
Pregnancy-Associated Plasma Protein (PAPP)-A2 in Physiology and Disease

 , , , , , , , ,
, , , , , , , ,  and
and
Abstract
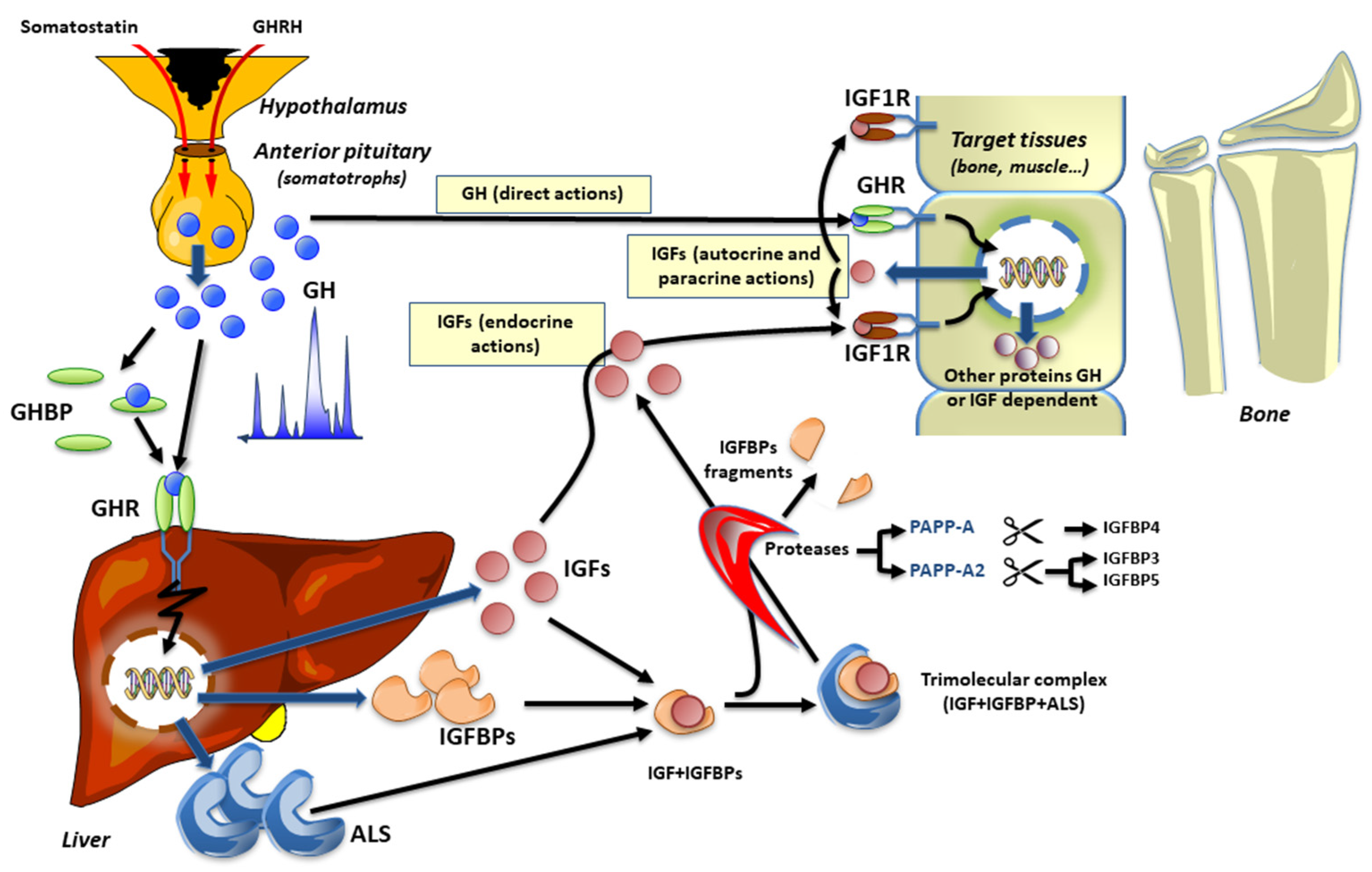
1. Introduction
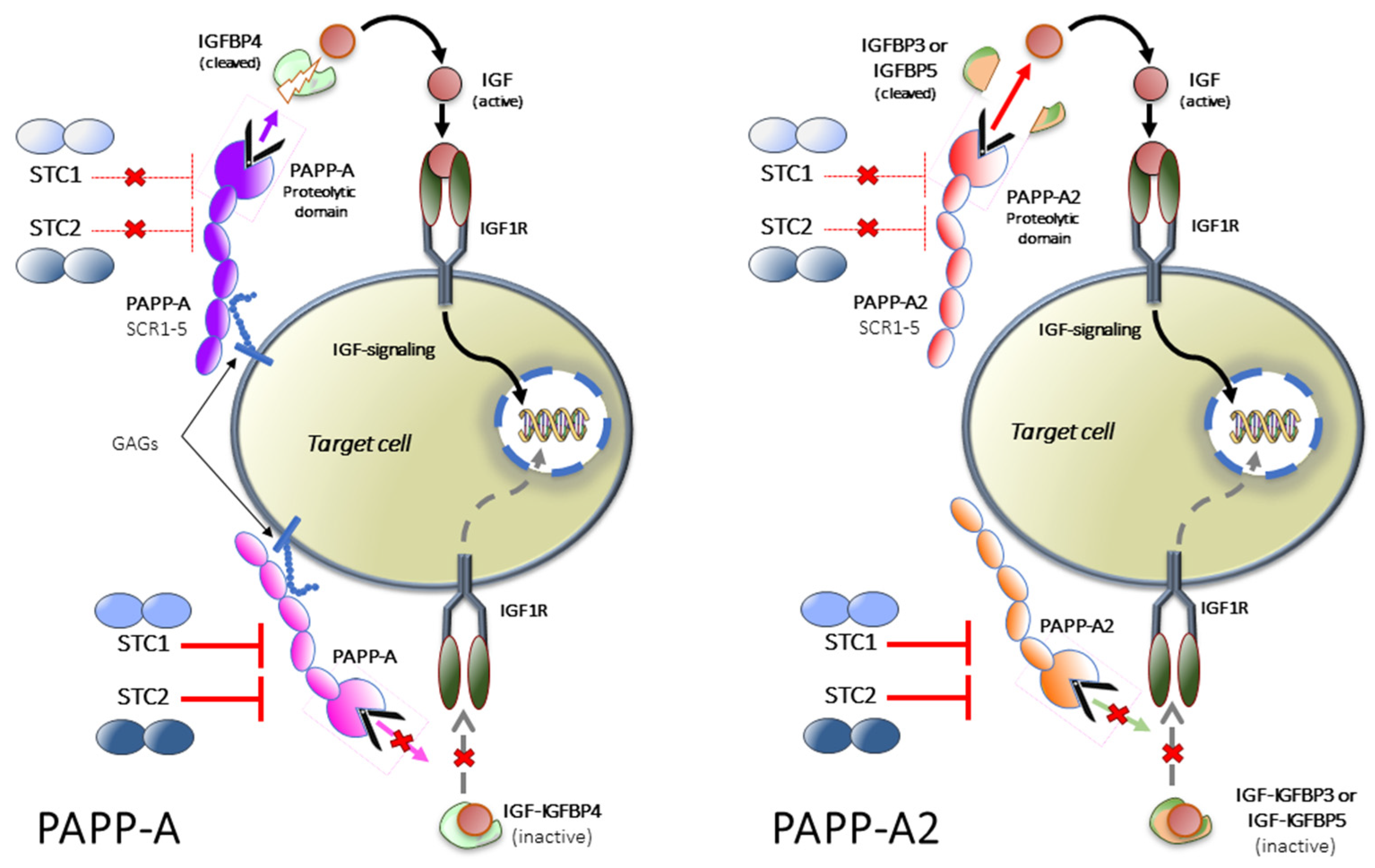
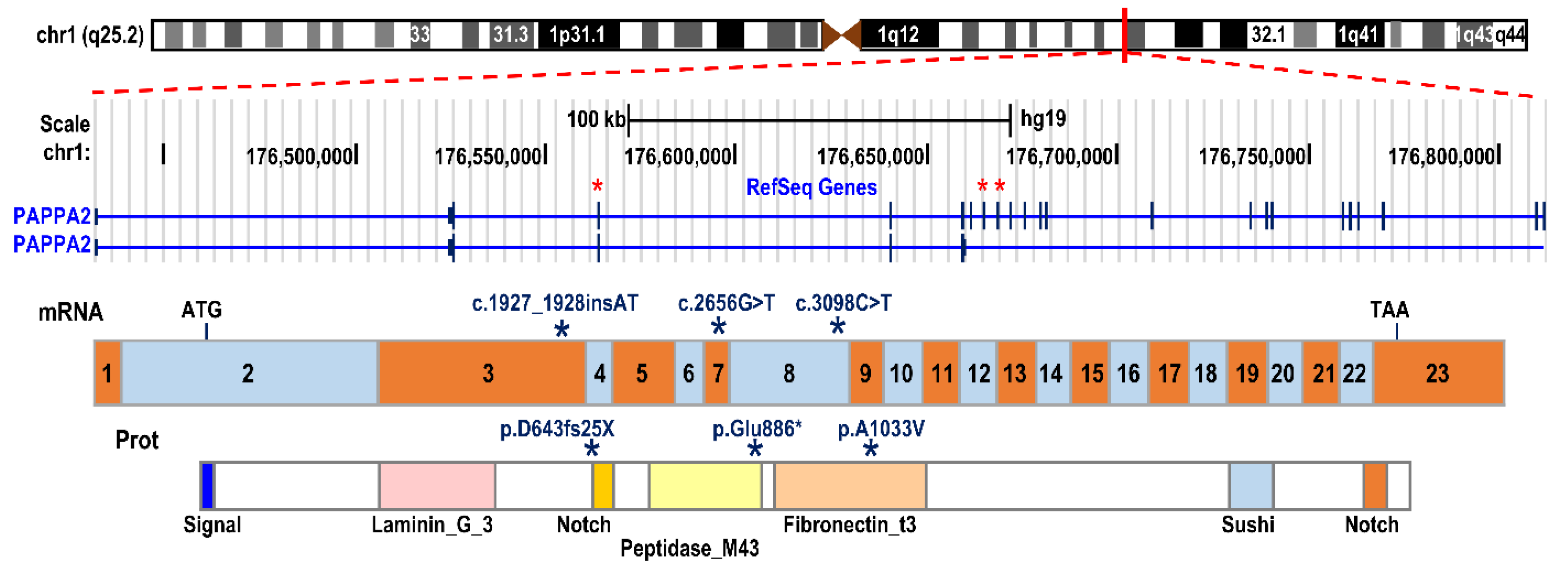
2. Mutations in the Human PAPP-A2 Gene
2.1. Clinical Phenotype
2.2. Auxology in PAPP-A2 Deficient Patients
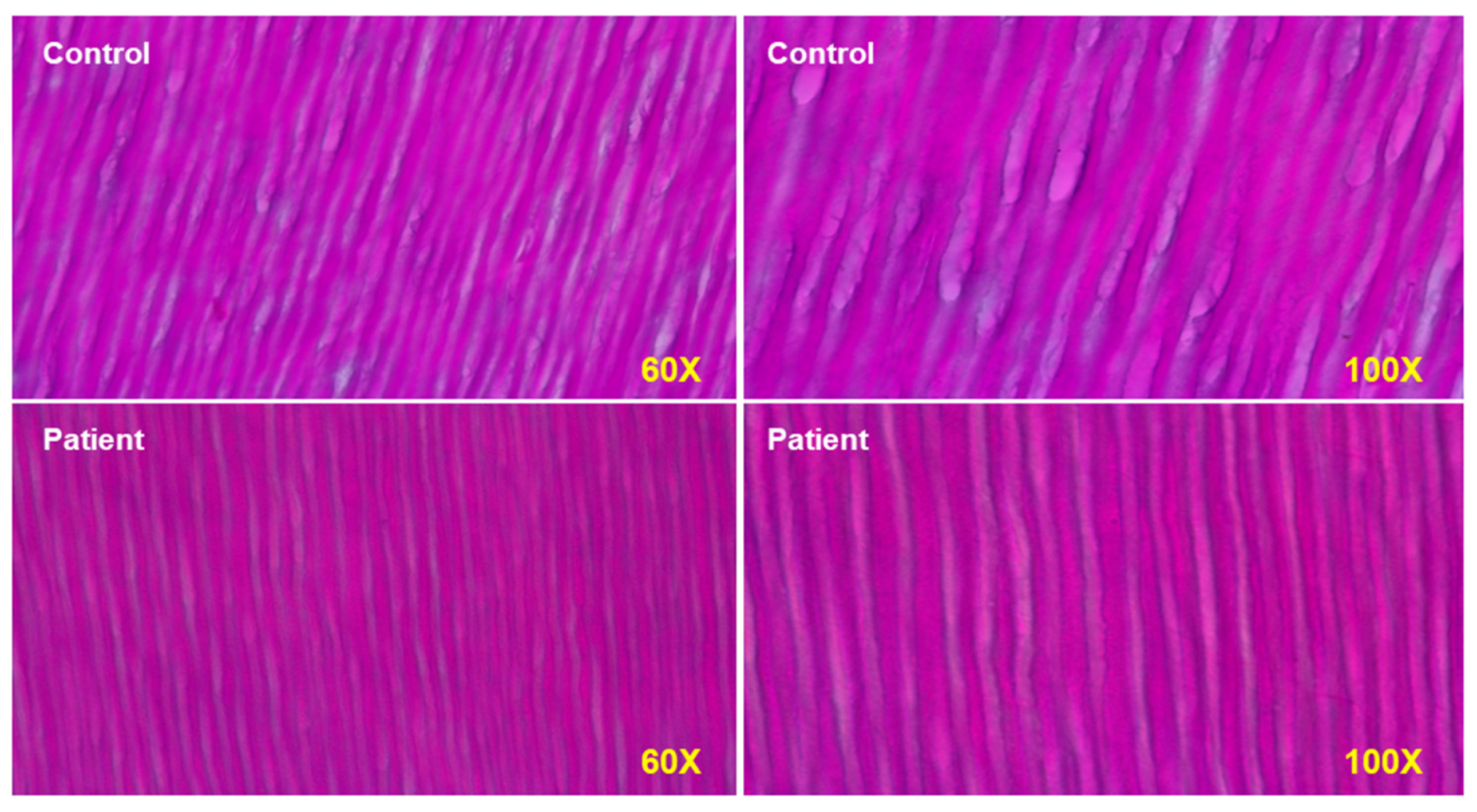
2.3. Bone and Teeth Abnormalities
2.4. Glucose Metabolism
2.5. Treatment
3. PAPP-A2 Mouse Models: What Are We Learning?
3.1. Expression Pattern of Pappa2 in Mice
3.2. Available Models and Their Differences in Growth and Body Weight
3.3. Differences in Hormonal Alterations in Available Models of Pappa2 Deficiency
3.4. Metabolic Disturbance in Available Models of Pappa2 Deficiency
3.5. Variations in Allometric and Bone Mineral Alterations in Mouse Models of Pappa2 Deficiency
3.6. Effects of Pappa2 Deficiency on Mechanisms Regulating Bone Development and Remodeling
3.7. Experimental Treatments in Mouse Models of Pappa2 Deficiency
4. Future Directions
4.1. Future Directions in Animal Models
4.2. Future Directions in Humans
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Lui, J.C.; Nilsson, O.; Baron, J. Recent research on the growth plate: Recent insights into the regulation of the growth plate. J. Mol. Endocrinol. 2014, 53, T1–T9. [Google Scholar] [CrossRef]
- Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H.; Kangawa, K. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 1999, 402, 656–660. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Asnicar, M.; Smith, R.G. Central and Peripheral Roles of Ghrelin on Glucose Homeostasis. Neuroendocrinology 2007, 86, 215–228. [Google Scholar] [CrossRef]
- Chanoine, J.-P.; De Waele, K.; Walia, P. Ghrelin and the growth hormone secretagogue receptor in growth and development. Int. J. Obes. 2009, 33 (Suppl. 1), S48–S52. [Google Scholar] [CrossRef] [PubMed]
- Coxam, V.; Bauchart, D.; Durand, D.; Davicco, M.-J.; Opmeer, F.; Barlet, J.-P. Nutrient effects on the hepatic production of somatomedin C (IGF1) in the milk-fed calf. Br. J. Nutr. 1989, 62, 425–437. [Google Scholar] [CrossRef]
- Brameld, J.; Gilmour, R.S.; Buttery, P.J. Glucose and Amino Acids Interact with Hormones to Control Expression of Insulin-Like Growth Factor-I and Growth Hormone Receptor mRNA in Cultured Pig Hepatocytes. J. Nutr. 1999, 129, 1298–1306. [Google Scholar] [CrossRef]
- Bailes, J.; Soloviev, M. Insulin-Like Growth Factor-1 (IGF-1) and Its Monitoring in Medical Diagnostic and in Sports. Biomolecules 2021, 11, 217. [Google Scholar] [CrossRef] [PubMed]
- Salmon, W.D., Jr.; Daughaday, W.H. A hormonally controlled serum factor which stimulates sulfate incorporation by cartilage in vitro. J. Lab. Clin. Med. 1957, 49, 825–836. [Google Scholar]
- Hintz, R.L.; Liu, F. Demonstration of specific plasma protein binding sites for somatomedin. J. Clin. Endocrinol. Metab. 1977, 45, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Shen, X.; Wan, C.; Zhao, Q.; Zhang, L.; Zhou, Q.; Deng, L. Effects of insulin and insulin-like growth factor 1 on osteoblast proliferation and differentiation: Differential signalling via Akt and ERK. Cell Biochem. Funct. 2012, 30, 297–302. [Google Scholar] [CrossRef]
- McKinnon, T.; Chakraborty, C.; Gleeson, L.M.; Chidiac, P.; Lala, P.K. Stimulation of human extravillous trophoblast migration by IGF-II is mediated by IGF type 2 receptor involving inhibitory G protein(s) and phosphorylation of MAPK. J. Clin. Endocrinol. Metab. 2001, 86, 3665–3674. [Google Scholar] [CrossRef]
