Methods for Studying Endometrial Pathology and the Potential of Atomic Force Microscopy in the Research of Endometrium



Abstract
1. Introduction
2. Cell Morphology and Topography—First Source of Information
3. Cell Metabolism—Gene and Protein Expression
4. Molecular Spectroscopy and Nano-Spectroscopy
5. AFM in Studies of Physical, Chemical, and Mechanical Properties of Endometrial Cells
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lv, S.; Wang, R.; Wang, Q.; Han, L.; Tuo, X.; Hou, H.; Liu, Y.; Shi, Z.; Wang, Q.; Li, Y.; et al. A novel solution configuration on liquid-based endometrial cytology. PLoS ONE 2018, 13, 1–13. [Google Scholar] [CrossRef] [PubMed]
- McAlpine, J.N.; Temkin, S.M.; Mackay, H.J. Endometrial cancer: Not your grandmother’s cancer. Cancer 2016, 122, 2787–2798. [Google Scholar] [CrossRef] [PubMed]
- Morice, P.; Leary, A.; Creutzberg, C.; Abu-Rustum, N.; Darai, E. Endometrial cancer. Lancet 2016, 387, 1094–1108. [Google Scholar] [CrossRef]
- Samir, C.; Kurtek, S.; Srivastava, A.; Canis, M. Elastic shape analysis of cylindrical surfaces for 3D/2D registration in endometrial tissue characterization. IEEE Trans. Med. Imaging 2014, 33, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.Y.; Yang, X.M.; Cheng, M.J.; Zhang, R.; Ye, J.; Yi, H.; Ao, J.P.; Zhang, Z.G.; Xu, C.J. Dysregulated cell mechanical properties of endometrial stromal cells from endometriosis patients. Int. J. Clin. Exp. Pathol. 2014, 7, 656–662. [Google Scholar]
- Kim, S.-M.; Kim, J.-S. A Review of Mechanisms of Implantation. Dev. Reprod. 2017, 21, 351–359. [Google Scholar] [CrossRef]
- Bahar, L.; Kahraman, S.; Eras, N.; Pirkevi, C. Comparison of endometrial biopsies of fertile women and women with repeated implantation failure at the ultrastructural level. Turk. J. Med. Sci. 2015, 45, 706–713. [Google Scholar] [CrossRef]
- Zhu, H.; Hou, C.C.; Luo, L.F.; Hu, Y.J.; Yang, W.X. Endometrial stromal cells and decidualized stromal cells: Origins, transformation and functions. Gene 2014, 551, 1–14. [Google Scholar] [CrossRef]
- Thie, M.; Röspel, R.; Dettmann, W.; Benoit, M.; Ludwig, M.; Gaub, H.E.; Denker, H.W. Interactions between trophoblast and uterine epithelium: Monitoring of adhesive forces. Hum. Reprod. 1998, 13, 3211–3219. [Google Scholar] [CrossRef]
- Thie, M.; Denker, H.W. In vitro studies on endometrial adhesiveness for trophoblast: Cellular dynamics in uterine epithelial cells. Cells Tissues Organs 2002, 172, 237–252. [Google Scholar] [CrossRef]
- Amant, F.; Moerman, P.; Neven, P.; Timmerman, D.; Van Limbergen, E.; Vergote, I. Endometrial cancer. Lancet 2005, 366, 491–505. [Google Scholar] [CrossRef]
- Parlatan, U.; Inanc, M.T.; Ozgor, B.Y.; Oral, E.; Bastu, E.; Unlu, M.B.; Basar, G. Raman spectroscopy as a non-invasive diagnostic technique for endometriosis. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Gupta, N.; Dey, P. Morphometric analysis of endometrial cells in liquid-based cervical cytology samples. Cytopathology 2017, 28, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Kirk, D.; King, R.J.B.; Heyes, J.; Peachey, L.; Hirsch, P.J.; Taylor, R.W.T. Normal human endometrium in cell culture—I. Separation and characterization of epithelial and stromal components in vitro. In Vitro 1978, 14, 651–662. [Google Scholar] [CrossRef]
