
Advanced Glycation End-Products (AGEs) and Their Soluble Receptor (sRAGE) in Women Suffering from Systemic Lupus Erythematosus (SLE)

 ,
,  ,
,
Abstract
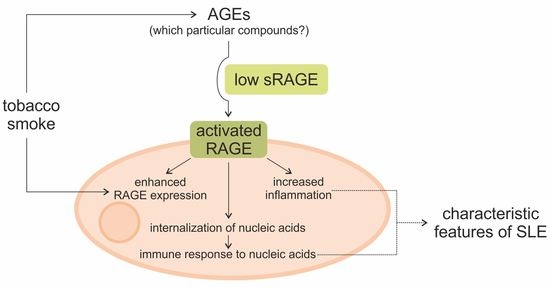
1. Introduction
2. Materials and Methods
2.1. Human Subjects
2.2. ELISA Assay
- OxiSelect™ Advanced Glycation End Product (AGE) Competitive ELISA Kit, catalogue number STA-817 (Cell Biolabs, Inc., San Diego, CA, USA) sensitivity 0.39 μg/mL; precision measured as coefficient of variation < 5% (intra-assay), <10% (inter-assay);
- OxiSelect™ N-epsilon-(Carboxymethyl) Lysine (CML) Competitive ELISA Kit, catalogue number STA-816 (Cell Biolabs, Inc., San Diego, CA, USA) sensitivity 0.05 μg/mL; precision measured as coefficient of variation < 6% (intra-assay), <10% (inter-assay);
- OxiSelect™ N-epsilon-(Carboxyethyl) Lysine (CEL) Competitive ELISA, catalogue number STA-813 (Cell Biolabs, Inc., San Diego, CA, USA) sensitivity 0.1 μg/mL; precision measured as coefficient of variation < 6% (intra-assay), <10% (inter-assay);
- PTD (Pentosidine) ELISA Kit, catalogue number E-EL-0091 (Elabscience, Houston, TX, USA) sensitivity 0.47 ng/mL; precision measured as coefficient of variation < 6% (intra-assay and inter-assay);
- RayBio® Human RAGE ELISA Kit, catalogue number ELH-RAGE (RayBiotech, Norcross, GA, USA) sensitivity 3 pg/mL; precision measured as coefficient of variation < 10% (intra-assay), <12% (inter-assay).
3. Results
3.1. Characteristics of the Human Subjects
3.2. ELISA Assay Results
3.3. Correlation of the Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Somers, E.C.; Marder, W.; Cagnoli, P.; Lewis, E.E.; DeGuire, P.; Gordon, C.; Helmick, C.G.; Wang, L.; Wing, J.J.; Dhar, J.P.; et al. Population-based incidence and prevalence of systemic lupus erythematosus: The Michigan lupus epidemiology and surveillance program. Arthritis Rheumatol. 2014, 66, 369–378. [Google Scholar] [CrossRef]
- Cooper, G.S.; Bynum, M.L.K.; Somers, E.C. Recent insights in the epidemiology of autoimmune diseases: Improved prevalence estimates and understanding of clustering of diseases. J. Autoimmun. 2009, 33, 197–207. [Google Scholar] [CrossRef]
- Costenbader, K.H.; Feskanich, D.; Stampfer, M.J.; Karlson, E.W. Reproductive and menopausal factors and risk of systemic lupus erythematosus in women. Arthritis Rheum. 2007, 56, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Catalina, M.D.; Owen, K.A.; Labonte, A.C.; Grammer, A.C.; Lipsky, P.E. The pathogenesis of systemic lupus erythematosus: Harnessing big data to understand the molecular basis of lupus. J. Autoimmun. 2020, 110, 102359. [Google Scholar] [CrossRef]
- Martens, H.A.; Nienhuis, H.L.A.; Gross, S.; Van Der Steege, G.; Brouwer, E.; Berden, J.H.M.; De Sévaux, R.G.L.; Derksen, R.H.W.M.; Voskuyl, A.E.; Berger, S.P.; et al. Receptor for advanced glycation end products (RAGE) polymorphisms are associated with systemic lupus erythematosus and disease severity in lupus nephritis. Lupus 2012, 21, 959–968. [Google Scholar] [CrossRef]
