
Neuroimmune Regulation of Surgery-Associated Metastases




Abstract
1. Introduction
2. The Neuronal Regulation of Tumor Progression
3. Tissue Damage and Wounding Affect Tumor Progression
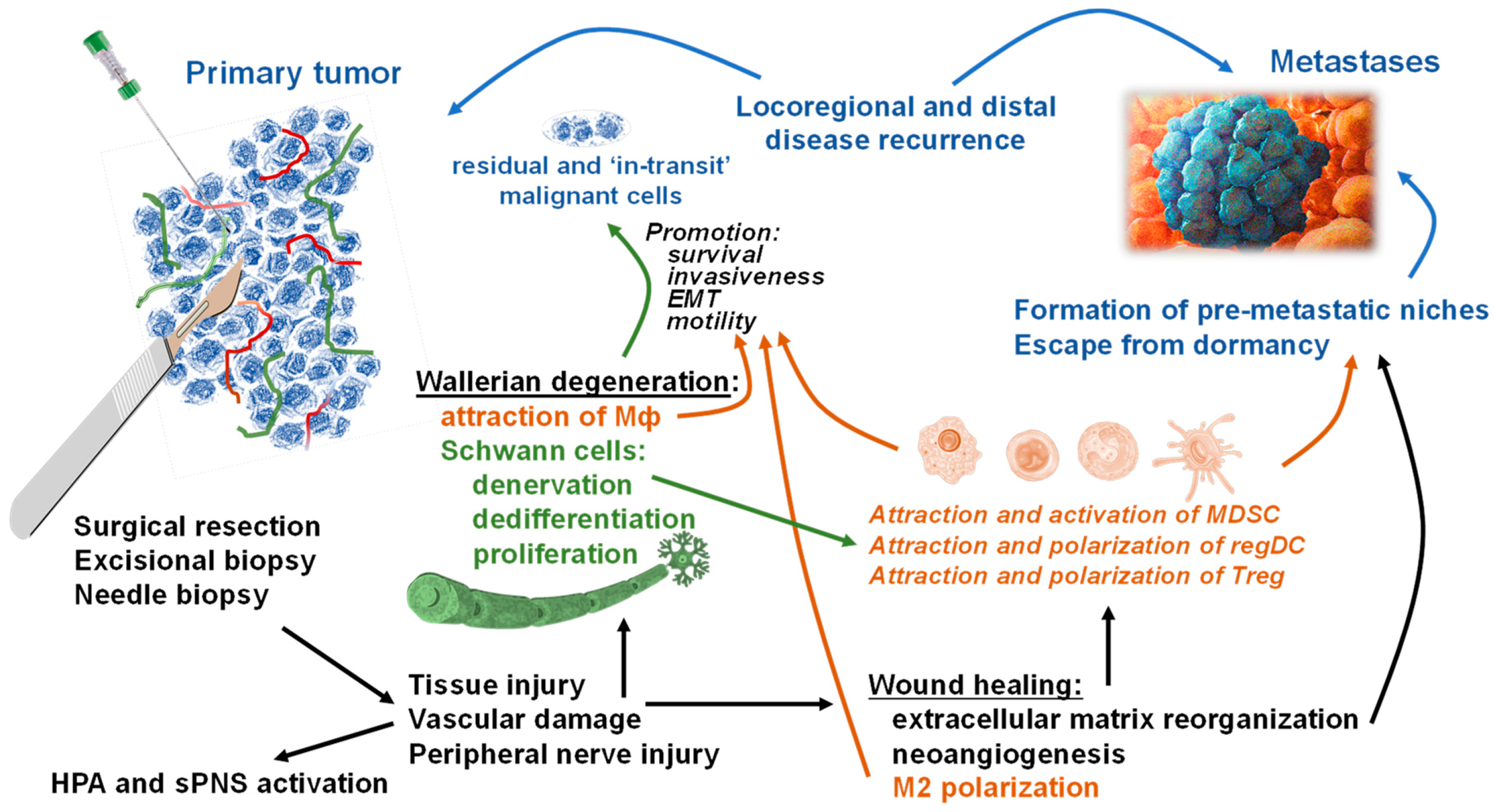
4. PNS Functioning in Surgery-Associated Metastasis Formation
5. Future Directions
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Cancer Society Official Page. Cancer Facts and Figures 2020. Available online: https://www.cancer.org/content/dam/cancer-org/research/cancer-facts-and-statistics/annual-cancer-facts-and-figures/2020/cancer-facts-and-figures-2020.pdf (accessed on 1 November 2020).
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA A Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Simpson-Herren, L.; Sanford, A.H.; Holmquist, J.P. Effects of surgery on the cell kinetics of residual tumor. Cancer Treat. Rep. 1976, 60, 1749–1760. [Google Scholar] [PubMed]
- Horg, S.A.; Rubin, P.; DeWys, W.D. Metastasis and disseminated disease. In Clinical Oncology for Medical Students and Physicians-A Multidisciplinary Approach, 6th ed.; American Cancer Society: New York, NY, USA, 1983; pp. 498–499. [Google Scholar]
- Steeg, P.S. Targeting metastasis. Nat. Rev. Cancer 2016, 16, 201–218. [Google Scholar] [CrossRef]
- Chaffer, C.L.; Weinberg, R.A. A perspective on cancer cell metastasis. Science 2011, 331, 1559–1564. [Google Scholar] [CrossRef]
- Dillekas, H.; Rogers, M.S.; Straume, O. Are 90% of deaths from cancer caused by metastases? Cancer Med 2019, 8, 5574–5576. [Google Scholar] [CrossRef] [PubMed]
- Pommier, A.; Anaparthy, N.; Memos, N.; Kelley, Z.L.; Gouronnec, A.; Yan, R.; Auffray, C.; Albrengues, J.; Egeblad, M.; Iacobuzio-Donahue, C.A.; et al. Unresolved endoplasmic reticulum stress engenders immune-resistant, latent pancreatic cancer metastases. Science 2018, 360. [Google Scholar] [CrossRef]
- Al-Sahaf, O.; Wang, J.H.; Browne, T.J.; Cotter, T.G.; Redmond, H.P. Surgical injury enhances the expression of genes that mediate breast cancer metastasis to the lung. Ann. Surg. 2010, 252, 1037–1043. [Google Scholar] [CrossRef]
- Lee, J.W.; Shahzad, M.M.; Lin, Y.G.; Armaiz-Pena, G.; Mangala, L.S.; Han, H.D.; Kim, H.S.; Nam, E.J.; Jennings, N.B.; Halder, J.; et al. Surgical stress promotes tumor growth in ovarian carcinoma. Clin. Cancer Res. 2009, 15, 2695–2702. [Google Scholar] [CrossRef]
- Demicheli, R.; Valagussa, P.; Bonadonna, G. Does surgery modify growth kinetics of breast cancer micrometastases? Br. J. Cancer 2001, 85, 490–492. [Google Scholar] [CrossRef]
- Demicheli, R.; Retsky, M.W.; Hrushesky, W.J.; Baum, M. Tumor dormancy and surgery-driven interruption of dormancy in breast cancer: Learning from failures. Nat. Clin. Pract. Oncol. 2007, 4, 699–710. [Google Scholar] [CrossRef]
- Kelsey, C.R.; Fornili, M.; Ambrogi, F.; Higgins, K.; Boyd, J.A.; Biganzoli, E.; Demicheli, R. Metastasis dynamics for non-small-cell lung cancer: Effect of patient and tumor-related factors. Clin. Lung Cancer 2013, 14, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Oosterling, S.J.; van der Bij, G.J.; van Egmond, M.; van der Sijp, J.R. Surgical trauma and peritoneal recurrence of colorectal carcinoma. Eur. J. Surg. Oncol. 2005, 31, 29–37. [Google Scholar] [CrossRef]
- van der Bij, G.J.; Oosterling, S.J.; Beelen, R.H.; Meijer, S.; Coffey, J.C.; van Egmond, M. The perioperative period is an underutilized window of therapeutic opportunity in patients with colorectal cancer. Ann. Surg. 2009, 249, 727–734. [Google Scholar] [CrossRef]
- Retsky, M.; Demicheli, R.; Hrushesky, W.; Baum, M.; Gukas, I. Surgery triggers outgrowth of latent distant disease in breast cancer: An inconvenient truth? Cancers 2010, 2, 305–337. [Google Scholar] [CrossRef]
- Retsky, M.; Demicheli, R.; Hrushesky, W.J.; Forget, P.; De Kock, M.; Gukas, I.; Rogers, R.A.; Baum, M.; Sukhatme, V.; Vaidya, J.S. Reduction of breast cancer relapses with perioperative non-steroidal anti-inflammatory drugs: New findings and a review. Curr. Med. Chem. 2013, 20, 4163–4176. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.; Torosian, M.H.; Boraas, M.C.; Sigurdson, E.R.; Hoffman, J.P.; Eisenberg, B.L.; Fowble, B. Local recurrence of breast cancer in the stereotactic core needle biopsy site: Case reports and review of the literature. Breast J. 2001, 7, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Hansen, N.M.; Ye, X.; Grube, B.J.; Giuliano, A.E. Manipulation of the primary breast tumor and the incidence of sentinel node metastases from invasive breast cancer. Arch. Surg. 2004, 139, 634–639; discussion 639–640. [Google Scholar] [CrossRef] [PubMed]
- Hocevar, M.; Dragonja, Z.; Pilko, G.; Gazic, B.; Zgajnar, J. Residual melanoma after an excisional biopsy is an independent prognostic factor for local recurrence and overall survival. Eur. J. Surg. Oncol. J. Eur. Soc. Surg. Oncol. Br. Assoc. Surg. Oncol. 2014, 40, 1271–1275. [Google Scholar] [CrossRef]
