
Astrocytic Regulation of Neural Circuits Underlying Behaviors

Abstract
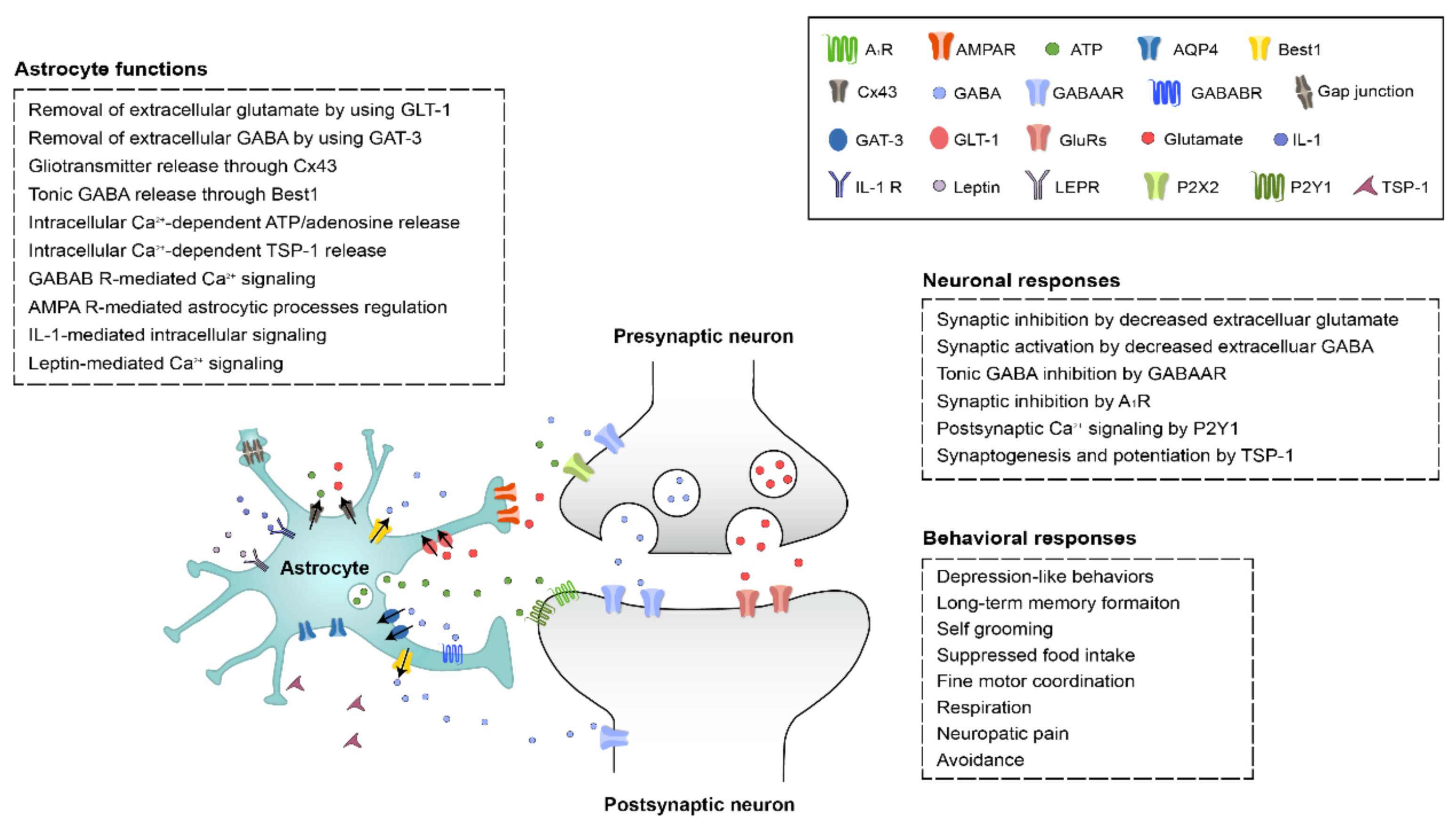
1. Introduction
2. Olfactory Bulb
3. Prefrontal Cortex
4. Motor and Somatosensory Cortices
5. Anterior Cingulate Cortex
6. Hippocampus
7. Striatum
8. Amygdala
9. Thalamus
10. Hypothalamus
10.1. Arcuate Nucleus
10.2. Suprachiasmatic Nucleus
11. Midbrain
11.1. Ventral Tegmental Area
11.2. Periaqueductal Grey
12. Medulla Oblongata
13. Cerebellum
14. Spinal Cord
15. Concluding Remarks

{kind=link}
{kind=link}
| Target Region | Manipulation | Change in Astrocytes | Consequence of Manipulation | Ref | |
|---|---|---|---|---|---|
| Circuit | Behavior | ||||
| OB | Aldh1L1-cre mice/ GFAP: FLEX-hM3Dq | n.d. | Odor-evoked Ca2+ in M/Tsș↓ | Odor detection accuracy in Go/No-Go task↑ | [32] |
| Aldh1L1-cre mice/ GFAP: FLEX-hM4Di | n.d. | Odor-evoked Ca2+ in M/Ts↑ | Odor detection accuracy in Go/No-Go task↓ | ||
| PFC | WT mice/L-AAA (astrocyte-specific cytotoxin) | GFAP+ cells↓ | Progressive neuronal loss | Spatial memory in Morris water maze test↓ Reversal learning in attentional set-shifting task↓ | [35] |
| WT mice/L-AAA | GFAP+ cells↓ | n.d. | Sucrose preference in SPT↓ Latency to feed in novelty suppressed feeding test↑ Immobility in FST↑ Escape latency in active avoidance test↑ | [25] | |
| WT mice/DHK (GLT-1 inhibitor) | n.d. | n.d. | Intracranial self-stimulation threshold↑ Latency to begin drinking sucrose in sucrose intake test↑ | [26] | |
| WT mice/ATP or ATP-γ-S | n.d. | n.d. | CSDS-induced social avoidance in SIT↓ CSDS-induced immobility in FST↓ CMS-induced fur condition↓ Post-CMS sucrose preference in SPT↑ | [24] | |
| IP3R2−/− mice | ATP release↓ | n.d. | Immobility in FST↑ Fur condition↓ Sucrose consumption in SPT↓ | ||
| GFAP-tTA;tetO-dnSNARE mice | ATP release↓ | n.d. | Immobility in FST↑ | ||
| GFAP-tTA;tetO-MrgprA1 mice | ATP release↑ | n.d. | Immobility in FST↓ Social avoidance in SIT↓ | ||
| WT mice/AAV GfaABC1D-hM3Dq | c-Fos+ cells↑ | n.d. | Ethanol consumption & preference↑ Locomotor activity (by low-dose ethanol)↑ Duration of LORR (by high-dose ethanol)↑ | [39] | |
| WT mice/AAV GfaABC1D::hPMCA2w/b | c-Fos+ cells↓ | n.d. | Ethanol consumption, preference↓ Duration of LORR (high-dose ethanol)↓ | ||
| MC | GLAST-creER;IP3R2fl/fl mice | Ca2+↓ | n.d. | Motor-skill learning in forelimb reaching task↓ | [40] |
| WT mice/fluorocitrate (astrocyte metabolic inhibitor) | Ca2+↓ | LTP↓, GluA1↓ | Motor-skill learning in forelimb reaching task↓ | ||
| SC | IP3R2−/− mice | Ca2+↓ | PSL-induced TSP-1 upregulation↓ | PSL-induced mechanical allodynia↓ | [41] |
| WT mice/BAPTA-AM (Ca2+ chelator) | n.d. | PSL-induced mechanical allodynia↓ | |||
| Fluoroacetate (astrocyte metabolic inhibitor) | n.d. | n.d. | PSL-induced mechanical allodynia↓ | ||
| ACC | L-AAA | GFAP intensity↓ | n.d. | CFA-induced place escape/avoidance↓ | [27] |
| GFAP-cre mice/ AAV CBA::FLEX-ChR2 | n.d. | n.d. | Non-rapid eye movement sleep↓ | [44] | |
| WT mice/SNAP-5114 (GAT-3 inhibitor) | GAT-3 activity↓ | n.d. | PSL-induced sleep disturbance↓ | [45] | |
| HP | WT mice/AAV GFAP::hM3Dq | Ca2+↑ | LTP↑ Neuronal c-Fos in CA1 (when learning is combined)↑ | Spatial memory in T-maze test↑ Contextual memory in fear conditioning test↑ | [47] |
| WT mice/AAV GFAP::hM4Di | Ca2+↓ | CA3→CA1 synaptic transmission↓ CA1→ACC neuronal activity↓ | Remote contextual memory in fear conditioning test↓ | [48] | |
| IL-1r−/− mice | n.d. | LTP↓ | Spatial memory in Morris water maze test↓ Contextual memory in fear conditioning test↓ | [50] | |
| IL-1r−/− mice/transplantation of WT NPCs | n.d. | LTP↑ | Spatial memory in Morris water maze test↑ Contextual memory in fear conditioning test↑ | ||
| GFAP-tTA;tetO-dnSNARE mice | n.d. | Sleep deprivation-induced LTP deficits↓ | Sleep deprivation-induced spatial memory impairment in SOR test↓ | [49] | |
| WT mice/CPT (adenosine A1Rantagonist) | n.d. | Sleep deprivation-induced LTP deficits↓ | Sleep deprivation-induced spatial memory impairment in SOR test↓ | ||
| AQP4−/− mice | n.d. | LTP↓ LTD↓ | Object recognition memory in object placement test↓ | [166] | |
| GFAP-cre;Cx43fl/fl;Cx30−/− mice | Edema, vacuolation | Widespread edema, vacuolation (white matter) | Spatial memory in SOR test↓ Motor coordination in rotarod and balance beam assays↓ | [52] | |
| STR | WT mice/AAV GfaABC1D::hPMCA2w/b | Ca2+↓ GAT-3↑ | Tonic GABA current↓ D1-MSN tonic inhibition↓ MSN excitability↓ | Duration of self-grooming↑ Travel distance in open field test↓ Time spent in center in open field test↓ | [55] |
| WT mice/AAV GfaABC1D::hM4Di | Ca2+↑ TSP-1↑ | MSN EPSC↑ MSN firing↑ | Travel distance in open field test↑ Investigatory activity in light/dark open field test↓ Exploration of novel object in NOR test↓ | [56] | |
| DRD1fl/fl mice/AAV GFAP::mCherry-cre | DA-evoked Ca2+↓ | DA-evoked synaptic depression↓ | Amphetamine-evoked locomotion enhancement↓ | [58] | |
| IP3R2−/− mice | DA-evoked Ca2+↓ | DA-evoked synaptic depression↓ | Amphetamine-evoked locomotion enhancement↓ | ||
| WT mice/AAV GFAP::hM3Dq | Intracellular Ca2+↑ ATP/adenosine release↑ | Synaptic depression↑ | n.d. | ||
| AMY | WT mice/AAV GFAP::hM3Dq | Intracellular Ca2+↑ | BLA→CeM excitatory synaptic transmission↓(by A1R↑) CeL→CeM inhibitory synaptic transmission↑(by A2R↑) CeM neuronal firing↓ | Fear memory formation in fear conditioning tes↓ | [73] |
| WT mice/DHK (GLT-1 inhibitor) | n.d. | n.d. | Intracranial self-stimulation threshold↑ Time spent in open arms in elevated plus maze test↓ Freezing behavior in fear conditioning test↑ | [74] | |
| WT mice/TAT-Cx43L2 (Cx43-hemichannel blocker) | Cx43 activity↓ | No effects on synaptic release | Fear memory consolidation in fear conditioning test↓ | [75] | |
| VB | WT mice/AAV Aldh1l1::cre and AAV pSico::DAO shRNA or AAV pSico::Aldh1a1 shRNA or AAV pSico::Best1 shRNA | Tonic GABA release↓ | Tonic GABA current↓ Temporal resolution↓ | Tactile discrimination in tactile-based NOR test↓ | [78] |
| THIP (GABAAR agonist) | Intracellular Ca2+↑ Tonic GABA release↑ | Tonic GABA current↑ Temporal resolution of stimulation-evoked TC neuronal firing↑ | Tactile discrimination in tactile-based NOR test↑ | ||
| LHb | WT rats/AAV GfaABC1D::Kir4.1 | RMP↓ | Extracellular K+↓ Neuronal RMP↓ Neuronal bursting↑ | Immobility in FST↑ Sucrose preference in SPT↓ | [79] |
| WT rats/AAV H1::Kir4.1-shRNA or AAV GfaABC1D::dnKir4.1 | Neuronal bursting↓ | Immobility in FST↓ Bar pressing in learned helplessness test↑ Sucrose preference in SPT↑ | |||
| ARH | GFAP-creERT2;Leprfl/fl mice | Number and length of projection↓ | Number of synapses onto POMC neurons↑ Glial coverage on the perikaryal membranes of POMC↓ mIPSC in POMC and AgRP neurons↑ mEPSC in AgRP neurons↑ | Leptin-induced suppression of food intake↓ Fasting- and ghrelin-induced food intake↑ | [84] |