- Bates, M.A.; Brandenberger, C.; Langohr, I.; Kumagai, K.; Harkema, J.R.; Holian, A.; Pestka, J.J. Silica triggers inflammation and ectopic lymphoid neogenesis in the lungs in parallel with accelerated onset of systemic autoimmunity and glomerulonephritis in the lupus-prone NZBWF1 mouse. PLoS ONE 2015, 10, 0125481. [Google Scholar] [CrossRef]
- Speyer, C.B.; Costenbader, K.H. Cigarette smoking and the pathogenesis of systemic lupus erythematosus. Expert Rev. Clin. Immunol. 2018, 14, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Barbhaiya, M.; Costenbader, K.H. Environmental exposures and the development of systemic lupus erythematosus. Curr. Opin. Rheumatol. 2016, 28, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Tsokos, G.C. Mechanisms of disease: Systemic lupus erythematosus. N. Engl. J. Med. 2011, 365, 2110–21121. [Google Scholar] [CrossRef]
- Yaniv, G.; Twig, G.; Shor, D.B.A.; Furer, A.; Sherer, Y.; Mozes, O.; Komisar, O.; Slonimsky, E.; Klang, E.; Lotan, E.; et al. A volcanic explosion of autoantibodies in systemic lupus erythematosus: A diversity of 180 different antibodies found in SLE patients. Autoimmun. Rev. 2015, 14, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Nienhuis, H.L.; de Leeuw, K.; Bijzet, J.; Smit, A.; Schalkwijk, C.G.; Graaff, R.; Kallenberg, C.G.; Bijl, M. Skin autofluorescence is increased in systemic lupus erythematosus but is not reflected by elevated plasma levels of advanced glycation endproducts. Rheumatology 2008, 47, 1554–1558. [Google Scholar] [CrossRef]
- Aguirre-Valencia, D.; Suárez-Avellaneda, A.; Ocampo-Piraquive, V.; Posso-Osorio, I.; Naranjo-Escobar, J.; Nieto-Aristizábal, I.; Tobón, G.J.; Cañas, C.A. Mortality in patients with systemic lupus erythematosus in Colombia: A case series. Clin. Rheumatol. 2019, 38, 1865–1871. [Google Scholar] [CrossRef]
- Tselios, K.; Gladman, D.D.; Sheane, B.J.; Su, J.; Urowitz, M. All-cause, cause-specific and age-specific standardised mortality ratios of patients with systemic lupus erythematosus in Ontario, Canada over 43 years (1971–2013). Ann. Rheum. Dis. 2019, 78, 802–806. [Google Scholar] [CrossRef] [PubMed]
- Ene, C.D.; Georgescu, S.R.; Tampa, M.; Matei, C.; Mitran, C.I.; Mitran, M.I.; Penescu, M.N.; Nicolae, I. Cellular response against oxidative stress, a novel insight into lupus nephritis pathogenesis. J. Pers. Med. 2021, 11, 693. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Mir, A.R.; Abidi, M.; Talha, M.; Zafar, A.; Habib, S.; Moinuddin. Methylglyoxal modified IgG generates autoimmune response in rheumatoid arthritis. Int. J. Biol. Macromol. 2018, 118, 15–23. [Google Scholar] [CrossRef]
- Sirois, C.M.; Jin, T.; Miller, A.L.; Bertheloot, D.; Nakamura, H.; Horvath, G.L.; Mian, A.; Jiang, J.; Schrum, J.; Bossaller, L.; et al. RAGE is a nucleic acid receptor that promotes inflammatory responses to DNA. J. Exp. Med. 2013, 210, 2447–2463. [Google Scholar] [CrossRef] [PubMed]
- Jyoti; Mir, A.R.; Habib, S.; Siddiqui, S.S.; Ali, A.; Moinuddin. Neo-epitopes on methylglyoxal modified human serum albumin lead to aggressive autoimmune response in diabetes. Int. J. Biol. Macromol. 2016, 86, 799–809. [Google Scholar] [CrossRef]
- Henning, C.; Glomb, M.A. Pathways of the Maillard reaction under physiological conditions. Glycoconj. J. 2016, 33, 499–512. [Google Scholar] [CrossRef]