- Pan-Castillo, B.; Gazze, S.A.; Thomas, S.; Lucas, C.; Margarit, L.; Gonzalez, D.; Francis, L.W.; Conlan, R.S. Morphophysical dynamics of human endometrial cells during decidualization. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 2235–2245. [Google Scholar] [CrossRef]
- Francis, L.W.; Lewis, P.D.; Gonzalez, D.; Ryder, T.A.; Webb, G.; Joels, L.A.; White, J.O.; Wright, C.J.; Conlan, R.S. Progesterone induces nano-scale molecular modifications on endometrial epithelial cell surfaces. Biol. Cell 2009, 101, 481–493. [Google Scholar] [CrossRef]
- Gonzalez, D.; Thackeray, H.; Lewis, P.D.; Mantani, A.; Brook, N.; Ahuja, K.; Margara, R.; Joels, L.; White, J.O.; Conlan, R.S. Loss of WT1 expression in the endometrium of infertile PCOS patients: A hyperandrogenic effect? J. Clin. Endocrinol. Metab. 2012, 97, 957–966. [Google Scholar] [CrossRef]
- Harada, M.; Osuga, Y.; Hirota, Y.; Koga, K.; Morimoto, C.; Hirata, T.; Yoshino, O.; Tsutsumi, O.; Yano, T.; Taketani, Y. Mechanical stretch stimulates interleukin-8 production in endometrial stromal cells: Possible implications in endometrium-related events. J. Clin. Endocrinol. Metab. 2005, 90, 1144–1148. [Google Scholar] [CrossRef]
- Trapero, C.; Vidal, A.; Fernández-Montolí, M.E.; Coroleu, B.; Tresserra, F.; Barri, P.; De Aranda, I.G.; Sévigny, J.; Ponce, J.; Matias-Guiu, X.; et al. Impaired expression of ectonucleotidases in ectopic and eutopic endometrial tissue is in favor of ATP accumulation in the tissue microenvironment in endometriosis. Int. J. Mol. Sci. 2019, 20, 5532. [Google Scholar] [CrossRef]
- Aliagas, E.; Vidal, A.; Texidó, L.; Ponce, J.; Condom, E.; Martín-Satué, M. High expression of ecto-nucleotidases CD39 and CD73 in human endometrial tumors. Mediat. Inflamm. 2014, 2014. [Google Scholar] [CrossRef]
- Gargett, C.E.; Schwab, K.E.; Zillwood, R.M.; Nguyen, H.P.T.; Wu, D. Isolation and culture of epithelial progenitors and mesenchymal stem cells from human endometrium. Biol. Reprod. 2009, 80, 1136–1145. [Google Scholar] [CrossRef] [PubMed]
- Notarstefano, V.; Gioacchini, G.; Byrne, H.J.; Zacà, C.; Sereni, E.; Vaccari, L.; Borini, A.; Carnevali, O.; Giorgini, E. Vibrational characterization of granulosa cells from patients affected by unilateral ovarian endometriosis: New insights from infrared and Raman microspectroscopy. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2019, 212, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Grady, M.E.; Composto, R.J.; Eckmann, D.M. Cell elasticity with altered cytoskeletal architectures across multiple cell types. J. Mech. Behav. Biomed. Mater. 2016, 61, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Hogan, B.; Babataheri, A.; Hwang, Y.; Barakat, A.I.; Husson, J. Characterizing Cell Adhesion by Using Micropipette Aspiration. Biophys. J. 2015, 109, 209–219. [Google Scholar] [CrossRef]
