
The Phosphoarginine Phosphatase PtpB from Staphylococcus aureus Is Involved in Bacterial Stress Adaptation during Infection

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Media, and Growth Conditions
2.2. Cloning and Expression of PtpA, PtpB and SecA Spot-Tagged Proteins in S. aureus
2.3. Cloning, Expression and Purification of Recombinant PtpB Proteins.
2.4. Construction of the S. aureus ptpB Deletion and Cis-Complementation Strains
2.5. Murine Abscess Model
2.6. Macrophage Culture and Infection
2.7. PtpB Phosphatase Activity Assay
2.8. H2O2 Susceptibility Assays
2.9. NO Susceptibility Assay
2.10. pH 5.5 Survival Assay
2.11. Measurement of Gene Expression by qRT-PCR
2.12. Statistical analyses
3. Results and Discussion
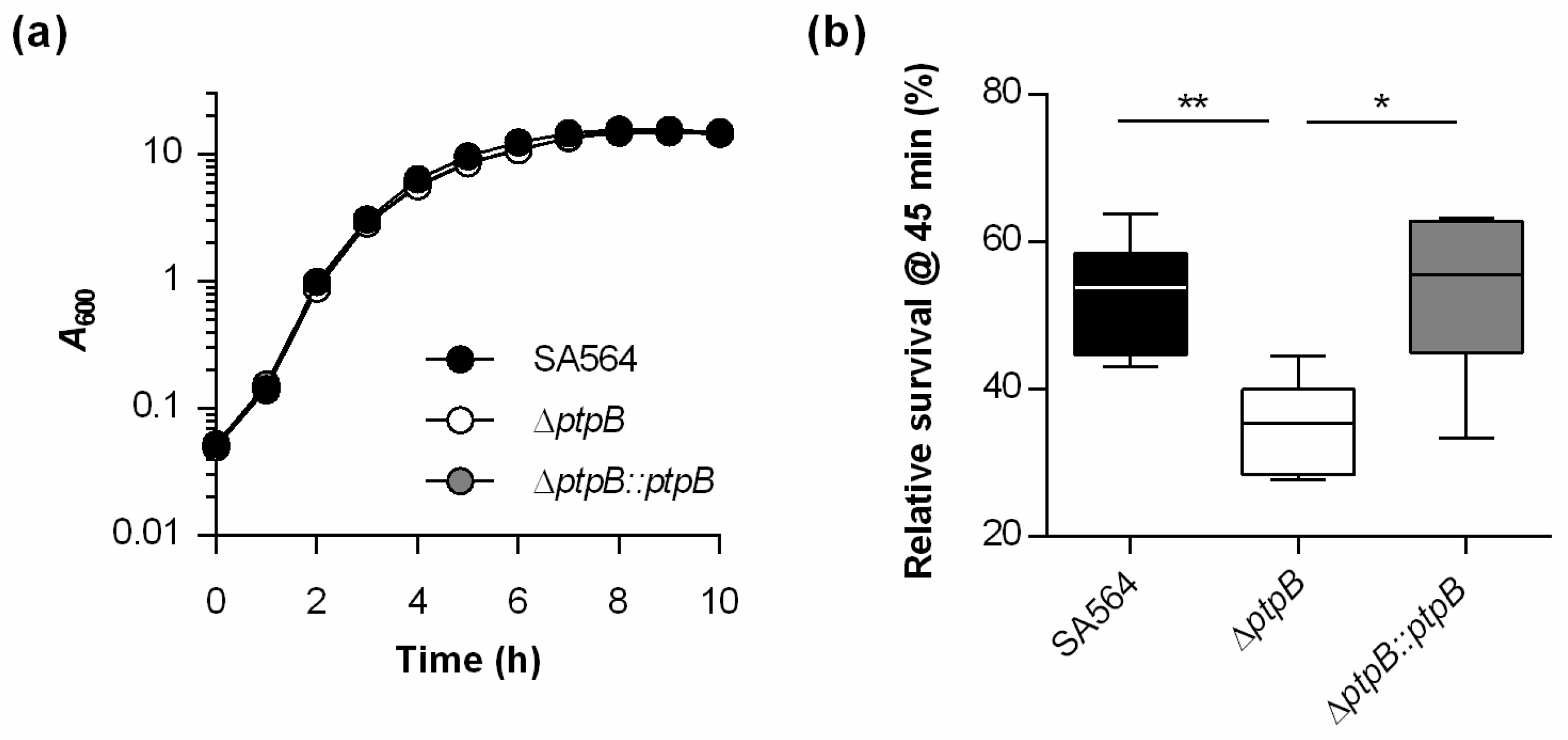
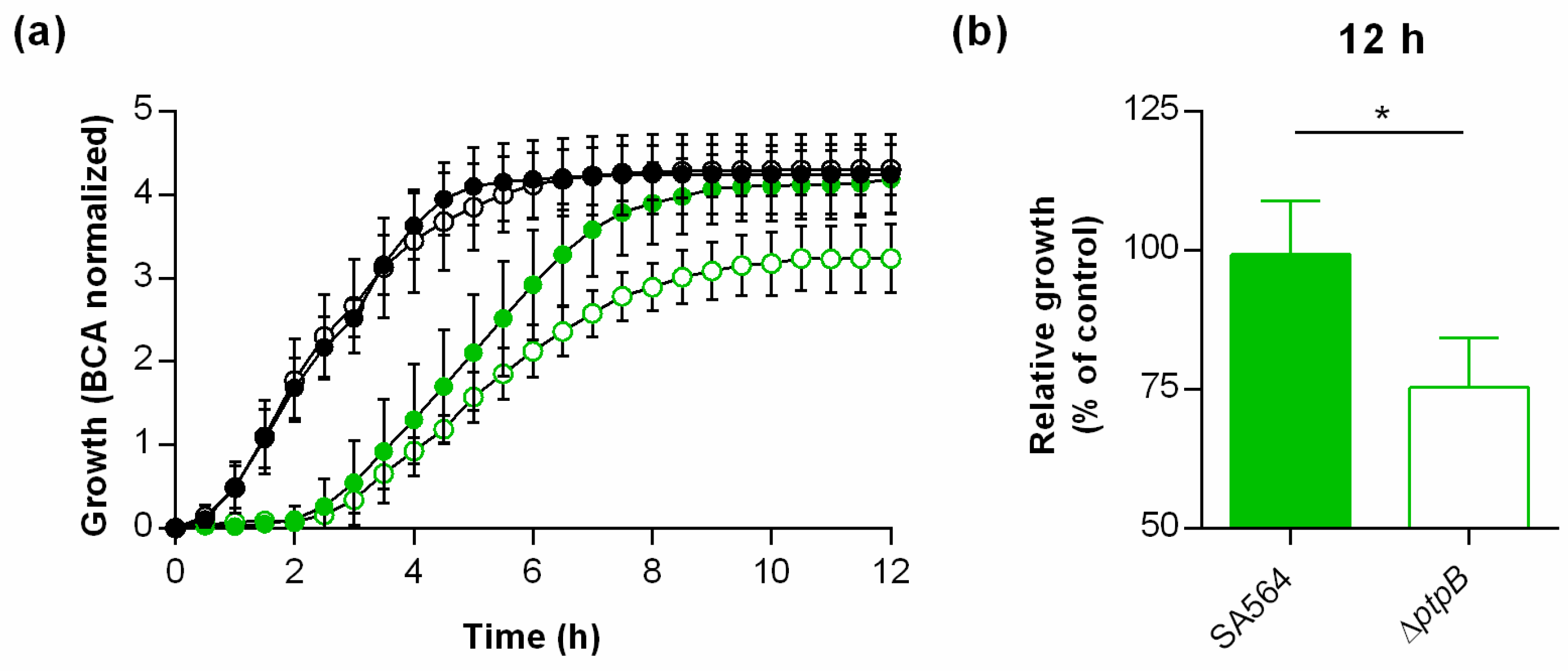
3.1. PtpB Does Not Affect the In Vitro Growth of S. aureus SA564 but Promotes Survival in Macrophages
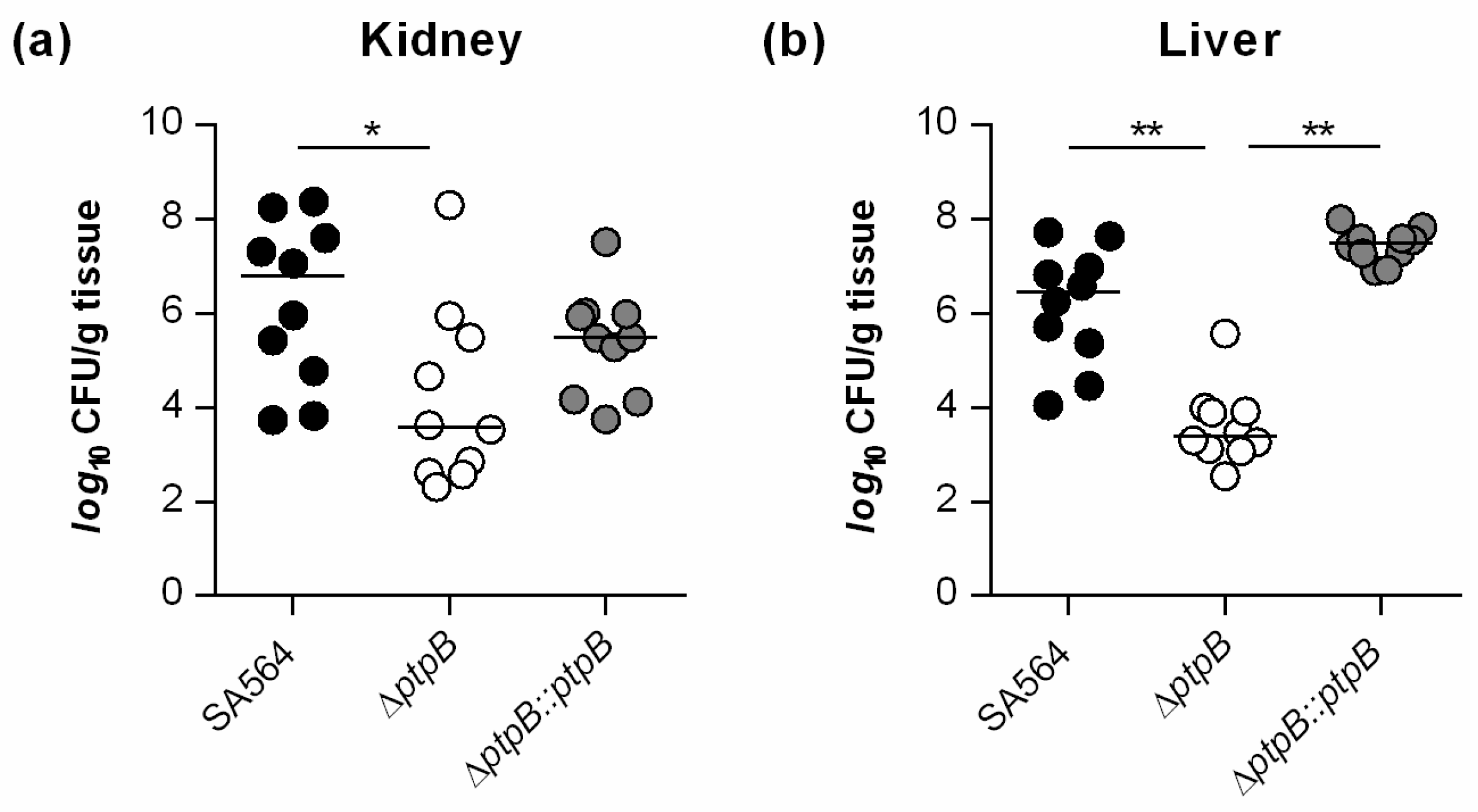
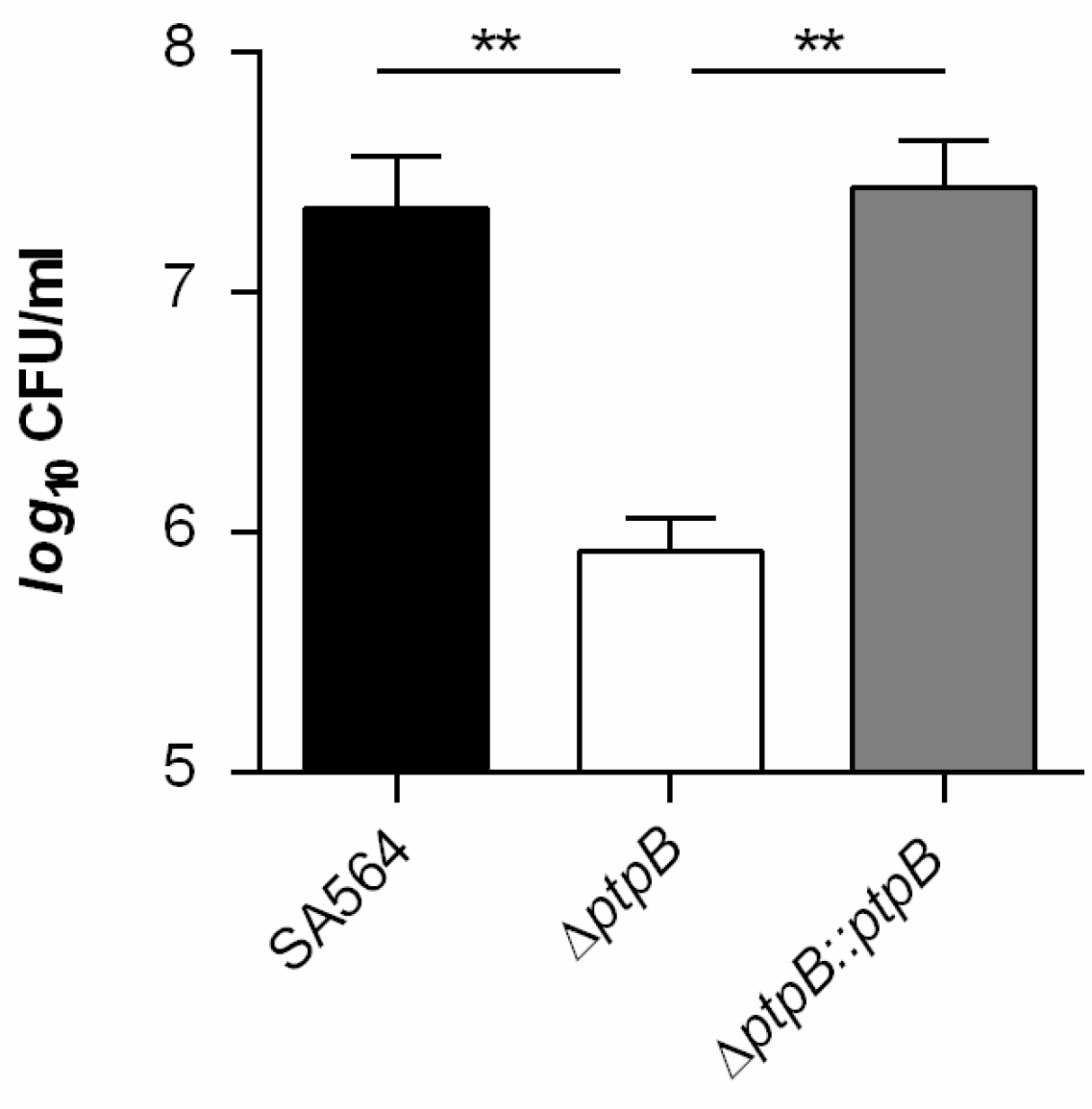
3.2. PtpB Contributes to Infectivity of S. aureus in a Murine Abscess Model
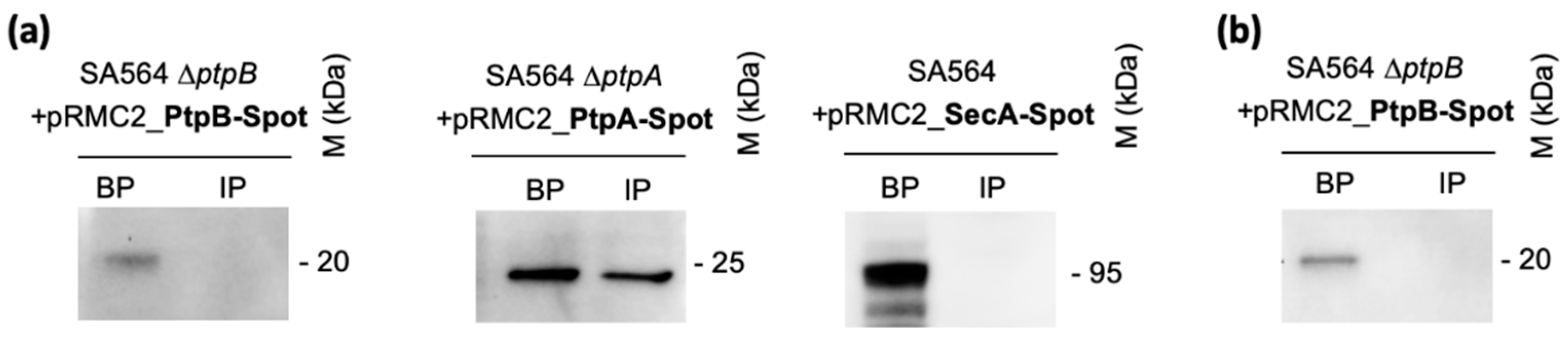
3.3. PtpB Is Not Secreted by S. aureus during In Vitro Growth and Upon Ingestion by Macrophages
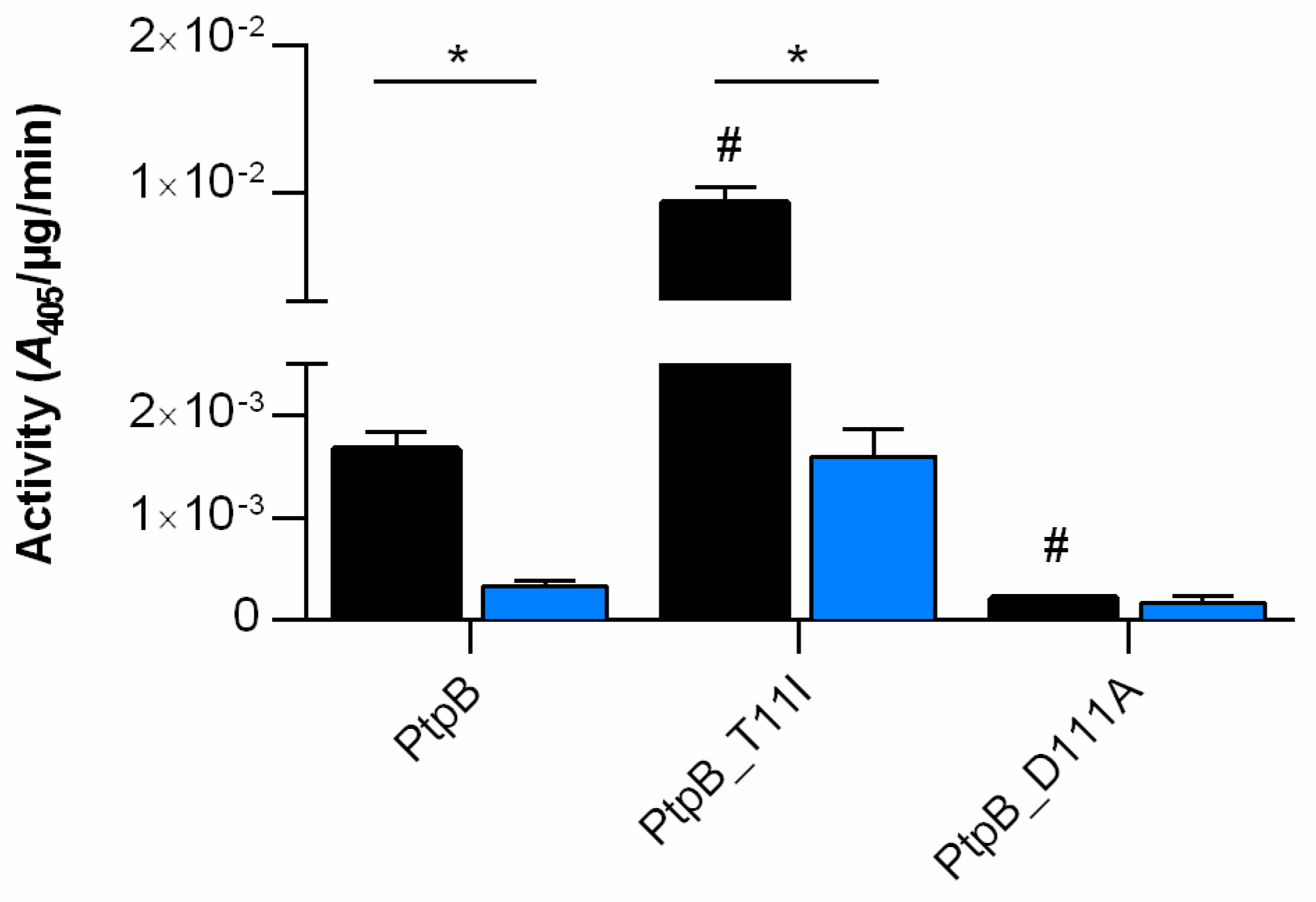
3.4. S. aureus PtpB Phosphatase Activity Depends on a Highly Specific Threonine Residue in Its Catalytic Loop
3.5. PtpB Affects the Capacity of S. aureus to Adapt to Oxidative Stress
3.6. PtpB Promotes the Capacity of S. aureus to Withstand NO˙ Stress.
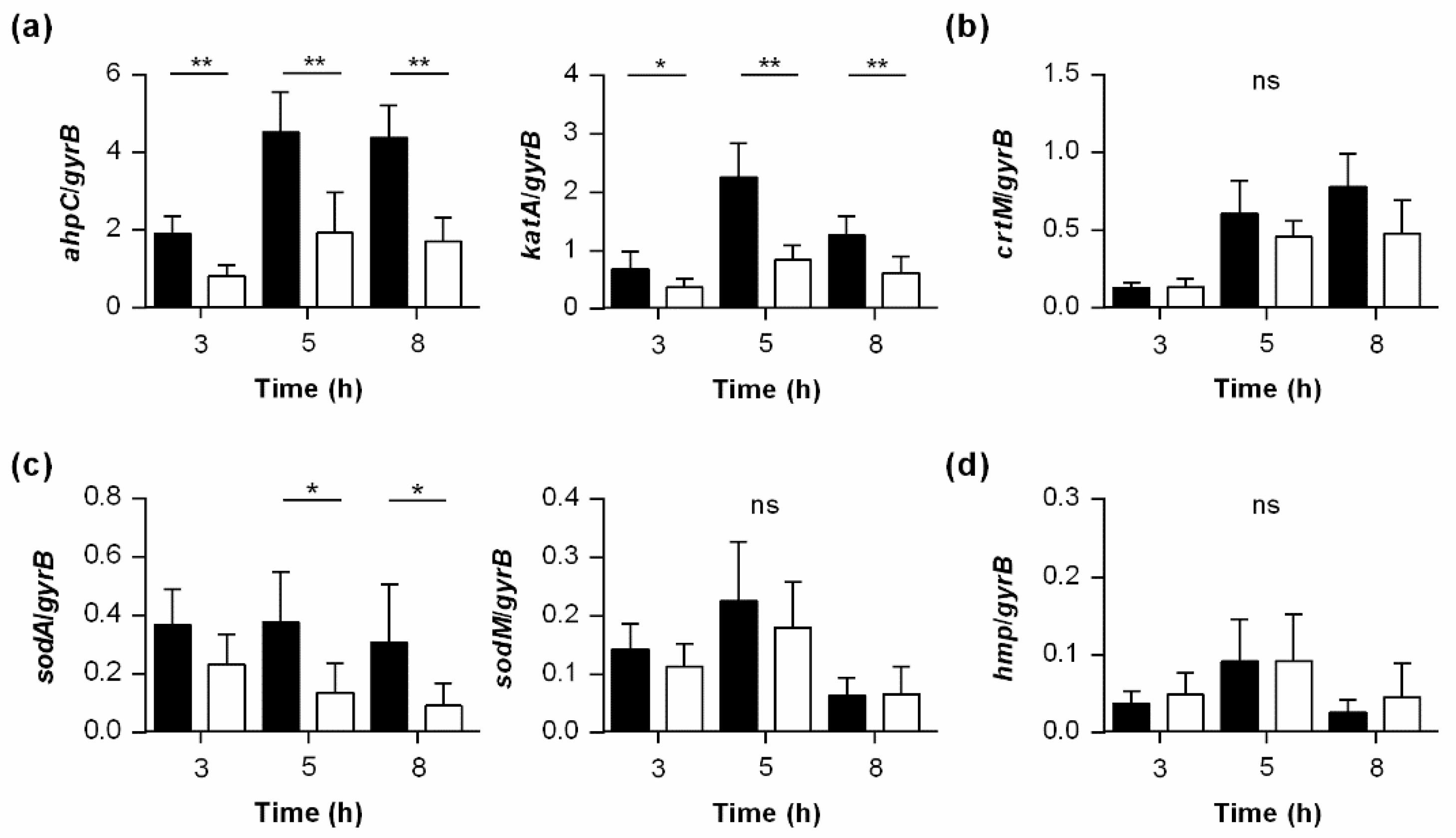
3.7. PtpB Alters the Transcription of Genes Encoding Factors Involved in the Detoxification of ROS
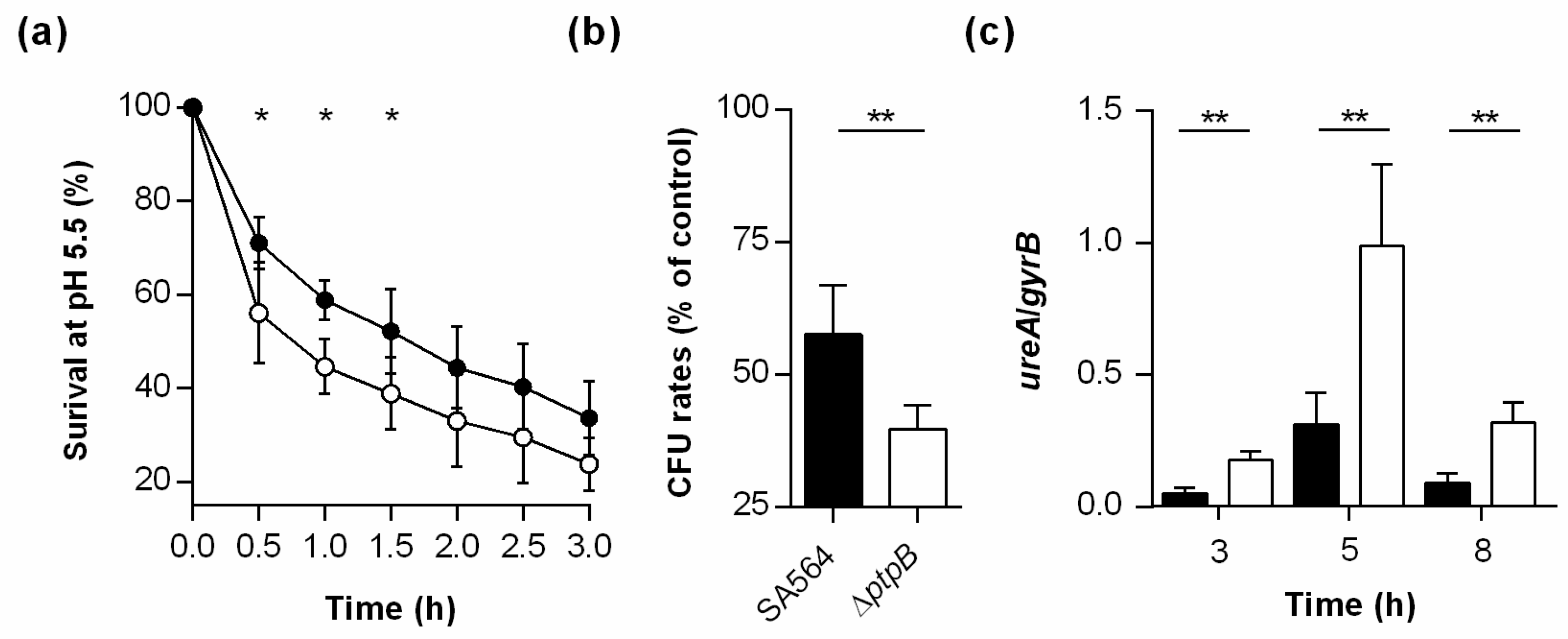
3.8. PtpB Contributes to the Survival Capacity of S. aureus Under Low pH.
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Gajdács, M. The Continuing Threat of Methicillin-Resistant Staphylococcus aureus. Antibiotics 2019, 8, 52. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, M.; Romby, P. Genetic Regulation. In Staphylococcus: Genetics and Physiology; Caister Academic Press: Poole, UK, 2016; pp. 301–334. [Google Scholar]
- Schlievert, P.M. Staphylococcal Virulence Factors. In Staphylococcus: Genetics and Physiology; Somervill, G.A., Ed.; Caister Academic Press: Lincoln, NE, USA, 2016; pp. 81–106. [Google Scholar]
- Fraunholz, M.; Sinha, B. Intracellular Staphylococcus aureus: Live-in and let die. Front. Cell. Infect. Microbiol. 2012, 2, 43. [Google Scholar] [CrossRef]
- Balasubramanian, D.; Harper, L.; Shopsin, B.; Torres, V.J. Staphylococcus aureus pathogenesis in diverse host environments. Pathog. Dis. 2017, 75. [Google Scholar] [CrossRef]