| WT mice/AAV GFAP::hM3Dq | n.d. | Activity of AgRP neurons↓ | Ghrelin-induced food intake↓ Leptin-induced suppression of food intake↓ | [85] | |
| WT mice/AAV GFAP::hM4Di | n.d. | n.d. | Ghrelin-induced food intake↑ Leptin-induced suppression of food intake↓ | ||
| WT mice/DPCPX (adenosine A1R antagonist) | n.d. | Activity of AgRP neurons↑ | Ghrelin-induced food intake↑ | ||
| SCN | Aldh1L1-cre;Bmal1fl/fl mice | n.d. | Altered circadian oscillation | Altered circadian wheel-running activity | [89] |
| GLAST-creERT2;Bmal1fl/fl mice | GAT-1, GAT-3↓ | Extracellular GABA level↑ Altered circadian oscillations of clock genes in neurons | Altered circadian wheel-running activity Cognitive deficits in NOR and SOL tests↑ | [90] | |
| Cry1/2−/− mice/AAV GFAP::cre + AAV Cry1:: FLEX-Cry1 | Recovery of TTFL | Recovery of circadian oscillation of extracellular glutamate | Recovery of circadian locomotor activity | [91] | |
| VTA | WT mice/AAV GfaABC1D::ChR2 | Na+, H+↑ K+↓ | GABAergic neuronal activity↑ Dopaminergic neuronal activity↓ | Real-time avoidance behavior in RTPP assay↑ Learned avoidance behavior in CPA assay↑ | [95] |
| GLT-1fl/fl mice/ AAV GfaABC1D::ChR2 + AAV GfaABC1D::cre | n.d. | ChR2-induced increase in GABAergic neuronal activity↓ Dopaminergic neuronal activity↑ | ChR2-induced increase in learned avoidance behavior in CPA assay↓ | ||
| PAG | WT mice/L-AAA (astrocytic cytotoxin) or SP600125 (JNK inhibitor) | BCP-induced GFAP upregulation↓ | n.d. | BCP-induced mechanical allodynia↓ | [98] |
| PreBötC | WT rats/AAV sGFAP::hM3Dq | Intracellular Ca2+↑ Vesicle fusion events↑ ATP release↑ | n.d. | Resting respiratory rate↑ Frequency of sighs↑ | [110] |
| WT rats/AAV sGFAP::dnSNARE or Tetanus toxin light chain (exocytosis inhibitor) | Vesicle fusion events↓ | n.d. | Resting respiratory rate↓ Frequency of sighs↓ | ||
| CB | GLAST-creERT2; Gria1fl/fl;Gria4fl/fl mice | BG process retraction | EPSC frequency of PC↓ PF-PC synapse density↓ | Fine motor coordination in ELT and PECT↓ | [120] |
| Best1−/− mice or MAOB−/− mice or WT mice/selegiline (MAOB inhibitor) | Tonic GABA release↓ | Excitability of GCs↑ PF-PC synaptic transmission↑ | Motor coordination in rotarod test↑ | [121] | |
| GFAP-MAOB TG mice | Tonic GABA release↑ | Excitability of GCs↓ PF-PC synaptic transmission↓ | Motor coordination in rotarod test↓ | ||
| Spinal cord | GFAP-cre;Stat3fl/fl mice or WT mice/AG490 (JAK inhibitor) | DCP-induced GFAP upregulation↓ DCP-induced LCN2 upregulation↓ | DCP-induced neuronal excitation↓ | DCP-induced scratching↓ | [122] |
| Hoxb8-cre;GLT-1fl/fl mice | n.d. | n.d. | PSL-induced mechanical allodynia↑ | [124] | |
| WT rats/AAV GfaABC1D::ChR2 | ATP release↑ | Adenosine A1R↑ Excitability of GABAergic interneurons↓ Excitability of NK1R+ projection neurons↑ | Mechanical allodynia↑ Thermal hyperalgesia↑ SNI-induced mechanical allodynia↑ | [128] | |
| Behavior | Target Region | Manipulation | Major Phenotype | Ref |
|---|---|---|---|---|
| Cognition | PFC | WT mice/L-AAA (astrocyte-specific cytotoxin) | Spatial memory in Morris water maze test↓ Reversal learning in attentional set-shifting task↓ | [35] |
| ACC | GFAP-cre mice/ AAV CBA::FLEX-ChR2 | Non-rapid eye movement sleep↓ | [44] | |
| HP | WT mice/AAV GFAP::hM3Dq | Spatial memory in T-maze test↑ Contextual memory in fear conditioning test↑ | [47] | |
| WT mice/AAV GFAP::hM4Di | Remote contextual memory in fear conditioning test↓ | [48] | ||
| IL-1r−/− mice | Spatial memory in Morris water maze test↓ Contextual memory in fear conditioning test↓ | [50] | ||
| IL-1r−/− mice/transplantation of WT NPCs | Spatial memory in Morris water maze test↑ Contextual memory in fear conditioning test↑ | [50] | ||
| GFAP-tTA;tetO-dnSNARE mice | Sleep deprivation-induced spatial memory impairment in SOR test↓ | [49] | ||
| WT mice/CPT (adenosine A1 receptor antagonist) | Sleep deprivation-induced spatial memory impairment in SOR test↓ | [49] | ||
| AQP4−/− mice | Object recognition memory in object placement test↓ | [166] | ||
| GFAP-cre;Cx43fl/fl;Cx30−/− mice | Spatial memory in SOR test↓ | [52] | ||
| AMY | WT mice/AAV GFAP::hM3Dq | Fear memory formation in fear conditioning test↓ | [73] | |
| WT mice/DHK (GLT-1 inhibitor) | Freezing behavior in fear conditioning test↑ | [74] | ||
| WT mice/TAT-Cx43L2 (Cx43-hemichannel blocker) | Fear memory consolidation in fear conditioning test↓ | [75] | ||
| SCN | GLAST-creERT2;Bmal1fl/fl mice | Altered circadian wheel-running activity Cognitive deficits in NOR and SOL tests↑ | [90] | |
| VTA | WT mice/AAV GfaABC1D::ChR2 | Real-time avoidance behavior in RTPP assay↑ Learned avoidance behavior in CPA assay↑ | [95] | |
| GLT-1fl/fl mice/ AAV GfaABC1D: ChR2 + AAV GfaABC1D: cre | ChR2-induced increase in learned avoidance behavior in CPA assay↓ | [95] | ||
| Circadian rhythm | SCN | Aldh1L1-cre;Bmal1fl/fl mice | Altered circadian wheel-running activity | [89] |
| GLAST-creERT2;Bmal1fl/fl mice | Altered circadian wheel-running activity | [90] | ||
| Cry1/2−/− mice/AAV GFAP::cre + AAV Cry1:: FLEX-Cry1 | Recovery of circadian locomotor activity | [91] | ||
| Emotion | PFC | WT mice/L-AAA | Sucrose preference in SPT↓ Latency to feed in novelty suppressed feeding test↑ Immobility in FST↑ Escape latency in active avoidance test↑ | [25] |
| WT mice/DHK (GLT-1 inhibitor) | Intracranial self-stimulation threshold↑ Latency to begin drinking sucrose in sucrose intake test↑ | [26] | ||
| WT mice/ATP or ATP-γ-S | CSDS-induced social avoidance in SIT↓ CSDS-induced immobility in FST↓ CMS-induced fur condition↓ Post-CMS sucrose preference in SPT↑ | [24] | ||
| IP3R2−/− mice | Immobility in FST↑ Fur condition↓ Sucrose consumption in SPT↓ | [24] | ||
| GFAP-tTA;tetO-dnSNARE mice | Immobility in FST↑ | [24] | ||
| GFAP-tTA;tetO-MrgprA1 mice | Immobility in FST↓ Social avoidance in SIT↓ | [24] | ||
| STR | WT mice/AAV GfaABC1D::hPMCA2w/b | Duration of self-grooming↑ Travel distance in open field test↓ Time spent in center in open field test↓ | [55] | |
| WT mice/AAV GfaABC1D::hM4Di | Travel distance in open field test↑ Investigatory activity in light/dark open field test↓ Exploration of novel object in NOR test↓ | [56] | ||
| AMY | WT mice/DHK (GLT-1 inhibitor) | Intracranial self-stimulation threshold↑ Time spent in open arms in elevated plus maze test↓ | [74] | |
| LHb | WT rats/AAV GfaABC1D::Kir4.1 | Immobility in FST↑ Sucrose preference in SPT↓ | [79] | |
| WT rats/AAV H1::Kir4.1-shRNA or AAV GfaABC1D::dnKir4.1 | Immobility in FST↓ Bar pressing in learned helplessness test↑ Sucrose preference in SPT↑ | [79] | ||
| Motor skill | MC | GLAST-creER;IP3R2fl/fl mice | Motor-skill learning in forelimb reaching task↓ | [40] |
