
Exogenous Application of ABA and NAA Alleviates the Delayed Coloring Caused by Puffing Inhibitor in Citrus Fruit



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
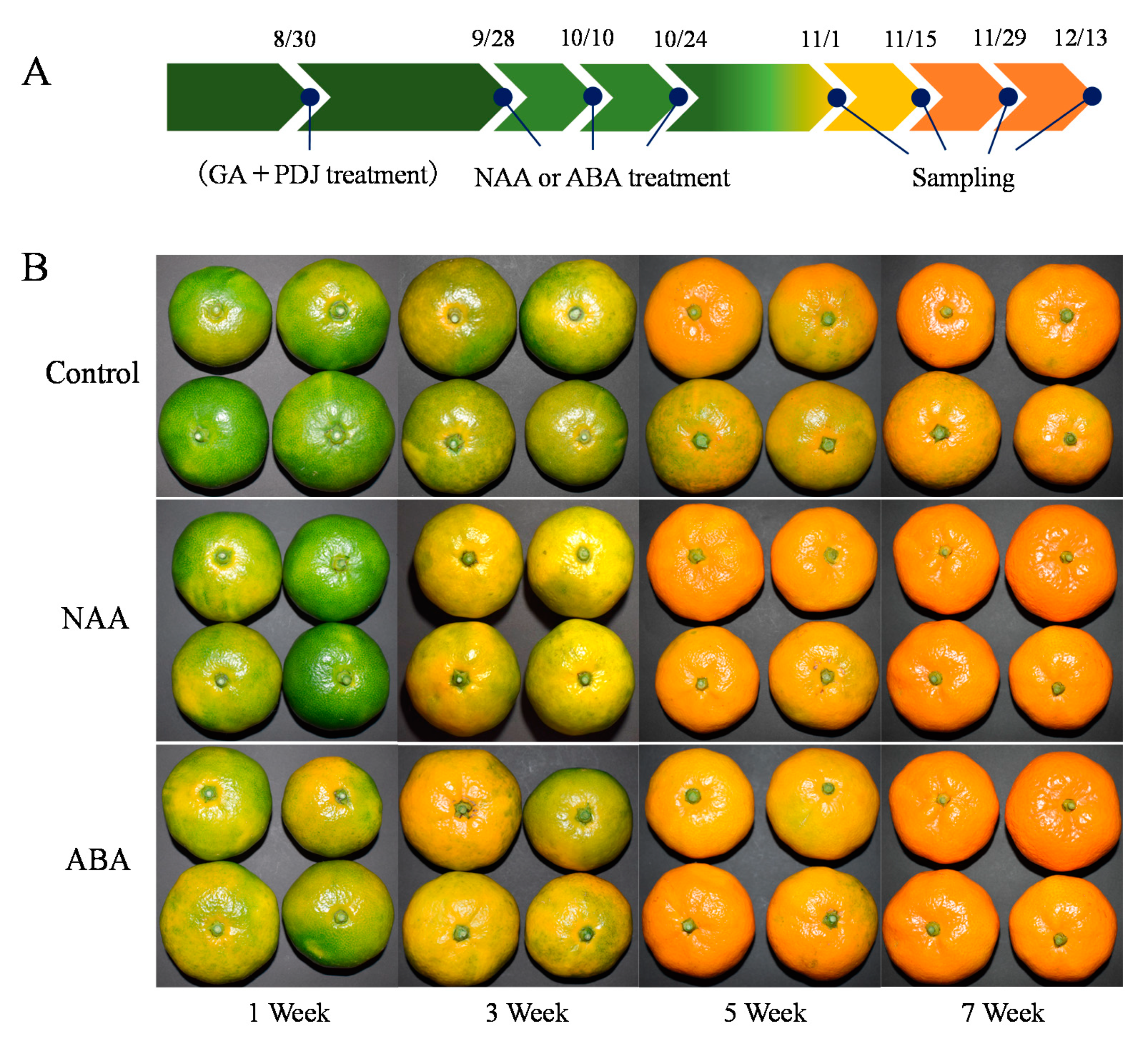
2.1. Plant Materials and Treatments
2.2. Color Measurement
2.3. Extraction and Determination of Chlorophylls
2.4. Extraction and Determination of Carotenoids
2.5. Total RNA Extraction and Real-Time Quantitative RT-PCR
2.6. Statistical Analysis
3. Results
3.1. Effects of NAA and ABA Spraying on the Color of the GA and PDJ-Treated Citrus Fruit
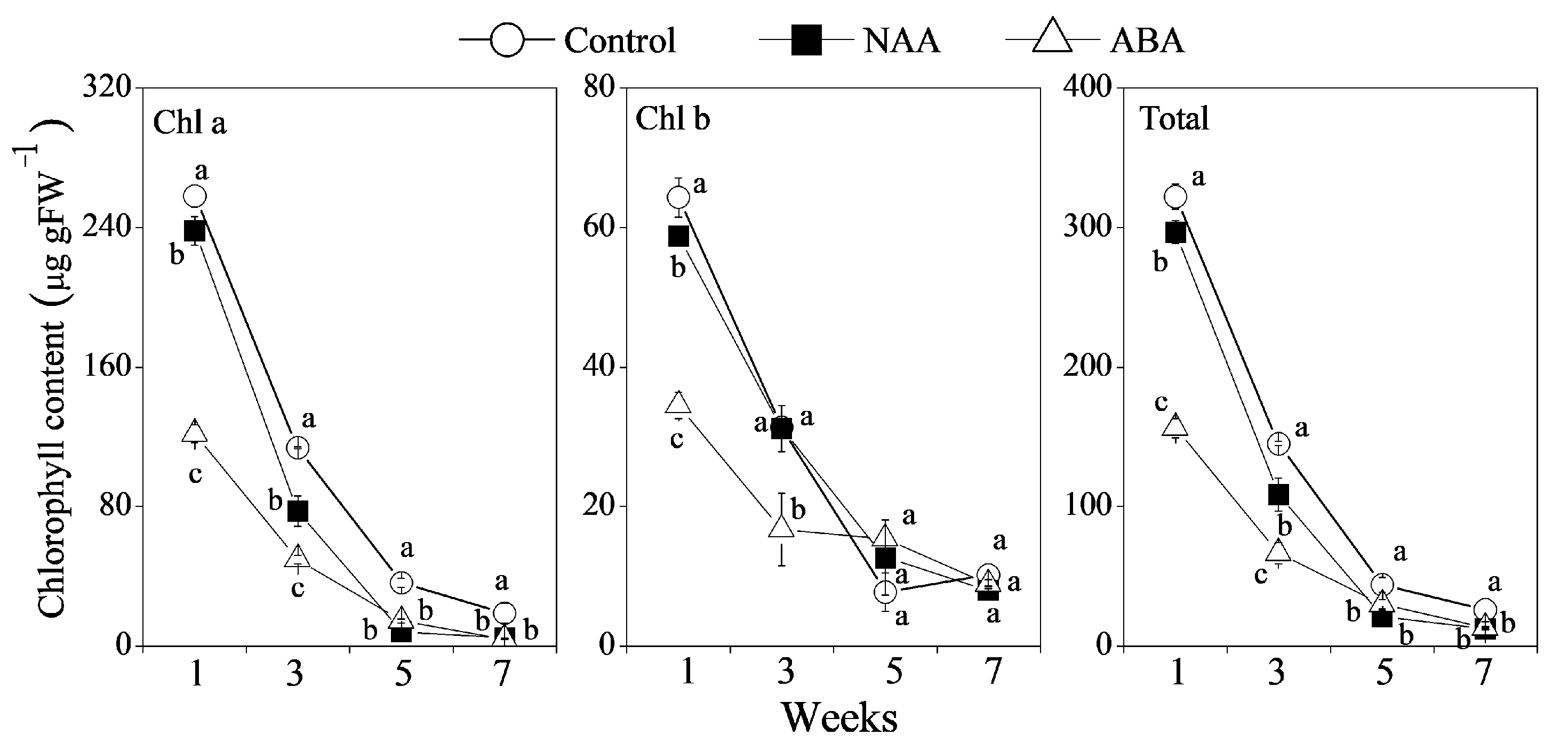
3.2. Effects of NAA and ABA Spraying on Chlorophyll Accumulation in the GA and PDJ-Treated Citrus Fruit
3.3. Effects of NAA and ABA Spraying on Carotenoid Accumulation in the GA and PDJ-Treated Citrus Fruit
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Martinelli, F.; Ibanez, A.M.; Reagan, R.L.; Davino, S.; Dandekar, A.M. Stress responses in citrus peel: Comparative analysis of host responses to Huanglongbing disease and puffing disorder. Sci. Hortic. 2015, 192, 409–420. [Google Scholar] [CrossRef]
- Kawase, K.; Suzuki, K.; Hirose, K. Use of Growth Regulators to Control Rind Puffing in Satsuma Mandarin Fruit; International Society of Citriculture: Shimizu, Japan, 1981; pp. 237–239. [Google Scholar]
- Garcialuis, A.; Agusti, M.; Almela, V.; Romero, E.; Guardiola, J.L. Effect of gibberellic-acid on ripening and peel puffing in satsuma mandarin. Sci. Hortic. 1985, 27, 75–86. [Google Scholar] [CrossRef]
- Shiraishi, M.; Mohammad, P.; Makita, Y.; Fujibuchi, M.; Manabe, T. Effects of calcium compounds on fruit puffing and the ultrastructural characteristics of the subepidermal cell walls of puffy and calcium-induced non-puffy Satsuma Mandarin fruits. J. Jap. Soc. Hortic. Sci. 1999, 68, 919–926. [Google Scholar] [CrossRef]
- Sato, K.; Ikoma, Y.; Matsumoto, H.; Nakajima, N. Effects of spray concentration and spray times of gibberellin and prohydrojasmon on peel puffing and peel color in Satsuma mandarin fruit. Hort. Res. 2015, 14, 419–426. [Google Scholar] [CrossRef]
- Iglesias, D.J.; Cercos, M.; Colmenero-Flores, J.M.; Naranjo, M.A.; Rios, G.; Carrera, E.; Ruiz-Rivero, O.; Lliso, I.; Morillon, R.; Tadeo, F.R.; et al. Physiology of citrus fruiting. Braz. J. Plant Physiol. 2007, 19, 333–362. [Google Scholar] [CrossRef]