- Jing, P.; Liu, Y.; Keeler, E.G.; Cruz, N.M.; Freedman, B.S.; Lin, L.Y. Optical tweezers system for live stem cell organization at the single-cell level. Biomed. Opt. Express 2018, 9, 771. [Google Scholar] [CrossRef]
- Yousafzai, M.S.; Coceano, G.; Mariutti, A.; Ndoye, F.; Amin, L.; Niemela, J.; Bonin, S.; Scoles, G.; Cojoc, D. Effect of neighboring cells on cell stiffness measured by optical tweezers indentation. J. Biomed. Opt. 2016, 21, 057004. [Google Scholar] [CrossRef]
- Hadjiantoniou, S.; Guolla, L.; Pelling, A.E. Mechanically induced deformation and strain dynamics in actin stress fibers. Commun. Integr. Biol. 2012, 5, 627–630. [Google Scholar] [CrossRef]
- Lopez, J.I.; Kang, I.; You, W.K.; McDonald, D.M.; Weaver, V.M. In situ force mapping of mammary gland transformation. Integr. Biol. 2011, 3, 910–921. [Google Scholar] [CrossRef]
- Arce, F.T.; Whitlock, J.L.; Birukova, A.A.; Birukov, K.G.; Arnsdorf, M.F.; Lal, R.; Garcia, J.G.N.; Dudek, S.M. Regulation of the micromechanical properties of pulmonary endothelium by S1P and thrombin: Role of cortactin. Biophys. J. 2008, 95, 886–894. [Google Scholar] [CrossRef]
- Wang, X.; Bleher, R.; Brown, M.E.; Garcia, J.G.N.; Dudek, S.M.; Shekhawat, G.S.; Dravid, V.P. Nano-Biomechanical Study of Spatio-Temporal Cytoskeleton Rearrangements that Determine Subcellular Mechanical Properties and Endothelial Permeability. Sci. Rep. 2015, 5, 1–11. [Google Scholar] [CrossRef]
- Wang, X.; Bleher, R.; Wang, L.; Garcia, J.G.N.; Dudek, S.M.; Shekhawat, G.S.; Dravid, V.P. Imatinib Alters Agonists-mediated Cytoskeletal Biomechanics in Lung Endothelium. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Caluori, G.; Pribyl, J.; Pesl, M.; Oliver-De La Cruz, J.; Nardone, G.; Skladal, P.; Forte, G. Advanced and rationalized atomic force microscopy analysis unveils specific properties of controlled cell mechanics. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Xia, Y.; Sandig, M.; Yang, J. Characterization of cell elasticity correlated with cell morphology by atomic force microscope. J. Biomech. 2012, 45, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Ketene, A.N.; Schmelz, E.M.; Roberts, P.C.; Agah, M. The effects of cancer progression on the viscoelasticity of ovarian cell cytoskeleton structures. Nanomed. Nanotechnol. Biol. Med. 2012, 8, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Neugebauer, U.; Rösch, P.; Schmitt, M.; Popp, J.; Julien, C.; Rasmussen, A.; Budich, C.; Deckert, V. On the way to nanometer-sized information of the bacterial surface by tip-enhanced Raman spectroscopy. ChemPhysChem 2006, 7, 1428–1430. [Google Scholar] [CrossRef]
- Olesen, F.; Rose, P.; Nafees, S.; Van Rijswijk, E.; Hiom, S.; Muth, C.; Beyer, M.; Neal, R.D. The Aarhus statement: Improving design and reporting of studies on early cancer diagnosis Clinical Studies. Br. J. Cancer 2012, 106, 1262–1267. [Google Scholar] [CrossRef]
- Costas, L.; Frias-Gomez, J.; Guardiola, M.; Benavente, Y.; Pineda, M.; Pavón, M.; Martínez, J.M.; Climent, M.; Barahona, M.; Canet, J.; et al. New perspectives on screening and early detection of endometrial cancer. Int. J. Cancer 2019, 145, 3194–3206. [Google Scholar] [CrossRef]