- Guler, H.-P.; Zapf, J.; Schmid, C.; Froesch, E.R. Insulin-like growth factors I and II in healthy man. Estimations of half-lives and production rates. Acta Endocrinol. 1989, 121, 753–758. [Google Scholar] [CrossRef]
- Yakar, S.; Werner, H.; Rosen, C.J. Insulin-like growth factors: Actions on the skeleton. J. Mol. Endocrinol. 2018, 61, T115–T137. [Google Scholar] [CrossRef]
- Bach, L.A. IGF-binding proteins. J. Mol. Endocrinol. 2018, 61, T11–T28. [Google Scholar] [CrossRef]
- Firth, S.M.; Ganeshprasad, U.; Baxter, R. Structural determinants of ligand and cell surface binding of insulin-like growth factor-binding protein-3. J. Biol. Chem. 1998, 273, 2631–2638. [Google Scholar] [CrossRef]
- Allard, J.B.; Duan, C. IGF-Binding Proteins: Why Do They Exist and Why Are There So Many? Front. Endocrinol. 2018, 9, 117. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Gluckman, P.; Yang, P.; Krissansen, G.; Sun, X.; Zhou, Y.; Wen, J.; Phillips, G.; Shorten, P.R.; McMahon, C.D.; et al. Cyclic glycine-proline regulates IGF-1 homeostasis by altering the binding of IGFBP-3 to IGF-1. Sci. Rep. 2014, 4, 4388. [Google Scholar] [CrossRef]
- Giudice, L.C.; Farrell, E.M.; Pham, H.; Rosenfeld, R.G. Identification of Insulin-Like Growth Factor-Binding Protein-3 (IGFBP-3) and IGFBP-2 in Human Follicular Fluid. J. Clin. Endocrinol. Metab. 1990, 71, 1330–1338. [Google Scholar] [CrossRef]
- Hossenlopp, P.; Segovia, B.; Lassarre, C.; Roghani, M.; Bredon, M.; Binoux, M. Evidence of Enzymatic Degradation of Insulin-Like Growth Factor-Binding Proteins in the 150K Complex during Pregnancy. J. Clin. Endocrinol. Metab. 1990, 71, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Fielder, P.J.; Thordarson, G.; Talamantes, F.; Rosenfeld, R.G. Characterization of Insulin-Like Growth Factor Binding Proteins (IGFBPs) during Gestation in Mice: Effects of Hypophysectomy and an IGFBP-Specific Serum Protease Activity. Endocrinology 1990, 127, 2270–2280. [Google Scholar] [CrossRef] [PubMed]
- Davenport, M.L.; Pucilowska, J.O.; Clemmons, D.R.; Lundblad, R.O.; Spencer, J.A.; Underwood, L.E. Tissue-specific expression of insulin-like growth factor binding protein-3 protease activity during rat pregnancy. Endocrinology 1992, 130, 2505–2512. [Google Scholar] [CrossRef]
- Lassarre, C.; Binoux, M. Insulin-like growth factor binding protein-3 is functionally altered in pregnancy plasma. Endocrinology 1994, 134, 1254–1262. [Google Scholar] [CrossRef][Green Version]
- Blat, C.; Villaudy, J.; Binoux, M. In vivo proteolysis of serum insulin-like growth factor (IGF) binding protein-3 results in increased availability of IGF to target cells. J. Clin. Investig. 1994, 93, 2286–2290. [Google Scholar] [CrossRef]
- Yan, X.; Baxter, R.; Firth, S.M. Involvement of Pregnancy-Associated Plasma Protein-A2 in Insulin-Like Growth Factor (IGF) Binding Protein-5 Proteolysis during Pregnancy: A Potential Mechanism for Increasing IGF Bioavailability. J. Clin. Endocrinol. Metab. 2010, 95, 1412–1420. [Google Scholar] [CrossRef]
- Byun, D.; Mohan, S.; Yoo, M.; Sexton, C.; Baylink, D.J.; Qin, X. Pregnancy-associated plasma protein-A accounts for the insulin-like growth factor (IGF)-binding protein-4 (IGFBP-4) proteolytic activity in human pregnancy serum and enhances the mitogenic activity of IGF by degrading IGFBP-4 in vitro. J. Clin. Endocrinol. Metab. 2001, 86, 847–854. [Google Scholar] [PubMed]