- Kato, M.; Ikoma, Y.; Matsumoto, H.; Sugiura, M.; Hyodo, H.; Yano, M. Accumulation of carotenoids and expression of carotenoid biosynthetic genes during maturation in citrus fruit. Plant Physiol. 2004, 134, 824–837. [Google Scholar] [CrossRef]
- Tadeo, F.R.; Cercos, M.; Colmenero-Flores, J.M.; Iglesias, D.J.; Naranjo, M.A.; Rios, G.; Carrera, E.; Ruiz-Rivero, O.; Lliso, I.; Morillon, R.; et al. Molecular physiology of development and quality of citrus. Adv. Bot. Res. 2008, 47, 147–223. [Google Scholar]
- Ríos, G.; Naranjo, M.A.; Rodrigo, M.J.; Alós, E.; Zacarías, L.; Cercós, M.; Talón, M. Identification of a GCC transcription factor responding to fruit colour change events in citrus through the transcriptomic analyses of two mutants. BMC Plant Biol. 2010, 10, 276. [Google Scholar] [CrossRef]
- Rodrigo, M.J.; Alquézar, B.; Alós, E.; Medina, V.; Carmona, L.; Bruno, M.; Al-Babili, S.; Zacarías, L. A novel carotenoid cleavage activity involved in the biosynthesis of citrus fruit-specific apocarotenoid pigments. J. Exp. Bot. 2013, 64, 4461–4478. [Google Scholar] [CrossRef]
- Bouvier, F.; Rahier, A.; Camara, B. Biogenesis, molecular regulation and function of plant isoprenoids. Prog. Lipid Res. 2005, 44, 357–429. [Google Scholar] [CrossRef]
- Pulido, P.; Perello, C.; Rodríguez-Concepción, M. New insights into plant isoprenoid metabolism. Mol. Plant 2012, 5, 964–967. [Google Scholar] [CrossRef]
- Kobayashi, K.; Masuda, T. Transcriptional regulation of tetrapyrrole biosynthesis in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 1811. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, M.J.; Marcos, J.F.; Zacarías, L. Biochemical and molecular analysis of carotenoid biosynthesis in flavedo of orange (Citrus sinensis L.) during fruit development and maturation. J. Agric. Food. Chem. 2004, 52, 6724–6731. [Google Scholar] [CrossRef] [PubMed]
- Alós, E.; Cercós, M.; Rodrigo, M.J.; Zacarías, L.; Talón, M. Regulation of color break in citrus fruits. Changes in pigment profiling and gene expression induced by gibberellins and nitrate, two ripening retardants. J. Agric. Food Chem. 2006, 54, 4888–4895. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, M.J.; Zacarías, L. Effect of postharvest ethylene treatment on carotenoid accumulation and the expression of carotenoid biosynthetic genes in the flavedo of orange (Citrus sinensis L. Osbeck) fruit. Postharvest Biol. Technol. 2007, 43, 14–22. [Google Scholar] [CrossRef]
