
Human Placenta Buffers the Fetus from Adverse Effects of Perceived Maternal Stress


 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Questionnaire-Based Measures
3. Biological Measures
3.1. Collection of Biospecimens
3.2. Analysis of Steroid Metabolites in Placenta and Cord Blood Using Ultra-Pure Liquid Chromatography (UPLC) Mass Spectrometry (MS)
3.3. Statistical Analysis
4. Results
4.1. Study Population Characteristics and Steroid Profiles in Maternal and Fetal Samples
4.2. Maternal or Fetal Cortisol and Cortisone Do Not Correlate with the Maternal PSS
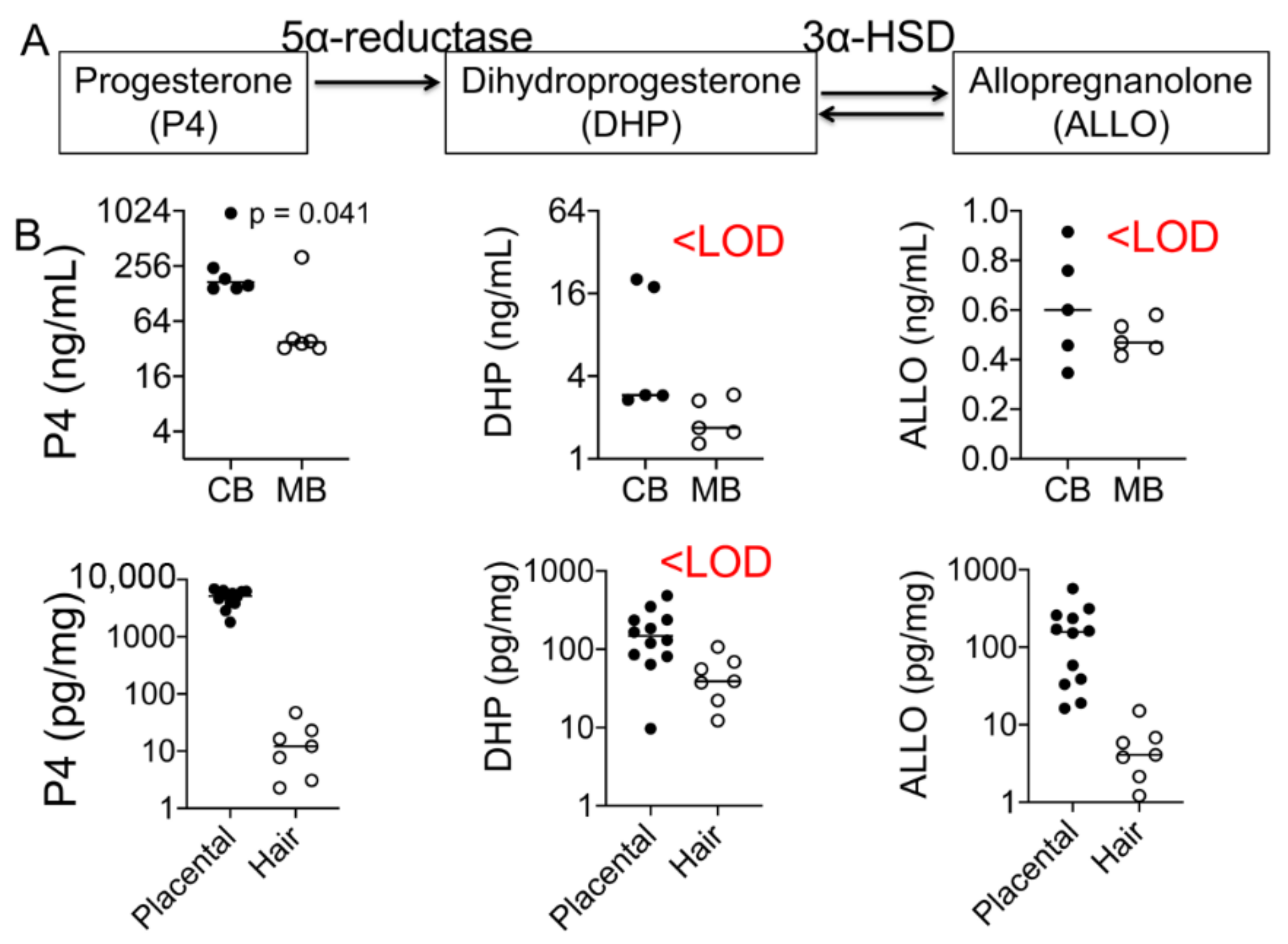
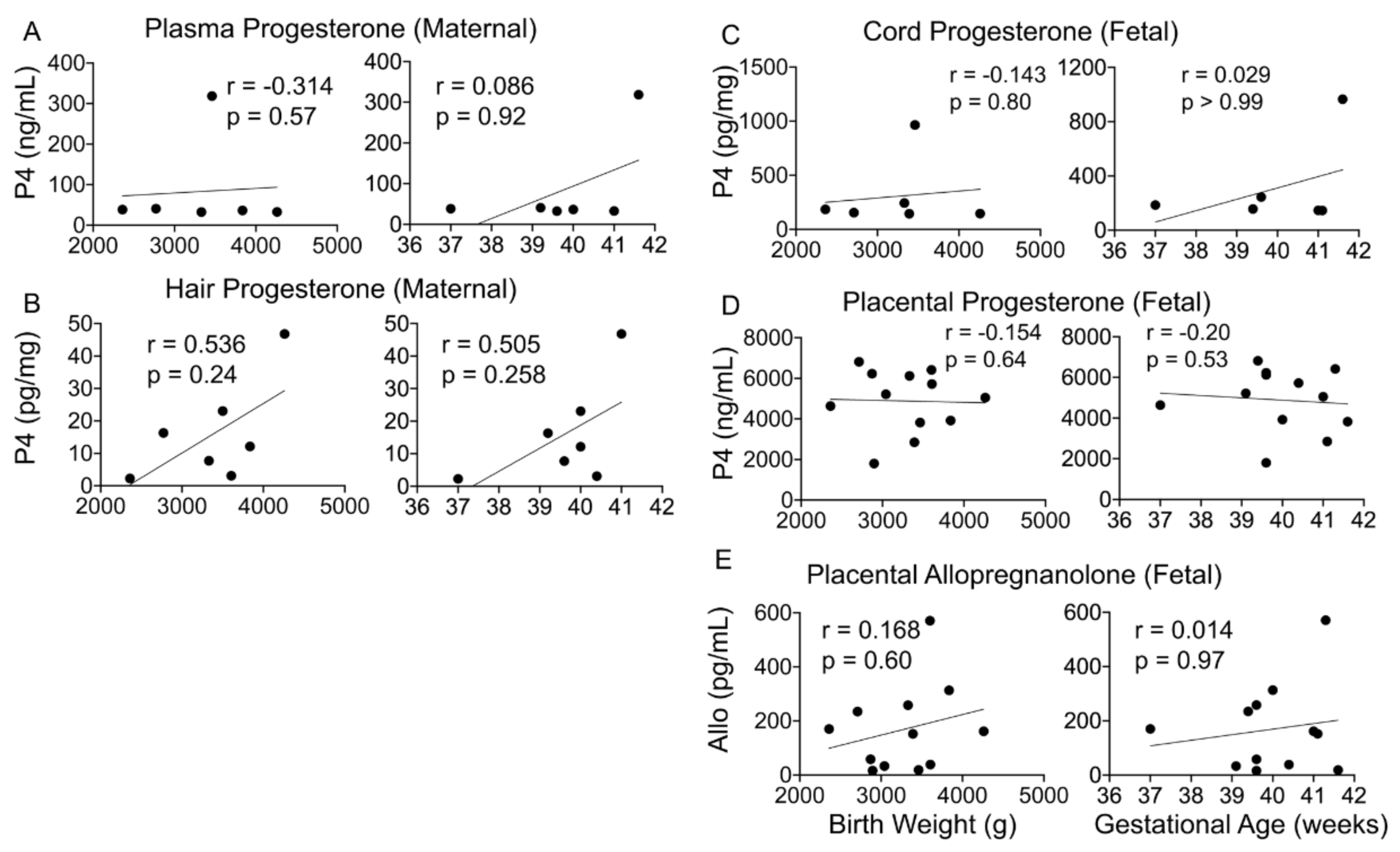
4.3. Progesterone and Its Metabolite Levels in Fetal and Maternal Samples
4.4. Maternal or Fetal Progesterone and ALLO Levels Do Not Associate with Birth Outcomes
4.5. Hair Progesterone Levels Positively Correlate with the Maternal Perceived Stress Score (PSS)
4.6. Maternal Dehydroepiandrosterone Sulfate (DHEAS) Correlates with the Perceived Stress Score
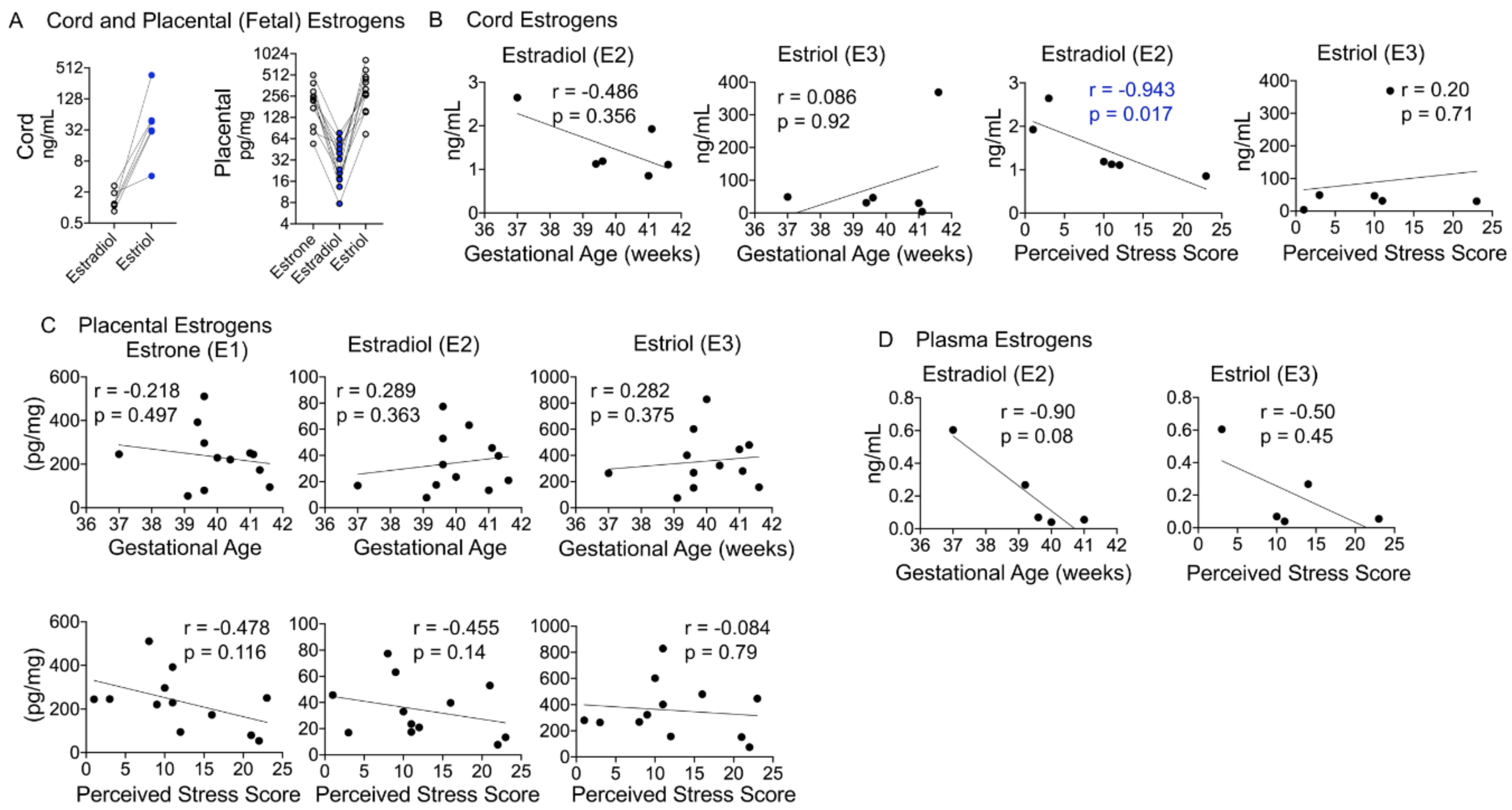
4.7. Fetal Estradiol Correlates with the Perceived Stress Score
5. Discussion
5.1. Relationships among Maternal and Fetal Hormones
5.2. Glucocorticoid Metabolites as Markers of Stress in a Healthy Pregnancy
5.3. Progesterone and Its Metabolites Correlate with Perceived Stress in a Healthy Pregnancy
5.4. Progesterone and Its Metabolites as Predictors of Birth Outcomes
5.5. Estrogen Metabolites and Their Relationship with Birth Outcomes and Maternal Stress
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ding, X.X.; Wu, Y.L.; Xu, S.J.; Zhu, R.P.; Jia, X.M.; Zhang, S.F.; Huang, K.; Zhu, P.; Hao, J.H.; Tao, F.B. Maternal anxiety during pregnancy and adverse birth outcomes: A systematic review and meta-analysis of prospective cohort studies. J. Affect. Disord. 2014, 159, 103–110. [Google Scholar] [CrossRef]
- Khashan, A.S.; McNamee, R.; Abel, K.M.; Mortensen, P.B.; Kenny, L.C.; Pedersen, M.G.; Webb, R.T.; Baker, P.N. Rates of preterm birth following antenatal maternal exposure to severe life events: A population-based cohort study. Hum. Reprod. 2009, 24, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Khashan, A.S.; McNamee, R.; Abel, K.M.; Pedersen, M.G.; Webb, R.T.; Kenny, L.C.; Mortensen, P.B.; Baker, P.N. Reduced infant birthweight consequent upon maternal exposure to severe life events. Psychosom. Med. 2008, 70, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. What is the placenta? Am. J. Obstet. Gynecol. 2015, 213, S6.e1–S6.e4. [Google Scholar] [CrossRef] [PubMed]