- Conover, C.A.; Boldt, H.B.; Bale, L.K.; Clifton, K.B.; Grell, J.A.; Mader, J.R.; Mason, E.J.; Powell, D.R. Pregnancy-Associated Plasma Protein-A2 (PAPP-A2): Tissue Expression and Biological Consequences of Gene Knockout in Mice. Endocrinology 2011, 152, 2837–2844. [Google Scholar] [CrossRef]
- Phang, D.; Rehage, M.; Bonafede, B.; Hou, D.; Xing, W.; Mohan, S.; Wergedal, J.E.; Qin, X. Inactivation of insulin-like-growth factors diminished the anabolic effects of pregnancy-associated plasma protein-A (PAPP-A) on bone in mice. Growth Horm. IGF Res. 2010, 20, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Ning, Y.; Schuller, A.G.P.; Conover, C.A.; Pintar, J.E. Insulin-like Growth Factor (IGF) Binding Protein-4 Is Both a Positive and Negative Regulator of IGF Activity in Vivo. Mol. Endocrinol. 2008, 22, 1213–1225. [Google Scholar] [CrossRef]
- Oxvig, C. The role of PAPP-A in the IGF system: Location, location, location. J. Cell Commun. Signal. 2015, 9, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Hjortebjerg, R.; Berryman, D.E.; Comisford, R.; List, E.O.; Oxvig, C.; Bjerre, M.; Frystyk, J.; Kopchick, J.J. Depot-specific and GH-dependent regulation of IGF binding protein-4, pregnancy-associated plasma protein-A, and stanniocalcin-2 in murine adipose tissue. Growth Horm. IGF Res. 2018, 39, 54–61. [Google Scholar] [CrossRef]
- Boldt, H.B.; Conover, C.A. Pregnancy-associated plasma protein-A (PAPP-A): A local regulator of IGF bioavailability through cleavage of IGFBPs. Growth Horm. IGF Res. 2007, 17, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Konev, A.A.; Smolyanova, T.I.; Kharitonov, A.V.; Serebryanaya, D.V.; Kozlovsky, S.V.; Kara, A.N.; Feygina, E.E.; Katrukha, A.G.; Postnikov, A.B. Characterization of endogenously circulating IGFBP-4 fragments—Novel biomarkers for cardiac risk assessment. Clin. Biochem. 2015, 48, 774–780. [Google Scholar] [CrossRef]
- Laursen, L.S.; Overgaard, M.T.; Søe, R.; Boldt, H.B.; Sottrup-Jensen, L.; Giudice, L.C.; Conover, C.A.; Oxvig, C. Pregnancy-associated plasma protein-A (PAPP-A) cleaves insulin-like growth factor binding protein (IGFBP)-5 independent of IGF: Implications for the mechanism of IGFBP-4 proteolysis by PAPP-A. FEBS Lett. 2001, 504, 36–40. [Google Scholar] [CrossRef]
- Hjortebjerg, R. IGFBP-4 and PAPP-A in normal physiology and disease. Growth Horm. IGF Res. 2018, 41, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Boldt, H.B.; Kjaer-Sorensen, K.; Overgaard, M.T.; Weyer, K.; Poulsen, C.B.; Sottrup-Jensen, L.; Conover, C.A.; Giudice, L.C.; Oxvig, C. The Lin12-Notch Repeats of Pregnancy-associated Plasma Protein-A Bind Calcium and Determine Its Proteolytic Specificity. J. Biol. Chem. 2004, 279, 38525–38531. [Google Scholar] [CrossRef]
- Rivera, S.; Khrestchatisky, M.; Kaczmarek, L.; Rosenberg, G.A.; Jaworski, D.M. Metzincin Proteases and Their Inhibitors: Foes or Friends in Nervous System Physiology? J. Neurosci. 2010, 30, 15337–15357. [Google Scholar] [CrossRef] [PubMed]
- Overgaard, M.T.; Boldt, H.B.; Laursen, L.S.; Sottrup-Jensen, L.; Conover, C.A.; Oxvig, C. Pregnancy-associated Plasma Protein-A2 (PAPP-A2), a Novel Insulin-like Growth Factor-binding Protein-5 Proteinase. J. Biol. Chem. 2001, 276, 21849–21853. [Google Scholar] [CrossRef] [PubMed]
- Glerup, S.; Boldt, H.B.; Overgaard, M.T.; Sottrup-Jensen, L.; Giudice, L.C.; Oxvig, C. Proteinase Inhibition by Proform of Eosinophil Major Basic Protein (pro-MBP) Is a Multistep Process of Intra- and Intermolecular Disulfide Rearrangements. J. Biol. Chem. 2005, 280, 9823–9832. [Google Scholar] [CrossRef]