| WT mice/fluorocitrate (astrocyte metabolic inhibitor) | Motor-skill learning in forelimb reaching task↓ | [40] | ||
| HP | GFAP-cre;Cx43fl/fl;Cx30−/− mice | Motor coordination in rotarod and balance beam assays↓ | [52] | |
| CB | GLAST-creERT2;Gria1fl/fl;Gria4fl/fl mice | Fine motor coordination in ELT and PECT↓ | [120] | |
| Best1−/− mice or MAOB−/− mice or WT mice/selegiline (MAOB inhibitor) | Motor coordination in rotarod test↑ | [121] | ||
| GFAP-MAOB TG mice | Motor coordination in rotarod test↓ | [121] | ||
| Sensory processing | OB | Aldh1L1-cre mice/ AAV GFAP::FLEX-hM3Dq | Odor detection accuracy in Go/No-Go task↑ | [32] |
| Aldh1L1-cre mice/ AAV GFAP::FLEX-hM4Di | Odor detection accuracy in Go/No-Go task↓ | [32] | ||
| SC | IP3R2−/− mice | PSL-induced mechanical allodynia↓ | [41] | |
| WT mice/BAPTA-AM (Ca2+ chelator) | PSL-induced mechanical allodynia↓ | [41] | ||
| Fluoroacetate (astrocyte metabolic inhibitor) | PSL-induced mechanical allodynia↓ | [41] | ||
| ACC | L-AAA | CFA-induced place escape/avoidance↓ | [27] | |
| WT mice/SNAP-5114 (GAT-3 inhibitor) | PSL-induced sleep disturbance↓ | [45] | ||
| VB | WT mice/AAV Aldh1l1::cre and AAV pSico::DAO shRNA or AAV pSico::Aldh1a1 shRNA or AAV pSico::Best1 shRNA | Tactile discrimination in tactile-based NOR test↓ | [78] | |
| THIP (GABAAR agonist) | Tactile discrimination in tactile-based NOR test↑ | [78] | ||
| PAG | WT mice/L-AAA (astrocytic cytotoxin) or SP600125 (JNK inhibitor) | BCP-induced mechanical allodynia↓ | [98] | |
| Spinal cord | GFAP-cre;Stat3fl/fl mice or WT mice/AG490 (JNK inhibitor) | DCP-induced scratching↓ | [122] | |
| Hoxb8-cre;GLT-1fl/fl mice | PSL-induced mechanical allodynia↑ | [124] | ||
| WT rats/AAV GfaABC1D::ChR2 | Mechanical allodynia Thermal hyperalgesia SNI-induced mechanical allodynia↑ | [128] | ||
| Appetite | ARH | GFAP-creERT2;Leprfl/fl mice | Leptin-induced suppression of food intake↓ Fasting- and ghrelin-induced food intake↑ | [84] |
| WT mice/AAV GFAP::hM3Dq | Ghrelin-induced food intake↓ Leptin-induced suppression of food intake↑ | [85] | ||
| WT mice/AAV GFAP::hM4Di | Ghrelin-induced food intake↑ Leptin-induced suppression of food intake↓ | [85] | ||
| WT mice/DPCPX (adenosine A1R antagonist) | Ghrelin-induced food intake↑ | [85] |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allen, N.J.; Lyons, D.A. Glia as architects of central nervous system formation and function. Science 2018, 362, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Zuo, Y.X.; Jiang, R.T. Astrocyte morphology: Diversity, plasticity, and role in neurological diseases. CNS Neurosci. Ther. 2019, 25, 665–673. [Google Scholar] [CrossRef]
- Yu, X.; Nagai, J.; Khakh, B.S. Improved tools to study astrocytes. Nat. Rev. Neurosci. 2020, 21, 121–138. [Google Scholar] [CrossRef]
- Chaboub, L.S.; Deneen, B. Developmental origins of astrocyte heterogeneity: The final frontier of CNS development. Dev. Neurosci. 2012, 34, 379–388. [Google Scholar] [CrossRef]
- Oberheim, N.A.; Goldman, S.A.; Nedergaard, M. Heterogeneity of astrocytic form and function. Methods Mol. Biol. 2012, 814, 23–45. [Google Scholar] [CrossRef]
- Bayraktar, O.A.; Fuentealba, L.C.; Alvarez-Buylla, A.; Rowitch, D.H. Astrocyte development and heterogeneity. Cold Spring Harb. Perspect. Biol. 2015, 7, a020362. [Google Scholar] [CrossRef]
- Zhang, Y.; Barres, B.A. Astrocyte heterogeneity: An underappreciated topic in neurobiology. Curr. Opin. Neurobiol. 2010, 20, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, A.; Hochgerner, H.; Lönnerberg, P.; Johnsson, A.; Memic, F.; van der Zwan, J.; Häring, M.; Braun, E.; Borm, L.; Manno, G.L.; et al. Molecular architecture of the mouse nervous system. Cell 2018, 174, 999–1014. [Google Scholar] [CrossRef]
- Khakh, B.S. Astrocyte-Neuron interactions in the striatum: Insights on identity, form, and function. Trends Neurosci. 2019, 42, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.J.; Philips, T.; Kim, N.; Dastgheyb, R.; Chen, Z.; Hsieh, Y.-C.; Daigle, J.G.; Datta, M.; Chew, J.; Vidensky, S.; et al. Molecularly defined cortical astroglia subpopulation modulates neurons via secretion of Norrin. Nat. Neurosci. 2019, 22, 741–752. [Google Scholar] [CrossRef]
- Tian, G.F.; Takano, T.; Lin, J.H.C.; Wang, X.; Bekar, L.; Nedergaard, M. Imaging of cortical astrocytes using 2-photon laser scanning microscopy in the intact mouse brain. Adv. Drug Deliv. Rev. 2006, 58, 773–787. [Google Scholar] [CrossRef]
- Tsacopoulos, M.; Magistretti, P.J. Metabolic coupling between glia and neurons. J. Neurosci. 1996, 16, 877–885. [Google Scholar] [CrossRef]
- Anderson, C.M.; Nedergaard, M. Astrocyte-mediated control of cerebral microcirculation. Trends Neurosci. 2003, 26, 340–344. [Google Scholar] [CrossRef]
- Janzer, R.C.; Raff, M.C. Astrocytes induce blood–brain barrier properties in endothelial cells. Nature 1987, 325, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Abbott, N.; Rönnbäck, L.; Hansson, E. Astrocyte-Endothelial interactions at the blood-brain barrier. Nat. Rev. Neurosci. 2006, 7, 41–53. [Google Scholar] [CrossRef]
- Simard, M.; Nedergaard, M. The neurobiology of glia in the context of water and ion homeostasis. Neuroscience 2004, 129, 877–896. [Google Scholar] [CrossRef]
- Cornell-Bell, A.H.; Finkbeiner, S.M. Ca2+ waves in astrocytes. Cell Calcium 1991, 12, 185–204. [Google Scholar] [CrossRef]
- Porter, J.T.; McCarthy, K.D. Astrocytic neurotransmitter receptors in situ and in vivo. Prog. Neurobiol. 1997, 51, 439–455. [Google Scholar] [CrossRef]
- Farhy-Tselnicker, I.; Allen, N.J. Astrocytes, neurons, synapses: A tripartite view on cortical circuit development. Neural Dev. 2018, 13, 7. [Google Scholar] [CrossRef] [PubMed]
- Guerra-Gomes, S.; Sousa, N.; Pinto, L.; Oliveira, J.F. Functional Roles of astrocyte calcium elevations: From synapses to behavior. Front. Cell. Neurosci. 2018, 11, 427. [Google Scholar] [CrossRef] [PubMed]
- Fiacco, T.A.; McCarthy, K.D. Multiple lines of evidence indicate that gliotransmission does not occur under physiological conditions. J. Neurosci. 2018, 38, 3–13. [Google Scholar] [CrossRef]
- Savtchouk, I.; Volterra, A. Gliotransmission: Beyond black-and-white. J. Neurosci. 2018, 38, 14–25. [Google Scholar] [CrossRef]
- Oliveira, J.F.; Sardinha, V.M.; Guerra-Gomes, S.; Araque, A.; Sousa, N. Do stars govern our actions? Astrocyte involvement in rodent behavior. Trends Neurosci. 2015, 38, 535–549. [Google Scholar] [CrossRef]
- Cao, X.; Li, L.P.; Wang, Q.; Wu, Q.; Hu, H.H.; Zhang, M.; Fang, Y.Y.; Zhang, J.; Li, S.J.; Xiong, W.C.; et al. Astrocyte-derived ATP modulates depressive-like behaviors. Nat. Med. 2013, 19, 773–777. [Google Scholar] [CrossRef] [PubMed]
- Banasr, M.; Duman, R.S. Glial loss in the prefrontal cortex is sufficient to induce depressive-like behaviors. Biol. Psychiatry 2008, 64, 863–870. [Google Scholar] [CrossRef] [PubMed]
