
Stroma-Mediated Resistance to S63845 and Venetoclax through MCL-1 and BCL-2 Expression Changes Induced by miR-193b-3p and miR-21-5p Dysregulation in Multiple Myeloma

 , , , ,
, , , ,  ,
,  , ,
, ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
3. Results
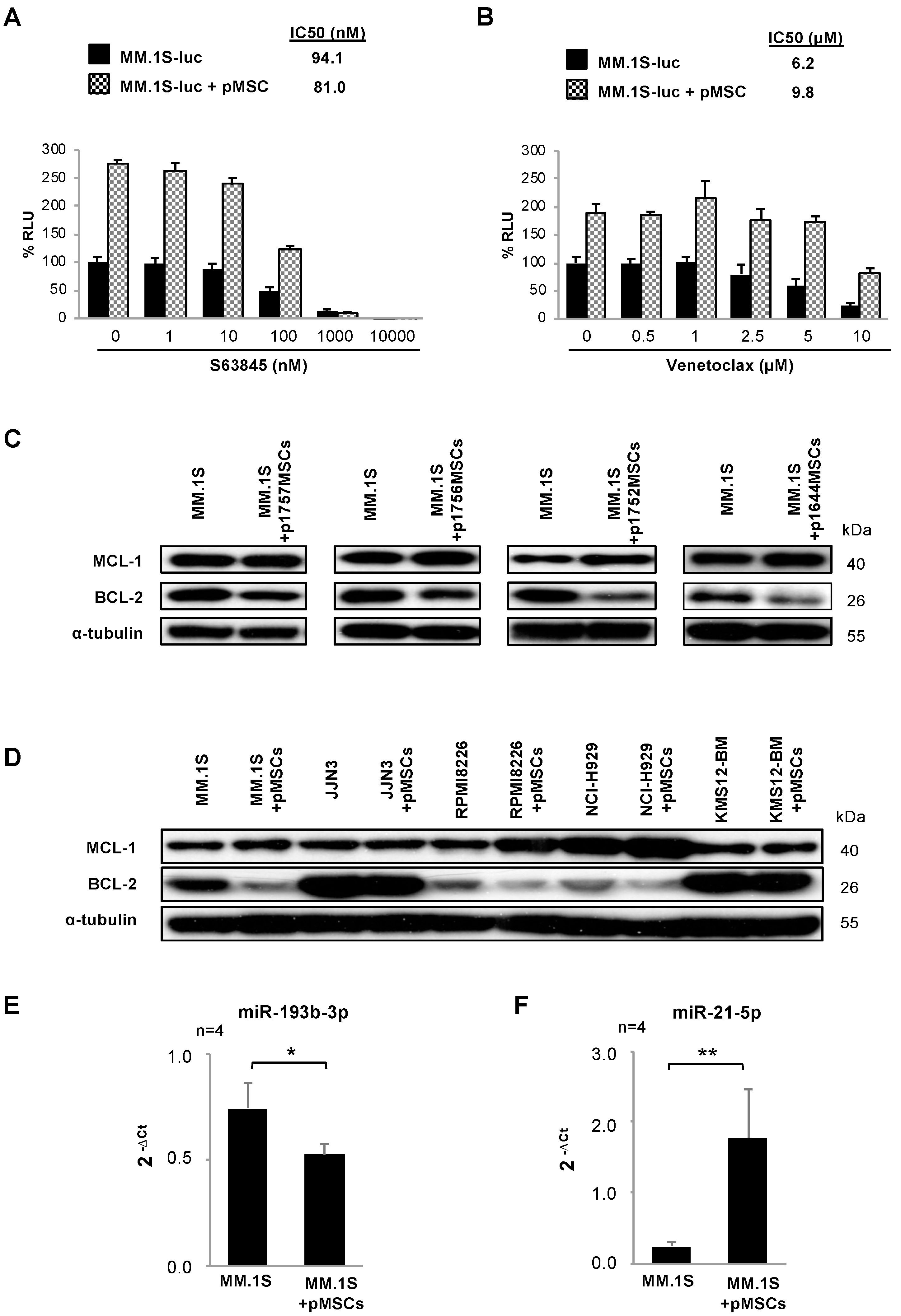
3.1. Co-Culture of MM.1S Cells with pMSCs Alters the Cytotoxic Effect of S63845 and Venetoclax in Monotherapy
3.2. pMSCs Modify the Expression of MCL-1 and BCL-2 in MM Cells
3.3. The BM Stromal Microenvironment Deregulates the Expression of miRNAs Potentially Modulating MCL-1 or BCL-2 Levels in MM.1S Cells
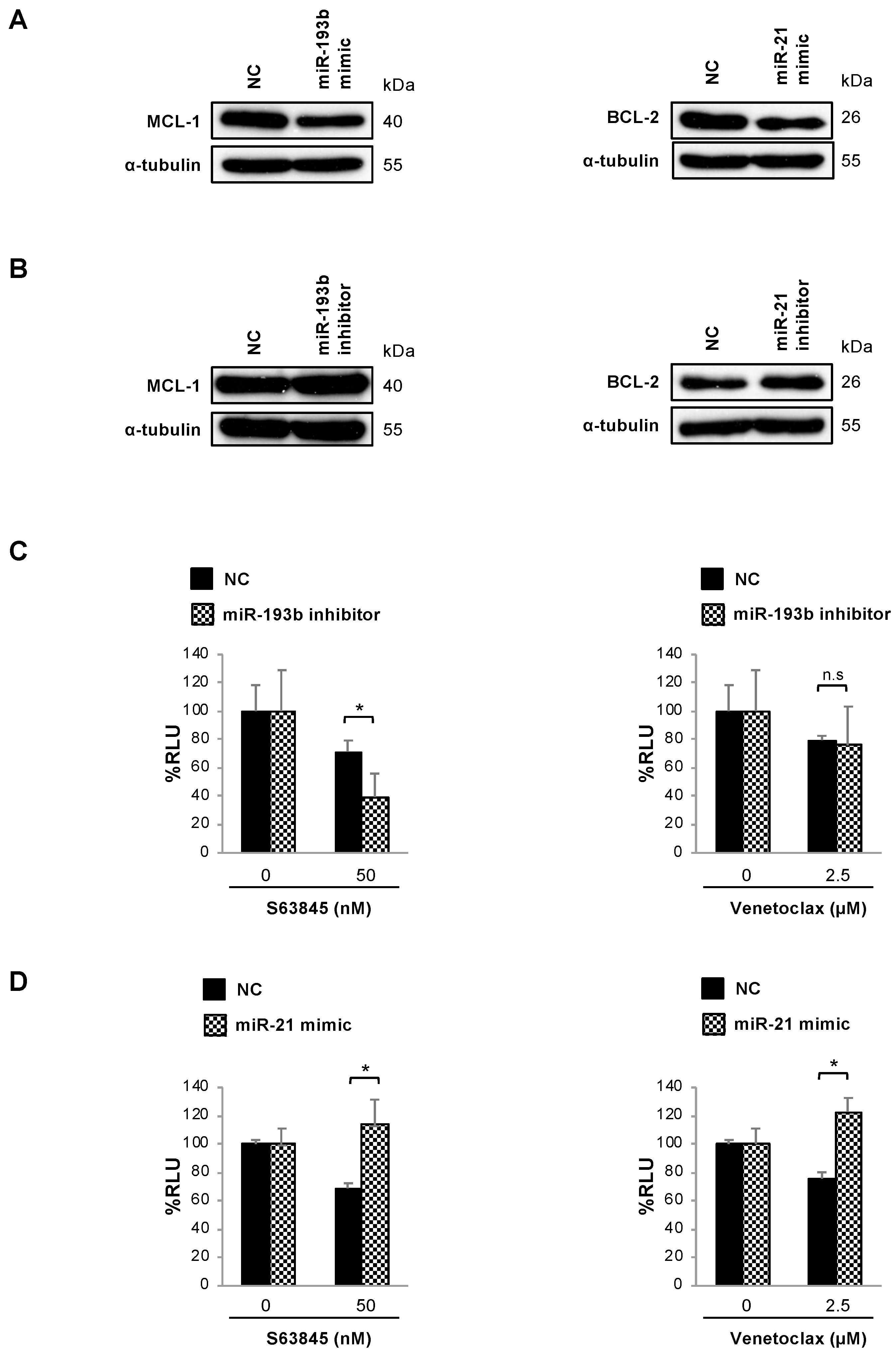
3.4. Changes in MCL-1 and BCL-2 Protein Expression Induced by pMSCs in MM Cells Are Partially Mediated by miR-193 and miR-21
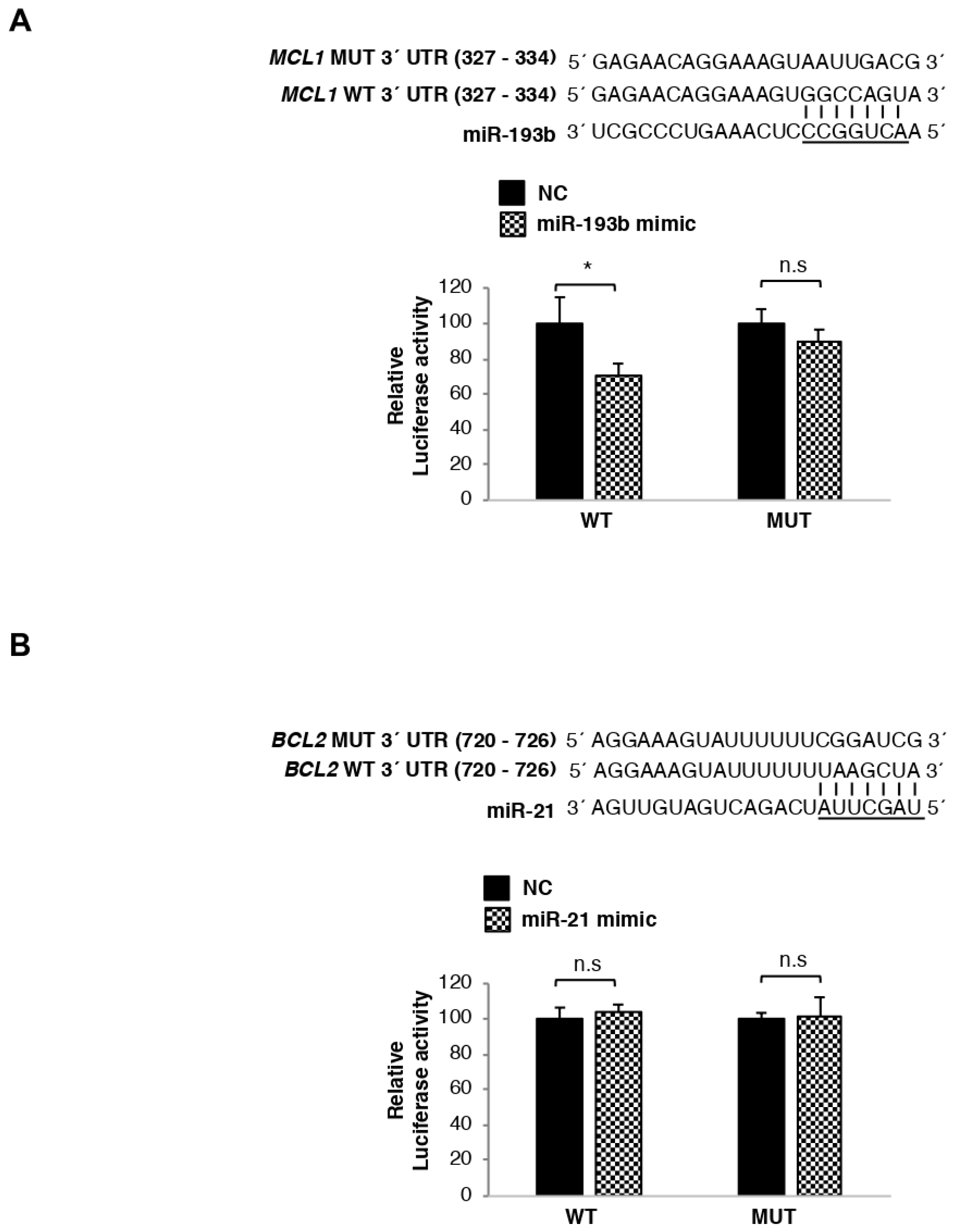
3.5. MCL1 mRNA Is Directly Regulated by miR-193b, Whereas BCL2 Transcript Is Not Targeted by miR-21
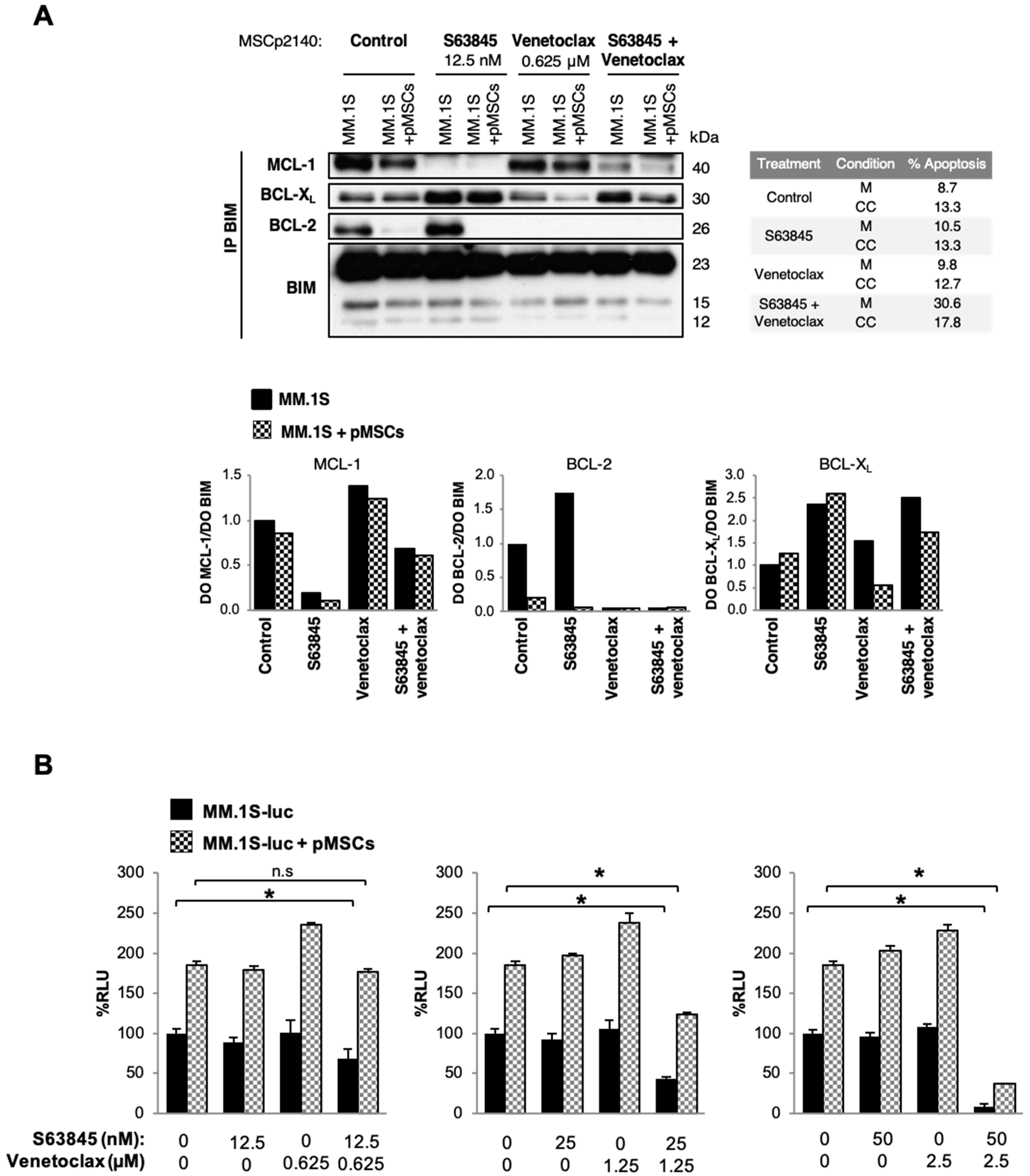
3.6. The Presence of Stromal Cells Modifies Interactions of MCL-1 and BCL-2 with BIM in Untreated and S63845- or Venetoclax-Treated MM.1S Cells
3.7. S63845 Potently Synergizes with Venetoclax in the Presence of pMSCs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moreau, P.; van de Donk, N.W.C.J.; San Miguel, J.; Lokhorst, H.; Nahi, H.; Ben-Yehuda, D.; Cavo, M.; Cook, G.; Delforge, M.; Einsele, H.; et al. Practical Considerations for the Use of Daratumumab, a Novel CD38 Monoclonal Antibody, in Myeloma. Drugs 2016, 76, 853–867. [Google Scholar] [CrossRef] [PubMed]
- Mateos, M.-V.; Ludwig, H.; Bazarbachi, A.; Beksac, M.; Bladé, J.; Boccadoro, M.; Cavo, M.; Delforge, M.; Dimopoulos, M.A.; Facon, T.; et al. Insights on Multiple Myeloma Treatment Strategies. Hemasphere 2018, 3, e163. [Google Scholar] [CrossRef]
- Hideshima, T.; Mitsiades, C.; Tonon, G.; Richardson, P.G.; Anderson, K.C. Understanding multiple myeloma pathogenesis in the bone marrow to identify new therapeutic targets. Nat. Rev. Cancer 2007, 7, 585–598. [Google Scholar] [CrossRef] [PubMed]
