression: Current perspectives. Sci. China Life Sci. 2020. [Google Scholar] [CrossRef]
- Zunszain, P.A.; Anacker, C.; Cattaneo, A.; Carvalho, L.A.; Pariante, C.M. Glucocorticoids, cytokines and brain abnormalities in depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 722–729. [Google Scholar] [CrossRef]
- Pariante, C.M. Why are depressed patients inflamed? A reflection on 20 years of research on depression, glucocorticoid resistance and inflammation. Eur. Neuropsychopharmacol. 2017, 27, 554–559. [Google Scholar] [CrossRef]
- Jou, S.-H.; Chiu, N.-Y.; Liu, C.-S. Mitochondrial dysfunction and psychiatric disorders. Chang Gung Med. J. 2009, 32, 370–379. [Google Scholar]
- Stork, C.; Renshaw, P.F. Mitochondrial dysfunction in bipolar disorder: Evidence from magnetic resonance spectroscopy research. Mol. Psychiatry 2005, 10, 900–919. [Google Scholar] [CrossRef] [PubMed]
- Detka, J.; Kurek, A.; Kucharczyk, M.; Głombik, K.; Basta-Kaim, A.; Kubera, M.; Lasoń, W.; Budziszewska, B. Brain glucose metabolism in an animal model of depression. Neuroscience 2015, 295. [Google Scholar] [CrossRef] [PubMed]
- Hage, M.P.; Azar, S.T. The Link between Thyroid Function and Depression. J. Thyroid Res. 2012, 2012, 1–8. [Google Scholar] [CrossRef]
- Bauer, M.; Berman, S.M.; Schlagenhauf, F.; Voytek, B.; Rasgon, N.; Mandelkern, M.A.; Whybrow, P.C.; London, E.D. Regional cerebral glucose metabolism and anxiety symptoms in bipolar depression: Effects of levothyroxine. Psychiatry Res. Neuroimaging 2010, 181, 71–76. [Google Scholar] [CrossRef]
- Bauer, M.; Heinz, A.; Whybrow, P.C. Thyroid hormones, serotonin and mood: Of synergy and significance in the adult brain. Mol. Psychiatry 2002, 7, 140–156. [Google Scholar] [CrossRef]
- De Escobar, G.M.; Obregón, M.J.; Del Rey, F.E. Role of thyroid hormone during early brain development. Eur. J. Endocrinol. Suppl. 2004, 151, 25–37. [Google Scholar] [CrossRef]
- Fernández-Lamo, I.; Montero-Pedrazuela, A.; Delgado-García, J.M.; Guadaño-Ferraz, A.; Gruart, A. Effects of thyroid hormone replacement on associative learning and hippocampal synaptic plasticity in adult hypothyroid rats. Eur. J. Neurosci. 2009, 30, 679–692. [Google Scholar] [CrossRef]
- Yousef, M.; Babür, E.; Delibaş, S.; Tan, B.; Çimen, A.; Dursun, N.; Süer, C. Adult-Onset Hypothyroidism Alters the Metaplastic Properties of Dentate Granule Cells by Decreasing Akt Phosphorylation. J. Mol. Neurosci. 2019, 68, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Loh, H.H.; Lim, L.L.; Yee, A.; Loh, H.S. Association between subclinical hypothyroidism and depression: An updated systematic review and meta-analysis 11 Medical and Health Sciences 1117 Public Health and Health Services 11 Medical and Health Sciences 1103 Clinical Sciences. BMC Psychiatry 2019, 19, 1–10. [Google Scholar] [CrossRef]
- Kelly, T. A hypothesis on the mechanism of action of high-dose thyroid in refractory mood disorders. Med. Hypotheses 2016, 97, 16–21. [Google Scholar] [CrossRef]
- Kelly, T.; Lieberman, D.Z. The use of triiodothyronine as an augmentation agent in treatment-resistant bipolar II and bipolar disorder NOS. J. Affect. Disord. 2009, 116, 222–226. [Google Scholar] [CrossRef]
- Pinna, G.; Brödel, O.; Visser, T.; Jeitner, A.; Grau, H.; Eravci, M.; Meinhold, H.; Baumgartner, A. Concentrations of seven iodothyronine metabolites in brain regions and the liver of the adult rat. Endocrinology 2002, 143, 1789–1800. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Głombik, K.; Detka, J.; Kurek, A.; Budziszewska, B. Impaired Brain Energy Metabolism: Involvement in Depression and Hypothyroidism. Front. Neurosci. 2020, 14, 1320. [Google Scholar] [CrossRef] [PubMed]