- John, C.S.; Smith, K.L.; Veer, A.V.; Gompf, H.S.; Carlezon, W.A., Jr.; Cohen, B.M.; Öngür, D.; Bechtholt-Gompf, A.J. Blockade of astrocytic glutamate uptake in the prefrontal cortex induces anhedonia. Neuropsychopharmacology 2012, 37, 2467–2475. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.L.; Dong, Y.L.; Zhang, Z.J.; Cao, D.L.; Xu, J.; Hui, J.; Zhu, L.; Gao, Y.J. Activation of astrocytes in the anterior cingulate cortex contributes to the affective component of pain in an inflammatory pain model. Brain Res. Bull. 2012, 87, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Chai, H.; Diaz-Castro, B.; Shigetomi, E.; Monte, E.; Octeau, J.C.; Yu, X.; Cohn, W.; Rajendran, P.S.; Vondriska, T.M.; Whitelegge, J.P.; et al. Neural circuit-specialized astrocytes: Transcriptomic, proteomic, morphological, and functional evidence. Neuron 2017, 95, 531–549.e9. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Lu, T.Y.; Chai, H.; Xu, J.; Huang, B.S.; Golshani, P.; Coppola, G.; Khakh, B.S. New transgenic mouse lines for selectively targeting astrocytes and studying calcium signals in astrocyte processes in situ and in vivo. Neuron 2016, 92, 1181–1195. [Google Scholar] [CrossRef] [PubMed]
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef]
- Nagayama, S.; Homma, R.; Imamura, F. Neuronal organization of olfactory bulb circuits. Front. Neural Circuits 2014, 8, 98. [Google Scholar] [CrossRef]
- Ung, K.; Tepe, B.; Pekarek, B.; Arenkiel, B.R.; Deneen, B. Parallel astrocyte calcium signaling modulate olfactory bulb responses. J. Neurosci. Res. 2020, 98, 1605–1618. [Google Scholar] [CrossRef] [PubMed]
- Ongür, D.; Price, J.L. The organization of networks within the orbital and medial prefrontal cortex of rats, monkeys and humans. Cereb. Cortex 2000, 10, 206–219. [Google Scholar] [CrossRef]
- Croxson, P.L.; Johansen-Berg, H.; Behrens, T.E.; Robson, M.D.; Pinsk, M.A.; Gross, C.G.; Richter, W.; Richter, M.C.; Kastner, S.; Rushworth, M.F. Quantitative investigation of connections of the prefrontal cortex in the human and macaque using probabilistic diffusion tractography. J. Neurosci. 2005, 25, 8854–8866. [Google Scholar] [CrossRef]
- Lima, A.; Sardinha, V.M.; Oliveira, A.F.; Reis, M.; Mota, C.; Silva, M.A.; Marques, F.; Cerqueira, J.J.; Pinto, L.; Sousa, N.; et al. Astrocyte pathology in the prefrontal cortex impairs the cognitive function of rats. Mol. Psychiatry 2014, 19, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Danbolt, N.C. Glutamate uptake. Prog. Neurobiol. 2001, 65, 1–105. [Google Scholar] [CrossRef]
- Peterson, A.R.; Binder, D.K. Astrocyte glutamate uptake and signaling as novel targets for antiepileptogenic therapy. Front. Neurol. 2020, 11, 1006. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Cao, X.; Zeng, Y.; Qin, X.; Zhu, M.; Ren, J.; Wu, Z.; Huang, Q.; Zhang, Y.; Wang, M.; et al. Astrocytic epoxyeicosatrienoic acid signaling in the medial prefrontal cortex modulates depressive-like behaviors. J. Neurosci. 2019, 39, 4606–4623. [Google Scholar] [CrossRef]
- Erickson, E.K.; DaCosta, A.J.; Mason, S.C.; Blednov, Y.A.; Mayfield, R.D.; Harris, R.A. Cortical astrocytes regulate ethanol consumption and intoxication in mice. Neuropsychopharmacology 2020, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Padmashri, R.; Suresh, A.; Boska, M.D.; Dunaevsky, A. Motor-skill learning is dependent on astrocytic activity. Neural Plast. 2015, 2015, 938023. [Google Scholar] [CrossRef]
- Kim, S.K.; Hayashi, H.; Ishikawa, T.; Shibata, K.; Shigetomi, E.; Shinozaki, Y.; Inada, H.; Roh, S.E.; Kim, S.J.; Lee, G.; et al. Cortical astrocytes rewire somatosensory cortical circuits for peripheral neuropathic pain. J. Clin. Investig. 2016, 126, 1983–1997. [Google Scholar] [CrossRef]
- Cao, H.; Gao, Y.-J.; Ren, W.-H.; Li, T.-T.; Duan, K.-Z.; Cui, Y.-H.; Cao, X.-H.; Zhao, Z.-Q.; Ji, R.-R.; Zhang, Y.-Q. Activation of extracellular signal-regulated kinase in the anterior cingulate cortex contributes to the induction and expression of affective pain. J. Neurosci. 2009, 29, 3307–3321. [Google Scholar] [CrossRef]
- Gu, D.; Zhou, M.; Han, C.; Lei, D.; Xie, S.; Yuan, Y.; Ma, T. Preoperative anxiety induces chronic postoperative pain by activating astrocytes in the anterior cingulate cortex region. Rev. Assoc. Med. Bras. 2019, 65, 1174–1180. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, A.; Hamada, A.; Suhara, Y.; Kawabe, R.; Yanase, M.; Kuzumaki, N.; Narita, M.; Matsui, R.; Okano, H.; Narita, M. Astrocytic activation in the anterior cingulate cortex is critical for sleep disorder under neuropathic pain. Synapse 2014, 68, 235–247. [Google Scholar] [CrossRef]
- Narita, M.; Niikura, K.; Nanjo-Niijura, K.; Narita, M.; Furuya, M.; Yamashita, A.; Saeki, M.; Matsushima, Y.; Imai, S.; Shimizu, T.; et al. Sleep disturbances in a neuropathic pain-like condition in the mouse are associated with altered GABAergic transmission in the cingulate cortex. Pain 2011, 152, 1358–1372. [Google Scholar] [CrossRef] [PubMed]
- Jarrard, L.E. On the role of the hippocampus in learning and memory in the rat. Behav. Neural Biol. 1993, 60, 9–26. [Google Scholar] [CrossRef]
- Adamsky, A.; Kol, A.; Kreisel, T.; Doron, A.; Ozeri-Engelhard, N.; Melcer, T.; Refaeli, R.; Horn, H.; Regev, L.; Groysman, M.; et al. Astrocytic activation generates de novo neuronal potentiation and memory enhancement. Cell 2018, 174, 59–71.e14. [Google Scholar] [CrossRef] [PubMed]
- Kol, A.; Adamsky, A.; Groysman, M.; Kreisel, T.; London, M.; Goshen, I. Astrocytes contribute to remote memory formation by modulating hippocampal-cortical communication during learning. Nat. Neurosci. 2020, 23, 1229–1239. [Google Scholar] [CrossRef]
- Florian, C.; Vecsey, C.G.; Halassa, M.M.; Haydon, P.G.; Abel, T. Astrocyte-Derived adenosine and A1 receptor activity contribute to sleep loss-induced deficits in hippocampal synaptic plasticity and memory in mice. J. Neurosci. 2011, 31, 6956–6962. [Google Scholar] [CrossRef] [PubMed]
- Menachem-Zidon, O.B.; Avital, A.; Ben-Menahem, Y.; Goshen, I.; Kreisel, T.; Shmueli, E.M.; Segal, M.; Hur, T.B.; Yirmiya, R. Astrocytes support hippocampal-dependent memory and long-term potentiation via interleukin-1 signaling. Brain Behav. Immun. 2011, 25, 1008–1016. [Google Scholar] [CrossRef]
- Scharfman, H.E.; Binder, D.K. Aquaporin-4 water channels and synaptic plasticity in the hippocampus. Neurochem. Int. 2013, 63, 702–711. [Google Scholar] [CrossRef] [PubMed]
- Lutz, S.E.; Zhao, Y.; Gulinello, M.; Lee, S.C.; Raine, C.S.; Brosnan, C.F. Deletion of astrocyte connexins 43 and 30 leads to a dysmyelinating phenotype and hippocampal CA1 vacuolation. J. Neurosci. 2009, 29, 7743–7752. [Google Scholar] [CrossRef] [PubMed]