- Barã¡k, I. Complexity of bacterial phosphorylation interaction network. Front. Microbiol. 2014, 5, 725. [Google Scholar] [CrossRef]
- Hunter, T. Protein kinases and phosphatases: The Yin and Yang of protein phosphorylation and signaling. Cell 1995, 80, 225–236. [Google Scholar] [CrossRef]
- Pereira, S.F.F.; Goss, L.; Dworkin, J. Eukaryote-Like Serine/Threonine Kinases and Phosphatases in Bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 192–212. [Google Scholar] [CrossRef]
- Barford, D.; Das, A.K.; Egloff, M.-P. The Structure and Mechanism of Protein Phosphatases: Insights into Catalysis and Regulation. Annu. Rev. Biophys. Biomol. Struct. 1998, 27, 133–164. [Google Scholar] [CrossRef] [PubMed]
- Perego, M.; Hanstein, C.; Welsh, K.M.; Djavakhishvili, T.; Glaser, P.; Hoch, J.A. Multiple protein-aspartate phosphatases provide a mechanism for the integration of diverse signals in the control of development in B. subtilis. Cell 1994, 79, 1047–1055. [Google Scholar] [CrossRef]
- Rigden, D.J. The histidine phosphatase superfamily: Structure and function. Biochem. J. 2007, 409, 333–348. [Google Scholar] [CrossRef] [PubMed]
- Klumpp, S.; Krieglstein, J. Reversible Phosphorylation of Histidine Residues in Proteins from Vertebrates. Sci. Signal. 2009, 2, pe13. [Google Scholar] [CrossRef]
- Sun, F.; Ding, Y.; Ji, Q.; Liang, Z.; Deng, X.; Wong, C.C.L.; Yi, C.; Zhang, L.; Xie, S.; Alvarez, S.; et al. Protein cysteine phosphorylation of SarA/MgrA family transcriptional regulators mediates bacterial virulence and antibiotic resistance. Proc. Natl. Acad. Sci. USA 2012, 109, 15461–15466. [Google Scholar] [CrossRef]
- Fuhrmann, J.; Mierzwa, B.; Trentini, D.B.; Spiess, S.; Lehner, A.; Charpentier, E.; Clausen, T. Structural Basis for Recognizing Phosphoarginine and Evolving Residue-Specific Protein Phosphatases in Gram-Positive Bacteria. Cell Rep. 2013, 3, 1832–1839. [Google Scholar] [CrossRef] [PubMed]
- Fuhrmann, J.; Schmidt, A.; Spiess, S.; Lehner, A.; Turgay, K.; Mechtler, K.; Charpentier, E.; Clausen, T. McsB Is a Protein Arginine Kinase That Phosphorylates and Inhibits the Heat-Shock Regulator CtsR. Science 2009, 324, 1323–1327. [Google Scholar] [CrossRef]
- Elsholz, A.K.W.; Turgay, K.; Michalik, S.; Hessling, B.; Gronau, K.; Oertel, D.; Mäder, U.; Bernhardt, J.; Becher, D.; Hecker, M.; et al. Global impact of protein arginine phosphorylation on the physiology of Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2012, 109, 7451–7456. [Google Scholar] [CrossRef]
- Sajid, A.; Arora, G.; Singhal, A.; Kalia, V.C.; Singh, Y. Protein Phosphatases of Pathogenic Bacteria: Role in Physiology and Virulence. Annu. Rev. Microbiol. 2015, 69, 527–547. [Google Scholar] [CrossRef]