- Maltepe, E.; Fisher, S.J. Placenta: The Forgotten Organ. Annu. Rev. Cell Dev. Biol. 2015, 31, 523–552. [Google Scholar] [CrossRef]
- Pepe, G.; Albrecht, E. Steroid endocrinology of pregnancy. In Gynecology and Obstetrics; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2008; Volume 5. [Google Scholar]
- Austin, M.P.; Leader, L. Maternal stress and obstetric and infant outcomes: Epidemiological findings and neuroendocrine mechanisms. Aust. N. Z. J. Obstet. Gynaecol. 2000, 40, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Wadhwa, P.D.; Entringer, S.; Buss, C.; Lu, M.C. The contribution of maternal stress to preterm birth: Issues and considerations. Clin. Perinatol. 2011, 38, 351–384. [Google Scholar] [CrossRef]
- Brown, L.D.; Hay, W.W., Jr. Impact of placental insufficiency on fetal skeletal muscle growth. Mol. Cell Endocrinol. 2016, 435, 69–77. [Google Scholar] [CrossRef]
- Catov, J.M.; Peng, Y.; Scifres, C.M.; Parks, W.T. Placental pathology measures: Can they be rapidly and reliably integrated into large-scale perinatal studies? Placenta 2015, 36, 687–692. [Google Scholar] [CrossRef]
- Derricott, H.; Jones, R.L.; Heazell, A.E. Investigating the association of villitis of unknown etiology with stillbirth and fetal growth restriction—A systematic review. Placenta 2013, 34, 856–862. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, X.; Rong, C.; Rui, C.; Ji, H.; Qian, Y.J.; Jia, R.; Sun, L. Distinct DNA methylomes of human placentas between pre-eclampsia and gestational diabetes mellitus. Cell Physiol. Biochem. 2014, 34, 1877–1889. [Google Scholar] [CrossRef]
- Rademacher, T.W.; Gumaa, K.; Scioscia, M. Preeclampsia, insulin signalling and immunological dysfunction: A fetal, maternal or placental disorder? J. Reprod. Immunol. 2007, 76, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Uuskula, L.; Mannik, J.; Rull, K.; Minajeva, A.; Koks, S.; Vaas, P.; Teesalu, P.; Reimand, J.; Laan, M. Mid-gestational gene expression profile in placenta and link to pregnancy complications. PLoS ONE 2012, 7, e49248. [Google Scholar] [CrossRef] [PubMed]