- Graybiel, A.M. Habits, rituals, and the evaluative brain. Annu. Rev. Neurosci. 2008, 31, 359–387. [Google Scholar] [CrossRef]
- Graybiel, A.M.; Grafton, S.T. The striatum: Where skills and habits meet. Cold Spring Harb. Perspect. Biol. 2015, 7, a021691. [Google Scholar] [CrossRef]
- Yu, X.; Taylor, A.M.W.; Nagai, J.; Golshani, P.; Evans, C.J.; Coppola, G.; Khakh, B.S. Reducing astrocyte calcium signaling in vivo alters striatal microcircuits and causes repetitive behavior. Neuron 2018, 99, 1170–1187.e9. [Google Scholar] [CrossRef]
- Nagai, J.; Rajbhandari, A.K.; Gangwani, M.R.; Hachisuka, A.; Coppola, G.; Masmanidis, S.C.; Fanselow, M.S.; Khakh, B.S. Hyperactivity with disrupted attention by activation of an astrocyte synaptogenic cue. Cell 2019, 177, 1280–1292.e20. [Google Scholar] [CrossRef]
- Lüscher, C. The emergence of a circuit model for addiction. Annu. Rev. Neurosci. 2016, 39, 257–276. [Google Scholar] [CrossRef] [PubMed]
- Corkrum, M.; Covelo, A.; Lines, J.; Bellocchio, L.; Pisansky, M.; Loke, K.; Quintana, R.; Rothwell, P.E.; Lujan, R.; Marsicano, G.; et al. Dopamine-Evoked synaptic regulation in the nucleus accumbens requires astrocyte activity. Neuron 2020, 105, 1036–1047.e5. [Google Scholar] [CrossRef]
- Fendt, M.; Fenselow, M.S. The neuroanatomical and neurochemical basis of conditioned fear. Neurosci. Biobehav. Rev. 1999, 23, 743–760. [Google Scholar] [CrossRef]
- LeDoux, J.E. Emotion circuits in the brain. Annu. Rev. Neurosci. 2000, 23, 155–184. [Google Scholar] [CrossRef]
- Balderston, N.L.; Schultz, D.H.; Hopkins, L.; Helmstetter, F.J. Functionally distinct amygdala subregions identified using DTI and high-resolution fMRI. Soc. Cogn. Affect. Neurosci. 2015, 10, 1615–1622. [Google Scholar] [CrossRef] [PubMed]
- LeDoux, J.E.; Farb, C.; Ruggiero, D.A. Topographic organization of neurons in the acoustic thalamus that project to the amygdala. J. Neurosci. 1990, 10, 1043–1054. [Google Scholar] [CrossRef]
- McDonald, A.J. Cortical pathways to the mammalian amygdala. Prog. Neurobiol. 1998, 55, 257–332. [Google Scholar] [CrossRef]
- Duvarci, S.; Pare, D. Amygdala microcircuits controlling learned fear. Neuron 2014, 82, 966–980. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, I.; Humeau, Y.; Grenier, F.; Ciocchi, S.; Herry, C.; Lüthi, A. Amygdala inhibitory circuits and the control of fear memory. Neuron 2009, 62, 757–771. [Google Scholar] [CrossRef]
- Tovote, P.; Esposito, M.S.; Botta, P.; Chaudun, F.; Fadok, J.P.; Markovic, M.; Wolff, S.B.E.; Ramakrishnan, C.; Fenno, L.; Deisseroth, K.; et al. Midbrain circuits for defensive behaviour. Nature 2016, 534, 206–212. [Google Scholar] [CrossRef]
- Sheline, Y.I.; Gado, M.H.; Price, J.L. Amygdala core nuclei volumes are decreased in recurrent major depression. Neuroreport 1998, 9, 2023–2028. [Google Scholar] [CrossRef] [PubMed]
- Mervaala, E.; Könönen, J.F.M.; Valkonen-Korhonen, M.; Vainio, P.; Partanen, K.; Partanen, J.; Tiihonen, J.; Viinamäki, H.; Karjalainen, A.K.; Lehtonen, J. Quantitative MRI of the hippocampus and amygdala in severe depression. Psychol. Med. 2000, 30, 117–125. [Google Scholar] [CrossRef]
- Blumberg, H.P.; Kaufman, J.; Martin, A.; Whiteman, R.; Zhang, J.H.; Gore, J.C.; Charney, D.S.; Krystal, J.H.; Peterson, B.S. Amygdala and hippocampal volumes in adolescents and adults with bipolar disorder. Arch. Gen. Psychatry 2003, 60, 1201–1208. [Google Scholar] [CrossRef]
- Rosso, I.M.; Killgore, W.D.S.; Cintron, C.M.; Gruber, S.A.; Tohen, M.; Yurgelun-Todd, D.A. Reduced amygdala volumes in first-episode bipolar disorder and correlation with cerebral white matter. Biol. Psychiatry 2007, 61, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Bowley, M.P.; Drevets, W.C.; Ongür, D.; Price, J.L. Low glial numbers in the amygdala in major depressive disorder. Biol. Psychiatry 2002, 52, 404–412. [Google Scholar] [CrossRef]
- Gosselin, R.-D.; Gibney, S.; O’Malley, D.; Dinan, T.G.; Cryan, J.F. Region specific decrease in glial fibrillary acidic protein immunoreactivity in the brain of a rat model of depression. Neuroscience 2009, 159, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Martin-Fernandez, M.; Jamison, S.; Robin, L.M.; Zhao, Z.; Martin, E.D.; Aguilar, J.; Benneyworth, M.A.; Marsicano, G.; Araque, A. Synapse-Specific astrocyte gating of amygdala-related behavior. Nat. Neurosci. 2017, 20, 1540–1548. [Google Scholar] [CrossRef]
- John, C.S.; Sypek, E.I.; Carlezon, W.A.; Cohen, B.M.; Öngür, D.; Bechtholt, A.J. Blockade of the GLT-1 transporter in the central nucleus of the amygdala induces both anxiety and depressive-like symptoms. Neuropsychopharmacology 2015, 40, 1700–1708. [Google Scholar] [CrossRef]
- Stehberg, J.; Moraga-Amaro, R.; Salazar, C.; Becerra, A.; Echeverría, C.; Orellana, J.A.; Bultynck, G.; Ponsaerts, R.; Leybaert, L.; Simon, F.; et al. Release of gliotransmitters through astroglial connexin 43 hemichannels is necessary for fear memory consolidation in the basolateral amygdala. FASEB J. 2012, 26, 3649–3657. [Google Scholar] [CrossRef]
- Boury-Jamot, B.; Carrard, A.; Martin, J.L.; Halfon, O.; Magistretti, P.J.; Boutrel, B. Disrupting astrocyte-neuron lactate transfer persistently reduces conditioned responses to cocaine. Mol. Psychiatry 2016, 21, 1070–1076. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Tao, Y.; Guo, X.; Cheng, D.; Wang, F.; Liu, X.; Ma, L. Fear conditioning downregulates Rac1 activity in the basolateral amygdala astrocytes to facilitate the formation of fear memory. Front. Mol. Neurosci. 2017, 10, 396. [Google Scholar] [CrossRef] [PubMed]
- Kwak, H.; Koh, W.; Kim, S.; Song, K.; Shin, J.-I.; Lee, J.M.; Lee, E.H.; Bae, J.Y.; Ha, G.E.; Oh, J.-E.; et al. Astrocytes control sensory acuity via tonic inhibition in the thalamus. Neuron 2020, 108, 691–706.e10. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Yang, Y.; Ni, Z.; Dong, Y.; Cai, G.; Foncelle, A.; Ma, S.; Sang, K.; Tang, S.; Li, Y.; et al. Astroglial Kir4.1 in the lateral habenula drives neuronal bursts in depression. Nature 2018, 554, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.G.; Powell, T.P. An electron microscopic study of the mode of termination of cortico-thalamic fibres within the sensory relay nuclei of the thalamus. Proc. R. Soc. Lond. B Biol. Sci. 1969, 172, 173–185. [Google Scholar] [CrossRef]
- Bourassa, J.; Pinault, D.; Deschênes, M. Corticothalamic projections from the cortical barrel field to the somatosensory thalamus in rats: A single-fibre study using biocytin as an anterograde tracer. Eur. J. Neurosci. 1995, 7, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Heimer, L. Thalamus. In The Human Brain and Spinal Cord; Springer: New York, NY, USA, 1983; pp. 331–335. [Google Scholar] [CrossRef]