- Whitmore, S.E.; Lamont, R.J. Tyrosine phosphorylation and bacterial virulence. Int. J. Oral Sci. 2012, 4, 1–6. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Flannagan, R.S.; Heit, B.; Heinrichs, D.E. Antimicrobial Mechanisms of Macrophages and the Immune Evasion Strategies of Staphylococcus aureus. Pathogens 2015, 4, 826–868. [Google Scholar] [CrossRef] [PubMed]
- Soulat, D.; Vaganay, E.; Duclos, B.; Genestier, A.-L.; Etienne, J.; Cozzone, A.J. Staphylococcus aureus Contains Two Low-Molecular-Mass Phosphotyrosine Protein Phosphatases. J. Bacteriol. 2002, 184, 5194–5199. [Google Scholar] [CrossRef]
- Everitt, R.G.; Didelot, X.; Batty, E.M.; Miller, R.R.; Knox, K.; Young, B.C.; Bowden, R.; Auton, A.; Votintseva, A.; Larner-Svensson, H.; et al. Mobile elements drive recombination hotspots in the core genome of Staphylococcus aureus. Nat. Commun. 2014, 5, 3956. [Google Scholar] [CrossRef]
- Brelle, S.; Baronian, G.; Huc-Brandt, S.; Zaki, L.G.; Cohen-Gonsaud, M.; Bischoff, M.; Molle, V. Phosphorylation-mediated regulation of the Staphylococcus aureus secreted tyrosine phosphatase PtpA. Biochem. Biophys. Res. Commun. 2016, 469, 619–625. [Google Scholar] [CrossRef]
- Gannoun-Zaki, L.; Pätzold, L.; Huc-Brandt, S.; Baronian, G.; Elhawy, M.I.; Gaupp, R.; Martin, M.; Blanc-Potard, A.-B.; Letourneur, F.; Bischoff, M.; et al. PtpA, a secreted tyrosine phosphatase from Staphylococcus aureus, contributes to virulence and interacts with coronin-1A during infection. J. Biol. Chem. 2018, 293, 15569–15580. [Google Scholar] [CrossRef] [PubMed]
- Junker, S.; Maaβ, S.; Otto, A.; Michalik, S.; Morgenroth, F.; Gerth, U.; Hecker, M.; Becher, D. Spectral Library Based Analysis of Arginine Phosphorylations in Staphylococcus aureus. Mol. Cell. Proteom. 2018, 17, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Junker, S.; Maaß, S.; Otto, A.; Hecker, M.; Becher, D. Toward the Quantitative Characterization of Arginine Phosphorylations in Staphylococcus aureus. J. Proteome Res. 2018, 18, 265–279. [Google Scholar] [CrossRef]
- Somerville, G.A.; Beres, S.B.; Fitzgerald, J.R.; DeLeo, F.R.; Cole, R.L.; Hoff, J.S.; Musser, J.M. In Vitro Serial Passage of Staphylococcus aureus: Changes in Physiology, Virulence Factor Production, and agr Nucleotide Sequence. J. Bacteriol. 2002, 184, 1430–1437. [Google Scholar] [CrossRef] [PubMed]
- Monk, I.R.; Tree, J.J.; Howden, B.P.; Stinear, T.P.; Foster, T.J. Complete Bypass of Restriction Systems for Major Staphylococcus aureus Lineages. mBio 2015, 6, e00308-15. [Google Scholar] [CrossRef] [PubMed]