- Ma, G.; Zhang, L.C.; Matsuta, A.; Matsutani, K.; Yamawaki, K.; Yahata, M.; Wahyudi, A.; Motohashi, R.; Kato, M. Enzymatic formation of β-citraurin from β-cryptoxanthin and zeaxanthin by carotenoid cleavage dioxygenase4 in the flavedo of citrus fruit. Plant Physiol. 2013, 163, 682–695. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Chen, C.; Yu, Q.; Gady, A.; Yu, Y.; Liang, G.; Gmitter, F.G., Jr. Comparison of carotenoid accumulation and biosynthetic gene expression between Valencia and Rohde Red Valencia sweet oranges. Plant Sci. 2014, 227, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Zhang, L.C.; Kitaya, Y.; Seoka, M.; Kudaka, R.; Yahata, M.; Yamawaki, K.; Shimada, T.; Fujii, H.; Endo, T.; et al. Blue LED light induces regreening in the flavedo of Valencia orange in vitro. Food Chem. 2021, 335, 127621. [Google Scholar] [CrossRef]
- Matsumoto, H.; Ikoma, Y.; Kato, M.; Nakajima, N.; Hasegawa, Y. Effect of postharvest temperature and ethylene on carotenoid accumulation in the flavedo and juice sacs of Satsuma Mandarin (Citrus unshiu Marc.) fruit. J. Agric. Food Chem. 2009, 57, 4724–4732. [Google Scholar] [CrossRef]
- Ma, G.; Zhang, L.C.; Kato, M.; Yamawaki, K.; Kiriiwa, Y.; Yahata, M.; Matsumoto, H. Effect of blue and red LED light irradiation on β-cryptoxanthin accumulation in the flavedo of citrus fruits. J. Agric. Food Chem. 2012, 60, 197–201. [Google Scholar] [CrossRef]
- Lado, J.; Alós, E.; Manzi, M.; Cronje, P.J.R.; Gómez-Cadenas, A.; Rodrigo, M.J.; Zacarías, L. Light regulation of carotenoid biosynthesis in the peel of mandarin and sweet orange fruits. Front. Plant Sci. 2019, 10, 1288. [Google Scholar] [CrossRef] [PubMed]
- Mitalo, O.W.; Otsuki, T.; Okada, R.; Obitsu, S.; Masuda, K.; Hojo, Y.; Matsuura, T.; Mori, I.C.; Abe, D.; Asiche, W.O.; et al. Low temperature modulates natural peel degreening in lemon fruit independently of endogenous ethylene. J. Exp. Bot. 2020, 71, 4778–4796. [Google Scholar] [CrossRef] [PubMed]
- García-Luís, A.; Herrero-Villén, A.; Guardiola, J.L. Effects of applications of gibberellic acid on late growth, maturation and pigmentation of the Clementine mandarin. Sci. Hortic. 1992, 49, 71–82. [Google Scholar] [CrossRef]
- Fujii, H.; Shimada, T.; Sugiyama, A.; Endo, T.; Nishikawa, F.; Nakano, M.; Ikoma, Y.; Shimizu, T.; Omura, M. Profiling gibberellin (GA3)-responsive genes in mature mandarin fruit using a citrus 22 K oligoarray. Sci. Hortic. 2008, 116, 291–298. [Google Scholar] [CrossRef]