- Reis, W.L.; Yi, C.-X.; Gao, Y.; Tschöp, M.H.; Stern, J.E. Brain innate immunity regulates hypothalamic arcuate neuronal activity and feeding behavior. Endocrinology 2015, 156, 1303–1315. [Google Scholar] [CrossRef]
- Kim, J.G.; Suyama, S.; Koch, M.; Jin, S.; Argente-Arizon, P.; Argente, J.; Liu, Z.-W.; Zimmer, M.R.; Jeong, J.K.; Szigeti-Buck, K.; et al. Leptin signaling in astrocytes regulates hypothalamic neuronal circuits and feeding. Nat. Neurosci. 2014, 17, 908–910. [Google Scholar] [CrossRef]
- Yang, L.; Qi, Y.; Yang, Y. Astrocytes control food intake by inhibiting AGRP neuron activity via adenosine A1 receptors. Cell Rep. 2015, 11, 798–807. [Google Scholar] [CrossRef] [PubMed]
- Douglass, J.D.; Dorfman, M.D.; Fasnacht, R.; Shaffer, L.D.; Thaler, J.P. Astrocyte IKKβ/NF-κB signaling is required for diet-induced obesity and hypothalamic inflammation. Mol. Metab. 2017, 6, 366–373. [Google Scholar] [CrossRef]
- Weaver, D.R. He suprachiasmatic nucleus: A 25-year retrospective. J. Biol. Rhythm. 1998, 13, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Kwon, I.; Choe, H.K.; Son, G.H.; Kim, K. Mammalian molecular clocks. Exp. Neurobiol. 2011, 20, 18–28. [Google Scholar] [CrossRef]
- Tso, C.F.; Simon, T.; Greenlaw, A.C.; Puri, T.; Mieda, M.; Herzog, E.D. Astrocytes Regulate Daily Rhythms in the suprachiasmatic nucleus and behavior. Curr. Biol. 2017, 27, 1055–1061. [Google Scholar] [CrossRef]
- Barca-Mayo, O.; Pons-Espinal, M.; Follert, P.; Armirotti, A.; Berdondini, L.; Tonelli, D.P. Astrocyte deletion of Bmal1 alters daily locomotor activity and cognitive functions via GABA signalling. Nat. Commun. 2017, 8, 14336. [Google Scholar] [CrossRef]
- Brancaccio, M.; Edwards, M.D.; Patton, A.P.; Smyllie, N.J.; Chesham, J.E.; Maywood, E.S.; Hastings, M.H. Cell-Autonomous clock of astrocytes drives circadian behavior in mammals. Science 2019, 363, 187–192. [Google Scholar] [CrossRef]
- Hastings, M.H.; Maywood, E.S.; Brancaccio, M. Generation of circadian rhythms in the suprachiasmatic nucleus. Nat. Rev. Neurosci. 2018, 19, 453–469. [Google Scholar] [CrossRef]
- Wise, R.A. Dopamine, learning and motivation. Nat. Rev. Neurosci. 2004, 5, 483–494. [Google Scholar] [CrossRef]
- Wise, R.A. Roles for nigrostriatal—Not just mesocorticolimbic—Dopamine in reward and addiction. Trends Neurosci. 2009, 32, 517–524. [Google Scholar] [CrossRef]
- Gomez, J.A.; Perkins, J.M.; Beaudoin, G.M.; Cook, N.B.; Quraishi, S.A.; Szoeke, E.A.; Thangamani, K.; Tschumi, C.W.; Wanat, M.J.; Maroof, A.M.; et al. Ventral tegmental area astrocytes orchestrate avoidance and approach behavior. Nat. Commun. 2019, 10, 1455. [Google Scholar] [CrossRef] [PubMed]
- Fields, H.L. Central nervous system mechanisms of pain modulation. Pain 1996, 2, 44–46. [Google Scholar]
- Ni, H.-D.; Yao, M.; Huang, B.; Xu, L.-S.; Zheng, Y.; Chu, Y.-X.; Wang, H.-Q.; Liu, M.-J.; Xu, S.-J.; Li, H.-B. Glial activation in the periaqueductal gray promotes descending facilitation of neuropathic pain through the p38 MAPK signaling pathway. J. Neurosci. Res. 2016, 94, 50–61. [Google Scholar] [CrossRef]
- Ni, H.-D.; Xu, L.S.; Wang, Y.; Li, H.; An, K.; Liu, M.; Liu, Q.; Deng, H.; He, Q.; Huang, B.; et al. Astrocyte activation in the periaqueductal gray promotes descending facilitation to cancer-induced bone pain through the JNK MAPK signaling pathway. Mol. Pain 2019, 15. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.; Wang, Y.; An, K.; Liu, Q.; Xu, L.; Zhu, C.; Deng, H.; He, Q.; Wang, T.; Xu, M.; et al. Crosstalk between NFκB-dependent astrocytic CXCL1 and neuron CXCR2 plays a role in descending pain facilitation. J. Neuroinflamm. 2019, 16, 1. [Google Scholar] [CrossRef]
- Smith, J.C.; Ellenberger, H.H.; Ballanyi, K.; Richter, D.W.; Feldman, J.L. Pre-Bötzinger complex: A brainstem region that may generate respiratory rhythm in mammals. Science 1991, 254, 726–729. [Google Scholar] [CrossRef]
- Lieske, S.P.; Thoby-Brisson, M.; Telgkamp, P.; Ramirez, J.M. Reconfiguration of the neural network controlling multiple breathing patterns: Eupnea, sighs and gasps. Nat. Neurosci. 2000, 3, 600–607. [Google Scholar] [CrossRef]
- Hayes, J.A.; Wang, X.; Del Negro, C.A. Cumulative lesioning of respiratory interneurons disrupts and precludes motor rhythms in vitro. Proc. Natl. Acad. Sci. USA 2012, 109, 8286–8291. [Google Scholar] [CrossRef]
- Wang, X.; Hayes, J.A.; REvill, A.L.; Song, H.; Kottick, A.; Vann, N.C.; LaMar, M.D.; Picardo, M.C.D.; Akins, V.T.; Funk, G.D.; et al. Laser ablation of Dbx1 neurons in the pre-Bötzinger complex stops inspiratory rhythm and impairs output in neonatal mice. eLife 2014, 3, e03427. [Google Scholar] [CrossRef] [PubMed]
- Ezure, K. Reflections on respiratory rhythm generation. Prog. Brain Res. 2004, 143, 67–74. [Google Scholar] [CrossRef]
- Feldman, J.L.A.; Del Negro, C.; Gray, P.A. Understanding the rhythm of breathing: So near, yet so far. Annu. Rev. Physiol. 2013, 75, 423–452. [Google Scholar] [CrossRef]
- Okada, Y.; Sasaki, T.; Oku, Y.; Takahashi, N.; Seki, M.; Ujita, S.; Tanaka, J.F.; Matsuki, N.; Ikegaya, Y. Preinspiratory calcium rise in putative pre-Bötzinger complex astrocytes. J. Physiol. 2012, 590, 4933–4944. [Google Scholar] [CrossRef]
- Oku, Y.; Fresemann, J.; Miwakeichi, F.; Hülsmann, S. Respiratory calcium fluctuations in low-frequency oscillating astrocytes in the pre-Bötzinger complex. Respir. Physiol. Neurobiol. 2016, 226, 11–17. [Google Scholar] [CrossRef]
- Hülsmann, S.; Oku, Y.; Zhang, W.; Richter, D.W. Metabolic coupling between glia and neurons is necessary for maintaining respiratory activity in transverse medullary slices of neonatal mouse. Eur. J. Neurosci. 2000, 12, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Huxtable, A.G.; Zwicker, J.D.; Alvares, T.S.; Ruangkittisakul, A.; Fang, X.; Hahn, L.B.; de Chaves, E.P.; Baker, G.B.; Ballanyi, K.; Funk, G.D. Glia contribute to the purinergic modulation of inspiratory rhythm-generating networks. J. Neurosci. 2010, 30, 3947–3958. [Google Scholar] [CrossRef]
- Sheikhbahaei, S.; Turovsky, E.A.; Hosford, P.S.; Hadjihambi, A.; Theparambil, S.M.; Liu, B.; Marina, N.; Teschemacher, A.G.; Kasparov, S.; Smith, J.C.; et al. Astrocytes modulate brainstem respiratory rhythm-generating circuits and determine exercise capacity. Nat. Commun. 2018, 9, 370. [Google Scholar] [CrossRef] [PubMed]
- Lorier, A.R.; Huxtable, A.G.; Robinson, D.M.; Lipski, J.; Housley, G.D.; Funk, G.D. P2Y1 receptor modulation of the pre-Bötzinger complex inspiratory rhythm generating network in vitro. J. Neurosci. 2007, 27, 993–1005. [Google Scholar] [CrossRef]
- Lorier, A.R.; Lipski, J.; Housley, G.D.; Greer, J.J.; Funk, G.D. ATP sensitivity of preBötzinger complex neurones in neonatal rat in vitro: Mechanism underlying a P2 receptor-mediated increase in inspiratory frequency. J. Physiol. 2008, 586, 1429–1446. [Google Scholar] [CrossRef] [PubMed]
