
Extracellular Soluble Membranes from Retinal Pigment Epithelial Cells Mediate Apoptosis in Macrophages


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Experimental Autoimmune Uveitis (EAU)
2.3. Extracellular Membrane Isolation
2.4. Caspase 3/7 Activity
2.5. TUNEL Assay
2.6. Immunoblotting for FasL
2.7. Statistical Analysis
3. Results
3.1. RPE ESMs Induce Apoptosis in RAW264.7 Macrophage Cells
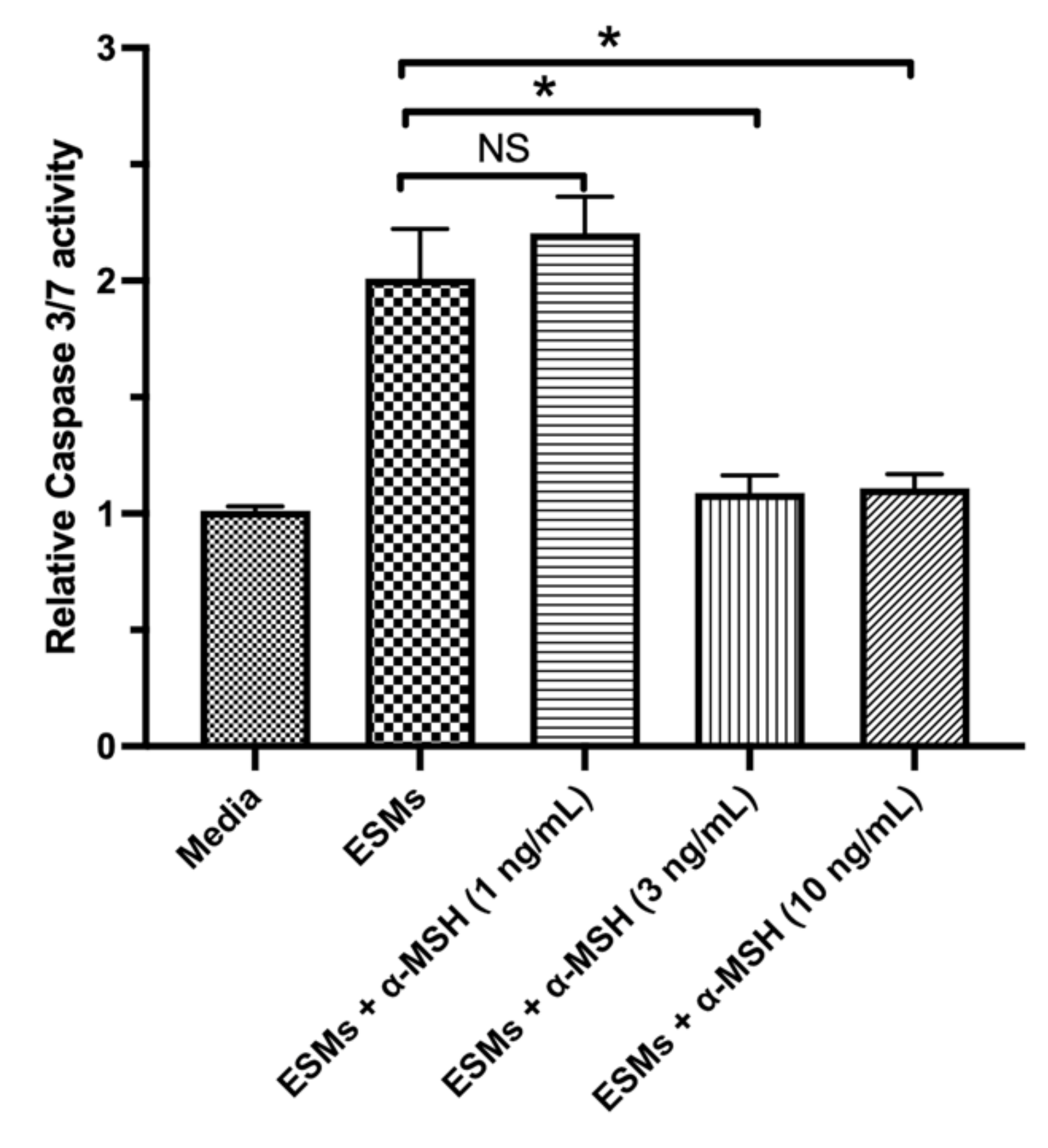
3.2. The Effects of α-MSH on RPE-ESM-Induced Apoptosis
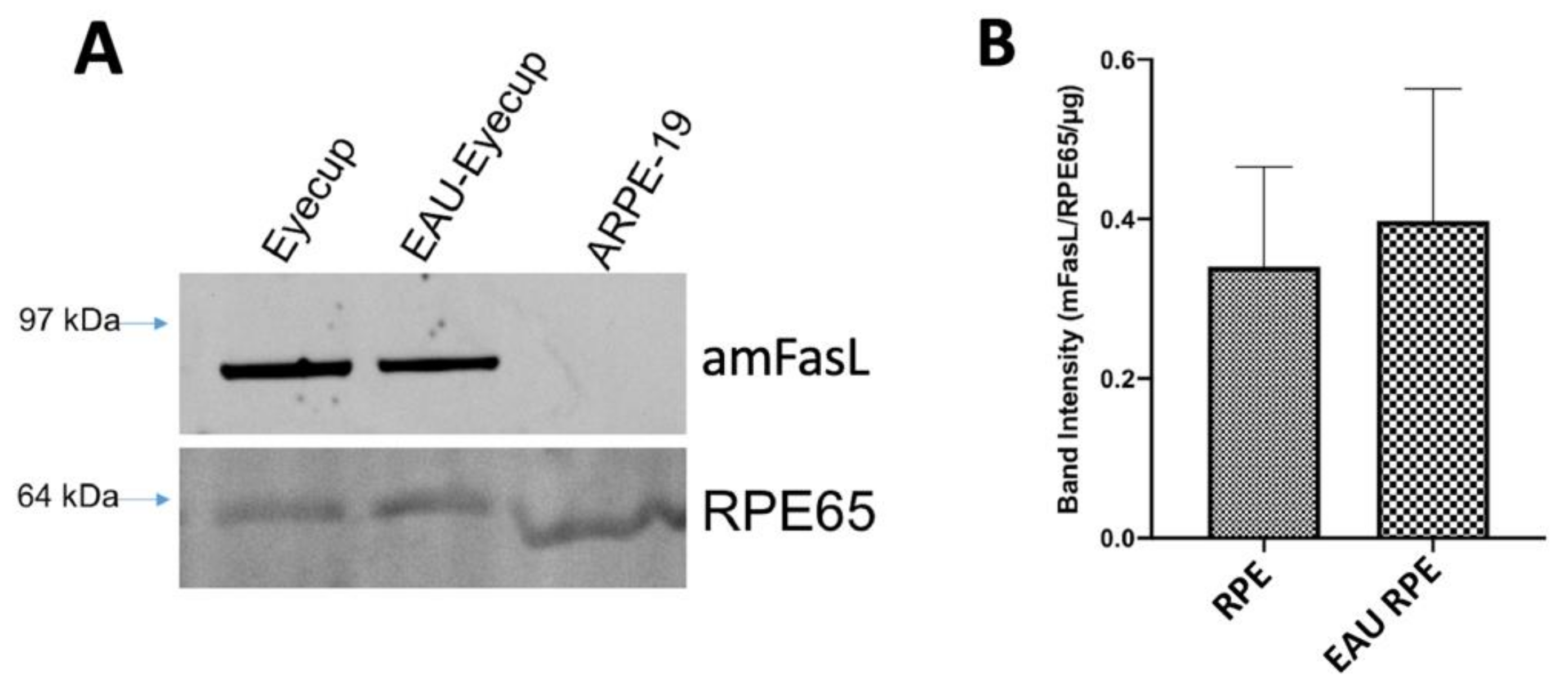
3.3. The Expression of Membrane FasL (mFasL) in RPE ESMs
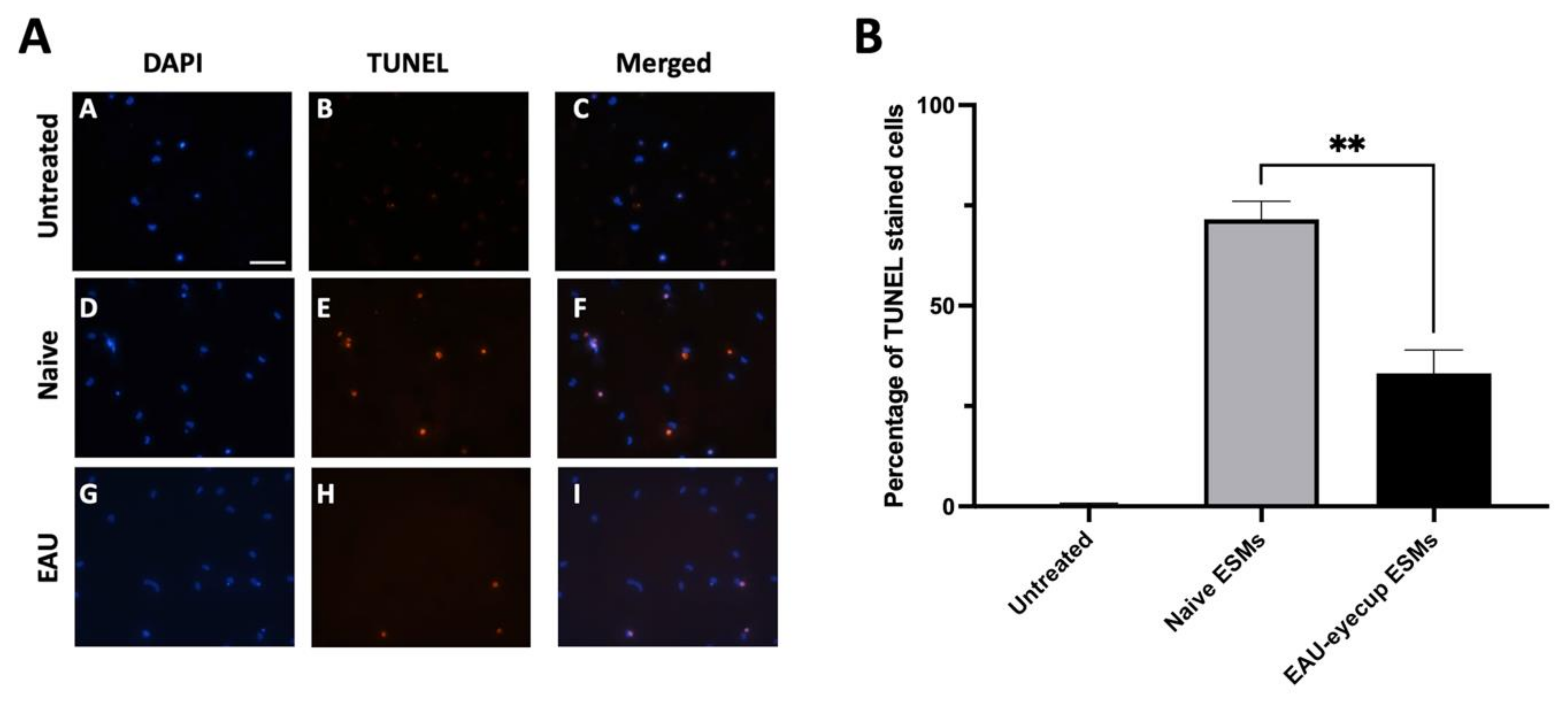
3.4. RPE-ESM Induction of Apoptosis in Primary Macrophages
4. Discussion
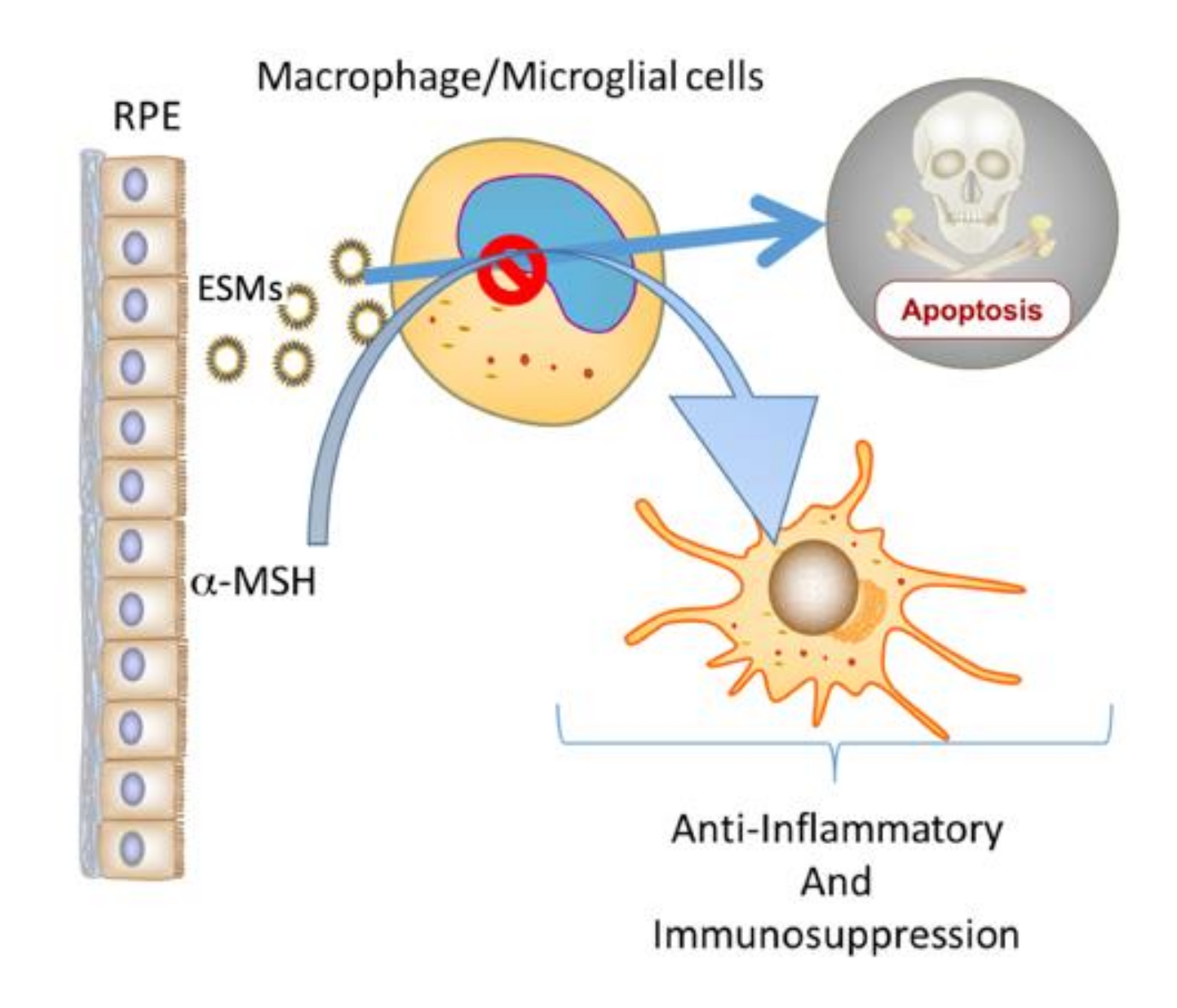
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zamiri, P.; Masli, S.; Kitaichi, N.; Taylor, A.W.; Streilein, J.W. Thrombospondin Plays a Vital Role in the Immune Privilege of the Eye. Investig. Opthalmology Vis. Sci. 2005, 46, 908–919. [Google Scholar] [CrossRef] [PubMed]
- Zamiri, P.; Masli, S.; Streilein, J.W.; Taylor, A.W. Pigment Epithelial Growth Factor Suppresses Inflammation by Modulating Macrophage Activation. Investig. Opthalmology Vis. Sci. 2006, 47, 3912–3918. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.H.; Taylor, A.W. The immune privileged retina mediates an alternative activation of J774A.1 cells. Ocul. Immunol. Inflamm. 2009, 17, 380–389. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kawanaka, N.; Taylor, A.W. Localized retinal neuropeptide regulation of macrophage and microglial cell functionality. J. Neuroimmunol. 