
Effects of a 6 Week Low-Dose Combined Resistance and Endurance Training on T Cells and Systemic Inflammation in the Elderly


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Participants
2.3. Body Characteristics and Physical Capacity
2.4. Exercise Training
2.5. Isolation of Peripheral Blood Mononuclear Cells
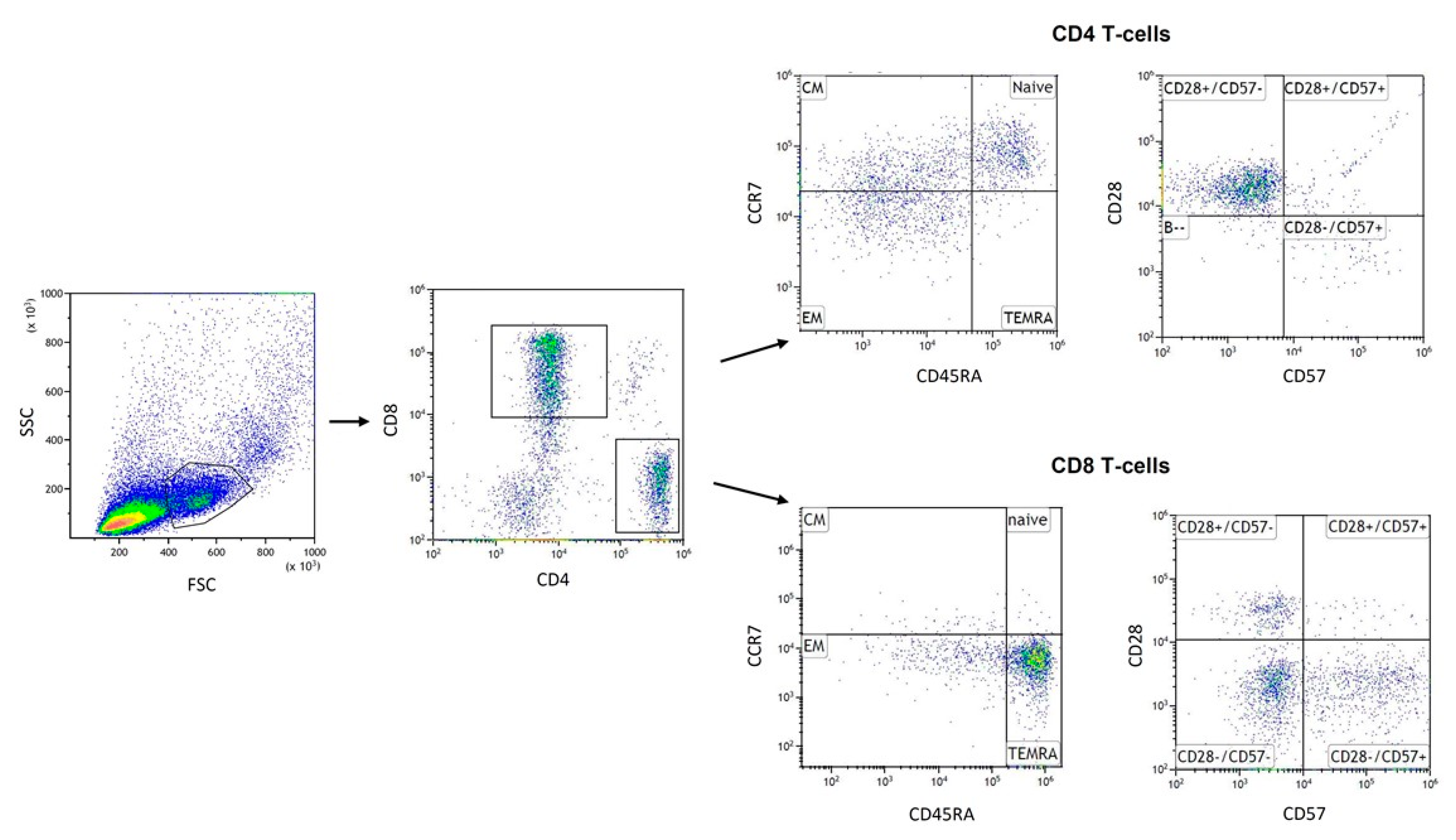
2.6. Flow Cytometry
2.7. Multiplex ELISA Luminex Assay
2.8. Statistical Analysis
3. Results
3.1. Effects of Training Intervention on Health Status and Physical Capacity
3.2. Effects of Training Intervention on T Cell Subpopulations
3.3. Effects of Training Intervention on Plasma Cytokine Levels
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gruver, A.L.; Hudson, L.L.; Sempowski, G.D. Immunosenescence of ageing. J. Pathol. 2007, 211, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Aiello, A.; Farzaneh, F.; Candore, G.; Caruso, C.; Davinelli, S.; Gambino, C.M.; Ligotti, M.E.; Zareian, N.; Accardi, G. Immunosenescence and Its Hallmarks: How to Oppose Aging Strategically? A Review of Potential Options for Therapeutic Intervention. Front. Immunol. 2019, 10, 2247. [Google Scholar] [CrossRef] [PubMed]
- Wikby, A.; Månsson, I.A.; Johansson, B.; Strindhall, J.; Nilsson, S.E. The immune risk profile is associated with age and gender: Findings from three Swedish population studies of individuals 20–100 years of age. Biogerontology 2008, 9, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Farber, D.L.; Yudanin, N.A.; Restifo, N.P. Human memory T cells: Generation, compartmentalization and homeostasis. Nat. Rev. Immunol. 2014, 14, 24–35. [Google Scholar] [CrossRef]
- Xu, W.; Larbi, A. Markers of T Cell Senescence in Humans. Int. J. Mol. Sci. 2017, 18, 1742. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, I.J.; Lalinde Ruiz, N.; Llano León, M.; Martínez Enríquez, L.; Montilla Velásquez, M.D.P.; Ortiz Aguirre, J.P.; Rodríguez Bohórquez, O.M.; Velandia Vargas, E.A.; Hernández, E.D.; Parra López, C.A. Immunosenescence Study of T Cells: A Systematic Review. Front. Immunol. 2020, 11, 604591. [Google Scholar] [CrossRef]
- Di Mitri, D.; Azevedo, R.I.; Henson, S.M.; Libri, V.; Riddell, N.E.; Macaulay, R.; Kipling, D.; Soares, M.V.D.; Battistini, L.; Akbar, A.N. Reversible senescence in human CD4+CD45RA+CD27- memory T cells. J. Immunol. 2011, 187, 2093–2100. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Capri, M.; Monti, D.; Giunta, S.; Olivieri, F.; Sevini, F.; Panourgia, M.P.; Invidia, L.; Celani, L.; Scurti, M.; et al. Inflammaging and anti-inflammaging: A systemic perspective on aging and longevity emerged from studies in humans. Mech. Ageing Dev. 2007, 128, 92–105. [Google Scholar] [CrossRef]