- Lomas, O.C.; Tahri, S.; Ghobrial, I.M. The microenvironment in myeloma. Curr. Opin. Oncol. 2020, 32, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Ocio, E.M.; Mitsiades, C.S.; Orlowski, R.; Anderson, K.C. Future agents and treatment directions in multiple myeloma. Expert Rev. Hematol. 2014, 7, 127–141. [Google Scholar] [CrossRef]
- McMillin, D.W.; Delmore, J.; Weisberg, E.; Negri, J.M.; Geer, D.C.; Klippel, S.; Mitsiades, N.; Schlossman, R.L.; Munshi, N.C.; Kung, A.L.; et al. Tumor cell-specific bioluminescence platform to identify stroma-induced changes to anticancer drug activity. Nat. Med. 2010, 16, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Faict, S.; Maes, K.; De Bruyne, E.; Van Valckenborgh, E.; Schots, R.; Vanderkerken, K.; Menu, E. Extracellular vesicle cross-talk in the bone marrow microenvironment: Implications in multiple myeloma. Oncotarget 2016, 7, 38927–38945. [Google Scholar] [CrossRef]
- McMillin, D.W.; Negri, J.M.; Mitsiades, C.S. The role of tumour-stromal interactions in modifying drug response: Challenges and opportunities. Nat. Rev. Drug Discov. 2013, 12, 217–228. [Google Scholar] [CrossRef]
- Raimondi, L.; De Luca, A.; Morelli, E.; Giavaresi, G.; Tagliaferri, P.; Tassone, P.; Amodio, N. MicroRNAs: Novel Crossroads between Myeloma Cells and the Bone Marrow Microenvironment. Biomed. Res. Int. 2016, 2016, 6504593. [Google Scholar] [CrossRef]
- Hao, M.; Zhang, L.; An, G.; Meng, H.; Han, Y.; Xie, Z.; Xu, Y.; Li, C.; Yu, Z.; Chang, H.; et al. Bone marrow stromal cells protect myeloma cells from bortezomib induced apoptosis by suppressing microRNA-15a expression. Leuk. Lymphoma 2011, 52, 1787–1794. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, C.; Ju, S.; Wang, Y.; Wang, H.; Zhong, R. Myeloma cell adhesion to bone marrow stromal cells confers drug resistance by microRNA-21 up-regulation. Leuk. Lymphoma 2011, 52, 1991–1998. [Google Scholar] [CrossRef]