- Rajani, V.; Zhang, Y.; Jalubula, V.; Rancic, V.; SheikhBahaei, S.; Zwicker, J.D.; Pagliardini, S.; Dickson, C.T.; Ballanyi, K.; Kasparov, S.; et al. Release of ATP by preBötzinger complex astrocytes contributes to the hypoxic ventilatory response via a Ca2+-dependent P2Y1 receptor mechanism. J. Physiol. 2018, 596, 3245–3269. [Google Scholar] [CrossRef]
- Ito, M. Mechanisms of motor learning in the cerebellum. Brain Res. 2000, 886, 237–245. [Google Scholar] [CrossRef]
- Wolf, U.; Rapoport, M.J.; Schweizer, T.A. Evaluating the affective component of the cerebellar cognitive affective syndrome. J. Neuropsychiatry Clin. Neurosci. 2009, 21, 245–253. [Google Scholar] [CrossRef]
- Das, G.D. Differentiation of Bergmann glia cells in the cerebellum. Brain Res. 1976, 110, 199–213. [Google Scholar] [CrossRef]
- Yuasa, S. Bergmann glial development in the mouse cerebellum as revealed by tenascin expression. Anat. Embryol. 1996, 194, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Grosche, J.; Kettenmann, H.; Reichenbach, A. Bergmann glial cells form distinct morphological structures to interact with cerebellar neurons. J. Neurosci. Res. 2002, 68, 138–149. [Google Scholar] [CrossRef]
- Spacek, J. Three-dimensional analysis of dendritic spines. Anat. Embryol. 1985, 171, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Saab, A.S.; Neumeyer, A.; Jahn, H.M.; Cupido, A.; Šimek, A.A.M.; Boele, H.-J.; Scheller, A.; Meur, K.L.; Götz, M.; Monyer, H.; et al. Bergmann glial AMPA receptors are required for fine motor coordination. Science 2012, 337, 749–753. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.; Min, J.O.; Kang, D.-S.; Kim, Y.S.; Jung, G.H.; Park, H.J.; Kim, S.; An, H.; Kwon, J.; Kim, J.; et al. Control of motor coordination by astrocytic tonic GABA release through modulation of excitation/inhibition balance in cerebellum. Proc. Natl. Acad. Sci. USA 2018, 115, 5004–5009. [Google Scholar] [CrossRef]
- Shiratori-Hayashi, M.; Koga, K.; Tozaki-Saitoh, H.; Kohro, Y.; Toyonaga, H.; Yamaguchi, C.; Hasegawa, A.; Nakahara, T.; Hachisuka, J.; Akira, S.; et al. STAT3-dependent reactive astrogliosis in the spinal dorsal horn underlies chronic itch. Nat. Med. 2015, 21, 927–931. [Google Scholar] [CrossRef]
- Liu, T.; Han, Q.; Chen, G.; Huang, Y.; Zhao, L.-X.; Berta, T.; Gao, Y.-J.; Ji, R.-R. Toll-like receptor 4 contributes to chronic itch, alloknesis and spinal astrocyte activation in male mice. Pain 2016, 157, 806–817. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Hiraoka, Y.; Ogawa, H.; Tanaka, K. Region-Specific deletions of the glutamate transporter GLT1 differentially affect nerve injury-induced neuropathic pain in mice. Glia 2018, 66, 1988–1998. [Google Scholar] [CrossRef]
- Hu, Y.; Li, W.; Lu, L.; Cai, J.; Xian, X.; Zhang, M.; Li, Q.; Li, L. An antinociceptive role for ceftriaxone in chronic neuropathic pain in rats. Pain 2010, 148, 284–301. [Google Scholar] [CrossRef]
- Peng, W.; Cotrina, M.L.; Han, X.; Yu, H.; Bekar, L.; Blum, L.; Takano, T.; Tian, G.-F.; Goldman, S.A.; Nedergaard, M. Systemic administration of an antagonist of the ATP-sensitive receptor P2X7 improves recovery after spinal cord injury. Proc. Natl. Acad. Sci. USA 2009, 106, 12489–12493. [Google Scholar] [CrossRef]
- Huang, C.; Han, X.; Li, X.; Lam, E.; Peng, W.; Lou, N.; Torres, A.; Yang, M.; Mauricio Garre, J.; Tian, G.-F.; et al. Critical role of connexin 43 in secondary expansion of traumatic spinal cord injury. J. Neurosci. 2012, 32, 3333–3338. [Google Scholar] [CrossRef] [PubMed]
- Nam, Y.; Kim, J.-H.; Kim, J.-H.; Jha, M.K.; Jung, J.Y.; Lee, M.-G.; Choi, I.-S.; Jang, I.S.; Lim, D.G.; Hwang, S.-H.; et al. Reversible induction of pain hypersensitivity following optogenetic stimulation of spinal astrocytes. Cell Rep. 2016, 17, 3049–3061. [Google Scholar] [CrossRef]
- Huang, A.Y.-S.; Woo, J.; Sardar, D.; Lozzi, B.; Huerta, N.A.B.; Lin, C.-C.J.; Felice, D.; Jain, A.; Paulucci-Holthauzen, A.; Deneen, B. Region-Specific transcriptional control of astrocyte function oversees local circuit activities. Neuron 2020, 106, 992–1008.e9. [Google Scholar] [CrossRef] [PubMed]
- Lozzi, B.; Huang, T.-W.; Sardar, D.; Huang, A.Y.-S.; Deneen, B. Regionally distinct astrocytes display unique transcription factor profiles in the adult brain. Front. Neurosci. 2020, 14, 61. [Google Scholar] [CrossRef]
- Morel, L.; Chiang, M.S.R.; Higashimori, H.; Shoneye, T.; Iyer, L.K.; Yelick, J.; Tai, A.; Yang, Y. Molecular and functional properties of regional astrocytes in the adult brain. J. Neurosci. 2017, 37, 8706–8717. [Google Scholar] [CrossRef]
- Saunders, A.; Macosko, E.Z.; Wysoker, A.; Goldman, M.; Krienen, F.M.; Rivera, H.; Bien, E.; Baum, M.; Bortolin, L.; Wang, S.; et al. Molecular diversity and specializations among the cells of the adult mouse brain. Cell 2018, 174, 1015–1030.e16. [Google Scholar] [CrossRef] [PubMed]
- Pannasch, U.; Rouach, N. Emerging role for astroglial networks in information processing: From synapse to behavior. Trends Neurosci. 2013, 36, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Aponte, Y.; Atasoy, D.; Sternson, S. AGRP neurons are sufficient to orchestrate feeding behavior rapidly and without training. Nat. Neurosci. 2011, 14, 351–355. [Google Scholar] [CrossRef]
- Carter, M.E.; Soden, M.E.; Zweifel, L.S.; Palmiter, R.D. Genetic identification of a neural circuit that suppresses appetite. Nature 2013, 503, 111–114. [Google Scholar] [CrossRef]
- Mu, D.; Deng, J.; Liu, K.F.; Wu, Z.Y.; Shi, Y.F.; Guo, W.M.; Mao, Q.Q.; Liu, X.J.; Li, H.; Sun, Y.G. A central neural circuit for itch sensation. Science 2017, 357, 695–699. [Google Scholar] [CrossRef]
- Chiang, M.C.; Bowen, A.; Schier, L.A.; Tupone, D.; Uddin, O.; Heinricher, M.M. Parabrachial complex: A hub for pain and aversion. J. Neurosci. 2019, 39, 8225–8230. [Google Scholar] [CrossRef]
- Füzesi, T.; Daviu, N.; Wamsteeker Cusulin, J.I.; Bonin, R.P.; Bains, J.S. Hypothalamic CRH neurons orchestrate complex behaviours after stress. Nat. Commun. 2016, 7, 11937. [Google Scholar] [CrossRef] [PubMed]
- Jennings, J.H.; Rizzi, G.; Stamatakis, A.M.; Ung, R.L.; Stuber, G.D. The inhibitory circuit architecture of the lateral hypothalamus orchestrates feeding. Science 2013, 341, 1517–1521. [Google Scholar] [CrossRef]
- Mangieri, L.R.; Lu, Y.; Xu, Y.; Cassidy, R.M.; Xu, Y.; Arenkiel, B.R.; Tong, Q. A neural basis for antagonistic control of feeding and compulsive behaviors. Nat. Commun. 2018, 9, 52. [Google Scholar] [CrossRef]
- Wilson, T.D.; Valdivia, S.; Khan, A.; Ahn, H.-S.; Adke, A.P.; Gonzalez, S.M.; Sugimura, Y.K.; Carrasquillo, Y. Dual and opposing functions of the central amygdala in the modulation of pain. Cell Rep. 2019, 29, 332–346.e5. [Google Scholar] [CrossRef]