- Franceschi, C.; Campisi, J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69 (Suppl. 1), S4–S9. [Google Scholar] [CrossRef]
- Minciullo, P.L.; Catalano, A.; Mandraffino, G.; Casciaro, M.; Crucitti, A.; Maltese, G.; Morabito, N.; Lasco, A.; Gangemi, S.; Basile, G. Inflammaging and Anti-Inflammaging: The Role of Cytokines in Extreme Longevity. Arch. Immunol. Ther. Exp. (Warsz) 2016, 64, 111–126. [Google Scholar] [CrossRef]
- Dugan, H.L.; Henry, C.; Wilson, P.C. Aging and influenza vaccine-induced immunity. Cell. Immunol. 2020, 348, 103998. [Google Scholar] [CrossRef]
- Nikolich-Zugich, J. Ageing and life-long maintenance of T-cell subsets in the face of latent persistent infections. Nat. Rev. Immunol. 2008, 8, 512–522. [Google Scholar] [CrossRef]
- Pera, A.; Campos, C.; López, N.; Hassouneh, F.; Alonso, C.; Tarazona, R.; Solana, R. Immunosenescence: Implications for response to infection and vaccination in older people. Maturitas 2015, 82, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Cunha, L.L.; Perazzio, S.F.; Azzi, J.; Cravedi, P.; Riella, L.V. Remodeling of the Immune Response With Aging: Immunosenescence and Its Potential Impact on COVID-19 Immune Response. Front. Immunol. 2020, 11, 1748. [Google Scholar] [CrossRef] [PubMed]
- Lian, J.; Yue, Y.; Yu, W.; Zhang, Y. Immunosenescence: A key player in cancer development. J. Hematol. Oncol. 2020, 13, 151. [Google Scholar] [CrossRef]
- Thewissen, M.; Somers, V.; Venken, K.; Linsen, L.; van Paassen, P.; Geusens, P.; Damoiseaux, J.; Stinissen, P. Analyses of immunosenescent markers in patients with autoimmune disease. Clin. Immunol. 2007, 123, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Costantini, E.; D’Angelo, C.; Reale, M. The Role of Immunosenescence in Neurodegenerative Diseases. Mediat. Inflamm. 2018, 2018, 6039171. [Google Scholar] [CrossRef]
- Prather, A.A.; Epel, E.S.; Portela Parra, E.; Coccia, M.; Puterman, E.; Aiello, A.E.; Dhabhar, F.S. Associations between chronic caregiving stress and T cell markers implicated in immunosenescence. Brain Behav. Immun. 2018, 73, 546–549. [Google Scholar] [CrossRef] [PubMed]