- Morelli, E.; Leone, E.; Cantafio, M.E.G.; Di Martino, M.T.; Amodio, N.; Biamonte, L.; Gullà, A.; Foresta, U.; Pitari, M.R.; Botta, C.; et al. Selective targeting of IRF4 by synthetic microRNA-125b-5p mimics induces anti-multiple myeloma activity in vitro and in vivo. Leukemia 2015, 29, 2173–2183. [Google Scholar] [CrossRef] [PubMed]
- Gulla, A.; Di Martino, M.T.; Gallo Cantafio, M.E.; Morelli, E.; Amodio, N.; Botta, C.; Pitari, M.R.; Lio, S.G.; Britti, D.; Stamato, M.A.; et al. A 13 mer LNA-i-miR-221 Inhibitor Restores Drug Sensitivity in Melphalan-Refractory Multiple Myeloma Cells. Clin. Cancer Res. 2016, 22, 1222–1233. [Google Scholar] [CrossRef] [PubMed]
- Abramson, H.N. The Multiple Myeloma Drug Pipeline-2018: A Review of Small Molecules and Their Therapeutic Targets. Clin. Lymphoma Myeloma Leuk. 2018, 18, 611–627. [Google Scholar] [CrossRef]
- Vogler, M.; Walter, H.S.; Dyer, M.J.S. Targeting anti-apoptotic BCL2 family proteins in haematological malignancies—From pathogenesis to treatment. Br. J. Haematol. 2017, 178, 364–379. [Google Scholar] [CrossRef] [PubMed]
- Del Gaizo Moore, V.; Brown, J.R.; Certo, M.; Love, T.M.; Novina, C.D.; Letai, A. Chronic lymphocytic leukemia requires BCL2 to sequester prodeath BIM, explaining sensitivity to BCL2 antagonist ABT-737. J. Clin. Investig. 2007, 117, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.A.; Matulis, S.M.; Conage-Pough, J.E.; Nooka, A.K.; Kaufman, J.L.; Lonial, S.; Boise, L.H. Bone marrow microenvironment–derived signals induce Mcl-1 dependence in multiple myeloma. Blood 2017, 129, 1969–1979. [Google Scholar] [CrossRef]
- Roberts, A.W.; Seymour, J.F.; Brown, J.R.; Wierda, W.G.; Kipps, T.J.; Khaw, S.L.; Carney, D.A.; He, S.Z.; Huang, D.C.S.; Xiong, H.; et al. Substantial Susceptibility of Chronic Lymphocytic Leukemia to BCL2 Inhibition: Results of a Phase I Study of Navitoclax in Patients With Relapsed or Refractory Disease. J. Clin. Oncol. 2012, 30, 488–496. [Google Scholar] [CrossRef]
- Kumar, S.; Kaufman, J.L.; Gasparetto, C.; Mikhael, J.; Vij, R.; Pegourie, B.; Benboubker, L.; Facon, T.; Amiot, M.; Moreau, P.; et al. Efficacy of venetoclax as targeted therapy for relapsed/refractory t(11;14) multiple myeloma. Blood 2017, 130, 2401–2409. [Google Scholar] [CrossRef]