2011, 232, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.W.; Ng, T.F. Negative regulators that mediate ocular immune privilege. J. Leukoc. Biol. 2018, 103, 1179–1187. [Google Scholar] [CrossRef]
- Griffith, T.S.; Brunner, T.; Fletcher, S.M.; Green, D.R.; Ferguson, T.A. Fas Ligand-Induced Apoptosis as a Mechanism of Immune Privilege. Science 1995, 270, 1189–1192. [Google Scholar] [CrossRef]
- Hueber, A.; Aduckathil, S.; Kociok, N.; Welsandt, G.; Dinslage, S.; Kirchhof, B.; Esser, P.J. Apoptosis-mediating receptor–ligand systems in human retinal pigment epithelial cells. Graefe’s Arch. Clin. Exp. Ophthalmol. 2002, 240, 551–556. [Google Scholar] [CrossRef]
- Ferguson, T.A.; Griffith, T.S. The Role of Fas Ligand and TNF-Related Apoptosis-Inducing Ligand (TRAIL) in the Ocular Immune Response. Chem. Immunol. Allergy 2007, 92, 140–154. [Google Scholar] [CrossRef]
- Sugita, S.; Horie, S.; Nakamura, O.; Futagami, Y.; Takase, H.; Keino, H.; Aburatani, H.; Katunuma, N.; Ishidoh, K.; Yamamoto, Y.; et al. Retinal Pigment Epithelium-Derived CTLA-2α Induces TGFβ-Producing T Regulatory Cells. J. Immunol. 2008, 181, 7525–7536. [Google Scholar] [CrossRef]
- Ke, Y.; Sun, D.; Jiang, G.; Kaplan, H.J.; Shao, H. PD-L1(hi) retinal pigment epithelium (RPE) cells elicited by inflammatory cytokines induce regulatory activity in uveitogenic T cells. J. Leukoc. Biol. 2010, 88, 1241–1249. [Google Scholar] [CrossRef]
- Wang, E.; Choe, Y.; Ng, T.F.; Taylor, A.W. Retinal Pigment Epithelial Cells Suppress Phagolysosome Activation in Macrophages. Investig. Opthalmology Vis. Sci. 2017, 58, 1266–1273. [Google Scholar] [CrossRef]