- Geiger, T.; Francois, P.; Liebeke, M.; Fraunholz, M.; Goerke, C.; Krismer, B.; Schrenzel, J.; Lalk, M.; Wolz, C. The Stringent Response of Staphylococcus aureus and Its Impact on Survival after Phagocytosis through the Induction of Intracellular PSMs Expression. PLOS Pathog. 2012, 8, e1003016. [Google Scholar] [CrossRef] [PubMed]
- Giachino, P.; Engelmann, S.; Bischoff, M. Sigma(B) Activity Depends on RsbU in Staphylococcus aureus. J. Bacteriol. 2001, 183, 1843–1852. [Google Scholar] [CrossRef]
- Corrigan, R.M.; Foster, T.J. An improved tetracycline-inducible expression vector for Staphylococcus aureus. Plasmid 2009, 61, 126–129. [Google Scholar] [CrossRef]
- Terpe, K. Overview of tag protein fusions: From molecular and biochemical fundamentals to commercial systems. Appl. Microbiol. Biotechnol. 2003, 60, 523–533. [Google Scholar] [CrossRef]
- Bae, T.; Schneewind, O. Allelic replacement in Staphylococcus aureus with inducible counter-selection. Plasmid 2006, 55, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Gaupp, R.; Wirf, J.; Wonnenberg, B.; Biegel, T.; Eisenbeis, J.; Graham, J.; Herrmann, M.; Lee, C.Y.; Beisswenger, C.; Wolz, C.; et al. RpiRc Is a Pleiotropic Effector of Virulence Determinant Synthesis and Attenuates Pathogenicity in Staphylococcus aureus. Infect. Immun. 2016, 84, 2031–2041. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, S.; Yamabe, M.; Yamaguchi, Y.; Kobayashi, Y.; Konno, T.; Tada, K. Establishment and characterization of a human acute monocytic leukemia cell line (THP-1). Int. J. Cancer 1980, 26, 171–176. [Google Scholar] [CrossRef]
- Stokes, R.W.; Doxsee, D. The Receptor-Mediated Uptake, Survival, Replication, and Drug Sensitivity of Mycobacterium tuberculosis within the Macrophage-like Cell Line THP-1: A Comparison with Human Monocyte-Derived Macrophages. Cell. Immunol. 1999, 197, 1–9. [Google Scholar] [CrossRef]
- Weinstein, M.P.; Patel, J.B.; Burnham, C.-A.; Campeau, S.; Conville, P.S.; Doarn, C.; Eliopoulos, G.M.; Galas, M.F.; Humphries, R.M.; Jenkins, S.G.; et al. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically in CLSI Standard M07. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Chatterjee, I.; Becker, P.; Grundmeier, M.; Bischoff, M.; Somerville, G.A.; Peters, G.; Sinha, B.; Harraghy, N.; Proctor, R.A.; Herrmann, M. Staphylococcus aureus ClpC Is Required for Stress Resistance, Aconitase Activity, Growth Recovery, and Death. J. Bacteriol. 2005, 187, 4488–4496. [Google Scholar] [CrossRef]
- Kubica, M.; Guzik, K.; Koziel, J.; Zarebski, M.; Richter, W.; Gajkowska, B.; Golda, A.; Maciag-Gudowska, A.; Brix, K.; Shaw, L.; et al. A Potential New Pathway for Staphylococcus aureus Dissemination: The Silent Survival of S. aureus Phagocytosed by Human Monocyte-Derived Macrophages. PLoS ONE 2008, 3, e1409. [Google Scholar] [CrossRef]