- Wuillème-Toumi, S.; Robillard, N.; Gomez, P.; Moreau, P.; Le Gouill, S.; Avet-Loiseau, H.; Harousseau, J.-L.; Amiot, M.; Bataille, R. Mcl-1 is overexpressed in multiple myeloma and associated with relapse and shorter survival. Leukemia 2005, 19, 1248–1252. [Google Scholar] [CrossRef] [PubMed]
- Derenne, S. Antisense strategy shows that Mcl-1 rather than Bcl-2 or Bcl-xL is an essential survival protein of human myeloma cells. Blood 2002, 100, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B. Myeloid cell factor-1 is a critical survival factor for multiple myeloma. Blood 2002, 99, 1885–1893. [Google Scholar] [CrossRef]
- Kotschy, A.; Szlavik, Z.; Murray, J.; Davidson, J.; Maragno, A.L.; Le Toumelin-Braizat, G.; Chanrion, M.; Kelly, G.L.; Gong, J.-N.; Moujalled, D.M.; et al. The MCL1 inhibitor S63845 is tolerable and effective in diverse cancer models. Nature 2016, 538, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Tron, A.E.; Belmonte, M.A.; Adam, A.; Aquila, B.M.; Boise, L.H.; Chiarparin, E.; Cidado, J.; Embrey, K.J.; Gangl, E.; Gibbons, F.D.; et al. Discovery of Mcl-1-specific inhibitor AZD5991 and preclinical activity in multiple myeloma and acute myeloid leukemia. Nat. Commun. 2018, 9, 5341. [Google Scholar] [CrossRef]
- Caenepeel, S.; Brown, S.P.; Belmontes, B.; Moody, G.; Keegan, K.S.; Chui, D.; Whittington, D.A.; Huang, X.; Poppe, L.; Cheng, A.C.; et al. AMG 176, a Selective MCL1 Inhibitor, is Effective in Hematological Cancer Models Alone and in Combination with Established Therapies. Cancer Discov. 2018, CD-18-0387. [Google Scholar] [CrossRef]
- Algarín, E.M.; Díaz-Tejedor, A.; Mogollón, P.; Hernández-García, S.; Corchete, L.A.; San-Segundo, L.; Martín-Sánchez, M.; González-Méndez, L.; Schoumacher, M.; Banquet, S.; et al. Preclinical evaluation of the simultaneous inhibition of MCL-1 and BCL-2 with the combination of S63845 and venetoclax in multiple myeloma. Haematologica 2019. [Google Scholar] [CrossRef]