- Głombik, K.; Detka, J.; Bobula, B.; Bąk, J.; Kusek, M.; Tokarski, K.; Budziszewska, B. Contribution of hypothyroidism to cognitive impairment and hippocampal synaptic plasticity regulation in an animal model of depression. Int. J. Mol. Sci. 2021, 22, 1599. [Google Scholar] [CrossRef]
- Whybrow, P.C. A Hypothesis of Thyroid-Catecholamine-Receptor Interaction. Arch. Gen. Psychiatry 1981, 38, 106. [Google Scholar] [CrossRef]
- Will, C.C.; Aird, F.; Redei, E.E. Selectively bred Wistar-Kyoto rats: An animal model of depression and hyper-responsiveness to antidepressants. Mol. Psychiatry 2003, 8, 925–932. [Google Scholar] [CrossRef]
- Aleksandrova, L.R.; Wang, Y.T.; Phillips, A.G. Evaluation of the Wistar-Kyoto rat model of depression and the role of synaptic plasticity in depression and antidepressant response. Neurosci. Biobehav. Rev. 2019, 105, 1–23. [Google Scholar] [CrossRef]
- Willner, P.; Gruca, P.; Lason, M.; Tota-Glowczyk, K.; Litwa, E.; Niemczyk, M.; Papp, M. Validation of chronic mild stress in the Wistar-Kyoto rat as an animal model of treatment-resistant depression. Behav. Pharmacol. 2019, 30, 239–250. [Google Scholar] [CrossRef]
- Siegmann, E.M.; Müller, H.H.O.; Luecke, C.; Philipsen, A.; Kornhuber, J.; Grömer, T.W. Association of depression and anxiety disorders with autoimmune thyroiditis: A systematic review and meta-analysis. JAMA Psychiatry 2018, 75, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Powell, C.L.; Davidson, A.R.; Brown, A.M. Universal Glia to Neurone Lactate Transfer in the Nervous System: Physiological Functions and Pathological Consequences. Biosensors 2020, 10, 183. [Google Scholar] [CrossRef]
- Yin, Y.N.; Hu, J.; Wei, Y.L.; Li, Z.L.; Luo, Z.C.; Wang, R.Q.; Yang, K.X.; Li, S.J.; Li, X.W.; Yang, J.M.; et al. Astrocyte-Derived Lactate Modulates the Passive Coping Response to Behavioral Challenge in Male Mice. Neurosci. Bull. 2020, 37, 1–14. [Google Scholar] [CrossRef]
- Chocyk, A.; Bobula, B.; Dudys, D.; Przyborowska, A.; Majcher-Maślanka, I.; Hess, G.; Wędzony, K. Early-life stress affects the structural and functional plasticity of the medial prefrontal cortex in adolescent rats. Eur. J. Neurosci. 2013, 38, 2089–2107. [Google Scholar] [CrossRef] [PubMed]
- Chamera, K.; Szuster-Głuszczak, M.; Trojan, E.; Basta-Kaim, A. Maternal Immune Activation Sensitizes Male Offspring Rats to Lipopolysaccharide-Induced Microglial Deficits Involving the Dysfunction of CD200-CD200R and CX3CL1-CX3CR1 Systems. Cells 2020, 9, 1676. [Google Scholar] [CrossRef]
- Trojan, E.; Głombik, K.; Ślusarczyk, J.; Budziszewska, B.; Kubera, M.; Roman, A.; Lasoń, W.; Basta-Kaim, A. The Beneficial Impact of Antidepressant Drugs on Prenatal Stress-Evoked Malfunction of the Insulin-Like Growth Factor-1 (IGF-1) Protein Family in the Olfactory Bulbs of Adult Rats. Neurotox. Res. 2016, 29. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Wernicke, C.; Hellmann, J.; Zięba, B.; Kuter, K.; Ossowska, K.; Frenzel, M.; Dencher, N.A.; Rommelspacher, H. 9-Methyl-β-carboline has restorative effects in an animal model of Parkinson’s disease. Pharmacol. Rep. 2010, 62, 35–53. [Google Scholar] [CrossRef]