- Stenqvist, A.-C.; Nagaeva, O.; Baranov, V.; Mincheva-Nilsson, L. Exosomes Secreted by Human Placenta Carry Functional Fas Ligand and TRAIL Molecules and Convey Apoptosis in Activated Immune Cells, Suggesting Exosome-Mediated Immune Privilege of the Fetus. J. Immunol. 2013, 191, 5515–5523. [Google Scholar] [CrossRef]
- Biasutto, L.; Chiechi, A.; Couch, R.; Liotta, L.A.; Espina, V. Retinal pigment epithelium (RPE) exosomes contain signaling phosphoproteins affected by oxidative stress. Exp. Cell Res. 2013, 319, 2113–2123. [Google Scholar] [CrossRef]
- Knickelbein, J.E.; Liu, B.; Arakelyan, A.; Zicari, S.; Hannes, S.; Chen, P.; Li, Z.; Grivel, J.-C.; Chaigne-Delalande, B.; Sen, H.N.; et al. Modulation of Immune Responses by Extracellular Vesicles From Retinal Pigment Epithelium. Investig. Opthalmol. Vis. Sci. 2016, 57, 4101–4107. [Google Scholar] [CrossRef][Green Version]
- Locke, C.J.; Congrove, N.R.; Dismuke, W.M.; Bowen, T.J.; Stamer, W.D.; McKay, B.S. Controlled exosome release from the retinal pigment epithelium in situ. Exp. Eye Res. 2014, 129, 1–4. [Google Scholar] [CrossRef]
- Wang, A.L.; Lukas, T.J.; Yuan, M.; Du, N.; Tso, M.O.; Neufeld, A.H. Autophagy and Exosomes in the Aged Retinal Pigment Epithelium: Possible Relevance to Drusen Formation and Age-Related Macular Degeneration. PLoS ONE 2009, 4, e4160. [Google Scholar] [CrossRef]