- Rajah, R.; Katz, L.; Nunn, S.; Solberg, P.; Beers, T.; Cohen, P. Insulin-like growth factor binding protein (IGFBP) proteases: Functional regulators of cell growth. Prog. Growth Factor Res. 1995, 6, 273–284. [Google Scholar] [CrossRef]
- Wagner, G.; Hampong, M.; Park, C.; Copp, D. Purification, characterization, and bioassay of teleocalcin, a glycoprotein from salmon corpuscles of Stannius. Gen. Comp. Endocrinol. 1986, 63, 481–491. [Google Scholar] [CrossRef]
- Kløverpris, S.; Mikkelsen, J.H.; Pedersen, J.H.; Jepsen, M.; Laursen, L.S.; Petersen, S.V.; Oxvig, C. Stanniocalcin-1 Potently Inhibits the Proteolytic Activity of the Metalloproteinase Pregnancy-associated Plasma Protein-A. J. Biol. Chem. 2015, 290, 21915–21924. [Google Scholar] [CrossRef]
- Varghese, R.; Gagliardi, A.D.; Bialek, P.E.; Yee, S.P.; Wagner, G.F.; Dimattia, G.E. Overexpression of human stanniocalcin affects growth and reproduction in transgenic mice. Endocrinology 2002, 143, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Yoshiko, Y.; De Luca, F. Stanniocalcin 1 Acts as a Paracrine Regulator of Growth Plate Chondrogenesis. J. Biol. Chem. 2006, 281, 5120–5127. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.C.-M.; Hook, J.; Lemckert, F.A.; McDonald, M.; Nguyen, M.-A.T.; Hardeman, E.C.; Little, D.G.; Gunning, P.W.; Reddel, R. The Murine Stanniocalcin 2 Gene Is a Negative Regulator of Postnatal Growth. Endocrinology 2008, 149, 2403–2410. [Google Scholar] [CrossRef]
- Jepsen, M.; Kløverpris, S.; Mikkelsen, J.H.; Pedersen, J.H.; Füchtbauer, E.-M.; Laursen, L.S.; Oxvig, C. Stanniocalcin-2 Inhibits Mammalian Growth by Proteolytic Inhibition of the Insulin-like Growth Factor Axis. J. Biol. Chem. 2015, 290, 3430–3439. [Google Scholar] [CrossRef] [PubMed]
- Marouli, E.; Graff, M.; Medina-Gomez, C.; Lo, K.S.; Wood, A.R.; Kjaer, T.R.; Fine, R.S.; Lu, Y.; Schurmann, C.; Highland, H.M.; et al. Rare and low-frequency coding variants alter human adult height. Nat. Cell Biol. 2017, 542, 186–190. [Google Scholar] [CrossRef]
- Gagliardi, A.D.; Kuo, E.Y.W.; Raulic, S.; Wagner, G.F.; DiMattia, G.E. Human stanniocalcin-2 exhibits potent growth-suppressive properties in transgenic mice independently of growth hormone and IGFs. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E92–E105. [Google Scholar] [CrossRef]
- Dauber, A.; Muñoz-Calvo, M.T.; Barrios, V.; Domené, H.M.; Kloverpris, S.; Serra-Juhé, C.; Desikan, V.; Pozo, J.; Muzumdar, R.; Martos-Moreno, G.Á.; et al. Mutations in pregnancy-associated plasma protein A2 cause short stature due to low IGF-I availability. EMBO Mol. Med. 2016, 8, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Babiker, A.; Al Noaim, K.; Al Swaid, A.; Al Fadhel, M.; Deeb, A.; Martín-Rivada, Á.; Barrios, V.; Pérez-Jurado, L.A.; Alfares, A.; Al Alwan, I.; et al. Short stature with low insulin-like growth factor 1 availability due to pregnancy-associated plasma protein A2 deficiency in a Saudi family. Clin. Genet. 2021, 100, 601–606. [Google Scholar] [CrossRef]
- Argente, J. Challenges in the Management of Short Stature. Horm. Res. Paediatr. 2016, 85, 2–10. [Google Scholar] [CrossRef]
- Argente, J.; Tatton-Brown, K.; Lehwalder, D.; Pfäffle, R. Genetics of Growth Disorders-Which Patients Require Genetic Testing? Front. Endocrinol. 2019, 10, 602. [Google Scholar] [CrossRef] [PubMed]