- Heitzer, E.; Haque, I.S.; Roberts, C.E.S.; Speicher, M.R. Current and future perspectives of liquid biopsies in genomics-driven oncology. Nat. Rev. Genet. 2019, 20, 71–88. [Google Scholar] [CrossRef]
- Pucci, C.; Martinelli, C.; Ciofani, G. Innovative approaches for cancer treatment: Current perspectives and new challenges. Ecancermedicalscience 2019, 13, 1–26. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Massou, S.; Nunes Vicente, F.; Wetzel, F.; Mehidi, A.; Strehle, D.; Leduc, C.; Voituriez, R.; Rossier, O.; Nassoy, P.; Giannone, G. Cell stretching is amplified by active actin remodelling to deform and recruit proteins in mechanosensitive structures. Nat. Cell Biol. 2020, 22, 1011–1023. [Google Scholar] [CrossRef] [PubMed]
- Oda, T.; Takeda, S.; Narita, A.; Maéda, Y. Structural Polymorphism of Actin. J. Mol. Biol. 2019, 431, 3217–3228. [Google Scholar] [CrossRef] [PubMed]
- Kim, L.Y.; Thompson, P.M.; Lee, H.T.; Pershad, M.; Campbell, S.L.; Alushin, G.M. The Structural Basis of Actin Organization by Vinculin and Metavinculin. J. Mol. Biol. 2016, 428, 10–25. [Google Scholar] [CrossRef] [PubMed]
- Klein, D.E.; Stayrook, S.E.; Shi, F.; Narayan, K.; Lemmon, M.A. Structural basis for EGFR ligand sequestration by Argos. Nature 2008, 453, 1271–1275. [Google Scholar] [CrossRef] [PubMed]
- Large, M.J.; Wetendorf, M.; Lanz, R.B.; Hartig, S.M.; Creighton, C.J.; Mancini, M.A.; Kovanci, E.; Lee, K.F.; Threadgill, D.W.; Lydon, J.P.; et al. The Epidermal Growth Factor Receptor Critically Regulates Endometrial Function during Early Pregnancy. PLoS Genet. 2014, 10, 1–18. [Google Scholar] [CrossRef]
- Long, D.A. The Raman Effect: A Unified Treatment of the Theory of Raman Scattering by Molecules; John Wiley & Sons, Ltd.: Chichester, UK, 2002; Volume 8, ISBN 0471490288. [Google Scholar]
- Wilkosz, N.; Czaja, M.; Seweryn, S.; Skirlinska-Nosek, K.; Szymonski, M.; Lipiec, E.; Sofinska, K. Molecular spectroscopic markers of abnormal protein aggregation. Molecules 2020, 25, 2498. [Google Scholar] [CrossRef]
- Butler, H.J.; Ashton, L.; Bird, B.; Cinque, G.; Curtis, K.; Dorney, J.; Esmonde-White, K.; Fullwood, N.J.; Gardner, B.; Martin-Hirsch, P.L.; et al. Using Raman spectroscopy to characterize biological materials. Nat. Protoc. 2016, 11, 664–687. [Google Scholar] [CrossRef]
- Stöckle, R.M.; Suh, Y.D.; Deckert, V.; Zenobi, R. Nanoscale chemical analysis by tip-enhanced Raman spectroscopy. Chem. Phys. Lett. 2000, 318, 131–136. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, Y.; Dong, Z.C.; Jiang, S.; Zhang, C.; Chen, L.G.; Zhang, L.; Liao, Y.; Aizpurua, J.; Luo, Y.; et al. Chemical mapping of a single molecule by plasmon-enhanced Raman scattering. Nature 2013, 498, 82–86. [Google Scholar] [CrossRef]
- Lipiec, E.W.; Wood, B.R. Tip-Enhanced Raman Scattering: Principles, Instrumentation, and the Application toe Biological Systems. In Encyclopedia of Analytical Chemistry; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 1–26. [Google Scholar]
- Böhme, R.; Cialla, D.; Richter, M.; Rösch, P.; Popp, J.; Deckert, V. Biochemical imaging below the diffraction limit—Probing cellular membrane related structures by tip-enhanced Raman spectroscopy (TERS). J. Biophotonics 2010, 3, 455–461. [Google Scholar] [CrossRef]