- Alcazar, O.; Hawkridge, A.M.; Collier, T.S.; Cousins, S.W.; Bhattacharya, S.K.; Muddiman, D.C.; Marin-Castano, M.E. Proteomics Characterization of Cell Membrane Blebs in Human Retinal Pigment Epithelium Cells. Mol. Cell. Proteom. 2009, 8, 2201–2211. [Google Scholar] [CrossRef]
- Taylor, A.W.; Dixit, S.; Yu, J. Retinal Pigment Epithelial Cell Line Suppression of Phagolysosome Activation. Int. J. Ophthalmol. Eye Sci. 2015, (Suppl. 2), 1–6. [Google Scholar]
- Lee, D.J.; Preble, J.; Lee, S.; Foster, C.S.; Taylor, A.W. MC5r and A2Ar Deficiencies During Experimental Autoimmune Uveitis Identifies Distinct T cell Polarization Programs and a Biphasic Regulatory Response. Sci. Rep. 2016, 6, 37790. [Google Scholar] [CrossRef]
- Patel, G.K.; Khan, M.A.; Zubair, H.; Srivastava, S.K.; Khushman, M.; Singh, S.; Singh, A.P. Comparative analysis of exosome isolation methods using culture supernatant for optimum yield, purity and downstream applications. Sci. Rep. 2019, 9, 5335. [Google Scholar] [CrossRef]
- Cheng, L.-B.; Cheng, L.; Bi, H.-E.; Zhang, Z.-Q.; Yao, J.; Zhou, X.-Z.; Jiang, Q. Alpha-melanocyte stimulating hormone protects retinal pigment epithelium cells from oxidative stress through activation of melanocortin 1 receptor–Akt–mTOR signaling. Biochem. Biophys. Res. Commun. 2014, 443, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.W. Alpha-Melanocyte Stimulating Hormone (α-MSH) Is a Post-Caspase Suppressor of Apoptosis in RAW 264.7 Macrophages. PLoS ONE 2013, 8, e74488. [Google Scholar] [CrossRef] [PubMed]