- Mathenge, E.G.; Dean, C.A.; Clements, D.; Vaghar-Kashani, A.; Photopoulos, S.; Coyle, K.M.; Giacomantonio, M.; Malueth, B.; Nunokawa, A.; Jordan, J.; et al. Core needle biopsy of breast cancer tumors increases distant metastases in a mouse model. Neoplasia 2014, 16, 950–960. [Google Scholar] [CrossRef]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef]
- Govaert, K.M.; Jongen, J.M.J.; Kranenburg, O.; Borel Rinkes, I.H.M. Surgery-induced tumor growth in (metastatic) colorectal cancer. Surg. Oncol. 2017, 26, 535–543. [Google Scholar] [CrossRef]
- Perry, J.A.; Douglas, H. Immunomodulatory Effects of Surgery, Pain, and Opioids in Cancer Patients. The Veterinary clinics of North America. Small Anim. Pract. 2019, 49, 981–991. [Google Scholar] [CrossRef]
- Ryall, C. Cancer infection and cancer recurrence: A anger to avoid in cancer operations. Lancet 1907, 2, 1311–1316. [Google Scholar] [CrossRef]
- Tyzzer, E.E. Factors in the Production and Growth of tumor Metastases. J. Med Res. 1913, 28, 309–332-1. [Google Scholar] [PubMed]
- Jones, F.S.; Rous, P. On the cause of the localization of secondary tumors at points of injury. J. Exp. Med. 1914, 20, 404–412. [Google Scholar] [CrossRef]
- Schatten, W.E. An experimental study of postoperative tumor metastases. I. Growth of pulmonary metastases following total removal of primary leg tumor. Cancer 1958, 11, 455–459. [Google Scholar] [CrossRef]
- Lewis, M.R.; Cole, W.H. Experimental increase of lung metastases after operative trauma (amputation of limb with tumor). AMA Arch. Surg. 1958, 77, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Morgenstern, L.; Yamakawa, T.; Seltzer, D. Carcinoma of the gastric stump. Am. J. Surg. 1973, 125, 29–38. [Google Scholar] [CrossRef]
- Lange, P.H.; Hekmat, K.; Bosl, G.; Kennedy, B.J.; Fraley, E.E. Acclerated growth of testicular cancer after cytoreductive surgery. Cancer 1980, 45, 1498–1506. [Google Scholar] [CrossRef]
- Moore, G.E. Debunking debulking. Surg. Gynecol. Obs. 1980, 150, 395–396. [Google Scholar]
- Hughes, E.S.; McDermott, F.T.; Polglase, A.L.; Johnson, W.R. Tumor recurrence in the abdominal wall scar tissue after large-bowel cancer surgery. Dis. Colon Rectum 1983, 26, 571–572. [Google Scholar] [CrossRef]
- Baker, D.G.; Masterson, T.M.; Pace, R.; Constable, W.C.; Wanebo, H. The influence of the surgical wound on local tumor recurrence. Surgery 1989, 106, 525–532. [Google Scholar] [PubMed]
- Hoskins, W.J. The influence of cytoreductive surgery on progression-free interval and survival in epithelial ovarian cancer. Baillieres Clin. Obs. Gynaecol. 1989, 3, 59–71. [Google Scholar] [CrossRef]
- Schuh, A.C.; Keating, S.J.; Monteclaro, F.S.; Vogt, P.K.; Breitman, M.L. Obligatory wounding requirement for tumorigenesis in v-jun transgenic mice. Nature 1990, 346, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Arai, K.; Asakura, T.; Nemir, P., Jr. Effect of local tumor removal and retained oncolysate on lung metastasis. J. Surg. Res. 1992, 53, 30–38. [Google Scholar] [CrossRef]
- Ramos, J.M.; Gupta, S.; Anthone, G.J.; Ortega, A.E.; Simons, A.J.; Beart, R.W., Jr. Laparoscopy and colon cancer. Is the port site at risk? A preliminary report. Arch. Surg. 1994, 129, 897–899; discussion 900. [Google Scholar] [CrossRef]
- Lee, J.Y.; Murphy, S.M.; Scanlon, E.F. Effect of trauma of implantation of metastatic tumor in bone in mice. J. Surg. Oncol. 1994, 56, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Watson, D.I. Abdominal wall metastasis after laparoscopic gastroenterostomy. Med. J. Aust. 1995, 163, 106–107. [Google Scholar] [CrossRef]
- Mitsudomi, T.; Nishioka, K.; Maruyama, R.; Saitoh, G.; Hamatake, M.; Fukuyama, Y.; Yaita, H.; Ishida, T.; Sugimachi, K. Kinetic analysis of recurrence and survival after potentially curative resection of nonsmall cell lung cancer. J. Surg. Oncol. 1996, 63, 159–165. [Google Scholar] [CrossRef]
- Hofer, S.O.; Shrayer, D.; Reichner, J.S.; Hoekstra, H.J.; Wanebo, H.J. Wound-induced tumor progression: A probable role in recurrence after tumor resection. Arch. Surg. 1998, 133, 383–389. [Google Scholar] [CrossRef]
- Karrison, T.G.; Ferguson, D.J.; Meier, P. Dormancy of mammary carcinoma after mastectomy. J. Natl. Cancer Inst. 1999, 91, 80–85. [Google Scholar] [CrossRef]
- Pidgeon, G.P.; Harmey, J.H.; Kay, E.; Da Costa, M.; Redmond, H.P.; Bouchier-Hayes, D.J. The role of endotoxin/lipopolysaccharide in surgically induced tumour growth in a murine model of metastatic disease. Br. J. Cancer 1999, 81, 1311–1317. [Google Scholar] [CrossRef] [PubMed]
- Maniwa, Y.; Kanki, M.; Okita, Y. Importance of the control of lung recurrence soon after surgery of pulmonary metastases. Am. J. Surg. 2000, 179, 122–125. [Google Scholar] [CrossRef]
- Da Costa, M.L.; Redmond, H.P.; Bouchier-Hayes, D.J. Taurolidine improves survival by abrogating the accelerated development and proliferation of solid tumors and development of organ metastases from circulating tumor cells released following surgery. J. Surg. Res. 2001, 101, 111–119. [Google Scholar] [CrossRef]