- Francis, L.W.; Lewis, P.D.; Wright, C.J.; Conlan, R.S. Atomic force microscopy comes of age. Biol. Cell 2010, 102, 133–143. [Google Scholar] [CrossRef]
- Allison, D.P.; Mortensen, N.P.; Sullivan, C.J.; Doktycz, M.J. Atomic force microscopy of biological samples. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2010, 2, 618–634. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.C.; Chiang, Y.W.; Yang, C.H.; Liou, J.W. Atomic force microscopy in biology and biomedicine. Tzu Chi Med. J. 2012, 24, 162–169. [Google Scholar] [CrossRef]
- Lal, R.; John, S.A. Biological applications of atomic force microscopy. Am. J. Physiol. Cell Physiol. 1994, 266, 1298–1304. [Google Scholar] [CrossRef]
- Kuznetsova, T.G.; Starodubtseva, M.N.; Yegorenkov, N.I.; Chizhik, S.A.; Zhdanov, R.I. Atomic force microscopy probing of cell elasticity. Micron 2007, 38, 824–833. [Google Scholar] [CrossRef]
- Rutherford, E.; Hill, A.; Hopkins, A. Adhesion in Physiological, Benign and Malignant Proliferative States of the Endometrium: Microenvironment and the Clinical Big Picture. Cells 2018, 7, 43. [Google Scholar] [CrossRef]
- Kashef, J.; Franz, C.M. Quantitative methods for analyzing cell-cell adhesion in development. Dev. Biol. 2015, 401, 165–174. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, K.K. Optical tweezers for single cells. J. R. Soc. Interface 2008, 5, 671–690. [Google Scholar] [CrossRef]
- Neuman, K.C.K.K.C.; Nagy, A. Single-molecule force spectroscopy: Optical tweezers, magnetic tweezers and atomic force microscopy. Nat. Methods 2008, 5, 491–505. [Google Scholar] [CrossRef]
- Fazal, F.M.; Block, S.M. Optical tweezers study life under tension. Nat. Photonics 2011, 5, 318–321. [Google Scholar] [CrossRef]
- Luo, Q.; Kuang, D.; Zhang, B.; Song, G. Biochimica et Biophysica Acta Cell stiffness determined by atomic force microscopy and its correlation with cell motility. BBA Gen. Subj. 2016, 1860, 1953–1960. [Google Scholar] [CrossRef] [PubMed]
- Haase, K.; Pelling, A.E. Investigating cell mechanics with atomic force microscopy. J. R. Soc. Interface 2015, 12. [Google Scholar] [CrossRef] [PubMed]
- Lekka, M. Discrimination Between Normal and Cancerous Cells Using AFM. Bionanoscience 2016, 6, 65–80. [Google Scholar] [CrossRef]
- Rebelo, L.M.; De Sousa, J.S.; Mendes Filho, J.; Radmacher, M. Comparison of the viscoelastic properties of cells from different kidney cancer phenotypes measured with atomic force microscopy. Nanotechnology 2013, 24, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhong, Y.; Ye, T.; Wang, D.; Mao, B. Discrimination Between Cervical Cancer Cells and Normal Cervical Cells Based on Longitudinal Elasticity Using Atomic Force Microscopy. Nanoscale Res. Lett. 2015, 10, 1–8. [Google Scholar] [CrossRef]
- Huang, L.; Ma, M.; Du, Z.; Liu, Z.; Gong, X. Quantitative evaluation of tissue stiffness around lesion by sound touch elastography in the diagnosis of benign and malignant breast lesions. PLoS ONE 2019, 14, 1–14. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