- Jourdan, M.; Veyrune, J.-L.; Vos, J.D.; Redal, N.; Couderc, G.; Klein, B. A major role for Mcl-1 antiapoptotic protein in the IL-6-induced survival of human myeloma cells. Oncogene 2003, 22, 2950–2959. [Google Scholar] [CrossRef] [PubMed]
- Conage-Pough, J.E.; Boise, L.H. Phosphorylation alters Bim-mediated Mcl-1 stabilization and priming. FEBS J. 2018, 285, 2626–2640. [Google Scholar] [CrossRef]
- Garcia-Gomez, A.; Quwaider, D.; Canavese, M.; Ocio, E.M.; Tian, Z.; Blanco, J.F.; Berger, A.J.; Ortiz-de-Solorzano, C.; Hernandez-Iglesias, T.; Martens, A.C.M.; et al. Preclinical Activity of the Oral Proteasome Inhibitor MLN9708 in Myeloma Bone Disease. Clin. Cancer Res. 2014, 20, 1542–1554. [Google Scholar] [CrossRef]
- Tessoulin, B.; Papin, A.; Gomez-Bougie, P.; Bellanger, C.; Amiot, M.; Pellat-Deceunynck, C.; Chiron, D. BCL2-Family Dysregulation in B-Cell Malignancies: From Gene Expression Regulation to a Targeted Therapy Biomarker. Front. Oncol. 2018, 8, 645. [Google Scholar] [CrossRef] [PubMed]
- Montero, J.; Letai, A. Why do BCL-2 inhibitors work and where should we use them in the clinic? Cell Death Differ. 2018, 25, 56–64. [Google Scholar] [CrossRef]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef]
- Del Gaizo Moore, V.; Letai, A. BH3 profiling – Measuring integrated function of the mitochondrial apoptotic pathway to predict cell fate decisions. Cancer Lett. 2013, 332, 202–205. [Google Scholar] [CrossRef]
- Gomez-Bougie, P.; Maiga, S.; Tessoulin, B.; Bourcier, J.; Bonnet, A.; Rodriguez, M.S.; Le Gouill, S.; Touzeau, C.; Moreau, P.; Pellat-Deceunynck, C.; et al. BH3-mimetic toolkit guides the respective use of BCL2 and MCL1 BH3-mimetics in myeloma treatment. Blood 2018, 132, 2656–2669. [Google Scholar] [CrossRef] [PubMed]
- Jourdan, M.; De Vos, J.; Mechti, N.; Klein, B. Regulation of Bcl-2-family proteins in myeloma cells by three myeloma survival factors: Interleukin-6, interferon-alpha and insulin-like growth factor 1. Cell Death Differ. 2000, 7, 1244–1252. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, H. LncRNA NEAT1 promotes dexamethasone resistance in multiple myeloma by targeting miR-193a/MCL1 pathway. J. Biochem. Mol. Toxicol. 2018, 32, e22008. [Google Scholar] [CrossRef]