- Li, T.S.; Kaneda, Y.; Ueda, K.; Hamano, K.; Zempo, N.; Esato, K. The influence of tumour resection on angiostatin levels and tumour growth--an experimental study in tumour-bearing mice. Eur. J. Cancer 2001, 37, 2283–2288. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Sawada, S.; Yoshioka, I.; Ohashi, Y.; Matsuo, M.; Harimaya, Y.; Tsukada, K.; Saiki, I. Increased surgical stress promotes tumor metastasis. Surgery 2003, 133, 547–555. [Google Scholar] [CrossRef]
- Shakhar, G.; Abudarham, N.; Melamed, R.; Schwartz, Y.; Rosenne, E.; Ben-Eliyahu, S. Amelioration of operation-induced suppression of marginating pulmonary NK activity using poly IC: A potential approach to reduce postoperative metastasis. Ann. Surg. Oncol. 2007, 14, 841–852. [Google Scholar] [CrossRef]
- Beecken, W.D.; Engl, T.; Jonas, D.; Blaheta, R.A. Expression of angiogenesis inhibitors in human bladder cancer may explain rapid metastatic progression after radical cystectomy. Int. J. Mol. Med. 2009, 23, 261–266. [Google Scholar]
- Goldfarb, Y.; Shapiro, H.; Singer, P.; Kalderon, Y.; Levi, B.; Glasner, A.; Benish, M.; Ben-Eliyahu, S. Fish oil attenuates surgery-induced immunosuppression, limits post-operative metastatic dissemination and increases long-term recurrence-free survival in rodents inoculated with cancer cells. Clin. Nutr. 2012, 31, 396–404. [Google Scholar] [CrossRef]
- Tham, M.; Khoo, K.; Yeo, K.P.; Kato, M.; Prevost-Blondel, A.; Angeli, V.; Abastado, J.P. Macrophage depletion reduces postsurgical tumor recurrence and metastatic growth in a spontaneous murine model of melanoma. Oncotarget 2015, 6, 22857–22868. [Google Scholar] [CrossRef]
- O’Connell, E.P.; O’Leary, D.P.; Fogarty, K.; Khan, Z.J.; Redmond, H.P. Predictors and patterns of melanoma recurrence following a negative sentinel lymph node biopsy. Melanoma Res. 2016, 26, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Pinson, H.; Cosyns, S.; Ceelen, W.P. The impact of surgical resection of the primary tumor on the development of synchronous colorectal liver metastasis: A systematic review. Acta Chir. Belg. 2018, 118, 203–211. [Google Scholar] [CrossRef]
- Zhang, Q.; Shan, F.; Li, Z.; Gao, J.; Li, Y.; Shen, L.; Ji, J.; Lu, M. A prospective study on the changes and clinical significance of pre-operative and post-operative circulating tumor cells in resectable gastric cancer. J. Transl. Med. 2018, 16, 171. [Google Scholar] [CrossRef]
- Grewal, S.; Korthouwer, R.; Bögels, M.; Braster, R.; Heemskerk, N.; Budding, A.E.; Pouw, S.M.; van Horssen, J.; Ankersmit, M.; Meijerink, J.; et al. Spillage of bacterial products during colon surgery increases the risk of liver metastases development in a rat colon carcinoma model. Oncoimmunology 2018, 7, e1461302. [Google Scholar] [CrossRef] [PubMed]
- Ou, H.; Huang, Y.; Xiang, L.; Chen, Z.; Fang, Y.; Lin, Y.; Cui, Z.; Yu, S.; Li, X.; Yang, D. Circulating Tumor Cell Phenotype Indicates Poor Survival and Recurrence After Surgery for Hepatocellular Carcinoma. Dig. Dis. Sci. 2018, 63, 2373–2380. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Zhu, Y.; Cui, Y.; Yang, Z.; Zhou, S.; Han, Y.; Yu, D.; Xiao, N.; Cao, X.; Li, Y.; et al. Circulating tumor cells in the pulmonary vein increase significantly after lobectomy: A prospective observational study. Thorac. Cancer 2019, 10, 163–169. [Google Scholar] [CrossRef]
- Balkwill, F.R.; Capasso, M.; Hagemann, T. The tumor microenvironment at a glance. J. Cell Sci. 2012, 125, 5591–5596. [Google Scholar] [CrossRef]
- Seifert, P.; Benedic, M.; Effert, P. Nerve fibers in tumors of the human urinary bladder. Virchows Arch. Int. J. Pathol. 2002, 440, 291–297. [Google Scholar] [CrossRef]
- Ayala, G.E.; Dai, H.; Powell, M.; Li, R.; Ding, Y.; Wheeler, T.M.; Shine, D.; Kadmon, D.; Thompson, T.; Miles, B.J.; et al. Cancer-related axonogenesis and neurogenesis in prostate cancer. Clin. Cancer Res. 2008, 14, 7593–7603. [Google Scholar] [CrossRef]
- Mravec, B.; Gidron, Y.; Hulin, I. Neurobiology of cancer: Interactions between nervous, endocrine and immune systems as a base for monitoring and modulating the tumorigenesis by the brain. Semin. Cancer Biol. 2008, 18, 150–163. [Google Scholar] [CrossRef]
- Seifert, P.; Spitznas, M. Axons in human choroidal melanoma suggest the participation of nerves in the control of these tumors. Am. J. Ophthalmol. 2002, 133, 711–713. [Google Scholar] [CrossRef]
- Magnon, C.; Hall, S.J.; Lin, J.; Xue, X.; Gerber, L.; Freedland, S.J.; Frenette, P.S. Autonomic nerve development contributes to prostate cancer progression. Science 2013, 341, 1236361. [Google Scholar] [CrossRef]
- Fernandez, E.V.; Price, D.K.; Figg, W.D. Prostate cancer progression attributed to autonomic nerve development: Potential for therapeutic prevention of localized and metastatic disease. Cancer Biol. Ther. 2013, 14, 1005–1006. [Google Scholar] [CrossRef] [PubMed]
- Keskinov, A.A.; Tapias, V.; Watkins, S.C.; Ma, Y.; Shurin, M.R.; Shurin, G.V. Impact of the Sensory Neurons on Melanoma Growth In Vivo. PLoS ONE 2016, 11, e0156095. [Google Scholar] [CrossRef] [PubMed]