- Wong, D.; Chao, J.D.; Av-Gay, Y. Mycobacterium tuberculosis-secreted phosphatases: From pathogenesis to targets for TB drug development. Trends Microbiol. 2013, 21, 100–109. [Google Scholar] [CrossRef]
- Vega, C.; Chou, S.; Engel, K.; Harrell, M.E.; Rajagopal, L.; Grundner, C. Structure and Substrate Recognition of the Staphylococcus aureus Protein Tyrosine Phosphatase PtpA. J. Mol. Biol. 2011, 413, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Sun, F.; Cho, H.; Yelavarthi, V.; Sohn, C.; He, C.; Schneewind, O.; Bae, T. CcpA Mediates Proline Auxotrophy and Is Required for Staphylococcus aureus Pathogenesis. J. Bacteriol. 2010, 192, 3883–3892. [Google Scholar] [CrossRef]
- Favazzo, L.J.; Gill, A.L.; Farnsworth, C.W.; Mooney, R.A.; Gill, S.R. The Response of nor and nos Contributes to Staphylococcus aureus Virulence and Metabolism. J. Bacteriol. 2019, 201, e00107-19. [Google Scholar] [CrossRef]
- Xu, S.X.; Gilmore, K.J.; Szabo, P.A.; Zeppa, J.J.; Baroja, M.L.; Haeryfar, S.M.M.; McCormick, J.K. Superantigens Subvert the Neutrophil Response To Promote Abscess Formation and Enhance Staphylococcus aureus Survival In Vivo. Infect. Immun. 2014, 82, 3588–3598. [Google Scholar] [CrossRef]
- Selbach, M.; Paul, F.E.; Brandt, S.; Guye, P.; Daumke, O.; Backert, S.; Dehio, C.; Mann, M. Host Cell Interactome of Tyrosine-Phosphorylated Bacterial Proteins. Cell Host Microbe 2009, 5, 397–403. [Google Scholar] [CrossRef]
- Canova, M.J.; Molle, V. Bacterial Serine/Threonine Protein Kinases in Host-Pathogen Interactions. J. Biol. Chem. 2014, 289, 9473–9479. [Google Scholar] [CrossRef] [PubMed]
- Popa, C.M.; Tabuchi, M.; Valls, M. Modification of Bacterial Effector Proteins Inside Eukaryotic Host Cells. Front. Cell. Infect. Microbiol. 2016, 6, 73. [Google Scholar] [CrossRef]
- Busche, T.; Hillion, M.; Van Loi, V.; Berg, D.; Walther, B.; Semmler, T.; Strommenger, B.; Witte, W.; Cuny, C.; Mellmann, A.; et al. Comparative Secretome Analyses of Human and Zoonotic Staphylococcus aureus Isolates CC8, CC22, and CC398. Mol. Cell. Proteom. 2018, 17, 2412–2433. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-K.; Ewis, H.E.; Zhang, X.; Lu, C.-D.; Hu, H.-J.; Pan, Y.; Abdelal, A.T.; Tai, P.C. Nonclassical Protein Secretion by Bacillus subtilis in the Stationary Phase Is Not Due to Cell Lysis. J. Bacteriol. 2011, 193, 5607–5615. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Dhar, R.; Das, A.K. Analyzing the catalytic mechanism of protein tyrosine phosphatase PtpB from Staphylococcus aureus through site-directed mutagenesis. Int. J. Biol. Macromol. 2009, 45, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Denu, J.M.; Dixon, J.E. Protein tyrosine phosphatases: Mechanisms of catalysis and regulation. Curr. Opin. Chem. Biol. 1998, 2, 633–641. [Google Scholar] [CrossRef]