- Duggal, N.A.; Niemiro, G.; Harridge, S.D.R.; Simpson, R.J.; Lord, J.M. Can physical activity ameliorate immunosenescence and thereby reduce age-related multi-morbidity? Nat. Rev. Immunol. 2019, 19, 563–572. [Google Scholar] [CrossRef]
- Weyh, C.; Krüger, K.; Strasser, B. Physical Activity and Diet Shape the Immune System during Aging. Nutrients 2020, 12, 622. [Google Scholar] [CrossRef]
- Duggal, N.A.; Pollock, R.D.; Lazarus, N.R.; Harridge, S.; Lord, J.M. Major features of immunesenescence, including reduced thymic output, are ameliorated by high levels of physical activity in adulthood. Aging Cell 2018, 17. [Google Scholar] [CrossRef]
- Philippe, M.; Gatterer, H.; Burtscher, M.; Weinberger, B.; Keller, M.; Grubeck-Loebenstein, B.; Fleckenstein, J.; Alack, K.; Krüger, K. Concentric and Eccentric Endurance Exercise Reverse Hallmarks of T-Cell Senescence in Pre-diabetic Subjects. Front. Physiol. 2019, 10, 684. [Google Scholar] [CrossRef] [PubMed]
- Fairey, A.S.; Courneya, K.S.; Field, C.J.; Bell, G.J.; Jones, L.W.; Mackey, J.R. Randomized controlled trial of exercise and blood immune function in postmenopausal breast cancer survivors. J. Appl. Physiol. 2005, 98, 1534–1540. [Google Scholar] [CrossRef] [PubMed]
- Flynn, M.G.; Fahlman, M.; Braun, W.A.; Lambert, C.P.; Bouillon, L.E.; Brolinson, P.G.; Armstrong, C.W. Effects of resistance training on selected indexes of immune function in elderly women. J. Appl. Physiol. 1999, 86, 1905–1913. [Google Scholar] [CrossRef]
- Dumville, J.C.; Hahn, S.; Miles, J.N.V.; Torgerson, D.J. The use of unequal randomisation ratios in clinical trials: A review. Contemp. Clin. Trials 2006, 27, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Feigenbaum, M.S.; Pollock, M.L. Prescription of resistance training for health and disease. Med. Sci. Sports Exerc. 1999, 31, 38–45. [Google Scholar] [CrossRef]
- Franz, I.-W. Bewegungs- und Sporttherapiein der ambulanten Lungen-sportgruppe. Dtsch. Z. Sportmed. 2001, 52, 44–45. [Google Scholar]
- McBride, J.A.; Striker, R. Imbalance in the game of T cells: What can the CD4/CD8 T-cell ratio tell us about HIV and health? PLoS Pathog. 2017, 13, e1006624. [Google Scholar] [CrossRef] [PubMed]
- Richard, C.; Wadowski, M.; Goruk, S.; Cameron, L.; Sharma, A.M.; Field, C.J. Individuals with obesity and type 2 diabetes have additional immune dysfunction compared with obese individuals who are metabolically healthy. BMJ Open Diabetes Res. Care 2017, 5, e000379. [Google Scholar] [CrossRef] [PubMed]