- Karlberg, J. A Biologically Oriented Mathematical Model (ICP) for Human Growth. Acta Paediatr. Scand. Suppl. 1989, 350, 70–94. [Google Scholar] [CrossRef] [PubMed]
- Hawkins-Carranza, F.G.; Muñoz-Calvo, M.T.; Martos-Moreno, G.Á.; Allo-Miguel, G.; Del Río, L.; Pozo, J.; Chowen, J.A.; Pérez-Jurado, L.A.; Argente, J. rhIGF-1 Treatment Increases Bone Mineral Density and Trabecular Bone Structure in Children with PAPP-A2 Deficiency. Horm. Res. Paediatr. 2018, 89, 200–204. [Google Scholar] [CrossRef]
- Maddux, B.A.; Chan, A.; Mandarino, L.J.; Goldfine, I.D.; De Filippis, E.A. IGF-Binding Protein-1 Levels Are Related to Insulin-Mediated Glucose Disposal and Are a Potential Serum Marker of Insulin Resistance. Diabetes Care 2006, 29, 1535–1537. [Google Scholar] [CrossRef][Green Version]
- Heath, K.E.; Argente, J.; Barrios, V.; Pozo, J.; Díaz-González, F.; Martos-Moreno, G.A.; Caimari, M.; Gracia, R.; Campos-Barros, A. Primary acid-labile subunit deficiency due to recessive IGFALS mutations results in postnatal growth deficit associated with low circulating insulin growth factor (IGF)-I, IGF binding protein-3 levels, and hyperinsulinemia. J. Clin. Endocrinol. Metab. 2008, 93, 1616–1624. [Google Scholar] [CrossRef]
- Muñoz-Calvo, M.T.; Barrios, V.; Pozo, J.; Chowen, J.; Martos-Moreno, G.Á.; Hawkins, F.; Dauber, A.; Domené, H.M.; Yakar, S.; Rosenfeld, R.G.; et al. Treatment With Recombinant Human Insulin-Like Growth Factor-1 Improves Growth in Patients With PAPP-A2 Deficiency. J. Clin. Endocrinol. Metab. 2016, 101, 3879–3883. [Google Scholar] [CrossRef]
- Martín-Rivada, Á.; Barrios, V.; Díaz-Guerra, G.M.; Pozo, J.; Díaz-Guerra, G.M.; Pozo, J.; Martos-Moreno, G.Á.; Argente, J. Adult height and long-term outcomes after rhIGF-1 therapy in two patients with PAPP-A2 deficiency. Growth Horm. IGF Res. 2021, 60-61, 101419. [Google Scholar] [CrossRef]
- Fujimoto, M.; Andrew, M.; Dauber, A. Disorders caused by genetic defects associated with GH-dependent genes: PAPPA2 defects. Mol. Cell. Endocrinol. 2020, 518, 110967. [Google Scholar] [CrossRef] [PubMed]
- Farr, M.; Strübe, J.; Geppert, H.-G.; Kocourek, A.; Mahne, M.; Tschesche, H. Pregnancy-associated plasma protein-E (PAPP-E). Biochim. Biophys. Acta BBA-Gene Struct. Expr. 2000, 1493, 356–362. [Google Scholar] [CrossRef]
- Page, N.; Butlin, D.; Lomthaisong, K.; Lowry, P. The Characterization of Pregnancy Associated Plasma Protein-E and the Identification of an Alternative Splice Variant. Placenta 2001, 22, 681–687. [Google Scholar] [CrossRef]
- Wagner, P.K.; Otomo, A.; Christians, J.K. Regulation of pregnancy-associated plasma protein A2 (PAPPA2) in a human placental trophoblast cell line (BeWo). Reprod. Biol. Endocrinol. 2011, 9, 48. [Google Scholar] [CrossRef]
- Christians, J.K.; Hoeflich, A.; Keightley, P.D. PAPPA2, an Enzyme That Cleaves an Insulin-Like Growth-Factor-Binding Protein, Is a Candidate Gene for a Quantitative Trait Locus Affecting Body Size in Mice. Genetics 2006, 173, 1547–1553. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Qiu, Q.; Haider, M.; Bell, M.; Gruslin, A.; Christians, J.K. Expression of pregnancy-associated plasma protein A2 during pregnancy in human and mouse. J. Endocrinol. 2009, 202, 337–345. [Google Scholar] [CrossRef]
- Chen, Y.; Lv, H.; Li, L.; Wang, E.; Zhang, L.; Zhao, Q. Expression of PAPP-A2 and IGF Pathway-Related Proteins in the Hip Joint of Normal Rat and Those with Developmental Dysplasia of the Hip. Int. J. Endocrinol. 2019, 2019, 7691531. [Google Scholar] [CrossRef]