- Bunimovich, Y.L.; Keskinov, A.A.; Shurin, G.V.; Shurin, M.R. Schwann cells: A new player in the tumor microenvironment. Cancer Immunol. Immunother. 2017, 66, 959–968. [Google Scholar] [CrossRef]
- Zhou, Y.; Shurin, G.V.; Zhong, H.; Bunimovich, Y.L.; Han, B.; Shurin, M.R. Schwann Cells Augment Cell Spreading and Metastasis of Lung Cancer. Cancer Res. 2018, 78, 5927–5939. [Google Scholar] [CrossRef]
- Martyn, G.V.; Shurin, G.V.; Keskinov, A.A.; Bunimovich, Y.L.; Shurin, M.R. Schwann cells shape the neuro-immune environs and control cancer progression. Cancer Immunol. Immunother. 2019, 68, 1819–1829. [Google Scholar] [CrossRef]
- Shurin, G.V.; Kruglov, O.; Ding, F.; Lin, Y.; Hao, X.; Keskinov, A.A.; You, Z.; Lokshin, A.E.; LaFramboise, W.A.; Falo, L.D., Jr.; et al. Melanoma-Induced Reprogramming of Schwann Cell Signaling Aids Tumor Growth. Cancer Res. 2019, 79, 2736–2747. [Google Scholar] [CrossRef] [PubMed]
- Shurin, M.R.; Shurin, G.V.; Zlotnikov, S.B.; Bunimovich, Y.L. The Neuroimmune Axis in the Tumor Microenvironment. J. Immunol. 2020, 204, 280–285. [Google Scholar] [CrossRef]
- Seifert, P.; Spitznas, M. Tumours may be innervated. Virchows Arch. Int. J. Pathol. 2001, 438, 228–231. [Google Scholar] [CrossRef]
- Ondicova, K.; Mravec, B. Role of nervous system in cancer aetiopathogenesis. Lancet. Oncol. 2010, 11, 596–601. [Google Scholar] [CrossRef]
- Li, S.; Sun, Y.; Gao, D. Role of the nervous system in cancer metastasis. Oncol. Lett. 2013, 5, 1101–1111. [Google Scholar] [CrossRef] [PubMed]
- Arese, M.; Bussolino, F.; Pergolizzi, M.; Bizzozero, L.; Pascal, D. Tumor progression: The neuronal input. Ann. Transl. Med. 2018, 6, 89. [Google Scholar] [CrossRef] [PubMed]
- Ceyhan, G.O.; Demir, I.E.; Altintas, B.; Rauch, U.; Thiel, G.; Müller, M.W.; Giese, N.A.; Friess, H.; Schäfer, K.-H. Neural invasion in pancreatic cancer: A mutual tropism between neurons and cancer cells. Biochem. Biophys. Res. Commun. 2008, 374, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Liebig, C.; Ayala, G.; Wilks, J.A.; Berger, D.H.; Albo, D. Perineural invasion in cancer. Cancer 2009, 115, 3379–3391. [Google Scholar] [CrossRef]
- Cole, S.W. Nervous system regulation of the cancer genome. Brain Behav. Immun. 2013, 30, S10–S18. [Google Scholar] [CrossRef] [PubMed]
- Jobling, P.; Pundavela, J.; Oliveira, S.M.; Roselli, S.; Walker, M.M.; Hondermarck, H. Nerve-Cancer Cell Cross-talk: A Novel Promoter of Tumor Progression. Cancer Res. 2015, 75, 1777–1781. [Google Scholar] [CrossRef]
- Rabben, H.L.; Zhao, C.M.; Hayakawa, Y.; Wang, T.C.; Chen, D. Vagotomy and Gastric Tumorigenesis. Curr. Neuropharmacol. 2016, 14, 967–972. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.M.; Hayakawa, Y.; Kodama, Y.; Muthupalani, S.; Westphalen, C.B.; Andersen, G.T.; Flatberg, A.; Johannessen, H.; Friedman, R.A.; Renz, B.W.; et al. Denervation suppresses gastric tumorigenesis. Sci. Transl. Med. 2014, 6, 250ra115. [Google Scholar] [CrossRef] [PubMed]
- Dolgin, E. Cancer–neuronal crosstalk and the startups working to silence it. Nat. Biotechnol. 2020, 38, 115–117. [Google Scholar] [CrossRef]
- Entschladen, F.; Drell, T.L.t.; Lang, K.; Joseph, J.; Zaenker, K.S. Tumour-cell migration, invasion, and metastasis: Navigation by neurotransmitters. Lancet. Oncol. 2004, 5, 254–258. [Google Scholar] [CrossRef]
- Sloan, E.K.; Priceman, S.J.; Cox, B.F.; Yu, S.; Pimentel, M.A.; Tangkanangnukul, V.; Arevalo, J.M.; Morizono, K.; Karanikolas, B.D.; Wu, L.; et al. The sympathetic nervous system induces a metastatic switch in primary breast cancer. Cancer Res. 2010, 70, 7042–7052. [Google Scholar] [CrossRef]
- Kim-Fuchs, C.; Le, C.P.; Pimentel, M.A.; Shackleford, D.; Ferrari, D.; Angst, E.; Hollande, F.; Sloan, E.K. Chronic stress accelerates pancreatic cancer growth and invasion: A critical role for beta-adrenergic signaling in the pancreatic microenvironment. Brain Behav. Immun. 2014, 40, 40–47. [Google Scholar] [CrossRef]
- Mancino, M.; Ametller, E.; Gascon, P.; Almendro, V. The neuronal influence on tumor progression. Biochim. Biophys. Acta 2011, 1816, 105–118. [Google Scholar] [CrossRef]
- Pimentel, M.; Chai, M.; Le, C.; Cole, S.; Sloan, E. Sympathetic nervous system regulation of metastasis. In Metastatic Cancer: Clinical and Biological Perspectives; Jandial, R., Ed.; Landes Bioscience: Austin, TX, USA, 2013; pp. 169–179. [Google Scholar]
- Amit, M.; Takahashi, H.; Dragomir, M.P.; Lindemann, A.; Gleber-Netto, F.O.; Pickering, C.R.; Anfossi, S.; Osman, A.A.; Cai, Y.; Wang, R.; et al. Loss of p53 drives neuron reprogramming in head and neck cancer. Nature 2020, 578, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Pouya, F.D.; Rasmi, Y.; Asl, E.R. Role of Neurotransmitters and Neuropeptides in Breast Cancer Metastasis. Biochem. (Mosc.) Suppl. Ser. A Membr. Cell Biol. 2020, 14, 107–116. [Google Scholar] [CrossRef]
- Tilan, J.; Kitlinska, J. Neuropeptide Y (NPY) in tumor growth and progression: Lessons learned from pediatric oncology. Neuropeptides 2016, 55, 55–66. [Google Scholar] [CrossRef]