- Valero, D.; Martínez-Romero, D.; Serrano, M.; Riquelme, F. Influence of postharvest treatment with putrescine and calcium on endogenous polyamines, firmness, and abscisic acid in lemon (Citrus lemon L. Burm cv. Verna). J. Agric. Food Chem. 1998, 46, 2102–2109. [Google Scholar] [CrossRef]
- Rodrigo, M.J.; Marcos, J.F.; Alférez, F.; Mallent, M.D.; Zacarías, L. Characterization of Pinalate, a novel Citrus sinensis mutant with a fruit specific alteration that results in yellow pigmentation and decreased ABA content. J. Exp. Bot. 2003, 54, 727–738. [Google Scholar] [CrossRef]
- Wang, X.; Yin, W.; Wu, J.; Chai, L.; Yi, H. Effects of exogenous abscisic acid on the expression of citrus fruit ripening-related genes and fruit ripening. Sci. Hortic. 2016, 201, 175–183. [Google Scholar] [CrossRef]
- Bermejo, A.; Granero, B.; Mesejo, C.; Reig, C.; Tejedo, V.; Agustí, M.; Primo-Millo, E.; Iglesias, D.J. Auxin and gibberellin interact in citrus fruit set. J. Plant Growth Regul. 2018, 37, 491–501. [Google Scholar] [CrossRef]
- Rehmana, M.; Singha, Z.; Khurshid, T. Pre-harvest spray application of abscisic acid (S-ABA) regulates fruit colour development and quality in early maturing M7 Navel orange. Sci. Hortic. 2018, 229, 1–9. [Google Scholar] [CrossRef]
- Pérez-Llorca, M.; Muñoz, P.; Müller, M.; Munné-Bosch, S. Biosynthesis, metabolism and function of auxin, salicylic acid and melatonin in climacteric and non-climacteric fruits. Front. Plant Sci. 2019, 10, 136. [Google Scholar] [CrossRef]
- Ma, G.; Zhang, L.; Kato, M.; Yamawaki, K.; Kiriiwa, Y.; Yahata, M.; Ikoma, Y.; Matsumoto, H. Effect of the combination of ethylene and red LED light irradiation on carotenoid accumulation and carotenogenic gene expression in the flavedo of citrus fruit. Postharvest Biol. Technol. 2015, 99, 99–104. [Google Scholar] [CrossRef]
- Moran, R. Formulae for determination of chlorophyllous pigments extracted with N,N-dimethylformamide. Plant Physiol. 1982, 69, 1376–1381. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Ikoma, Y. Effects of elevated temperatures during the flowering to physiological fruit drop stage and at the fruit maturation stage on fruit quality of the satsuma mandarin. J. Agric. Meteorol. 2020, 76, 29–35. [Google Scholar] [CrossRef]
- Sugiura, T.; Kuroda, H.; Sugiura, H. Influence of the current state of global warming on fruit tree growth in Japan. Hort. Res. 2007, 6, 257–263. [Google Scholar] [CrossRef]