- Maisto, R.; Gesualdo, C.; Trotta, M.C.; Grieco, P.; Testa, F.; Simonelli, F.; Barcia, J.M.; D’Amico, M.; Di Filippo, C.; Rossi, S. Melanocortin receptor agonists MCR 1-5 protect photoreceptors from high-glucose damage and restore antioxidant enzymes in primary retinal cell culture. J. Cell. Mol. Med. 2016, 21, 968–974. [Google Scholar] [CrossRef] [PubMed]
- Naveh, N. Melanocortins applied intravitreally delay retinal dystrophy in Royal College of Surgeons rats. Graefe’s Arch. Clin. Exp. Ophthalmol. 2003, 241, 1044–1050. [Google Scholar] [CrossRef]
- Kaminski, V.D.L.; Ellwanger, J.H.; Chies, J.A.B. Extracellular vesicles in host-pathogen interactions and immune regulation—Exosomes as emerging actors in the immunological theater of pregnancy. Heliyon 2019, 5, e02355. [Google Scholar] [CrossRef]
- Holler, N.; Tardivel, A.; Kovacsovics-Bankowski, M.; Hertig, S.; Gaide, O.; Martinon, F.; Tinel, A.; Deperthes, D.; Calderara, S.; Schulthess, T.; et al. Two Adjacent Trimeric Fas Ligands Are Required for Fas Signaling and Formation of a Death-Inducing Signaling Complex. Mol. Cell. Biol. 2003, 23, 1428–1440. [Google Scholar] [CrossRef]
- Maisto, R.; Oltra, M.; Vidal-Gil, L.; Martínez-Gil, N.; Sancho-Pellúz, J.; Di Filippo, C.; Rossi, S.; D’amico, M.; Barcia, J.M.; Romero, F.J. ARPE-19-derived VEGF-containing exosomes promote neovascularization in HUVEC: The role of the melanocortin receptor 5. Cell Cycle 2019, 18, 413–424. [Google Scholar] [CrossRef]
- Clemson, C.M.; Yost, J.; Taylor, A.W. The Role of Alpha-MSH as a Modulator of Ocular Immunobiology Exemplifies Mechanistic Differences between Melanocortins and Steroids. Ocul. Immunol. Inflamm. 2017, 25, 179–189. [Google Scholar] [CrossRef]