- Amiri, N.; Christians, J.K. PAPP-A2 expression by osteoblasts is required for normal postnatal growth in mice. Growth Horm. IGF Res. 2015, 25, 274–280. [Google Scholar] [CrossRef]
- Kjaer-Sorensen, K.; Engholm, D.H.; Jepsen, M.R.; Morch, M.G.; Weyer, K.; Hefting, L.L.; Skov, L.L.; Laursen, L.S.; Oxvig, C. Papp-a2 modulates development of cranial cartilage and angiogenesis in zebrafish embryos. J. Cell Sci. 2014, 127 Pt 23, 5027–5037. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fujimoto, M.; Andrew, M.; Liao, L.; Zhang, D.; Yildirim, G.; Sluss, P.; Kalra, B.; Kumar, A.; Yakar, S.; Hwa, V.; et al. Low IGF-I Bioavailability Impairs Growth and Glucose Metabolism in a Mouse Model of Human PAPPA2 p.Ala1033Val Mutation. Endocrinology 2019, 160, 1363–1376. [Google Scholar] [CrossRef]
- Christians, J.K.; de Zwaan, D.; Fung, S.H.Y. Pregnancy Associated Plasma Protein A2 (PAPP-A2) Affects Bone Size and Shape and Contributes to Natural Variation in Postnatal Growth in Mice. PLoS ONE 2013, 8, e56260. [Google Scholar] [CrossRef]
- Christians, J.K.; Bath, A.K.; Amiri, N. Pappa2 deletion alters IGFBPs but has little effect on glucose disposal or adiposity. Growth Horm. IGF Res. 2015, 25, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Rubio, L.; Vargas, A.; Rivera, P.; López-Gambero, A.; Tovar, R.; Christians, J.; Martín-De-Las-Heras, S.; de Fonseca, F.R.; Chowen, J.; Argente, J.; et al. Recombinant IGF-1 Induces Sex-Specific Changes in Bone Composition and Remodeling in Adult Mice with Pappa2 Deficiency. Int. J. Mol. Sci. 2021, 22, 4048. [Google Scholar] [CrossRef]
- Rogowska, M.D.; Pena, U.N.V.; Binning, N.; Christians, J.K. Recovery of the maternal skeleton after lactation is impaired by advanced maternal age but not by reduced IGF availability in the mouse. PLoS ONE 2021, 16, e0256906. [Google Scholar] [CrossRef]
- Christians, J.K.; King, A.Y.; Rogowska, M.D.; Hessels, S.M. Pappa2 deletion in mice affects male but not female fertility. Reprod. Biol. Endocrinol. 2015, 13, 109. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.L.; DeMambro, V.E.; Guntur, A.R.; Le, P.; Nagano, K.; Baron, R.; de Paula, F.J.A.; Motyl, K.J. High fat diet attenuates hyperglycemia, body composition changes, and bone loss in male streptozotocin-induced type 1 diabetic mice. J. Cell. Physiol. 2018, 233, 1585–1600. [Google Scholar] [CrossRef] [PubMed]
- Yanagihara, G.R.; Shimano, R.C.; Tida, J.A.; Yamanaka, J.S.; Fukada, S.Y.; Issa, J.P.M.; Tavares, J.M.R.S. Influence of High-Fat Diet on Bone Tissue: An Experimental Study in Growing Rats. J. Nutr. Health Aging 2017, 21, 1337–1343. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Wu, Y.-W.; Ren, Y.-Z. The impact of a high fat diet on bones: Potential mechanisms. Food Funct. 2021, 12, 963–975. [Google Scholar] [CrossRef]
- Kawai, M.; Rosen, C.J. The IGF-I regulatory system and its impact on skeletal and energy homeostasis. J. Cell. Biochem. 2010, 111, 14–19. [Google Scholar] [CrossRef]
- Christians, J.K.; Amiri, N.; Schipilow, J.D.; Zhang, S.W.; May-Rashke, K.I. Pappa2 deletion has sex- and age-specific effects on bone in mice. Growth Horm. IGF Res. 2019, 44, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Mohan, S.; Yakar, S. Does the GH/IGF-1 axis contribute to skeletal sexual dimorphism? Evidence from mouse studies. Growth Horm. IGF Res. 2016, 27, 7–17. [Google Scholar] [CrossRef]
- Olson, L.E.; Ohlsson, C.; Mohan, S. The Role of GH/IGF-I-Mediated Mechanisms in Sex Differences in Cortical Bone Size in Mice. Calcif. Tissue Int. 2010, 88, 1–8. [Google Scholar] [CrossRef]