- Nigro, C.; Leone, A.; Fiory, F.; Prevenzano, I.; Nicolò, A.; Mirra, P.; Beguinot, F.; Miele, C. Dicarbonyl stress at the crossroads of healthy and unhealthy aging. Cells 2019, 8, 749. [Google Scholar] [CrossRef]
- Henning, C.; Liehr, K.; Girndt, M.; Ulrich, C.; Glomb, M.A. Analysis and chemistry of novel protein oxidation markers in vivo. J. Agric. Food Chem. 2018, 66, 4692–4701. [Google Scholar] [CrossRef]
- Vytášek, R.; Šedová, L.; Vilím, V. Increased concentration of two different advanced glycation end products detected by enzyme immunoassays with new monoclonal antibodies in sera of patients with rheumatoid arthritis. BMC Musculoskelet. Disord. 2010, 11, 83. [Google Scholar] [CrossRef]
- Chen, D.Y.; Chen, Y.M.; Lin, C.C.; Hsieh, C.W.; Wu, Y.C.; Hung, W.T.; Chen, H.H.; Lan, J.L. The potential role of advanced glycation end products (AGEs) and soluble receptors for AGEs (sRAGE) in the pathogenesis of adult-onset Still’s disease. BMC Musculoskelet. Disord. 2015, 16, 111. [Google Scholar] [CrossRef]
- De Groot, L.; Posthumus, M.D.; Kallenberg, C.G.M.; Bijl, M. Risk factors and early detection of atherosclerosis in rheumatoid arthritis. Eur. J. Clin. Investig. 2010, 40, 835–842. [Google Scholar] [CrossRef]
- Prasad, C.; Davis, K.E.; Imrhan, V.; Juma, S.; Vijayagopal, P. Advanced glycation end products and risks for chronic diseases: Intervening through lifestyle modification. Am. J. Lifestyle Med. 2019, 13, 384–404. [Google Scholar] [CrossRef] [PubMed]
- Gopal, P.; Reynaert, N.L.; Scheijen, J.L.J.M.; Engelen, L.; Schalkwijk, C.G.; Franssen, F.M.E.; Wouters, E.F.M.; Rutten, E.P.A. Plasma advanced glycation end products and skin autofluorescence are increased in COPD. Eur. Respir. J. 2014, 43, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Fishman, S.L.; Sonmez, H.; Basman, C.; Singh, V.; Poretsky, L. The role of advanced glycation end products in the development of coronary artery disease in patients with and without diabetes mellitus: A review. Mol. Med. 2018, 24. [Google Scholar] [CrossRef] [PubMed]
- De Leeuw, K.; Graaff, R.; de Vries, R.; Dullaart, R.P.; Smit, A.J.; Kallenberg, C.G.; Bijl, M. Accumulation of advanced glycation endproducts in patients with systemic lupus erythematosus. Rheumatology 2007, 46, 1551–1556. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-García, J.; Requena, J.R.; Rodríguez-Segade, S. Increased concentrations of serum pentosidine in rheumatoid arthritis. Clin. Chem. 1998, 44, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Zieman, S.J.; Melenovsky, V.; Clattenburg, L.; Corretti, M.C.; Capriotti, A.; Gerstenblith, G.; Kass, D.A. Advanced glycation endproduct crosslink breaker (alagebrium) improves endothelial function in patients with isolated systolic hypertension. J. Hypertens. 2007, 25, 577–583. [Google Scholar] [CrossRef]
- Dobi, A.; Bravo, S.B.; Veeren, B.; Paradela-Dobarro, B.; Álvarez, E.; Meilhac, O.; Viranaicken, W.; Baret, P.; Devin, A.; Rondeau, P. Advanced glycation end products disrupt human endothelial cells redox homeostasis: New insights into reactive oxygen species production. Free Radic. Res. 2019, 53, 150–169. [Google Scholar] [CrossRef]