- Lam, L.T.; Lu, X.; Zhang, H.; Lesniewski, R.; Rosenberg, S.; Semizarov, D. A MicroRNA Screen to Identify Modulators of Sensitivity to BCL2 Inhibitor ABT-263 (Navitoclax). Mol. Cancer Ther. 2010, 9, 2943–2950. [Google Scholar] [CrossRef]
- Slomp, A.; Moesbergen, L.M.; Gong, J.; Cuenca, M.; von dem Borne, P.A.; Sonneveld, P.; Huang, D.C.S.; Minnema, M.C.; Peperzak, V. Multiple myeloma with 1q21 amplification is highly sensitive to MCL-1 targeting. Blood Adv. 2019, 3, 4202–4214. [Google Scholar] [CrossRef]
- Sims, E.K.; Lakhter, A.; Anderson-Baucum, E.; Kono, T.; Tong, X.; Evans-Molina, C. MicroRNA 21 targets BCL2 mRNA to increase cell apoptosis in rat and human beta cells. Diabetologia 2017, 60, 1057–1065. [Google Scholar] [CrossRef]
- Wickramasinghe, N.S.; Manavalan, T.T.; Dougherty, S.M.; Riggs, K.A.; Li, Y.; Klinge, C.M. Estradiol downregulates miR-21 expression and increases miR-21 target gene expression in MCF-7 breast cancer cells. Nucleic Acids Res. 2009, 37, 2584–2595. [Google Scholar] [CrossRef]
- Liu, K.; Du, J.; Ruan, L. MicroRNA-21 regulates the viability and apoptosis of diffuse large B-cell lymphoma cells by upregulating B cell lymphoma-2. Exp. Ther. Med. 2017. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Zhao, Y.-P.; Zhou, L.; Zhang, T.-P.; Chen, G. Bcl-2 Upregulation Induced by miR-21 Via a Direct Interaction Is Associated with Apoptosis and Chemoresistance in MIA PaCa-2 Pancreatic Cancer Cells. Arch. Med. Res. 2011, 42, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yan, L.; Zhang, W.; Wang, H.; Chen, W.; Hu, N.; Ou, H. miR-21 inhibitor suppresses proliferation and migration of nasopharyngeal carcinoma cells through down-regulation of BCL2 expression. Int. J. Clin. Exp. Pathol. 2014, 7, 3478–3487. [Google Scholar] [PubMed]