- Rao, G.; Patel, P.S.; Idler, S.P.; Maloof, P.; Gascon, P.; Potian, J.A.; Rameshwar, P. Facilitating role of preprotachykinin-I gene in the integration of breast cancer cells within the stromal compartment of the bone marrow: A model of early cancer progression. Cancer Res. 2004, 64, 2874–2881. [Google Scholar] [CrossRef]
- Ohmori, H.; Fujii, K.; Sasahira, T.; Luo, Y.; Isobe, M.; Tatsumoto, N.; Kuniyasu, H. Methionine-enkephalin secreted by human colorectal cancer cells suppresses T lymphocytes. Cancer Sci. 2009, 100, 497–502. [Google Scholar] [CrossRef]
- Ishimaru, H.; Kageyama, Y.; Hayashi, T.; Nemoto, T.; Eishi, Y.; Kihara, K. Expression of matrix metalloproteinase-9 and bombesin/gastrin-releasing peptide in human prostate cancers and their lymph node metastases. Acta Oncol. 2002, 41, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Kuol, N.; Stojanovska, L.; Apostolopoulos, V.; Nurgali, K. Role of the nervous system in cancer metastasis. J. Exp. Clin. Cancer Res. 2018, 37, 5. [Google Scholar] [CrossRef]
- Alkire, B.C.; Raykar, N.P.; Shrime, M.G.; Weiser, T.G.; Bickler, S.W.; Rose, J.A.; Nutt, C.T.; Greenberg, S.L.; Kotagal, M.; Riesel, J.N.; et al. Global access to surgical care: A modelling study. Lancet Glob. Health 2015, 3, e316–e323. [Google Scholar] [CrossRef]
- Griffiths, J.D. The dissemination of cancer cells during operative procedures. Ann. R. Coll. Surg. Engl. 1960, 27, 14–44. [Google Scholar] [PubMed]
- Marie, P.; Clunet, J. Frequence des m6tastases visc6rale chez les souris cancereuses apr6s ablation chirurgicale de leur tumneur. Bulletin L’association Francaise Pour l’etude Cancer 1910, 3, 19–23. [Google Scholar]
- Clunet, J. Recherches Expdrimentales Sur Les Tumeurs Malignes; Steinheil: Paris, French, 1910. [Google Scholar]
- Ryall, C. The technique of cancer operations with reference to the danger of caneer infection. Br. Med. J. 1908, 2, 1005. [Google Scholar]
- Yamaguchi, K.; Takagi, Y.; Aoki, S.; Futamura, M.; Saji, S. Significant detection of circulating cancer cells in the blood by reverse transcriptase-polymerase chain reaction during colorectal cancer resection. Ann. Surg. 2000, 232, 58–65. [Google Scholar] [CrossRef]
- Ebenezer, G.J.; McArthur, J.C.; Thomas, D.; Murinson, B.; Hauer, P.; Polydefkis, M.; Griffin, J.W. Denervation of skin in neuropathies: The sequence of axonal and Schwann cell changes in skin biopsies. Brain 2007, 130, 2703–2714. [Google Scholar] [CrossRef] [PubMed]
- Abramovitch, R.; Marikovsky, M.; Meir, G.; Neeman, M. Stimulation of tumour growth by wound-derived growth factors. Br. J. Cancer 1999, 79, 1392–1398. [Google Scholar] [CrossRef] [PubMed]
- Skipper, D.; Jeffrey, M.J.; Cooper, A.J.; Alexander, P.; Taylor, I. Enhanced growth of tumour cells in healing colonic anastomoses and laparotomy wounds. Int. J. Colorectal Dis. 1989, 4, 172–177. [Google Scholar] [CrossRef]
- Jiang, W.G.; Puntis, M.C.; Hallett, M.B. Molecular and cellular basis of cancer invasion and metastasis: Implications for treatment. Br. J. Surg. 1994, 81, 1576–1590. [Google Scholar] [CrossRef]
- Svendsen, M.N.; Werther, K.; Nielsen, H.J.; Kristjansen, P.E. VEGF and tumour angiogenesis. Impact of surgery, wound healing, inflammation and blood transfusion. Scand. J. Gastroenterol. 2002, 37, 373–379. [Google Scholar] [CrossRef]
- Berguer, R.; Bravo, N.; Bowyer, M.; Egan, C.; Knolmayer, T.; Ferrick, D. Major surgery suppresses maximal production of helper T-cell type 1 cytokines without potentiating the release of helper T-cell type 2 cytokines. Arch. Surg. 1999, 134, 540–544. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Herbert, T.B. Health psychology: Psychological factors and physical disease from the perspective of human psychoneuroimmunology. Annu. Rev. Psychol. 1996, 47, 113–142. [Google Scholar] [CrossRef]
- Moynihan, J.A.; Ader, R. Psychoneuroimmunology: Animal models of disease. Psychosom. Med. 1996, 58, 546–558. [Google Scholar] [CrossRef]
- Neeman, E.; Ben-Eliyahu, S. Surgery and stress promote cancer metastasis: New outlooks on perioperative mediating mechanisms and immune involvement. Brain Behav. Immun. 2013, 30, S32–S40. [Google Scholar] [CrossRef] [PubMed]
- Ceelen, W.; Pattyn, P.; Mareel, M. Surgery, wound healing, and metastasis: Recent insights and clinical implications. Crit. Rev. Oncol. Hematol. 2014, 89, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Coffey, J.C.; Wang, J.H.; Smith, M.J.; Bouchier-Hayes, D.; Cotter, T.G.; Redmond, H.P. Excisional surgery for cancer cure: Therapy at a cost. Lancet Oncol. 2003, 4, 760–768. [Google Scholar] [CrossRef]
- Nan Tie, E.; Henderson, M.A.; Gyorki, D.E. Management of in-transit melanoma metastases: A review. ANZ J. Surg. 2019, 89, 647–652. [Google Scholar] [CrossRef]
- Blakely, A.M.; Comissiong, D.S.; Vezeridis, M.P.; Miner, T.J. Suboptimal Compliance with National Comprehensive Cancer Network Melanoma Guidelines: Who Is at Risk? Am. J. Clin. Oncol. 2018, 41, 754–759. [Google Scholar] [CrossRef]