- Campbell, P.T.; Wener, M.H.; Sorensen, B.; Wood, B.; Chen-Levy, Z.; Potter, J.D.; McTiernan, A.; Ulrich, C.M. Effect of exercise on in vitro immune function: A 12-month randomized, controlled trial among postmenopausal women. J. Appl. Physiol. 2008, 104, 1648–1655. [Google Scholar] [CrossRef]
- Kapasi, Z.F.; Ouslander, J.G.; Schnelle, J.F.; Kutner, M.; Fahey, J.L. Effects of an exercise intervention on immunologic parameters in frail elderly nursing home residents. J. Gerontol. A Biol. Sci. Med. Sci. 2003, 58, 636–643. [Google Scholar] [CrossRef]
- Shimizu, K.; Suzuki, N.; Imai, T.; Aizawa, K.; Nanba, H.; Hanaoka, Y.; Kuno, S.; Mesaki, N.; Kono, I.; Akama, T. Monocyte and T-cell responses to exercise training in elderly subjects. J. Strength Cond. Res. 2011, 25, 2565–2572. [Google Scholar] [CrossRef]
- Fletcher, J.M.; Vukmanovic-Stejic, M.; Dunne, P.J.; Birch, K.E.; Cook, J.E.; Jackson, S.E.; Salmon, M.; Rustin, M.H.; Akbar, A.N. Cytomegalovirus-specific CD4+ T cells in healthy carriers are continuously driven to replicative exhaustion. J. Immunol. 2005, 175, 8218–8225. [Google Scholar] [CrossRef] [PubMed]
- Minuzzi, L.G.; Rama, L.; Chupel, M.U.; Rosado, F.; Dos Santos, J.V.; Simpson, R.; Martinho, A.; Paiva, A.; Teixeira, A.M. Effects of lifelong training on senescence and mobilization of T lymphocytes in response to acute exercise. Exerc. Immunol. Rev. 2018, 24, 72–84. [Google Scholar]
- Bautmans, I.; Salimans, L.; Njemini, R.; Beyer, I.; Lieten, S.; Liberman, K. The effects of exercise interventions on the inflammatory profile of older adults: A systematic review of the recent literature. Exp. Gerontol. 2021, 146, 111236. [Google Scholar] [CrossRef]
- Pedersen, B.K. Anti-inflammatory effects of exercise: Role in diabetes and cardiovascular disease. Eur. J. Clin. Investig. 2017, 47, 600–611. [Google Scholar] [CrossRef]
- Zheng, G.; Qiu, P.; Xia, R.; Lin, H.; Ye, B.; Tao, J.; Chen, L. Effect of Aerobic Exercise on Inflammatory Markers in Healthy Middle-Aged and Older Adults: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Front. Aging Neurosci. 2019, 11, 98. [Google Scholar] [CrossRef]
- Pakiz, B.; Flatt, S.W.; Bardwell, W.A.; Rock, C.L.; Mills, P.J. Effects of a weight loss intervention on body mass, fitness, and inflammatory biomarkers in overweight or obese breast cancer survivors. Int. J. Behav. Med. 2011, 18, 333–341. [Google Scholar] [CrossRef]