- Morales, A.A.; Kurtoglu, M.; Matulis, S.M.; Liu, J.; Siefker, D.; Gutman, D.M.; Kaufman, J.L.; Lee, K.P.; Lonial, S.; Boise, L.H. Distribution of Bim determines Mcl-1 dependence or codependence with Bcl-xL/Bcl-2 in Mcl-1-expressing myeloma cells. Blood 2011, 118, 1329–1339. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MCL1 mRNA | BCL2 mRNA |
|---|---|
| hsa-miR-125a-5p | hsa-miR-16-5p |
| hsa-miR-4319 | hsa-miR-6838-5p |
| hsa-miR-125b-5p | hsa-miR-195-5p |
| hsa-miR-29a-3p | hsa-miR-424-5p |
| hsa-miR-29b-3p | hsa-miR-15a-5p |
| hsa-miR-29c-3p | hsa-miR-15b-5p |
| hsa-miR-526b-3p | hsa-miR-497-5p |
| hsa-miR-106a-5p | hsa-miR-4262 |
| hsa-miR-20b-5p | hsa-miR-181d-5p |
| hsa-miR-93-5p | hsa-miR-181b-5p |
| hsa-miR-519d-3p | hsa-miR-181a-5p |
| hsa-miR-17-5p | hsa-miR-181c-5p |
| hsa-miR-20a-5p | hsa-miR-125a-5p |
| hsa-miR-106b-5p | hsa-miR-125b-5p |
| hsa-miR-520f-3p | hsa-miR-4319 |
| hsa-miR-302c-3p.2 | hsa-miR-153-3p |
| hsa-miR-133a-3p.1 | hsa-miR-182-5p |
| hsa-miR-133b | hsa-miR-30d-5p |
| hsa-miR-133a-3p.2 | hsa-miR-30e-5p |
| hsa-miR-373-3p | hsa-miR-30a-5p |
| hsa-miR-372-3p | hsa-miR-30b-5p |
| hsa-miR-520a-3p | hsa-miR-30c-5p |
| hsa-miR-520d-3p | hsa-miR-449a |
| hsa-miR-520e | hsa-miR-449b-5p |
| hsa-miR-302d-3p | hsa-miR-34a-5p |
| hsa-miR-302b-3p | hsa-miR-34c-5p |
| hsa-miR-302c-3p.1 | hsa-miR-96-5p |
| hsa-miR-520c-3p | hsa-miR-1271-5p |
| hsa-miR-302a-3p | hsa-miR-200b-3p |
| hsa-miR-520b | hsa-miR-429 |
| hsa-miR-302e | hsa-miR-200c-3p |
| hsa-miR-135a-5p | hsa-miR-365b-3p |
| hsa-miR-135b-5p | hsa-miR-365a-3p |
| hsa-miR-153-3p | hsa-miR-204-5p |
| hsa-miR-4262 | hsa-miR-211-5p |
| hsa-miR-181b-5p | hsa-miR-140-3p.2 |
| hsa-miR-181a-5p | hsa-miR-21-5p |
| hsa-miR-181d-5p | hsa-miR-590-5p |
| hsa-miR-181c-5p | hsa-miR-6088 |
| hsa-miR-5590-3p | hsa-miR-143-3p |
| hsa-miR-142-5p | hsa-miR-4770 |
| hsa-miR-193a-3p | hsa-miR-383-5p.2 |
| hsa-miR-193b-3p | hsa-miR-383-5p.1 |
| hsa-miR-4465 | hsa-miR-23b-3p |
| hsa-miR-26b-5p | hsa-miR-130a-5p |
| hsa-miR-26a-5p | hsa-miR-23c |
| hsa-miR-1297 | hsa-miR-23a-3p |
| hsa-miR-101-3p.1 | hsa-miR-202-5p |
| hsa-miR-101-3p.2 | hsa-miR-448 |