- Mangold, A.R.; Skinner, R.; Dueck, A.C.; Sekulic, A.; Pockaj, B.A. Risk Factors Predicting Positive Margins at Primary Wide Local Excision of Cutaneous Melanoma. Dermatol. Surg. 2016, 42, 646–652. [Google Scholar] [CrossRef]
- Mathew, G.; Watson, D.I.; Rofe, A.M.; Baigrie, C.F.; Ellis, T.; Jamieson, G.G. Wound metastases following laparoscopic and open surgery for abdominal cancer in a rat model. Br. J. Surg. 1996, 83, 1087–1090. [Google Scholar] [CrossRef] [PubMed]
- Spelt, L.; Andersson, B.; Nilsson, J.; Andersson, R. Prognostic models for outcome following liver resection for colorectal cancer metastases: A systematic review. Eur. J. Surg. Oncol. 2012, 38, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Demicheli, R.; Dillekas, H.; Straume, O.; Biganzoli, E. Distant metastasis dynamics following subsequent surgeries after primary breast cancer removal. Breast Cancer Res. 2019, 21, 57. [Google Scholar] [CrossRef] [PubMed]
- Qadri, S.S.; Wang, J.H.; Coffey, J.C.; Alam, M.; O’Donnell, A.; Aherne, T.; Redmond, H.P. Can surgery for cancer accelerate the progression of secondary tumors within residual minimal disease at both local and systemic levels? Ann. Thorac. Surg. 2005, 80, 1046–1050; discussion 1041–1050. [Google Scholar] [CrossRef] [PubMed]
- Mehlen, P.; Puisieux, A. Metastasis: A question of life or death. Nat. Rev. Cancer 2006, 6, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, T.; Goto, T. Links between Inflammation and Postoperative Cancer Recurrence. J. Clin. Med. 2021, 10, 228. [Google Scholar] [CrossRef]
- Antonio, N.; Bonnelykke-Behrndtz, M.L.; Ward, L.C.; Collin, J.; Christensen, I.J.; Steiniche, T.; Schmidt, H.; Feng, Y.; Martin, P. The wound inflammatory response exacerbates growth of pre-neoplastic cells and progression to cancer. EMBO J. 2015, 34, 2219–2236. [Google Scholar] [CrossRef]
- Murthy, S.M.; Goldschmidt, R.A.; Rao, L.N.; Ammirati, M.; Buchmann, T.; Scanlon, E.F. The influence of surgical trauma on experimental metastasis. Cancer 1989, 64, 2035–2044. [Google Scholar] [CrossRef]
- Wang, J.; Yang, L.; Yu, L.; Wang, Y.Y.; Chen, R.; Qian, J.; Hong, Z.P.; Su, X.S. Surgery-induced monocytic myeloid-derived suppressor cells expand regulatory T cells in lung cancer. Oncotarget 2017, 8, 17050–17058. [Google Scholar] [CrossRef]
- Tai, L.H.; Zhang, J.; Auer, R.C. Preventing surgery-induced NK cell dysfunction and cancer metastases with influenza vaccination. Oncoimmunology 2013, 2, e26618. [Google Scholar] [CrossRef]
- Tai, L.H.; de Souza, C.T.; Bélanger, S.; Ly, L.; Alkayyal, A.A.; Zhang, J.; Rintoul, J.L.; Ananth, A.A.; Lam, T.; Breitbach, C.J.; et al. Preventing postoperative metastatic disease by inhibiting surgery-induced dysfunction in natural killer cells. Cancer Res. 2013, 73, 97–107. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, D.P.; O’Leary, E.; Foley, N.; Cotter, T.G.; Wang, J.H.; Redmond, H.P. Effects of surgery on the cancer stem cell niche. Eur. J. Surg. oncol. 2016, 42, 319–325. [Google Scholar] [CrossRef]
- Hiller, J.G.; Perry, N.J.; Poulogiannis, G.; Riedel, B.; Sloan, E.K. Perioperative events influence cancer recurrence risk after surgery. Nature reviews. Clin. Oncol. 2018, 15, 205–218. [Google Scholar] [CrossRef]
- Sullivan, R.; Peppercorn, J.; Sikora, K.; Zalcberg, J.; Meropol, N.J.; Amir, E.; Khayat, D.; Boyle, P.; Autier, P.; Tannock, I.F.; et al. Delivering affordable cancer care in high-income countries. Lancet Oncol. 2011, 12, 933–980. [Google Scholar] [CrossRef]
- Chéret, J.; Lebonvallet, N.; Buhé, V.; Carre, J.L.; Misery, L.; Le Gall-Ianotto, C. Influence of sensory neuropeptides on human cutaneous wound healing process. J. Dermatol. Sci. 2014, 74, 193–203. [Google Scholar] [CrossRef]
- Blais, M.; Mottier, L.; Germain, M.A.; Bellenfant, S.; Cadau, S.; Berthod, F. Sensory neurons accelerate skin reepithelialization via substance P in an innervated tissue-engineered wound healing model. Tissue Eng. Part A 2014, 20, 2180–2188. [Google Scholar] [CrossRef] [PubMed]
- Ricon, I.; Hanalis-Miller, T.; Haldar, R.; Jacoby, R.; Ben-Eliyahu, S. Perioperative biobehavioral interventions to prevent cancer recurrence through combined inhibition of beta-adrenergic and cyclooxygenase 2 signaling. Cancer 2019, 125, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, J.M.; McIver, C.M.; Stephenson, S.A.; Hewett, P.J.; Rieger, N.; Hardingham, J.E. Identification of early-stage colorectal cancer patients at risk of relapse post-resection by immunobead reverse transcription-PCR analysis of peritoneal lavage fluid for malignant cells. Clin. Cancer Res. 2006, 12, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, P.; Xu, Y.; Yan, J.; Liu, Z.; Lau, W.B.; Lau, B.; Li, Y.; Zhao, X.; Wei, Y.; et al. Surgical stress and cancer progression: The twisted tango. Mol. Cancer 2019, 18, 132. [Google Scholar] [CrossRef]
- Brown, D.C.; Purushotham, A.D.; Birnie, G.D.; George, W.D. Detection of intraoperative tumor cell dissemination in patients with breast cancer by use of reverse transcription and polymerase chain reaction. Surgery 1995, 117, 95–101. [Google Scholar] [CrossRef]