- Zhang, Z.-Y. Mechanistic Studies on Protein Tyrosine Phosphatases. Prog. Nucleic Acid. Res. Mol. Biol. 2003, 73, 171–220. [Google Scholar] [CrossRef]
- Peters, G.H.; Frimurer, T.M.; Olsen, O.H. Electrostatic Evaluation of the Signature Motif (H/V)CX5R(S/T) in Protein−Tyrosine Phosphatases. Biochemistry 1998, 37, 5383–5393. [Google Scholar] [CrossRef]
- Grundner, C.; Ng, H.-L.; Alber, T. Mycobacterium tuberculosis Protein Tyrosine Phosphatase PtpB Structure Reveals a Diverged Fold and a Buried Active Site. Structure 2005, 13, 1625–1634. [Google Scholar] [CrossRef] [PubMed]
- Fuhrmann, J.; Subramanian, V.; Thompson, P.R. Targeting the Arginine Phosphatase YwlE with a Catalytic Redox-Based Inhibitor. ACS Chem. Biol. 2013, 8, 2024–2032. [Google Scholar] [CrossRef] [PubMed]
- Fuhrmann, J.; Subramanian, V.; Kojetin, D.J.; Thompson, P.R. Activity-Based Profiling Reveals a Regulatory Link between Oxidative Stress and Protein Arginine Phosphorylation. Cell Chem. Biol. 2016, 23, 967–977. [Google Scholar] [CrossRef]
- Abuaita, B.H.; Schultz, T.L.; O’Riordan, M.X. Mitochondria-Derived Vesicles Deliver Antimicrobial Reactive Oxygen Species to Control Phagosome-Localized Staphylococcus aureus. Cell Host Microbe 2018, 24, 625–636. [Google Scholar] [CrossRef]
- Gaupp, R.; Ledala, N.; Somerville, G.A. Staphylococcal Response to Oxidative Stress. Front. Cell Infect. Microbiol. 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, K.; Coutts, G.; Jonsson, I.-M.; Tarkowski, A.; Kokai-Kun, J.F.; Mond, J.J.; Foster, S.J. Catalase (KatA) and Alkyl Hydroperoxide Reductase (AhpC) Have Compensatory Roles in Peroxide Stress Resistance and Are Required for Survival, Persistence, and Nasal Colonization in Staphylococcus aureus. J. Bacteriol. 2006, 189, 1025–1035. [Google Scholar] [CrossRef]
- Clauditz, A.; Resch, A.; Wieland, K.-P.; Peschel, A.; Götz, F. Staphyloxanthin Plays a Role in the Fitness of Staphylococcus aureus and Its Ability To Cope with Oxidative Stress. Infect. Immun. 2006, 74, 4950–4953. [Google Scholar] [CrossRef]
- Pelz, A.; Wieland, K.-P.; Putzbach, K.; Hentschel, P.; Albert, K.; Götz, F. Structure and Biosynthesis of Staphyloxanthin from Staphylococcus aureus. J. Biol. Chem. 2005, 280, 32493–32498. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.Y.; Essex, A.; Buchanan, J.T.; Datta, V.; Hoffman, H.M.; Bastian, J.F.; Fierer, J.; Nizet, V. Staphylococcus aureus golden pigment impairs neutrophil killing and promotes virulence through its antioxidant activity. J. Exp. Med. 2005, 202, 209–215. [Google Scholar] [CrossRef]