- Głombik, K.; Detka, J.; Góralska, J.; Kurek, A.; Solnica, B.; Budziszewska, B. Brain Metabolic Alterations in Rats Showing Depression-Like and Obesity Phenotypes. Neurotox. Res. 2020, 37. [Google Scholar] [CrossRef] [PubMed]
- Pilhatsch, M.; Stamm, T.J.; Stahl, P.; Lewitzka, U.; Berghöfer, A.; Sauer, C.; Gitlin, M.; Frye, M.A.; Whybrow, P.C.; Bauer, M. Treatment of bipolar depression with supraphysiologic doses of levothyroxine: A randomized, placebo-controlled study of comorbid anxiety symptoms. Int. J. Bipolar Disord. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Lane, S.; Korsak, A.; Paton, J.F.R.; Gourine, A.V.; Kasparov, S.; Teschemacher, A.G. Lactate-mediated glia-neuronal signalling in the mammalian brain. Nat. Commun. 2014, 5, 1–13. [Google Scholar] [CrossRef]
- Pellerin, L.; Magistretti, P.J. Glutamate uptake into astrocytes stimulates aerobic glycolysis: A mechanism coupling neuronal activity to glucose utilization. Proc. Natl. Acad. Sci. USA 1994, 91, 10625–10629. [Google Scholar] [CrossRef]
- Suzuki, A.; Stern, S.A.; Bozdagi, O.; Huntley, G.W.; Walker, R.H.; Magistretti, P.J.; Alberini, C.M. Astrocyte-neuron lactate transport is required for long-term memory formation. Cell 2011, 144, 810–823. [Google Scholar] [CrossRef]
- Pellerin, L.; Magistretti, P.J. Food for Thought: Challenging the Dogmas. J. Cereb. Blood Flow Metab. 2003, 23, 1282–1286. [Google Scholar] [CrossRef] [PubMed]
- Morland, C.; Lauritzen, K.H.; Puchades, M.; Holm-Hansen, S.; Andersson, K.; Gjedde, A.; Attramadal, H.; Storm-Mathisen, J.; Bergersen, L.H. The lactate receptor, G-protein-coupled receptor 81/hydroxycarboxylic acid receptor 1: Expression and action in brain. J. Neurosci. Res. 2015, 93, 1045–1055. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Ding, X.; Song, Q.; Han, Z.; Yao, H.; Ding, J.; Hu, G. Lactate enhances Arc/arg3.1 expression through hydroxycarboxylic acid receptor 1-β-arrestin2 pathway in astrocytes. Neuropharmacology 2020, 171, 108084. [Google Scholar] [CrossRef] [PubMed]
- Zorov, D.; Vorobjev, I.; Popkov, V.; Babenko, V.; Zorova, L.; Pevzner, I.; Silachev, D.; Zorov, S.; Andrianova, N.; Plotnikov, E. Lessons from the Discovery of Mitochondrial Fragmentation (Fission): A Review and Update. Cells 2019, 8, 175. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Glucocorticoids, depression, and mood disorders: Structural remodeling in the brain. Metabolism 2005, 54, 20–23. [Google Scholar] [CrossRef]
- Yu, S.; Holsboer, F.; Almeida, O.F.X. Neuronal actions of glucocorticoids: Focus on depression. J. Steroid Biochem. Mol. Biol. 2008, 108, 300–309. [Google Scholar] [CrossRef] [PubMed]
- De Kloet, E.R.; DeRijk, R.H.; Meijer, O.C. Therapy insight: Is there an imbalanced response of mineralocorticoid and glucocorticoid receptors in depression? Nat. Clin. Pract. Endocrinol. Metab. 2007, 3, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Pariante, C.M.; Lightman, S.L. The HPA axis in major depression: Classical theories and new developments. Trends Neurosci. 2008, 31, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Kurek, A.; Głombik, K.; Detka, J.; Basta-Kaim, A.; Kubera, M.; Lasoń, W.; Budziszewska, B. Regulators of glucocorticoid receptor function in an animal model of depression and obesity. J. Neuroendocrinol. 2018, 30, e12591. [Google Scholar] [CrossRef]