- Veerapen, M.K.; Pelaez, L.; Potter, J.E.; Duthely, L.; Birusingh, R.; Rampersaud, E.; Bodamer, O.A.; Rodriguez, M.M. Bridging the gaps between the histopathological and demographic risk factors of preterm birth in a unique Miami inner-city population. Fetal Pediatr. Pathol. 2014, 33, 226–233. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Burns, E.R.; Farr, S.L.; Howards, P.P.; Centers for Disease Control and Prevention (CDC). Stressful life events experienced by women in the year before their infants’ births—United States, 2000–2010. Morb. Mortal. Wkly. Rep. 2015, 64, 247–251. [Google Scholar]
- Ghaemmaghami, P.; Dainese, S.M.; La Marca, R.; Zimmermann, R.; Ehlert, U. The association between the acute psychobiological stress response in second trimester pregnant women, amniotic fluid glucocorticoids, and neonatal birth outcome. Dev. Psychobiol. 2014, 56, 734–747. [Google Scholar] [CrossRef]
- Richoux, S.; Weiss, S. Women’s perceived stress and stress hormones during pregnancy: Are they related to one another? Presentation at the Annual Conference of Sigma Theta Tau International, Melbourne, Australia, 19–23 July 2018. [Google Scholar]
- Bolten, M.I.; Wurmser, H.; Buske-Kirschbaum, A.; Papousek, M.; Pirke, K.M.; Hellhammer, D. Cortisol levels in pregnancy as a psychobiological predictor for birth weight. Arch. Womens Ment. Health 2011, 14, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Goedhart, G.; Vrijkotte, T.G.; Roseboom, T.J.; van der Wal, M.F.; Cuijpers, P.; Bonsel, G.J. Maternal cortisol and offspring birthweight: Results from a large prospective cohort study. Psychoneuroendocrinology 2010, 35, 644–652. [Google Scholar] [CrossRef]
- Sze, Y.; Gill, A.C.; Brunton, P.J. Sex-dependent changes in neuroactive steroid concentrations in the rat brain following acute swim stress. J. Neuroendocrinol. 2018, 30, e12644. [Google Scholar] [CrossRef]
- Wilsterman, K.; Gotlieb, N.; Kriegsfeld, L.J.; Bentley, G.E. Pregnancy stage determines the effect of chronic stress on ovarian progesterone synthesis. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E987–E994. [Google Scholar] [CrossRef]
- Tulchinsky, D.; Hobel, C.J. Plasma human chorionic gonadotropin, estrone, estradiol, estriol, progesterone, and 17 alpha-hydroxyprogesterone in human pregnancy. 3. Early normal pregnancy. Am. J. Obstet. Gynecol. 1973, 117, 884–893. [Google Scholar] [CrossRef]