- Tan, K.C.B.; Chow, W.S.; Ai, V.H.G.; Metz, C.; Bucala, R.; Lam, K.S.L. Advanced glycation end products and endothelial dysfunction in type 2 diabetes. Diabetes Care 2002, 25, 1055–1059. [Google Scholar] [CrossRef]
- Deluyker, D.; Evens, L.; Bito, V. Advanced glycation end products (AGEs) and cardiovascular dysfunction: Focus on high molecular weight AGEs. Amino Acids 2017, 49, 1535–1541. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Son, M.; Choi, J.; Lee, S.; Byun, K. sRAGE prolonged stem cell survival and suppressed RAGE-related inflammatory cell and T lymphocyte accumulations in an Alzheimer’s disease model. Biochem. Biophys. Res. Commun. 2018, 495, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Lange, J.N.; Wood, K.D.; Knight, J.; Assimos, D.G.; Holmes, R.P. Glyoxal formation and its role in endogenous oxalate synthesis. Adv. Urol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.T.; Wu, T.H.; Lin, C.S.; Lee, C.S.; Wei, Y.H.; Tsai, C.Y.; Chang, D.M. The pathogenesis of systemic lupus erythematosus—From the viewpoint of oxidative stress and mitochondrial dysfunction. Mitochondrion 2016, 30, 1–7. [Google Scholar] [CrossRef]
- Shah, D.; Mahajan, N.; Sah, S.; Nath, S.K.; Paudyal, B. Oxidative stress and its biomarkers in systemic lupus erythematosus. J. Biomed. Sci. 2014, 21, 23. [Google Scholar] [CrossRef]
- Smallwood, M.J.; Nissim, A.; Knight, A.R.; Whiteman, M.; Haigh, R.; Winyard, P.G. Oxidative stress in autoimmune rheumatic diseases. Free Radic. Biol. Med. 2018, 125, 3–14. [Google Scholar] [CrossRef]
- Nasiri, R.; Field, M.J.; Zahedi, M.; Moosavi-Movahedi, A.A. Comparative DFT study to determine if α-oxoaldehydes are precursors for pentosidine formation. J. Phys. Chem. A 2012, 116, 2986–2996. [Google Scholar] [CrossRef] [PubMed]
- Raupbach, J.; Ott, C.; Koenig, J.; Grune, T. Proteasomal degradation of glycated proteins depends on substrate unfolding: Preferred degradation of moderately modified myoglobin. Free Radic. Biol. Med. 2020, 152, 516–524. [Google Scholar] [CrossRef]
- Bayoumy, N.; El-Shabrawi, M.; Nada, H. A soluble receptor for advanced glycation end product levels in patients with systemic lupus erythematosus. Turkish J. Rheumatol. 2013, 28, 101–108. [Google Scholar] [CrossRef]
- Ma, C.Y.; Ma, J.L.; Jiao, Y.L.; Li, J.F.; Wang, L.C.; Yang, Q.R.; You, L.; Cui, B.; Chen, Z.J.; Zhao, Y.R. The plasma level of soluble receptor for advanced glycation end products is decreased in patients with systemic lupus erythematosus. Scand. J. Immunol. 2012, 75, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Bobek, D.; Grčević, D.; Kovačić, N.; Lukić, K.K.; Jelušić, M. The presence of high mobility group box-1 and soluble receptor for advanced glycation end products in juvenile idiopathic arthritis and juvenile systemic lupus erythematosus. Pediatr. Rheumatol. 2014, 12, 1–9. [Google Scholar] [CrossRef]