- Binder, E.B. The role of FKBP5, a co-chaperone of the glucocorticoid receptor in the pathogenesis and therapy of affective and anxiety disorders. Psychoneuroendocrinology 2009, 34, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Talhada, D.; Feiteiro, J.; Costa, A.R.; Talhada, T.; Cairraõ, E.; Wieloch, T.; Englund, E.; Santos, C.R.; Goncąlves, I.; Ruscher, K. Triiodothyronine modulates neuronal plasticity mechanisms to enhance functional outcome after stroke. Acta Neuropathol. Commun. 2019, 7, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Wang, Y.; Furukawa, K.; Fu, W.; Ouyang, X.; Mattson, M.P. Evidence that caspase-1 is a negative regulator of AMPA receptor-mediated long-term potentiation at hippocampal synapses. J. Neurochem. 2006, 97, 1104–1110. [Google Scholar] [CrossRef]
- Sahin, E. Hypothyroidism Increases Expression of Sterile Inflammation Proteins in Rat Heart Tissue. Acta Endocrinol. 2019, 15, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-Y.; Davis, F.B.; Luidens, M.K.; Mousa, S.A.; Cao, J.H.; Zhou, M.; Davis, P.J. Molecular Basis for Certain Neuroprotective Effects of Thyroid Hormone. Front. Mol. Neurosci. 2011, 4, 1–6. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Brain Structure | WKY | WKY + PTU | WKY + PTU + VEN | WKY + PTU + L-T4 | WKY + PTU + VEN + L-T4 |
|---|---|---|---|---|---|---|
| HCAR1 | Frontal Cortex | 1.00 ± 0.10 | 0.94 ± 0.07 | 0.97 ± 0.11 | 1.01 ± 0.11 | 0.93 ± 0.08 |
| Hippocampus | 1.00 ± 0.08 | 0.92 ± 0.08 | 0.92 ± 0.09 | 0.98 ± 0.09 | 1.00 ± 0.11 | |
| MCT2 | Frontal Cortex | 1.00 ± 0.11 | 1.08 ± 0.13 | 0.95 ± 0.07 | 1.10 ± 0.09 | 1.00 ± 0.08 |
| Hippocampus | 1.00 ± 0.08 | 0.89 ± 0.06 | 0.90 ± 0.04 | 0.96 ± 0.07 | 0.85 ± 0.05 | |
| MCT4 | Frontal Cortex | 1.00 ± 0.07 | 0.80 ± 0.08 | 0.91 ± 0.07 | 1.02 ± 0.08 | 0.87 ± 0.08 |
| Hippocampus | 1.00 ± 0.08 | 0.82 ± 0.08 | 0.86 ± 0.06 | 0.89 ± 0.08 | 0.84 ± 0.04 | |
 | ||||||
| Protein | Brain Structure | WKY | WKY + PTU | WKY + PTU + VEN | WKY + PTU + L-T4 | WKY + PTU + VEN + L-T4 |
|---|---|---|---|---|---|---|
| Bax | Frontal Cortex | 1.00 ± 0.08 | 0.94 ± 0.07 | 0.92 ± 0.05 | 0.99 ± 0.06 | 0.98 ± 0.07 |
| Hippocampus | 1.00 ± 0.06 | 1.01 ± 0.05 | 1.01 ± 0.07 | 1.00 ± 0.06 | 1.02 ± 0.06 | |
| beclin 1 | Frontal Cortex | 1.00 ± 0.07 | 0.91 ± 0.07 | 0.88 ± 0.08 | 0.87 ± 0.07 | 0.88 ± 0.07 |
| Hippocampus | 1.00 ± 0.07 | 0.91 ± 0.04 | 0.92 ± 0.06 | 0.94 ± 0.06 | 0.88 ± 0.05 | |
 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Głombik, K.; Detka, J.; Budziszewska, B. Venlafaxine and L-Thyroxine Treatment Combination: Impact on Metabolic and Synaptic Plasticity Changes in an Animal Model of Coexisting Depression and Hypothyroidism. Cells 2021, 10, 1394. https://doi.org/10.3390/cells10061394
Głombik K, Detka J, Budziszewska B. Venlafaxine and L-Thyroxine Treatment Combination: Impact on Metabolic and Synaptic Plasticity Changes in an Animal Model of Coexisting Depression and Hypothyroidism. Cells. 2021; 10(6):1394. https://doi.org/10.3390/cells10061394
Chicago/Turabian StyleGłombik, Katarzyna, Jan Detka, and Bogusława Budziszewska. 2021. "Venlafaxine and L-Thyroxine Treatment Combination: Impact on Metabolic and Synaptic Plasticity Changes in an Animal Model of Coexisting Depression and Hypothyroidism" Cells 10, no. 6: 1394. https://doi.org/10.3390/cells10061394
APA StyleGłombik, K., Detka, J., & Budziszewska, B. (2021). Venlafaxine and L-Thyroxine Treatment Combination: Impact on Metabolic and Synaptic Plasticity Changes in an Animal Model of Coexisting Depression and Hypothyroidism. Cells, 10(6), 1394. https://doi.org/10.3390/cells10061394