- Tal, R.; Taylor, H.S.; Burney, R.O.; Mooney, S.B.; Giudice, L.C. Endocrinology of Pregnancy. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., Dungan, K., Grossman, A., Hershman, J.M., Kaltsas, G., Koch, C., Kopp, P., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Csapo, A.I.; Pulkkinen, M.O.; Kaihola, H.L. The relationship between the timing of luteectomy and the incidence of complete abortions. Am. J. Obstet. Gynecol. 1974, 118, 985–989. [Google Scholar] [CrossRef]
- Sauer, M.V.; Paulson, R.J.; Lobo, R.A. A preliminary report on oocyte donation extending reproductive potential to women over 40. N. Engl. J. Med. 1990, 323, 1157–1160. [Google Scholar] [CrossRef]
- Brunton, P.J.; Russell, J.A.; Hirst, J.J. Allopregnanolone in the brain: Protecting pregnancy and birth outcomes. Prog. Neurobiol. 2014, 113, 106–136. [Google Scholar] [CrossRef]
- Schumacher, M.; Mattern, C.; Ghoumari, A.; Oudinet, J.P.; Liere, P.; Labombarda, F.; Sitruk-Ware, R.; De Nicola, A.F.; Guennoun, R. Revisiting the roles of progesterone and allopregnanolone in the nervous system: Resurgence of the progesterone receptors. Prog. Neurobiol. 2014, 113, 6–39. [Google Scholar] [CrossRef] [PubMed]
- Freitas-de-Melo, A.; Damian, J.P.; Hotzel, M.J.; Banchero, G.; Ungerfeld, R. Progesterone pretreatment increases the stress response to social isolation in ewes. Hormones 2016, 15, 81–87. [Google Scholar] [CrossRef]
- Park, M.H.; Rehman, S.U.; Kim, I.S.; Choi, M.S.; Yoo, H.H. Stress-induced changes of neurosteroid profiles in rat brain and plasma under immobilized condition. J. Pharm. Biomed. Anal. 2017, 138, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Pepe, G.J.; Albrecht, E.D. Actions of placental and fetal adrenal steroid hormones in primate pregnancy. Endocr. Rev. 1995, 16, 608–648. [Google Scholar] [CrossRef] [PubMed]
- Pepe, G.J.; Albrecht, E.D. Regulation of functional differentiation of the placental villous syncytiotrophoblast by estrogen during primate pregnancy. Steroids 1999, 64, 624–627. [Google Scholar] [CrossRef]
- Pepe, G.J.; Johnson, D.K.; Albrecht, E.D. The effects of estrogen on cortisol metabolism in female baboons. Steroids 1982, 39, 471–477. [Google Scholar] [CrossRef]