- Zhang, L.C.; Ma, G.; Kato, M.; Yamawaki, K.; Takagi, T.; Kiriiwa, Y.; Ikoma, Y.; Matsumoto, H.; Yoshioka, T.; Nesumi, H. Regulation of carotenoid accumulation and the expression of carotenoid metabolic genes in citrus juice sacs in vitro. J. Exp. Bot. 2012, 63, 871–886. [Google Scholar] [CrossRef] [PubMed]
- Alquézar, B.; Zacarías, L.; Rodrigo, M.J. Molecular and functional characterization of a novel chromoplast-specific lycopene β-cyclase from Citrus and its relation to lycopene accumulation. J. Exp. Bot. 2009, 60, 1783–1797. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, G.; Shirai, Y.; Kato, M.; Yamawaki, K.; Ikoma, Y.; Matsumoto, H. Expression and functional analysis of two lycopene β-cyclases from citrus fruits. Planta 2012, 236, 1315–1325. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Zhang, L.C.; Yungyuen, W.; Tsukamoto, I.; Iijima, N.; Oikawa, M.; Yamawaki, K.; Yahata, M.; Kato, M. Expression and functional analysis of citrus carotene hydroxylases: Unraveling the xanthophyll biosynthesis in citrus fruits. BMC Plant Biol. 2016, 16, 148. [Google Scholar] [CrossRef]
- Oberholster, R.; Cowan, A.K.; Molnar, P.; Toth, G. Biochemical basis of color as an aesthetic quality in Citrus sinensis. J. Agric. Food Chem. 2001, 49, 303–307. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, G.; Zhang, L.; Kudaka, R.; Inaba, H.; Furuya, T.; Kitamura, M.; Kitaya, Y.; Yamamoto, R.; Yahata, M.; Matsumoto, H.; et al. Exogenous Application of ABA and NAA Alleviates the Delayed Coloring Caused by Puffing Inhibitor in Citrus Fruit. Cells 2021, 10, 308. https://doi.org/10.3390/cells10020308
Ma G, Zhang L, Kudaka R, Inaba H, Furuya T, Kitamura M, Kitaya Y, Yamamoto R, Yahata M, Matsumoto H, et al. Exogenous Application of ABA and NAA Alleviates the Delayed Coloring Caused by Puffing Inhibitor in Citrus Fruit. Cells. 2021; 10(2):308. https://doi.org/10.3390/cells10020308
Chicago/Turabian StyleMa, Gang, Lancui Zhang, Rin Kudaka, Hayato Inaba, Takuma Furuya, Minami Kitamura, Yurika Kitaya, Risa Yamamoto, Masaki Yahata, Hikaru Matsumoto, and et al. 2021. "Exogenous Application of ABA and NAA Alleviates the Delayed Coloring Caused by Puffing Inhibitor in Citrus Fruit" Cells 10, no. 2: 308. https://doi.org/10.3390/cells10020308
APA StyleMa, G., Zhang, L., Kudaka, R., Inaba, H., Furuya, T., Kitamura, M., Kitaya, Y., Yamamoto, R., Yahata, M., Matsumoto, H., & Kato, M. (2021). Exogenous Application of ABA and NAA Alleviates the Delayed Coloring Caused by Puffing Inhibitor in Citrus Fruit. Cells, 10(2), 308. https://doi.org/10.3390/cells10020308