- Astengo, M.; Dahl, A.; Karlsson, T.; Mattsson-Hultén, L.; Wiklund, O.; Wennerblom, B. Physical training after percutaneous coronary intervention in patients with stable angina: Effects on working capacity, metabolism, and markers of inflammation. Eur. J. Cardiovasc. Prev. Rehabil. 2010, 17, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Palmefors, H.; DuttaRoy, S.; Rundqvist, B.; Börjesson, M. The effect of physical activity or exercise on key biomarkers in atherosclerosis—A systematic review. Atherosclerosis 2014, 235, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Trøseid, M.; Lappegård, K.T.; Claudi, T.; Damås, J.K.; Mørkrid, L.; Brendberg, R.; Mollnes, T.E. Exercise reduces plasma levels of the chemokines MCP-1 and IL-8 in subjects with the metabolic syndrome. Eur. Heart J. 2004, 25, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Kochumon, S.; Al Madhoun, A.; Al-Rashed, F.; Thomas, R.; Sindhu, S.; Al-Ozairi, E.; Al-Mulla, F.; Ahmad, R. Elevated adipose tissue associated IL-2 expression in obesity correlates with metabolic inflammation and insulin resistance. Sci. Rep. 2020, 10, 16364. [Google Scholar] [CrossRef]
- Cerqueira, É.; Marinho, D.A.; Neiva, H.P.; Lourenço, O. Inflammatory Effects of High and Moderate Intensity Exercise-A Systematic Review. Front. Physiol. 2019, 10, 1550. [Google Scholar] [CrossRef]
- Tam, C.S.; Garnett, S.P.; Cowell, C.T.; Heilbronn, L.K.; Lee, J.W.; Wong, M.; Baur, L.A. IL-6, IL-8 and IL-10 levels in healthy weight and overweight children. Horm. Res. Paediatr. 2010, 73, 128–134. [Google Scholar] [CrossRef]
- Brenner, D.R.; Ruan, Y.; Adams, S.C.; Courneya, K.S.; Friedenreich, C.M. The impact of exercise on growth factors (VEGF and FGF2): Results from a 12-month randomized intervention trial. Eur. Rev. Aging Phys. Act. 2019, 16, 8. [Google Scholar] [CrossRef]
- Zhang, Q.; Fang, W.; Ma, L.; Wang, Z.-D.; Yang, Y.-M.; Lu, Y.-Q. VEGF levels in plasma in relation to metabolic control, inflammation, and microvascular complications in type-2 diabetes: A cohort study. Medicine (Baltimore) 2018, 97, e0415. [Google Scholar] [CrossRef] [PubMed]
- Smets, P.; Devauchelle-Pensec, V.; Rouzaire, P.-O.; Pereira, B.; Andre, M.; Soubrier, M. Vascular endothelial growth factor levels and rheumatic diseases of the elderly. Arthritis Res. Ther. 2016, 18, 283. [Google Scholar] [CrossRef]