- Namba, K.; Kitaichi, N.; Nishida, T.; Taylor, A.W. Induction of regulatory T cells by the immunomodulating cytokines al-pha-melanocyte-stimulating hormone and transforming growth factor-beta 2. J. Leukocyte Biol. 2002, 72, 946–952. [Google Scholar]
- Taylor, A.W.; Alard, P.; Yee, D.G.; Streilein, J.W. Aqueous humor induces transforming growth factor-ß (TGF-ß)-producing regulatory T-cells. Curr. Eye Res. 1997, 16, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.W.; Lee, D.J. The Alpha-Melanocyte Stimulating Hormone Induces Conversion of Effector T Cells into Treg Cells. J. Transplant. 2011, 2011, 246856. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.W.; Streilein, J.W.; Cousins, S.W. Identification of alpha-melanocyte stimulating hormone as a potential immunosuppressive factor in aqueous humor. Curr. Eye Res. 1992, 11, 1199–1206. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.W.; Streilein, J.W.; Cousins, S.W. Alpha-Melanocyte-Stimulating Hormone Suppresses Antigen-Stimulated T Cell Production of Gamma-lnterferon. Neuroimmunomodulation 1994, 1, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.J.; Taylor, A.W. Both MC5r and A2Ar Are Required for Protective Regulatory Immunity in the Spleen of Post–Experimental Autoimmune Uveitis in Mice. J. Immunol. 2013, 191, 4103–4111. [Google Scholar] [CrossRef]
- Lee, D.J.; Taylor, A.W. Following EAU Recovery There Is an Associated MC5r-Dependent APC Induction of Regulatory Immunity in the Spleen. Investig. Opthalmology Vis. Sci. 2011, 52, 8862–8867. [Google Scholar] [CrossRef]