- Olivier, A.C.; Lemaire, S.; Van Bambeke, F.; Tulkens, P.M.; Oldfield, E. Role of RsbU and Staphyloxanthin in Phagocytosis and Intracellular Growth of Staphylococcus aureus in Human Macrophages and Endothelial Cells. J. Infect. Dis. 2019, 200, 1367–1370. [Google Scholar] [CrossRef] [PubMed]
- Clements, M.O.; Watson, S.P.; Foster, S.J. Characterization of the Major Superoxide Dismutase of Staphylococcus aureus and Its Role in Starvation Survival, Stress Resistance, and Pathogenicity. J. Bacteriol. 1999, 181, 3898–3903. [Google Scholar] [CrossRef] [PubMed]
- Valderas, M.W.; Hart, M.E. Identification and Characterization of a Second Superoxide Dismutase Gene (sodM) from Staphylococcus aureus. J. Bacteriol. 2001, 183, 3399–3407. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.R.; Dunman, P.M.; Fang, F.C. The nitrosative stress response of Staphylococcus aureus is required for resistance to innate immunity. Mol. Microbiol. 2006, 61, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Flannagan, R.S.; Kuiack, R.C.; McGavin, M.J.; Heinrichs, D.E. Staphylococcus aureus Uses the GraXRS Regulatory System To Sense and Adapt to the Acidified Phagolysosome in Macrophages. mBio 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Bhinderwala, F.; Lehman, M.K.; Thomas, V.C.; Chaudhari, S.S.; Yamada, K.J.; Foster, K.W.; Powers, R.; Kielian, T.; Fey, P.D. Urease is an essential component of the acid response network of Staphylococcus aureus and is required for a persistent murine kidney infection. PLOS Pathog. 2019, 15, e1007538. [Google Scholar] [CrossRef]
- Boortz, C. YwlE and the Regulation of the Oxidative Stress Response in Bacillus anthracis. In Ph.D. Thesis; Texas Christian University: Fort Worth, TX, USA, 2016. [Google Scholar]
- Moldovan, A.; Fraunholz, M.J. In or out: Phagosomal escape of Staphylococcus aureus. Cell. Microbiol. 2019, 21, e12997. [Google Scholar] [CrossRef] [PubMed]
- Russell, D.G.; VanderVen, B.C.; Glennie, S.; Mwandumba, H.; Heyderman, R.S. The macrophage marches on its phagosome: Dynamic assays of phagosome function. Nat. Rev. Immunol. 2009, 9, 594–600. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Description 1 | Reference or Source |
|---|---|---|
| S. aureus | ||
| SA564 | S. aureus clinical isolate, wild type | [27] |
| SA564 ΔptpB | SA564 ΔptpB::lox66-erm(B)-lox71; ErmR | This study |
| SA564 ΔptpB::ptpB | cis-complemented SA564 ΔptpB derivative | This study |
| E. coli | ||
| BL21(DE3)Star | E. coli strain allowing a high-level recombinant protein expression. IPTG-inducible T7 RNA polymerase | Invitrogen |
| IM08B | E. coli DC10B derivative harboring hsdS of S. aureus strain NRS384, Δdcm | [28] |
| TOP10 | E. coli derivative ultra-competent cells used for general cloning | Invitrogen |