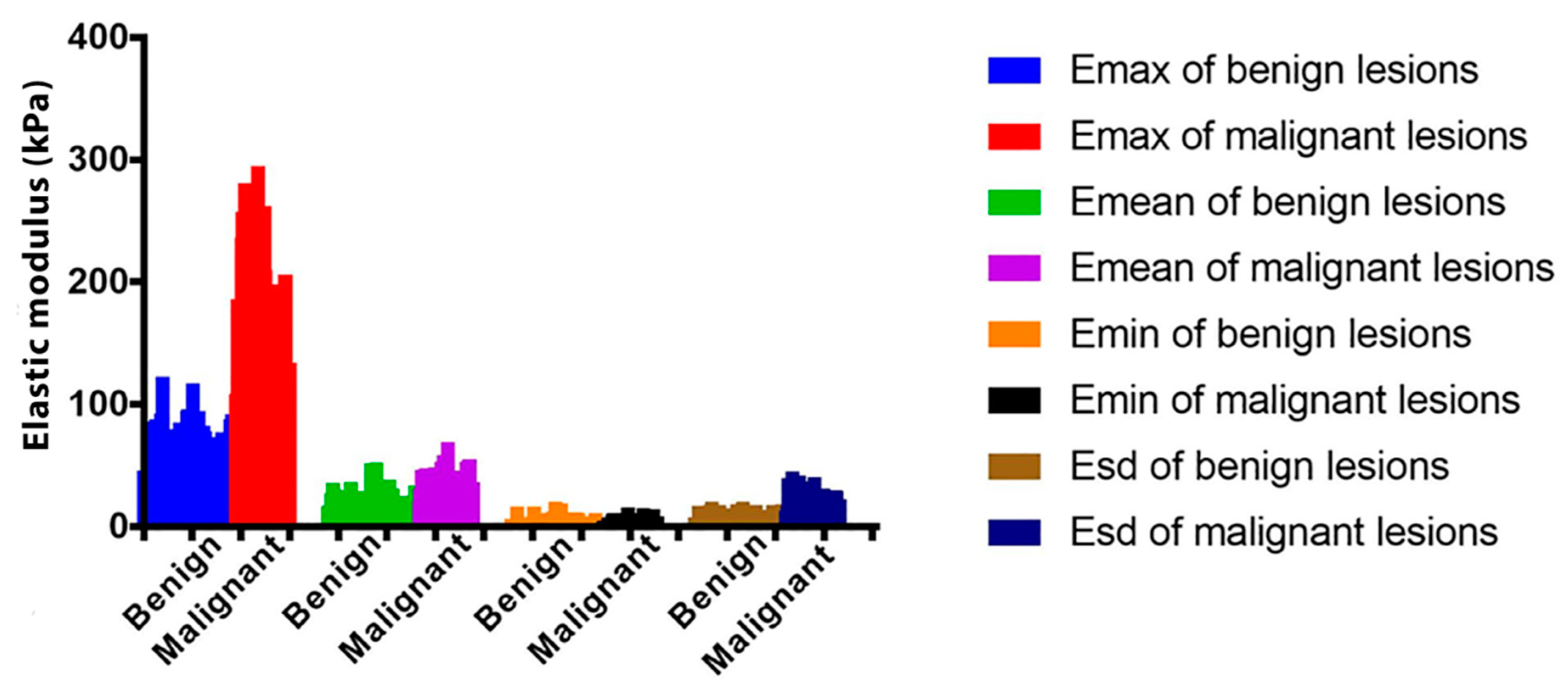{kind=link}
| Technique | Example of Application | |
|---|---|---|
| Microscopic techniques | Optical microscopy | Classification of benign, atypical and malignant cells [13] |
| Transmission and scanning electron microscopy | Analysis of structure differences between epithelial and stromal cells [14] | |
| Transmission electron microscopy | Observation of ultrastructural differences between endometrial cell from fertile woman and from woman with repeated implantation failure [7] | |
| Confocal microscopy | Molecules tracking as source of information on the course of cellular processes e.g., Observation of actin expression changes during endometriosis [5] Tracking of mesenchymal to epithelial transition markers [15] Analysis of phase of menstrual cycle [16] | |
| Atomic force microscopy | Height and roughness measurement, e.g., changes after progesterone treatment [16] or during decidualization [15] | |
| Time-lapse video-microscopy | Change of migration ability resulting from endometriosis [5] | |
| Gene and protein expression analysis (both complete research and research support) | Reverse-transcription polymerase chain reaction and Western blot | Confirmation of the ongoing cellular processes based on the expression of gen or protein [5,16,17] Monitoring of stretch-induced protein secretion in the context of uterine contractility [18] |
| Real-time polymerase chain reaction | Observation of gene expression changes during subsequent cycles e.g., in decidualization [15] | |
| Enzymatic tests | Enzyme-linked immunosorbent assay | Analysis of surface markers e.g., in decidualization [15] Monitoring of stretch-induced protein secretion in the context of uterine contractility [18] |
| Histochemistry | In situ activity analysis of ectonucleotidases as markers of endometriosis and endometrial cancer [19,20] | |
| Analytical technique | Flow cytometry | Analysis of surface markers, assessment of cell quality, e.g., in cell phenotype determination [21] |
| Vibration technique | Raman spectroscopy | Chemical composition of sample, e.g., in endometriosis diagnosis based on Raman spectrum [12] |
| Infrared micro-spectroscopy | Chemical composition of sample, e.g., changes caused by endometriosis [22] | |
| Mechanical properties measurement | Atomic force microscopy | Determination of Young’s modulus and adhesion of decidualized cells [15] Determination of adhesion in implantation process [9] |
| Technique | Example of Application | |
|---|---|---|
| Microscopic techniques | Phase contrast microscopy | Observation of actin filament and microtubules impact on human umbilical vein endothelial cells, chondrocytes, fibroblasts, fibrosarcoma and hepatocellular carcinoma cells [23] |
| Mechanical properties measurement | Micropipette aspiration | Determination of bovine aortic endothelial cells adhesion [24] |
| Optical tweezers | Intercellular adhesion of the early embryo epithelial cells determined by cell displacement [25] Assessment of the influence of neighboring cells on the stiffness of breast cancer cells [26] | |