| hsa-miR-325-3p | hsa-miR-342-3p |
| hsa-miR-873-5p.1 | hsa-miR-503-5p |
| hsa-miR-381-3p | hsa-miR-382-3p |
| hsa-miR-300 | hsa-miR-219a-2-3p |
| hsa-miR-374c-5p | |
| hsa-miR-655-3p | |
| hsa-miR-323a-3p | |
| hsa-miR-6835-3p | |
| hsa-miR-224-5p |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Algarín, E.M.; Quwaider, D.; Campos-Laborie, F.J.; Díaz-Tejedor, A.; Mogollón, P.; Vuelta, E.; Martín-Sánchez, M.; San-Segundo, L.; González-Méndez, L.; Gutiérrez, N.C.; et al. Stroma-Mediated Resistance to S63845 and Venetoclax through MCL-1 and BCL-2 Expression Changes Induced by miR-193b-3p and miR-21-5p Dysregulation in Multiple Myeloma. Cells 2021, 10, 559. https://doi.org/10.3390/cells10030559
Algarín EM, Quwaider D, Campos-Laborie FJ, Díaz-Tejedor A, Mogollón P, Vuelta E, Martín-Sánchez M, San-Segundo L, González-Méndez L, Gutiérrez NC, et al. Stroma-Mediated Resistance to S63845 and Venetoclax through MCL-1 and BCL-2 Expression Changes Induced by miR-193b-3p and miR-21-5p Dysregulation in Multiple Myeloma. Cells. 2021; 10(3):559. https://doi.org/10.3390/cells10030559
Chicago/Turabian StyleAlgarín, Esperanza M., Dalia Quwaider, Francisco J. Campos-Laborie, Andrea Díaz-Tejedor, Pedro Mogollón, Elena Vuelta, Montserrat Martín-Sánchez, Laura San-Segundo, Lorena González-Méndez, Norma C. Gutiérrez, and et al. 2021. "Stroma-Mediated Resistance to S63845 and Venetoclax through MCL-1 and BCL-2 Expression Changes Induced by miR-193b-3p and miR-21-5p Dysregulation in Multiple Myeloma" Cells 10, no. 3: 559. https://doi.org/10.3390/cells10030559
APA StyleAlgarín, E. M., Quwaider, D., Campos-Laborie, F. J., Díaz-Tejedor, A., Mogollón, P., Vuelta, E., Martín-Sánchez, M., San-Segundo, L., González-Méndez, L., Gutiérrez, N. C., García-Sanz, R., Paíno, T., De Las Rivas, J., Ocio, E. M., & Garayoa, M. (2021). Stroma-Mediated Resistance to S63845 and Venetoclax through MCL-1 and BCL-2 Expression Changes Induced by miR-193b-3p and miR-21-5p Dysregulation in Multiple Myeloma. Cells, 10(3), 559. https://doi.org/10.3390/cells10030559