- Tang, K.T.; Hsieh, T.Y.; Chao, Y.H.; Lin, M.X.; Chen, Y.H.; Chen, D.Y.; Lin, C.C. Plasma levels of high-mobility group box 1 and soluble receptor for advanced glycation end products in primary antiphospholipid antibody syndrome patients. PLoS ONE 2017, 12, e0178404. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.L.; Wong, C.K.; Szeto, C.C.; Li, E.K.; Cai, Z.; Tam, L.S. Members of the receptor for advanced glycation end products axis as potential therapeutic targets in patients with lupus nephritis. Lupus 2015, 24, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Abou-Raya, A.N.; Kamel, M.A.N.; Sayed, E.A.G.; El-Sharkawy, A.A.H. The plasma level of soluble receptor for advanced glycation end products in systemic lupus erythematosus patients and its relation to disease activity. Alexandria J. Med. 2016, 52, 151–157. [Google Scholar] [CrossRef][Green Version]
- Manganelli, V.; Truglia, S.; Capozzi, A.; Alessandri, C.; Riitano, G.; Spinelli, F.R.; Ceccarelli, F.; Mancuso, S.; Garofalo, T.; Longo, A.; et al. Alarmin HMGB1 and soluble RAGE as new tools to evaluate the risk stratification in patients with the antiphospholipid syndrome. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Chavakis, T.; Bierhaus, A.; Nawroth, P.P. RAGE (receptor for advanced glycation end products): A central player in the inflammatory response. Microbes Infect. 2004, 6, 1219–1225. [Google Scholar] [CrossRef]
- Munguia-Realpozo, P.; Mendoza-Pinto, C.; Sierra Benito, C.; Escarcega, R.O.; Garcia-Carrasco, M.; Mendez Martinez, S.; Etchegaray Morales, I.; Galvez Romero, J.L.; Ruiz-Arguelles, A.; Cervera, R. Systemic lupus erythematosus and hypertension. Autoimmun. Rev. 2019, 18. [Google Scholar] [CrossRef] [PubMed]
- Zeller, C.; Appenzeller, S. Cardiovascular disease in systemic lupus erythematosus: The role of traditional and lupus related risk factors. Curr. Cardiol. Rev. 2008, 4, 116–122. [Google Scholar] [CrossRef]
- Van Waateringe, R.P.; Mook-Kanamori, M.J.; Slagter, S.N.; Van Der Klauw, M.M.; Van Vliet-Ostaptchouk, J.V.; Graaff, R.; Lutgers, H.L.; Suhre, K.; El-Din Selim, M.M.; Mook-Kanamori, D.O.; et al. The association between various smoking behaviors, cotinine biomarkers and skin autofluorescence, a marker for advanced glycation end product accumulation. PLoS ONE 2017, 12, e0179330. [Google Scholar] [CrossRef]
- Mesaros, C.; Arora, J.S.; Wholer, A.; Vachani, A.; Blair, I.A. 8-oxo-2′-deoxyguanosine as a biomarker of tobacco-smoking-induced oxidative stress. Free Radic. Biol. Med. 2012, 53, 610–617. [Google Scholar] [CrossRef]
- Lee, H.T.; Lin, C.S.; Lee, C.S.; Tsai, C.Y.; Wei, Y.H. Increased 8-hydroxy-2′-deoxyguanosine in plasma and decreased mRNA expression of human 8-oxoguanine DNA glycosylase 1, anti-oxidant enzymes, mitochondrial biogenesis-related proteins and glycolytic enzymes in leucocytes in patients with systemic lupus ery. Clin. Exp. Immunol. 2014, 176, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, P.R.; Kasteler, S.D.; Cosio, M.G.; Sturrock, A.; Huecksteadt, T.; Hoidal, J.R. RAGE: Developmental expression and positive feedback regulation by Egr-1 during cigarette smoke exposure in pulmonary epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 294. [Google Scholar] [CrossRef]