| Atomic force microscopy | Detecting fibroblast cell inhomogeneities based on point cell pressing [27] Force mapping: Young’s modulus maps e.g., of mammary gland cancer [28], primary human pulmonary artery endothelial cells [29,30,31], epithelial-like breast carcinoma cells [32] and elasticity maps e.g., of human aortic endothelial cells [33] Elasticity measurements of human umbilical vein endothelial cells, chondrocytes, fibroblasts, fibrosarcoma and hepatocellular carcinoma cells [23] Differentiation of early, intermediate and late cancer stage base on elasticity and viscosity measurement [34] | |
| Combination with atomic force microscopy | Tip-enhanced Raman spectroscopy | Chemical composition of sample with higher resolution, e.g., peptides and polysaccharides on bacteria surface [35] |
| Elastic Modulus E (Pa) of Luminal Breast Cancer Cells MCF-7 | Elastic Modulus E (Pa) of Normal and Myoepithelial Cells HBL-100 | |||
|---|---|---|---|---|
| Isolation | Contact | Isolation | Contact | |
| Region above nucleus (L1) | 39 ± 8 | 20 ± 11 | 36 ± 11 | 30 ± 11 |
| Cytoplasm in intermediate position (L2) | 21 ± 10 | 18 ± 11 | 27 ± 10 | 25 ± 9 |
| Region near the leading edge (L3) | 16 ± 6 | 14 ± 7 | 19 ± 8 | 21 ± 8 |
| Measurements with JAr Covered Microbeads | HEC-1-A Cells | RL95-2 Cells |
|---|---|---|
| Distance range for soft repulsion observation | 4.0 ± 0.3 μm | 3.4 ± 0.4 μm |
| Indentation forces during the start of hard repulsion | 1.0 ± 0.2 nN | 0.7 ± 0.2 nN |
| Adhesive maximum after one minute contact | 7.1 ± 2 nN | 4.2 ± 2 nN |
| Adhesive maximum after one minute contact | 16 ± 4 nN | - |
| Early Stage of Cancer | Intermediate Stage of Cancer | Late Stage of Cancer | |
|---|---|---|---|
| Elastic modulus peak maximum (kPa) | 0.652 | 0.477 | 0.382 |
| Average elastic modulus (kPa) | 1.097 ± 0.682 | 0.796 ± 0.441 | 0.549 ± 0.281 |
| Viscosity rate peak maximum (Pa∙s) | 69.69 | 60.40 | 25.53 |
| Average Young modulus (kPa) | 0.554 ± 0.349 | 0.472 ± 0.306 | 0.395 ± 0.136 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurek, A.; Kłosowicz, E.; Sofińska, K.; Jach, R.; Barbasz, J. Methods for Studying Endometrial Pathology and the Potential of Atomic Force Microscopy in the Research of Endometrium. Cells 2021, 10, 219. https://doi.org/10.3390/cells10020219
Kurek A, Kłosowicz E, Sofińska K, Jach R, Barbasz J. Methods for Studying Endometrial Pathology and the Potential of Atomic Force Microscopy in the Research of Endometrium. Cells. 2021; 10(2):219. https://doi.org/10.3390/cells10020219
Chicago/Turabian StyleKurek, Agnieszka, Estera Kłosowicz, Kamila Sofińska, Robert Jach, and Jakub Barbasz. 2021. "Methods for Studying Endometrial Pathology and the Potential of Atomic Force Microscopy in the Research of Endometrium" Cells 10, no. 2: 219. https://doi.org/10.3390/cells10020219
APA StyleKurek, A., Kłosowicz, E., Sofińska, K., Jach, R., & Barbasz, J. (2021). Methods for Studying Endometrial Pathology and the Potential of Atomic Force Microscopy in the Research of Endometrium. Cells, 10(2), 219. https://doi.org/10.3390/cells10020219