- Trøseid, M.; Lappegård, K.T.; Mollnes, T.E.; Arnesen, H.; Seljeflot, I. The effect of exercise on serum levels of interleukin-18 and components of the metabolic syndrome. Metab. Syndr. Relat. Disord. 2009, 7, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.I.R.; Gomes, P.S.C. Movement velocity in resistance training. Sports Med. 2003, 33, 427–438. [Google Scholar] [CrossRef]
- Ouyang, Q.; Wagner, W.M.; Zheng, W.; Wikby, A.; Remarque, E.J.; Pawelec, G. Dysfunctional CMV-specific CD8(+) T cells accumulate in the elderly. Exp. Gerontol. 2004, 39, 607–613. [Google Scholar] [CrossRef]
- Simpson, R.J.; Bigley, A.B.; Spielmann, G.; LaVoy, E.C.P.; Kunz, H.; Bollard, C.M. Human cytomegalovirus infection and the immune response to exercise. Exerc. Immunol. Rev. 2016, 22, 8–27. [Google Scholar] [PubMed]
- Turner, J.E.; Aldred, S.; Witard, O.C.; Drayson, M.T.; Moss, P.M.; Bosch, J.A. Latent cytomegalovirus infection amplifies CD8 T-lymphocyte mobilisation and egress in response to exercise. Brain Behav. Immun. 2010, 24, 1362–1370. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| TG | CG | |
|---|---|---|
| n (male) | 30 (14) | 10 (7) |
| Anthropometrics | ||
| Age (years) | 70.4 ± 5.3 | 69.8 ± 4.4 |
| Height (cm) | 170.0 ± 7.3 | 171.9 ± 7.3 |
| Weight (kg) | 80.1 ± 11.7 | 77.1 ± 15.8 |
| BMI (kg/m2) | 27.7 ± 3.4 | 25.9 ± 3.5 |
| Body fat (%) | 31.6 ± 7.4 | 25.4 ± 8.6 |
| Health status n (%) | ||
| Participants with no pathologies | 10 (33.3) | 3 (30) |
| Participants with one pathology | 10 (33.3) | 4 (40) |
| Participats with pathologies ≥2 | 10 (33.3) | 3 (30) |
| Pathologies n (%) | ||
| Hypertension | 9 (30) | 5 (50) |
| Cardiovascular disease | 3 (10) | 1 (10) |
| Asthma | 2 (13.7) | 0 (0) |
| Hypercholesterolemia | 3 (10) | 1 (10) |
| Thyroid Disease | 4 (13.3) | 1 (10) |
| Mental disease | 2 (6.7) | 0 (0) |
| Other a | 7 (23.3) | 2 (20) |
| Status of medicine intake n (%) | ||
| Participants with no medicine | 10 (33.3) | 3 (30) |
| Participants with one medicine | 9 (30) | 4 (40) |
| Participats with medicine ≥ 2 | 11 (36.7) | 3 (30) |
| Use of prescribed medicine n (%) | ||
| Anti-hypertensive drugs | 11 (36.7) | 5 (50) |
| Anticoagulants | 5 (16.7) | 2 (20) |
| Statins | 2 (6.7) | 1 (10) |
| Inhaled sympathomimetics | 2 (6.7) | 0 (0) |
| Thyroid drugs | 4 (13.3) | 1 (10) |
| Psychotropic drugs | 3 (10) | 0 (0) |
| Other a | 5 (16.7) | 2 (20) |
| TG | CG | Time Effect | Time × Group Interaction | |||