- Haubensak, W.; Kunwar, P.S.; Cai, H.; Ciocchi, S.; Wall, N.R.; Ponnusamy, R.; Biag, J.; Dong, H.-W.; Deisseroth, K.; Callaway, E.M.; et al. Genetic dissection of an amygdala microcircuit that gates conditioned fear. Nature 2010, 468, 270–276. [Google Scholar] [CrossRef]
- Hong, W.; Kim, D.W.; Anderson, D.J. Antagonistic control of social versus repetitive self-grooming behaviors by separable amygdala neuronal subsets. Cell 2014, 158, 1348–1361. [Google Scholar] [CrossRef]
- Araque, A.; Carmignoto, G.; Haydon, P.G.; Oliet, S.H.; Robitaille, R.; Volterra, A. Gliotransmitters travel in time and space. Neuron 2014, 81, 728–739. [Google Scholar] [CrossRef]
- De Pittà, M.; Brunel, N.; Volterra, A. Astrocytes: Orchestrating synaptic plasticity? Neuroscience 2016, 323, 43–61. [Google Scholar] [CrossRef]
- Min, R.; Nevian, T. Astrocyte signaling controls spike timing-dependent depression at neocortical synapses. Nat. Neurosci. 2012, 15, 746–753. [Google Scholar] [CrossRef]
- Savtchouk, I.; Carriero, G.; Volterra, A. Studying axon-astrocyte functional interactions by 3D two-photon Ca2+ imaging: A practical guide to experiments and “Big Data” analysis. Front. Cell. Neurosci. 2018, 12, 98. [Google Scholar] [CrossRef] [PubMed]
- Volterra, A.; Meldolesi, J. Astrocytes, from brain glue to communication elements: The revolution continues. Nat. Rev. Neurosci. 2005, 6, 626–640. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Bajo-Grañeras, R.; Moratalla, R.; Perea, G.; Araque, A. Circuit-Specific signaling in astrocyte-neuron networks in basal ganglia pathways. Science 2015, 349, 730–734. [Google Scholar] [CrossRef]
- Mor, D.; Bembrick, A.L.; Austin, P.J.; Wyllie, P.M.; Creber, N.J.; Denyer, G.S.; Keay, K.A. Anatomically specific patterns of glial activation in the periaqueductal gray of the sub-population of rats showing pain and disability following chronic constriction injury of the sciatic nerve. Neuroscience 2010, 166, 1167–1184. [Google Scholar] [CrossRef] [PubMed]
- Thaler, J.P.; Yi, C.-X.; Schur, E.A.; Guyenet, S.J.; Hwang, B.H.; Dietrich, M.O.; Zhao, X.; Sarruf, D.A.; Izgur, V.; Maravilla, K.R.; et al. Obesity is associated with hypothalamic injury in rodents and humans. J. Clin. Investig. 2012, 122, 153–162. [Google Scholar] [CrossRef]
- Bazargani, N.; Attwell, D. Astrocyte calcium signaling: The third wave. Nat. Neurosci. 2016, 19, 182–189. [Google Scholar] [CrossRef]
- Shigetomi, E.; Patel, S.; Khakh, B.S. Probing the complexities of astrocyte calcium signaling. Trends Cell Biol. 2016, 26, 300–312. [Google Scholar] [CrossRef]
- Srinivasan, R.; Huang, B.S.; Venugopal, S.; Johnston, A.D.; Chai, H.; Zeng, H.; Golshani, P.; Khakh, B.S. Ca(2+) signaling in astrocytes from Ip3r2(−/−) mice in brain slices and during startle responses in vivo. Nat. Neurosci. 2015, 18, 708–717. [Google Scholar] [CrossRef]
- Uchiyama, T.; Yoshikawa, F.; Hishida, A.; Furuichi, T.; Mikoshiba, K. A novel recombinant hyperaffinity inositol 1,4,5-trisphosphate (IP3) absorbent traps IP3, resulting in specific inhibition of IP3-mediated Calcium Signaling. J. Biol. Chem. 2002, 277, 8106–8113. [Google Scholar] [CrossRef] [PubMed]
- Rungta, R.L.; Bernier, L.-P.; Dissing-Olesen, L.; Groten, C.J.; LeDue, J.M.; Ko, R.; Drissler, S.; MacVicar, B.A. Ca2+ transients in astrocyte fine processes occur via Ca2+ influx in the adult mouse hippocampus. Glia 2016, 64, 2093–2103. [Google Scholar] [CrossRef] [PubMed]
- Stobart, J.L.; Ferrari, K.D.; Barret, M.J.P.; Stobart, M.J.; Looser, Z.J.; Saab, A.S.; Weber, B. Long-Term in vivo calcium imaging of astrocytes reveals distinct cellular compartment responses to sensory stimulation. Cereb. Cortex 2018, 28, 184–198. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, T.; Sun, G.Y.; Ding, S. Specific disruption of astrocytic Ca2+ signaling pathway in vivo by adeno-associated viral transduction. Neuroscience 2010, 170, 992–1003. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Shin, P.-Y.; Gomi, H.; Yoshida, T.; Nakai, J.; Ando, R.; Furuichi, T.; Mikoshiba, K.; Semyanov, A.; Itohara, S. Astrocytic Ca2+ signals are required for the functional integrity of tripartite synapses. Mol. Brain 2013, 6, 6. [Google Scholar] [CrossRef]
- Bang, J.; Kim, H.Y.; Lee, H. Optogenetic and chemogenetic approaches for studying astrocytes and gliotransmitters. Exp. Neurobiol. 2016, 25, 205–221. [Google Scholar] [CrossRef]
- Nagai, J.; Yu, X.; Papouin, T.; Cheong, E.; Freeman, M.R.; Monk, K.R.; Hastings, M.H.; Haydon, P.G.; Rowitch, D.; Shaham, S.; et al. Behaviorally consequential astrocytic regulation of neural circuits. Neuron 2020. [Google Scholar] [CrossRef]
- Armbruster, B.N.; Li, X.; Pausch, M.H.; Herlitze, S.; Roth, B.L. Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc. Natl. Acad. Sci. USA 2007, 104, 5163–5168. [Google Scholar] [CrossRef]
- Octeau, J.C.; Gangwani, M.R.; Allam, S.L.; Tran, D.; Huang, S.; Hoang-Trong, T.M.; Golshani, P.; Rumbell, T.H.; Kozloski, J.R.; Khakh, B.S. Transient, consequential increases in extracellular potassium ions accompany channelrhodopsin2 excitation. Cell Rep. 2019, 27, 2249–2261.e7. [Google Scholar] [CrossRef] [PubMed]
- Woo, D.H.; Han, K.S.; Shim, J.W.; Yoon, B.E.; Kim, E.; Bae, J.Y.; Oh, S.J.; Hwang, E.M.; Marmorstein, A.D.; Bae, Y.C.; et al. TREK-1 and Best1 channels mediate fast and slow glutamate release in astrocytes upon GPCR activation. Cell 2012, 151, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Romanos, J.; Benke, D.; Pietrobon, D.; Zeilhofer, H.U.; Santello, M. Astrocyte dysfunction increases cortical dendritic excitability and promotes cranial pain in familial migraine. Sci. Adv. 2020, 6, eaaz1584. [Google Scholar] [CrossRef]
- Skucas, V.A.; Mathews, I.B.; Yang, J.; Cheng, Q.; Treister, A.; Duffy, A.M.; Verkman, A.S.; Hempstead, B.L.; Wood, M.A.; Binder, D.K.; et al. Impairment of select forms of spatial memory and neurotrophin-dependent synaptic plasticity by deletion of glial aquaporin-4. J. Neurosci. 2011, 31, 6392–6397. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, S.-N.; Lee, J.S.; Seo, K.; Lee, H. Astrocytic Regulation of Neural Circuits Underlying Behaviors. Cells 2021, 10, 296. https://doi.org/10.3390/cells10020296
Hwang S-N, Lee JS, Seo K, Lee H. Astrocytic Regulation of Neural Circuits Underlying Behaviors. Cells. 2021; 10(2):296. https://doi.org/10.3390/cells10020296
Chicago/Turabian StyleHwang, Sun-Nyoung, Jae Seung Lee, Kain Seo, and Hyosang Lee. 2021. "Astrocytic Regulation of Neural Circuits Underlying Behaviors" Cells 10, no. 2: 296. https://doi.org/10.3390/cells10020296
APA StyleHwang, S.-N., Lee, J. S., Seo, K., & Lee, H. (2021). Astrocytic Regulation of Neural Circuits Underlying Behaviors. Cells, 10(2), 296. https://doi.org/10.3390/cells10020296