- Vanderschueren, D.; Venken, K.; Ophoff, J.; Bouillon, R.; Boonen, S. Clinical Review: Sex steroids and the periosteum--reconsidering the roles of androgens and estrogens in periosteal expansion. J. Clin. Endocrinol. Metab. 2006, 91, 378–382. [Google Scholar] [CrossRef]
- Nuti, R.; Martini, G.; Merlotti, D.; De Paola, V.; Valleggi, F.; Gennari, L. Bone metabolism in men: Role of aromatase activity. J. Endocrinol. Investig. 2007, 30 (Suppl. 6), 18–23. [Google Scholar]
- Schutz, H.; Donovan, E.R.; Hayes, J.P. Effects of parity on pelvic size and shape dimorphism in Mus. J. Morphol. 2009, 270, 834–842. [Google Scholar] [CrossRef]
- Jia, J.; Li, L.; Zhao, Q.; Zhang, L.; Ru, J.; Liu, X.; Li, Q.; Shi, L. Association of a single nucleotide polymorphism in pregnancy-associated plasma protein-A2 with developmental dysplasia of the hip: A case–control study. Osteoarthr. Cartil. 2012, 20, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Christians, J.K.; Shergill, H.K.; Albert, A.Y.K. Sex-dependent effects of prenatal food and protein restriction on offspring physiology in rats and mice: Systematic review and meta-analyses. Biol. Sex Differ. 2021, 12, 21. [Google Scholar] [CrossRef]
- Ivaska, K.K.; Lenora, J.; Gerdhem, P.; Åkesson, K.; Väänänen, H.K.; Obrant, K.J. Serial Assessment of Serum Bone Metabolism Markers Identifies Women with the Highest Rate of Bone Loss and Osteoporosis Risk. J. Clin. Endocrinol. Metab. 2008, 93, 2622–2632. [Google Scholar] [CrossRef]
- Wickramasinghe, S.; Rincon, G.; Medrano, J. Variants in the pregnancy-associated plasma protein-A2 gene on Bos taurus autosome 16 are associated with daughter calving ease and productive life in Holstein cattle. J. Dairy Sci. 2011, 94, 1552–1558. [Google Scholar] [CrossRef]
- Cabrera-Salcedo, C.; Mizuno, T.; Tyzinski, L.; Andrew, M.; Vinks, A.; Frystyk, J.; Wasserman, H.; Gordon, C.M.; Hwa, V.; Backeljauw, P.; et al. Pharmacokinetics of IGF-1 in PAPP-A2-Deficient Patients, Growth Response, and Effects on Glucose and Bone Density. J. Clin. Endocrinol. Metab. 2017, 102, 4568–4577. [Google Scholar] [CrossRef] [PubMed]
- Sandstedt, J.; Törnell, J.; Norjavaara, E.; Isaksson, O.G.; Ohlsson, C. Elevated levels of growth hormone increase bone mineral content in normal young mice, but not in ovariectomized mice. Endocrinology 1996, 137, 3368–3374. [Google Scholar] [CrossRef] [PubMed]
- Narusawa, K.I.; Nakamura, T.; Suzuki, K.; Matsuoka, Y.; Lee, L.J.; Tanaka, H.; Seino, Y. The effects of recombinant human insulin-like growth factor (rhIGF)-1 and rhIGF-1/IGF binding protein-3 administration on rat osteopenia induced by ovariectomy with concomitant bilateral sciatic neurectomy. J. Bone Miner Res. 1995, 10, 1853–1864. [Google Scholar] [CrossRef] [PubMed]
- Åberg, D. Role of the Growth Hormone/Insulin-Like Growth Factor 1 Axis in Neurogenesis. Endocr. Dev. 2010, 17, 63–76. [Google Scholar]
- Ashpole, N.M.; Sanders, J.E.; Hodges, E.L.; Yan, H.; Sonntag, W.E. Growth hormone, insulin-like growth factor-1 and the aging brain. Exp. Gerontol. 2015, 68, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Argente, J.; Chowen, J.A.; Pérez-Jurado, L.A.; Frystyk, J.; Oxvig, C. One level up: Abnormal proteolytic regulation of IGF activity plays a role in human pathophysiology. EMBO Mol. Med. 2017, 9, 1338–1345. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
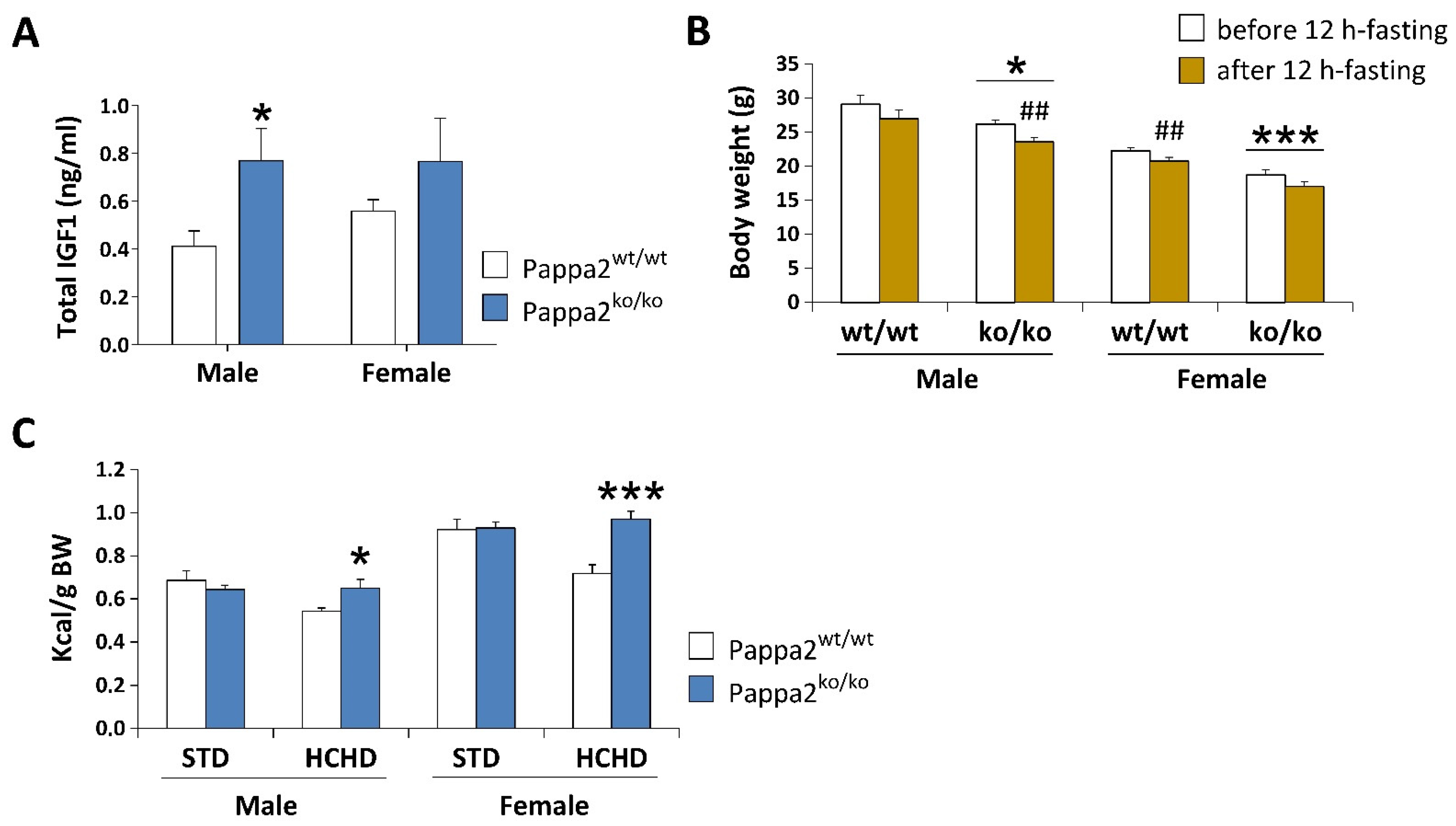{kind=link}