|---|---|---|---|---|---|---|
| T1 | T2 | T1 | T2 | p | p | |
| BMI (kg/m2) | 27.7 ± 3.5 | 27.7 ± 3.6 | 25.9 ± 3.5 | 25.8 ± 3.4 | 0.627 | 0.769 |
| Visceral fat mass (m2) | 123.4 ± 41.6 | 124.6 ±44.2 | 97.0 ± 36.7 | 101.3 ± 39.9 | 0.142 | 0.390 |
| Body fat (%) | 31.6 ± 7.4 | 31.6 ± 7.5 | 25.4 ± 8.6 | 26.3 ± 9.0 | 0.299 | 0.307 |
| Sceletal muscle mas (kg) | 30.1 ± 5.4 | 30.1 ± 5.3 | 31.7 ± 73 | 31.1 ± 6.8 | 0.112 | 0.139 |
| Back extension (kg) | 46.77 ± 17.4 | 65.00 ± 20.8 * | 50.33 ± 22.4 | 49.11 ± 19.3 | 0.007 | 0.002 |
| Abdominal crunch (kg) | 30.73 ± 11.8 | 37.3 ± 11.9 * | 34.33 ± 8.1 | 37.00 ± 13.4 | 0.004 | 0.205 |
| Leg extension (kg) | 56.4 ± 21.9 | 65.5 ± 21.6 * | 51.7 ± 15.8 | 63.9 ± 18.1 * | ≤0.001 | 0.218 |
| Leg flexion (kg) | 33.7 ± 16.0 | 38.5 ± 16.0 * | 32.4 ± 15.5 | 35.2 ± 14.5 | 0.032 | 0.499 |
| Chest press (kg) | 50.9 ± 22.0 | 64.4 ± 23.1 * | 54.3 ± 17.2 | 65.3 ± 18.9 * | ≤0.001 | 0.555 |
| Seated row (kg) | 55.1 ± 19.2 | 66.5 ± 18.3 * | 62.8 ± 19.8 | 70.9 ± 24.7 * | ≤0.001 | 0.542 |
| TG | CG | Time Effect | Time × Group Interaction | |||
|---|---|---|---|---|---|---|
| T1 | T2 | T1 | T2 | p | p | |
| T cells (cells/μL) | ||||||
| CD4+ | 431.63 ± 629.0 | 819.88 ± 1049.5 | 437.10 ± 275.3 | 964.43 ± 962.4 | 0.088 | 0.989 |
| CD8+ | 443.88 ± 705.6 | 541.38 ± 855.2 | 410.29 ± 376.4 | 685.86 ± 528.4 | 0.394 | 0.463 |
| CD4+/CD8+ ratio | 1.15 ± 0.4 | 2.27 ± 1.5 * | 1.20 ± 0.5 | 1.44 ± 0.7 | 0.021 | 0.109 |
| T cell subsets (%) | ||||||
| CD4+ naive | 27.94 ± 6.5 | 26.52 ± 12.6 | 24.50 ± 9.2 | 25.31 ± 13.2 | 0.905 | 0.659 |
| CD4+ CM | 34.02 ± 7.0 | 38.15 ± 7.8 | 43.50 ± 10.7 | 39.92 ± 9.4 | 0.894 | 0.094 |
| CD4+ EM | 35.21 ± 11.2 | 33.74 ± 14.9 | 28.27 ± 6.7 | 30.97 ± 8.3 | 0.859 | 0.549 |
| CD4+ T-EMRA | 1.25 ± 0.7 | 1.52 ± 0.8 | 3.85 ± 3.1 | 3.84 ± 4.4 | 0.783 | 0.765 |
| CD4+ CD28−CD57+ | 2.04 ± 2.1 | 2.37 ± 2.9 | 6.00 ± 6.0 | 5.69 ± 5.7 | 0.982 | 0.636 |
| CD8+ naive | 6.25 ± 3.1 | 5.48 ± 3.6 | 5.82 ± 3.1 | 6.93 ± 4.0 | 0.362 | 0.201 |
| CD8+ CM | 9.30 ± 5.4 | 6.72 ± 5.3 | 8.64 ± 5.1 | 9.29 ± 4.1 | 0.645 | 0.441 |
| CD8+ EM | 37.59 ± 16.4 | 35.20 ± 10.3 | 39.89 ± 12.7 | 29.56 ± 11.3 * | 0.042 | 0.183 |
| CD8+ TEMRA | 46.90 ± 10.7 | 50.35 ± 15.3 | 48.35 ± 11.3 | 54.03 ± 9.3 | 0.143 | 0.715 |
| CD8+ CD28−CD57+ | 40.48 ± 13.2 | 41.44 ± 11.0 | 35.00 ± 13.7 | 44.16 ± 16.7 * | 0.046 | 0.096 |
| TG | CG | Time Effect | Time × Group Interaction | |||
|---|---|---|---|---|---|---|