| Plasmids | ||
| pBASE6 | E. coli–S. aureus temperature-sensitive suicide shuttle vector, secY counterselection; bla cat | [29] |
| pBASE6 ptpB comp | pBASE6 derivative harboring the C-terminal region of NWMN_2020, ptpB, and the N-terminal region of glyA; bla, cat | This study |
| pBT | S. aureus suicide plasmid; tet(L) | [30] |
| pBT ptpB KO | pBT derivative harboring the genomic regions flanking ptpB and lox66-erm(B)-lox71; tet(L), erm(B) | This study |
| pET19b | E. coli vector for IPTG inducible protein expression; bla | Novagen |
| pET19b_PtpB | pET19b derivative used to express HAT-tagged fusion of S. aureus PtpB WT in E. coli; bla | This study |
| pET19b_PtpB_D111A | pET19b derivative used to express HAT-tagged fusion of S. aureus PtpB_D111A in E. coli; bla | This study |
| pET19b_PtpB_T11I | pET19b derivative used to express HAT-tagged fusion of S. aureus PtpB_T11I in E. coli; bla | This study |
| pRMC2 | E. coli–S. aureus shuttle vector, Tetracycline-inducible expression; bla, tet | [31] |
| pRMC2_PtpA-Spot | pRMC2 derivative used to express C-terminal Spot-tagged fusion of S. aureus PtpA; bla, tet | This study |
| pRMC2_PtpB-Spot | pRMC2 derivative used to express C-terminal Spot-tagged fusion of S. aureus PtpB; bla, tet | This study |
| pRMC2_SecA-Spot | pRMC2 derivative used to express C-terminal Spot-tagged fusion of S. aureus SecA; bla, tet | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elhawy, M.I.; Huc-Brandt, S.; Pätzold, L.; Gannoun-Zaki, L.; Abdrabou, A.M.M.; Bischoff, M.; Molle, V. The Phosphoarginine Phosphatase PtpB from Staphylococcus aureus Is Involved in Bacterial Stress Adaptation during Infection. Cells 2021, 10, 645. https://doi.org/10.3390/cells10030645
Elhawy MI, Huc-Brandt S, Pätzold L, Gannoun-Zaki L, Abdrabou AMM, Bischoff M, Molle V. The Phosphoarginine Phosphatase PtpB from Staphylococcus aureus Is Involved in Bacterial Stress Adaptation during Infection. Cells. 2021; 10(3):645. https://doi.org/10.3390/cells10030645
Chicago/Turabian StyleElhawy, Mohamed Ibrahem, Sylvaine Huc-Brandt, Linda Pätzold, Laila Gannoun-Zaki, Ahmed Mohamed Mostafa Abdrabou, Markus Bischoff, and Virginie Molle. 2021. "The Phosphoarginine Phosphatase PtpB from Staphylococcus aureus Is Involved in Bacterial Stress Adaptation during Infection" Cells 10, no. 3: 645. https://doi.org/10.3390/cells10030645
APA StyleElhawy, M. I., Huc-Brandt, S., Pätzold, L., Gannoun-Zaki, L., Abdrabou, A. M. M., Bischoff, M., & Molle, V. (2021). The Phosphoarginine Phosphatase PtpB from Staphylococcus aureus Is Involved in Bacterial Stress Adaptation during Infection. Cells, 10(3), 645. https://doi.org/10.3390/cells10030645