- Cai, S.; Yang, Q.; Hou, M.; Han, Q.; Zhang, H.; Wang, J.; Qi, C.; Bo, Q.; Ru, Y.; Yang, W.; et al. A-Melanocyte-Stimulating Hormone Protects Early Diabetic Retina from Blood-Retinal Barrier Breakdown and Vascular Leakage via MC4R. Cell. Physiol. Biochem. 2018, 45, 505–522. [Google Scholar] [CrossRef]
- Zhang, Y.; Bo, Q.; Wu, W.; Xu, C.; Yu, G.; Ma, S.; Yang, Q.; Cao, Y.; Han, Q.; Ru, Y.; et al. α-Melanocyte-stimulating hormone prevents glutamate excitotoxicity in developing chicken retina via MC4R-mediated down-regulation of microRNA-194. Sci. Rep. 2015, 5, 15812. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanjiv, N.; Osathanugrah, P.; Fraser, E.; Ng, T.F.; Taylor, A.W. Extracellular Soluble Membranes from Retinal Pigment Epithelial Cells Mediate Apoptosis in Macrophages. Cells 2021, 10, 1193. https://doi.org/10.3390/cells10051193
Sanjiv N, Osathanugrah P, Fraser E, Ng TF, Taylor AW. Extracellular Soluble Membranes from Retinal Pigment Epithelial Cells Mediate Apoptosis in Macrophages. Cells. 2021; 10(5):1193. https://doi.org/10.3390/cells10051193
Chicago/Turabian StyleSanjiv, Nayan, Pawarissara Osathanugrah, Emma Fraser, Tat Fong Ng, and Andrew W. Taylor. 2021. "Extracellular Soluble Membranes from Retinal Pigment Epithelial Cells Mediate Apoptosis in Macrophages" Cells 10, no. 5: 1193. https://doi.org/10.3390/cells10051193
APA StyleSanjiv, N., Osathanugrah, P., Fraser, E., Ng, T. F., & Taylor, A. W. (2021). Extracellular Soluble Membranes from Retinal Pigment Epithelial Cells Mediate Apoptosis in Macrophages. Cells, 10(5), 1193. https://doi.org/10.3390/cells10051193