- Miller, W.L.; Auchus, R.J. The molecular biology, biochemistry, and physiology of human steroidogenesis and its disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Kamarck, T.; Mermelstein, R. A global measure of perceived stress. J. Health Soc. Behav. 1983, 24, 385–396. [Google Scholar] [CrossRef]
- La Frano, M.R.; Carmichael, S.L.; Ma, C.; Hardley, M.; Shen, T.; Wong, R.; Rosales, L.; Borkowski, K.; Pedersen, T.L.; Shaw, G.M.; et al. Impact of post-collection freezing delay on the reliability of serum metabolomics in samples reflecting the California mid-term pregnancy biobank. Metabolomics 2018, 14, 151. [Google Scholar] [CrossRef]
- Dahlerup, B.R.; Egsmose, E.L.; Siersma, V.; Mortensen, E.L.; Hedegaard, M.; Knudsen, L.E.; Mathiesen, L. Maternal stress and placental function, a study using questionnaires and biomarkers at birth. PLoS ONE 2018, 13, e0207184. [Google Scholar] [CrossRef]
- La Marca-Ghaemmaghami, P.; La Marca, R.; Dainese, S.M.; Haller, M.; Zimmermann, R.; Ehlert, U. The association between perceived emotional support, maternal mood, salivary cortisol, salivary cortisone, and the ratio between the two compounds in response to acute stress in second trimester pregnant women. J. Psychosom. Res. 2013, 75, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Pepe, G.J.; Albrecht, E.D. Comparison of cortisol-cortisone interconversion in vitro by the human and baboon placenta. Steroids 1984, 44, 229–240. [Google Scholar] [CrossRef]
- Copper, R.L.; Goldenberg, R.L.; Das, A.; Elder, N.; Swain, M.; Norman, G.; Ramsey, R.; Cotroneo, P.; Collins, B.A.; Johnson, F.; et al. The preterm prediction study: Maternal stress is associated with spontaneous preterm birth at less than thirty-five weeks’ gestation. National Institute of Child Health and Human Development Maternal-Fetal Medicine Units Network. Am. J. Obstet. Gynecol. 1996, 175, 1286–1292. [Google Scholar] [CrossRef]
- Dunkel Schetter, C. Psychological science on pregnancy: Stress processes, biopsychosocial models, and emerging research issues. Annu. Rev. Psychol. 2011, 62, 531–558. [Google Scholar] [CrossRef] [PubMed]
- Entringer, S.; Buss, C.; Wadhwa, P.D. Prenatal stress, development, health and disease risk: A psychobiological perspective-2015 Curt Richter Award Paper. Psychoneuroendocrinology 2015, 62, 366–375. [Google Scholar] [CrossRef]
- Farr, S.L.; Dietz, P.M.; O’Hara, M.W.; Burley, K.; Ko, J.Y. Postpartum anxiety and comorbid depression in a population-based sample of women. J. Womens Health 2014, 23, 120–128. [Google Scholar] [CrossRef]