- Retsky, M.W.; Demicheli, R.; Hrushesky, W.J.; Baum, M.; Gukas, I.D. Dormancy and surgery-driven escape from dormancy help explain some clinical features of breast cancer. APMIS 2008, 116, 730–741. [Google Scholar] [CrossRef]
- Baum, M.; Demicheli, R.; Hrushesky, W.; Retsky, M. Does surgery unfavourably perturb the "natural history" of early breast cancer by accelerating the appearance of distant metastases? Eur. J. Cancer 2005, 41, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Kim, R. Effects of surgery and anesthetic choice on immunosuppression and cancer recurrence. J. Transl. Med. 2018, 16, 8. [Google Scholar] [CrossRef]
- Sood, A.K.; Bhatty, R.; Kamat, A.A.; Landen, C.N.; Han, L.; Thaker, P.H.; Li, Y.; Gershenson, D.M.; Lutgendorf, S.; Cole, S.W. Stress hormone-mediated invasion of ovarian cancer cells. Clin. Cancer Res. 2006, 12, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.V.; Kim, S.J.; Donovan, E.L.; Chen, M.; Gross, A.C.; Webster Marketon, J.I.; Barsky, S.H.; Glaser, R. Norepinephrine upregulates VEGF, IL-8, and IL-6 expression in human melanoma tumor cell lines: Implications for stress-related enhancement of tumor progression. Brain Behav. Immun. 2009, 23, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.Y.; Whited, J.L. Parallels between wound healing, epimorphic regeneration and solid tumors. Development 2020, 147, dev181636. [Google Scholar] [CrossRef]
- Conforti, L.; Gilley, J.; Coleman, M.P. Wallerian degeneration: An emerging axon death pathway linking injury and disease. Nat. Rev. Neurosci. 2014, 15, 394–409. [Google Scholar] [CrossRef]
- Gaudet, A.D.; Popovich, P.G.; Ramer, M.S. Wallerian degeneration: Gaining perspective on inflammatory events after peripheral nerve injury. J. Neuroinflamm. 2011, 8, 110. [Google Scholar] [CrossRef] [PubMed]
- Whalley, K. Glia: Schwann cells provide life support for axons. Nat. Rev. Neurosci. 2014, 15, 698–699. [Google Scholar] [CrossRef]
- Stratton, J.A.; Shah, P.T. Macrophage polarization in nerve injury: Do Schwann cells play a role? Neural Regen. Res. 2016, 11, 53–57. [Google Scholar] [CrossRef]
- Wong, K.M.; Babetto, E.; Beirowski, B. Axon degeneration: Make the Schwann cell great again. Neural Regen. Res. 2017, 12, 518–524. [Google Scholar] [CrossRef]
- Qu, W.R.; Zhu, Z.; Liu, J.; Song, D.B.; Tian, H.; Chen, B.P.; Li, R.; Deng, L.X. Interaction between Schwann cells and other cells during repair of peripheral nerve injury. Neural Regen. Res. 2021, 16, 93–98. [Google Scholar] [CrossRef]
- Parfejevs, V.; Antunes, A.T.; Sommer, L. Injury and stress responses of adult neural crest-derived cells. Dev. Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Parfejevs, V.; Debbache, J.; Shakhova, O.; Schaefer, S.M.; Glausch, M.; Wegner, M.; Suter, U.; Riekstina, U.; Werner, S.; Sommer, L. Injury-activated glial cells promote wound healing of the adult skin in mice. Nat. Commun. 2018, 9, 236. [Google Scholar] [CrossRef]
- Johnston, A.P. Schwann cells: An emerging player in tissue regeneration. Stem Cell Investig. 2017, 4, 14. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, S.H.; Shurin, G.V.; Khosravi, H.; Kazi, R.; Kruglov, O.; Shurin, M.R.; Bunimovich, Y.L. Immunomodulation by Schwann cells in disease. Cancer Immunol. Immunother. 2020, 69, 245–253. [Google Scholar] [CrossRef]
- Sroka, I.C.; Chopra, H.; Das, L.; Gard, J.M.; Nagle, R.B.; Cress, A.E. Schwann Cells Increase Prostate and Pancreatic Tumor Cell Invasion Using Laminin Binding A6 Integrin. J. Cell. Biochem. 2016, 117, 491–499. [Google Scholar] [CrossRef]
- Deborde, S.; Omelchenko, T.; Lyubchik, A.; Zhou, Y.; He, S.; McNamara, W.F.; Chernichenko, N.; Lee, S.Y.; Barajas, F.; Chen, C.H.; et al. Schwann cells induce cancer cell dispersion and invasion. J. Clin. Investig. 2016, 126, 1538–1554. [Google Scholar] [CrossRef] [PubMed]
- Peeters, C.F.; de Waal, R.M.; Wobbes, T.; Ruers, T.J. Metastatic dormancy imposed by the primary tumor: Does it exist in humans? Ann. Surg. Oncol. 2008, 15, 3308–3315. [Google Scholar] [CrossRef] [PubMed]
- Di Gianni, P.; Franco, M.; Meiss, R.P.; Vanzulli, S.; Piazzon, I.; Pasqualini, C.D.; Bustuoabad, O.D.; Ruggiero, R.A. Inhibition of metastases by a serum factor associated to concomitant resistance induced by unrelated murine tumors. Oncol. Rep. 1999, 6, 1073–1084. [Google Scholar] [CrossRef]
- Bonfil, R.D.; Ruggiero, R.A.; Bustuoabad, O.D.; Meiss, R.P.; Pasqualini, C.D. Role of concomitant resistance in the development of murine lung metastases. Int. J. Cancer 1988, 41, 415–422. [Google Scholar] [CrossRef]
- Day, S.; Myers, W.; Stansly, P.; Garattini, S.; Lewis, M.; Sugarbaker, E.; Thornthwaite, J.; Ketcham, A. Inhibitory effect of a primary tumor on metastasis. In Cancer Invasion and Metastasis. Biological Mechanisms and Therapy; Raven Press: New York, NY, USA, 1977; pp. 227–240. [Google Scholar]
- NCRAS. Chemotherapy, Radiotherapy and Surgical Tumour Resections in England; The National Cancer Registration and Analysis Service: London, UK, 2020.