| Molecules (pg/mL) | T1 | T2 | T1 | T2 | p | p |
| CCL-2 | 210.33 ± 91.4 | 190.20 ± 55.7 | 216.10 ± 83.6 | 212.37 ± 128.9 | 0.276 | 0.579 |
| CXCL-13 | 33.17 ± 11.9 | 28.89 ± 11.7 | 44.51 ± 29.5 | 44.89 ± 31.4 | 0.198 | 0.126 |
| ICAM-1 | 284,625.45 ± 135410.5 | 281,453.53 ± 112,499.9 | 354,343.68 ± 204,000.2 | 371,220.30 ± 168,159.4 | 0.610 | 0.601 |
| IL-1α | 3.96 ± 1.2 | 3.77 ± 1.1 | 3.69 ± 1.4 | 3.32 ± 1.0 | 0.376 | 0.646 |
| IL-1ra | 315.43 ± 189.2 | 280.59 ± 152.8 | 298.25 ± 124.3 | 266.30 ± 106.0 | 0.056 | 0.962 |
| IL-2 | 3.69 ± 2.0 | 1.98 ± 1.7 * | 2.80 ± 1.3 | 0.83 ± 0.7 * | 0.000 | 0.789 |
| IL-6 | 1.13 ± 0.3 | 0.98 ± 0.2 * | 1.17 ± 0.5 | 1.03 ± 0.4 | 0.008 | 0.862 |
| IL-8 | 3.05 ± 1.5 | 2.34 ± 0.7 * | 2.60 ± 0.8 | 2.45 ± 1.1 | 0.003 | 0.456 |
| IL-10 | 1.01 ± 0.4 | 0.87 ± 0.3 * | 1.07 ± 0.4 | 1.04 ± 0.2 | 0.024 | 0.486 |
| IL-18 | 179.71 ± 86.4 | 180.96 ± 80.5 | 216.97 ± 73.8 | 207.25 ± 60.1 | 0.640 | 0.544 |
| Leptin | 12,180.74 ± 11,654.7 | 14,804.82 ± 17,771.0 | 6095.97 ± 6350.4 | 6862.91 ± 10161.0 | 0.248 | 0.365 |
| Resistin | 7593.69 ± 2885.4 | 7634.37 ± 3263.1 | 7445.41 ± 1668.5 | 7770.8 ± 2068.7 | 0.477 | 0.580 |
| TNF-α | 2.55 ± 0.6 | 2.47 ± 0.5 | 2.49 ± 0.4 | 2.44 ± 0.5 | 0.423 | 0.646 |
| VEGF | 22.06 ± 16.1 | 14.99 ± 6.5 * | 15.49 ± 5.9 | 15.49 ± 7.0 | 0.021 | 0.232 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Despeghel, M.; Reichel, T.; Zander, J.; Krüger, K.; Weyh, C. Effects of a 6 Week Low-Dose Combined Resistance and Endurance Training on T Cells and Systemic Inflammation in the Elderly. Cells 2021, 10, 843. https://doi.org/10.3390/cells10040843
Despeghel M, Reichel T, Zander J, Krüger K, Weyh C. Effects of a 6 Week Low-Dose Combined Resistance and Endurance Training on T Cells and Systemic Inflammation in the Elderly. Cells. 2021; 10(4):843. https://doi.org/10.3390/cells10040843
Chicago/Turabian StyleDespeghel, Michael, Thomas Reichel, Johannes Zander, Karsten Krüger, and Christopher Weyh. 2021. "Effects of a 6 Week Low-Dose Combined Resistance and Endurance Training on T Cells and Systemic Inflammation in the Elderly" Cells 10, no. 4: 843. https://doi.org/10.3390/cells10040843
APA StyleDespeghel, M., Reichel, T., Zander, J., Krüger, K., & Weyh, C. (2021). Effects of a 6 Week Low-Dose Combined Resistance and Endurance Training on T Cells and Systemic Inflammation in the Elderly. Cells, 10(4), 843. https://doi.org/10.3390/cells10040843