- Hobel, C.J.; Goldstein, A.; Barrett, E.S. Psychosocial stress and pregnancy outcome. Clin. Obstet. Gynecol. 2008, 51, 333–348. [Google Scholar] [CrossRef]
- Witt, W.P.; Litzelman, K.; Cheng, E.R.; Wakeel, F.; Barker, E.S. Measuring stress before and during pregnancy: A review of population-based studies of obstetric outcomes. Matern. Child Health J. 2014, 18, 52–63. [Google Scholar] [CrossRef] [PubMed]
- American College of Obstetricians and Gynecologists Committee on Health Care for Undeserved Women. ACOG Committee Opinion No. 343: Psychosocial risk factors: Perinatal screening and intervention. Obstet. Gynecol. 2006, 108, 469–477. [Google Scholar] [CrossRef]
- Albrecht, E.D.; Aberdeen, G.W.; Pepe, G.J. The role of estrogen in the maintenance of primate pregnancy. Am. J. Obstet. Gynecol. 2000, 182, 432–438. [Google Scholar] [CrossRef]
- Soldin, O.P.; Guo, T.; Weiderpass, E.; Tractenberg, R.E.; Hilakivi-Clarke, L.; Soldin, S.J. Steroid hormone levels in pregnancy and 1 year postpartum using isotope dilution tandem mass spectrometry. Fertil. Steril. 2005, 84, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Tulchinsky, D.; Korenman, S.G. The plasma estradiol as an index of fetoplacental function. J. Clin. Investig. 1971, 50, 1490–1497. [Google Scholar] [CrossRef]
- Musana, J.W.; Cohen, C.R.; Kuppermann, M.; Gerona, R.; Wanyoro, A.; Aguilar, D.; Santos, N.; Temmerman, M.; Weiss, S.J. Association of differential symptoms of stress to hair cortisol and cortisone concentrations among pregnant women in Kenya. Stress 2020, 23, 556–566. [Google Scholar] [CrossRef]
- Schreier, H.M.; Enlow, M.B.; Ritz, T.; Coull, B.A.; Gennings, C.; Wright, R.O.; Wright, R.J. Lifetime exposure to traumatic and other stressful life events and hair cortisol in a multi-racial/ethnic sample of pregnant women. Stress 2016, 19, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Schury, K.; Koenig, A.M.; Isele, D.; Hulbert, A.L.; Krause, S.; Umlauft, M.; Kolassa, S.; Ziegenhain, U.; Karabatsiakis, A.; Reister, F.; et al. Alterations of hair cortisol and dehydroepiandrosterone in mother-infant-dyads with maternal childhood maltreatment. BMC Psychiatry 2017, 17, 213. [Google Scholar] [CrossRef] [PubMed]