- Alieva, M.; van Rheenen, J.; Broekman, M.L.D. Potential impact of invasive surgical procedures on primary tumor growth and metastasis. Clin. Exp. Metastasis 2018, 35, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Sorski, L.; Melamed, R.; Levi, B.; Matzner, P.; Lavon, H.; Rosenne, E.; Shaashua, L.; Ricon, I.; Sandbank, E.; Benbenishty, A.; et al. Prevention of liver metastases through perioperative acute CpG-C immune stimulation. Cancer Immunol. Immunother. 2020, 69, 2021–2031. [Google Scholar] [CrossRef]
- Melhem-Bertrandt, A.; Chavez-Macgregor, M.; Lei, X.; Brown, E.N.; Lee, R.T.; Meric-Bernstam, F.; Sood, A.K.; Conzen, S.D.; Hortobagyi, G.N.; Gonzalez-Angulo, A.M. Beta-blocker use is associated with improved relapse-free survival in patients with triple-negative breast cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2011, 29, 2645–2652. [Google Scholar] [CrossRef] [PubMed]
- Forget, P.; Vandenhende, J.; Berliere, M.; Machiels, J.P.; Nussbaum, B.; Legrand, C.; De Kock, M. Do intraoperative analgesics influence breast cancer recurrence after mastectomy? A retrospective analysis. Anesth. Analg. 2010, 110, 1630–1635. [Google Scholar] [CrossRef] [PubMed]
- Krall, J.A.; Reinhardt, F.; Mercury, O.A.; Pattabiraman, D.R.; Brooks, M.W.; Dougan, M.; Lambert, A.W.; Bierie, B.; Ploegh, H.L.; Dougan, S.K.; et al. The systemic response to surgery triggers the outgrowth of distant immune-controlled tumors in mouse models of dormancy. Sci. Transl. Med. 2018, 10, eaan3464. [Google Scholar] [CrossRef]
- Hiller, J.G.; Cole, S.W.; Crone, E.M.; Byrne, D.J.; Shackleford, D.M.; Pang, J.B.; Henderson, M.A.; Nightingale, S.S.; Ho, K.M.; Myles, P.S.; et al. Preoperative beta-Blockade with Propranolol Reduces Biomarkers of Metastasis in Breast Cancer: A Phase II Randomized Trial. Clin. Cancer Res. 2020, 26, 1803–1811. [Google Scholar] [CrossRef] [PubMed]
- Choy, C.; Raytis, J.L.; Smith, D.D.; Duenas, M.; Neman, J.; Jandial, R.; Lew, M.W. Inhibition of β2-adrenergic receptor reduces triple-negative breast cancer brain metastases: The potential benefit of perioperative β-blockade. Oncol. Rep. 2016, 35, 3135–3142. [Google Scholar] [CrossRef]
- Sorski, L.; Melamed, R.; Matzner, P.; Lavon, H.; Shaashua, L.; Rosenne, E.; Ben-Eliyahu, S. Reducing liver metastases of colon cancer in the context of extensive and minor surgeries through β-adrenoceptors blockade and COX2 inhibition. Brain Behav. Immun. 2016, 58, 91–98. [Google Scholar] [CrossRef]
- Tai, L.H.; Alkayyal, A.A.; Leslie, A.L.; Sahi, S.; Bennett, S.; Tanese de Souza, C.; Baxter, K.; Angka, L.; Xu, R.; Kennedy, M.A.; et al. Phosphodiesterase-5 inhibition reduces postoperative metastatic disease by targeting surgery-induced myeloid derived suppressor cell-dependent inhibition of Natural Killer cell cytotoxicity. Oncoimmunology 2018, 7, e1431082. [Google Scholar] [CrossRef]


{kind=link}
| Cancer | Species | Notes | Year | Reference |
|---|---|---|---|---|
| different | human | surgery for primary tumors | 1907 | [25] |
| carcinoma | mouse | removal of s.c. tumors | 1913 | [26] |
| carcinoma | mouse | localization of tumors at points of injury | 1914 | [27] |
| melanoma, sarcoma | mouse | amputation of the tumor-bearing leg | 1958 | [28] |
| sarcoma | mouse | pulmonary metastases after amputation of the limb bearing the tumor | 1958 | [29] |
| carcinoma of the gastric stump | human | appearance of a second primary lesion after resection for malignancy | 1973 | [30] |
| lung carcinoma | mouse | noncurative excision | 1976 | [3] |
| testicular cancer | human | cytoreductive surgery | 1980 | [31] |
| ovarian cancer | human | cytoreductive surgery | 1980 | [32] |
| colon cancer | human | abdominal wall recurrence after colectomy | 1983 | [33] |
| melanoma, sarcoma | mouse | tumor cell growth at the surgical wound site | 1989 | [34] |
| ovarian cancer | human | cytoreductive surgery | 1989 | [35] |
| dermal fibrosarcoma | mouse | spontaneous tumors at abnormal wound repair sites | 1990 | [36] |
| melanoma | mice | local tumor removal | 1992 | [37] |
| colon cancer | human | recurrence after colectomy | 1994 | [38] |
| mammary adenocarcinoma | mouse | tumor injection after surgery, frequency of tumor formation at the site of bone wound | 1994 | [39] |
| gastric cancer | human | abdominal wall metastasis after laparoscopic gastroenterostomy | 1995 | [40] |
| NSCLC | human | potentially curative resection | 1996 | [41] |
| melanoma | mouse | tumor growth in adjacent subcutaneous tissue next to a surgical wound | 1998 | [42] |
| breast cancer | human | radical mastectomy | 1999 | [43] |
| mammary carcinoma | mouse | lung metastases after open surgery | 1999 | [44] |
| lung cancer | human | surgery | 2000 | [45] |
| breast cancer | human | mastectomy | 2001 | [11] |
| melanoma | mouse | surgery and immunostimulation | 2001 | [46] |
| lung carcinoma | mouse | surgical tumor resection | 2001 | [47] |
| breast cancer | human | local recurrence in the core needle biopsy site | 2001 | [18] |
| colon carcinoma | mouse | lung metastasis after 5 kinds of surgical stress of different degree | 2003 | [48] |
| breast cancer | human | needle core biopsy and the incidence of sentinel node metastases | 2004 | [19] |
| colorectal cancer | human | local recurrences after curative surgery | 2005 | [14] |
| ovarian carcinoma | mouse | laparotomy and mastectomy as surgical stress | 2009 | [49] |
| colorectal cancer | human | resection of the primary tumor | 2009 | [10] |
| bladder carcinoma | human | surgical removal and rapid metastatic progression | 2009 | [15] |
| mammary adenocarcinoma | mouse | resection of primary tumor | 2010 | [50] |
| breast cancer | human | surgery | 2010 | [9] |
| NSCLC | human | surgery for early-stage cancer | 2013 | [16,17] |
| cutaneous melanoma | human | excisional biopsy and local recurrences | 2014 | [51] |
| mammary adenocarcinoma | mouse | distant metastases after core needle biopsy | 2014 | [51] |
| melanoma | human | sentinel lymph node biopsy and disease recurrence (distant metastases) | 2016 | [13] |
| colorectal cancer | human | surgery, partial hepatectomy | 2017 | [20] |
| pancreatic ductal adenocarcinoma | human | resection of primary tumor | 2018 | [21] |
| colorectal cancer | human | colorectal liver metastases after colorectal surgery | 2018 | [52] |
| gastric cancer | human | circulating tumor cells after resection | 2018 | [53] |
| hepatocellular carcinoma | human | circulating tumor cells after radical surgery | 2018 | [23] |
| lung cancer | human | circulating tumor cells after resection of a lung lobe | 2019 | [8] |
| gastric cancer | human | liver metastases after gastrectomy | 2020 | [54] |
| pancreatic acinar cell carcinoma | human | gastric and lymph node metastases after pancreatoduodenectomy | 2020 | [55] |
| squamous cell carcinoma of tongue | mouse | surgery induces hypoxia, CD11b+ cell infiltration and lymph node metastasis | 2020 | [56] |
| colon cancer | mouse | Immunotherapy decreases surgery-induced liver metastases | 2020 | [57] |
| papillary thyroid cancer | human | recurrence in subcutaneous area and lymph node metastasis after core needle biopsy | 2021 | [58] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shurin, M.R.; Baraldi, J.H.; Shurin, G.V. Neuroimmune Regulation of Surgery-Associated Metastases. Cells 2021, 10, 454. https://doi.org/10.3390/cells10020454
Shurin MR, Baraldi JH, Shurin GV. Neuroimmune Regulation of Surgery-Associated Metastases. Cells. 2021; 10(2):454. https://doi.org/10.3390/cells10020454
Chicago/Turabian StyleShurin, Michael R., James H. Baraldi, and Galina V. Shurin. 2021. "Neuroimmune Regulation of Surgery-Associated Metastases" Cells 10, no. 2: 454. https://doi.org/10.3390/cells10020454
APA StyleShurin, M. R., Baraldi, J. H., & Shurin, G. V. (2021). Neuroimmune Regulation of Surgery-Associated Metastases. Cells, 10(2), 454. https://doi.org/10.3390/cells10020454