| Constitutive Pappa2 KO | Constitutive Pappa2 KO | Constitutive Induction of Pappa2 KO | Conditional Pappa2 KO in Osteoblasts | Constitutive Human PAPP-A2 KI | |
|---|---|---|---|---|---|
| Main references | Conover et al. 2011 | Christians et al. 2013, 2015a, 2019; Rogowska et al. 2021; Rubio et al. 2021 | Christians et al. 2015a | Amiri and Christians, 2015 | Fujimoto et al. 2019 |
| Auxological parameters | |||||
| Body weight (BW) | Ns 1 in males Reduction in females | Reduction | ns | Reduction | Reduction |
| Body length | Ns in males Reduction in females | Reduction | -- | Reduction in tail | Reduction |
| Organ size | Ns in males Increase in females | ns | -- | -- | Increase in liver |
| Body composition | -- | -- | -- | -- | Higher fat mass. Lower lean mass |
| Hormonal parameters | |||||
| Free Igf1 | Decrease | Decrease | -- | -- | Decrease |
| Total Igf1 | Increase | Increase | -- | -- | Increase |
| Igfbp5 | -- | Increase in plasma Ns in liver and kidney Increase in ovaries Ns in tibia | ns | ns | -- |
| Igfbp3 | -- | Decrease in serum Ns in liver and kidney Increase in tibia | Decrease | -- | Increase |
| Igfals | -- | Ns in tibia | -- | -- | Increase |
| Energy metabolism | |||||
| Glucose tolerance | -- | ns | -- | -- | Intolerant |
| Insulin sensitivity | -- | ns | -- | -- | Resistant |
| Adiposity | ns | -- | -- | -- | |
| BW loss | -- | Increase in fasting | -- | -- | -- |
| Caloric intake | -- | Increase in HCHD 2 | -- | -- | -- |
| Allometric parameters | |||||
| Femur length | ns | Reduction | -- | Reduction | Reduction |
| Femur weight | -- | Reduction | -- | -- | -- |
| Other bone dimensions | -- | Defects | -- | Defects | -- |
| Bone shape | -- | Defects (pelvic girdle and mandible) | -- | Defects (pelvic girdle and mandible) | -- |
| Bone mineral properties | |||||
| Bone mineral content | Decreases in trabecular and cortical femur | Decrease in trabecular femur Increase in cortical femur | -- -- | -- -- | Decrease in trabecular femur Ns in cortical femur |
| Bone mineral composition | -- | Alterations in male femur | -- | -- | -- |
| Collagen maturity | Decrease in female femur | -- | -- | -- | |
| Bone remodeling | |||||
| Bone formation and resorption | -- | Decreases in female serum Increases in female tibia | -- | -- | -- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrios, V.; Chowen, J.A.; Martín-Rivada, Á.; Guerra-Cantera, S.; Pozo, J.; Yakar, S.; Rosenfeld, R.G.; Pérez-Jurado, L.A.; Suárez, J.; Argente, J. Pregnancy-Associated Plasma Protein (PAPP)-A2 in Physiology and Disease. Cells 2021, 10, 3576. https://doi.org/10.3390/cells10123576
Barrios V, Chowen JA, Martín-Rivada Á, Guerra-Cantera S, Pozo J, Yakar S, Rosenfeld RG, Pérez-Jurado LA, Suárez J, Argente J. Pregnancy-Associated Plasma Protein (PAPP)-A2 in Physiology and Disease. Cells. 2021; 10(12):3576. https://doi.org/10.3390/cells10123576
Chicago/Turabian StyleBarrios, Vicente, Julie A. Chowen, Álvaro Martín-Rivada, Santiago Guerra-Cantera, Jesús Pozo, Shoshana Yakar, Ron G. Rosenfeld, Luis A. Pérez-Jurado, Juan Suárez, and Jesús Argente. 2021. "Pregnancy-Associated Plasma Protein (PAPP)-A2 in Physiology and Disease" Cells 10, no. 12: 3576. https://doi.org/10.3390/cells10123576
APA StyleBarrios, V., Chowen, J. A., Martín-Rivada, Á., Guerra-Cantera, S., Pozo, J., Yakar, S., Rosenfeld, R. G., Pérez-Jurado, L. A., Suárez, J., & Argente, J. (2021). Pregnancy-Associated Plasma Protein (PAPP)-A2 in Physiology and Disease. Cells, 10(12), 3576. https://doi.org/10.3390/cells10123576