- Smy, L.; Shaw, K.; Amstutz, U.; Smith, A.; Berger, H.; Carleton, B.; Koren, G. Hair cortisol as a hypothalamic-pituitary-adrenal axis biomarker in pregnant women with asthma: A retrospective observational study. BMC Pregnancy Childbirth 2016, 16, 176. [Google Scholar] [CrossRef]
- Yellon, S.M.; Dobyns, A.E.; Beck, H.L.; Kurtzman, J.T.; Garfield, R.E.; Kirby, M.A. Loss of progesterone receptor-mediated actions induce preterm cellular and structural remodeling of the cervix and premature birth. PLoS ONE 2013, 8, e81340. [Google Scholar] [CrossRef] [PubMed]
- Norman, J.E.; Marlow, N.; Messow, C.M.; Shennan, A.; Bennett, P.R.; Thornton, S.; Robson, S.C.; McConnachie, A.; Petrou, S.; Sebire, N.J.; et al. Vaginal progesterone prophylaxis for preterm birth (the OPPTIMUM study): A multicentre, randomised, double-blind trial. Lancet 2016, 387, 2106–2116. [Google Scholar] [CrossRef]
- Romero, R.; Conde-Agudelo, A.; Da Fonseca, E.; O’Brien, J.M.; Cetingoz, E.; Creasy, G.W.; Hassan, S.S.; Nicolaides, K.H. Vaginal progesterone for preventing preterm birth and adverse perinatal outcomes in singleton gestations with a short cervix: A meta-analysis of individual patient data. Am. J. Obstet. Gynecol. 2018, 218, 161–180. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Gestational Age | BMI | Infant Weight (g) | Infant Sex | Apgar Score | Mode of Delivery |
|---|---|---|---|---|---|---|
| P-002 | 39 + 4 | 24.8 | 2710 | F | 9 9 | NSVD |
| P-006 | 39 + 2 | 27.6 | 2772 | F | 9 9 | NSVD |
| P-016 | 39 + 6 | 26.3 | 3330 | M | 9 9 | NSVD |
| P-022 | 37 + 0 | 26.9 | 2360 | F | 9 9 | NSVD |
| P-023 | 41 + 3 | 23.3 | 3600 | M | 8 9 | NSVD |
| P-026 | 41 + 1 | 27.0 | 3390 | M | 8 9 | NSVD |
| P-030 | 40 + 0 | 26.7 | 3500 | F | 8 9 | NSVD |
| P-032 | 40 + 0 | 29.0 | 3835 | F | 9 9 | LTCS |
| P-034 | 41 + 0 | 52.2 | 4260 | F | 7 9 | NSVD |
| P-037 | 39 + 3 | 24.7 | 3525 | M | 9 9 | VAVD |
| P-038 | 41 + 6 | 33.8 | 3460 | F | 8 9 | NSVD |
| P-041 | 39 + 6 | 36.6 | 2895 | F | 4 9 | NSVD |
| P-043 | 40 + 4 | 27.1 | 3605 | F | 8 9 | NSVD |
| P-046 | 39 + 1 | 27.1 | 2850 | F | 1 9 | LTCS |
| P-048 | 39 + 6 | 22.3 | 2870 | F | 4 9 | NSVD |
| P-049 | 39 + 1 | 24.2 | 3040 | M | 9 9 | NSVD |
| P-051 | 39 + 3 | 35.5 | 4160 | M | 3 8 | LTCS |
| Metabolite | LOD (ng/mL) | Cord Plasma ± SEM (ng/mL) | Maternal Plasma ± SEM (ng/mL) | LOD (pg/mg) | Placenta Mean ± SEM (pg/mg) | Hair Mean ± SEM (pg/mg) | ||
|---|---|---|---|---|---|---|---|---|
| Patient X | Average ± SEM | Patient X | Average ± SEM | |||||
| Cortisol (CRTL) | 1.811 | 31.68 | 22.18 ± 5.82 | 50.68 | 57.96 ± 24.81 | 36.22 | <LOD (<7.06) | <LOD (<0.76) |
| Cortisone (CRTN) | 0.360 | 167.78 | 52.21 ± 3.62 | 361.72 | 10.15 ± 2.21 | 7.20 | 370.82 ± 47.05 | <LOD (<0.98) |
| Progesterone (P4) | 0.393 | 965.64 | 175.66 ± 18.67 | 318.46 | 36.28 ± 1.64 | 7.86 | 4884.46 ± 443.97 | 15.92 ± 5.85 |
| Allo-pregnanolone (ALLO) | 4.774 | 1.02 | <LOD (<0.72) | 12.3 | <LOD (<0.50) | 95.48 | 168.88 ± 46.73 | <LOD (<7.31) |
| Dihydroprogesterone (DHP) | 39.530 | 24.44 | <LOD (<13.33) | 5.59 | <LOD (<2.32) | 790.60 | <LOD (<200.00) | <LOD (<60.88) |
| Estrone (E1) | 2.702 | 4.99 | <LOD (<3.3) | 69.22 | <LOD (<0.32) | 54.03 | 232.78 ± 37.47 | <LOD (<1.28) |
| Estradiol (E2) | 0.680 | 1.11 | 1.55 ± 0.33 | 18.27 | 3.96 ± 0.56 | 13.61 | 34.32 ± 6.29 | <LOD (<0.12) |
| Estriol (E3) | 2.882 | 369.04 | 32.28 ± 8.04 | 674.53 | <LOD (<1.72) | 57.64 | 356.34 ± 61.12 | <LOD (<0.83) |
| Dehydroepiandrosterone (DHEA) | 7.205 | 0.407 | <LOD (<0.30) | 0.03 | <LOD (<0.48) | 144.10 | <LOD (<1.08) | <LOD (<2.25) |
| Dehydroepiandrosterone sulfate (DHEAS) | 18.424 | 2983.48 | 387.47 ± 45.83 | 4969.63 | 52.28 ± 30.13 | 368.49 | <LOD (<1.53) | <LOD (<4.39) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vuppaladhadiam, L.; Lager, J.; Fiehn, O.; Weiss, S.; Chesney, M.; Hasdemir, B.; Bhargava, A. Human Placenta Buffers the Fetus from Adverse Effects of Perceived Maternal Stress. Cells 2021, 10, 379. https://doi.org/10.3390/cells10020379
Vuppaladhadiam L, Lager J, Fiehn O, Weiss S, Chesney M, Hasdemir B, Bhargava A. Human Placenta Buffers the Fetus from Adverse Effects of Perceived Maternal Stress. Cells. 2021; 10(2):379. https://doi.org/10.3390/cells10020379
Chicago/Turabian StyleVuppaladhadiam, Lahari, Jeannette Lager, Oliver Fiehn, Sandra Weiss, Margaret Chesney, Burcu Hasdemir, and Aditi Bhargava. 2021. "Human Placenta Buffers the Fetus from Adverse Effects of Perceived Maternal Stress" Cells 10, no. 2: 379. https://doi.org/10.3390/cells10020379
APA StyleVuppaladhadiam, L., Lager, J., Fiehn, O., Weiss, S., Chesney, M., Hasdemir, B., & Bhargava, A. (2021). Human Placenta Buffers the Fetus from Adverse Effects of Perceived Maternal Stress. Cells, 10(2), 379. https://doi.org/10.3390/cells10020379