- Wang, J.; Kay, A.B.; Fletcher, J.; Formica, M.K.; McAlindon, T.E. Alcohol consumption is not protective for systemic lupus erythematosus. Ann. Rheum. Dis. 2009, 68, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Pan, H.F.; Ye, D.Q.; Su, H.; Li, X.P. Moderate alcohol drinking might be protective for systemic lupus erythematosus: A systematic review and meta-analysis. Clin. Rheumatol. 2008, 27, 1557–1563. [Google Scholar] [CrossRef]
- Barbhaiya, M.; Lu, B.; Sparks, J.A.; Malspeis, S.; Chang, S.C.; Karlson, E.W.; Costenbader, K.H. Influence of alcohol consumption on the risk of systemic lupus erythematosus among women in the Nurses’ Health Study Cohorts. Arthritis Care Res. 2017, 69, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Cozier, Y.C.; Barbhaiya, M.; Castro-Webb, N.; Conte, C.; Tedeschi, S.K.; Leatherwood, C.; Costenbader, K.H.; Rosenberg, L. Relationship of cigarette smoking and alcohol consumption to incidence of systemic lupus erythematosus in a prospective cohort study of black women. Arthritis Care Res. 2019, 71, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Perrone, A.; Giovino, A.; Benny, J.; Martinelli, F. Advanced glycation end products (AGEs): Biochemistry, signaling, analytical methods, and epigenetic effects. Oxid. Med. Cell. Longev. 2020, 2020, 3818196. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Parameter | SLE Patients (n = 31) | Control Group (n = 26) | Statistical Significance |
|---|---|---|---|
| mean age (±SD) (years) | 56.39 (±11.36) | 51.88 (±11.05) | p = 0.14 |
| current or past regular smokers % (n) | 54.84 (17) | 26.92 (7) | p < 0.05 |
| mean time of smoking (±SD) (years) | 23.00 (±11.16) | 16.71 (±10.61) | p = 0.31 |
| passive smokers % (n) | 41.94 (13) | 26.92 (7) | p = 0.24 |
| drinking alcohol % (n) | 61.29 (19) | 84.62 (22) | p = 0.05 |
| mean disease duration (±SD) (years) | 12.61 (±8.49) | NA | NA |
| creatinine (μmol/L) | 69.44 (±19.76) | NA | NA |
| mean GFR (±SD) (mL/min/1.73m2) | 91.17 (±19.86) | NA | NA |
| mean SLEDAI-2K score (±SD) | 11.45 (±7.28) | NA | NA |
| medication for SLE | |||
| receiving any medication or SLE % (n) | 90.32 (28) | NA | NA |
| antimalarial % (n) (chloroquine, hydroxychloroquine) | 22.58 (7) | NA | NA |
| corticosteroids % (n) (methylprednisolone, prednisone) | 35.48 (11) | NA | NA |
| antimetabolites % (n) (azathioprine, methotrexate) | 45.16 (14) | NA | NA |
| manifestations incidence since the time of diagnosis | |||
| rash % (n) | 93.55 (29) | NA | NA |
| photosensitivity % (n) | 80.65 (25) | NA | NA |
| oral ulcers % (n) | 25.81 (8) | NA | NA |
| nonerosive arthritis % (n) | 90.32 (28) | NA | NA |
| pleuritis or pericarditis % (n) | 0 (0) | NA | NA |
| renal disorder % (n) | 6.45 (2) | NA | NA |
| neurologic disorder % (n) | 3.23 (1) | NA | NA |
| hematologic disorder % (n) | 45.16 (14) | NA | NA |
| cardiovascular disorder % (n) | 80.65 (25) | NA | NA |
| immunological disorder /ANA % (n) | 93.55 (29) | NA | NA |
| Mean Concentration (±SD) | SLE Patients (n = 31) | Control Group (n = 26) | Statistical Significance |
|---|---|---|---|
| AGEs (µg/mL) | 30.51 (±6.80) | 24.02 (±8.50) | p < 0.01 |
| CML (µg/mL) | 0.31 (±0.09) | 0.30 (±0.16) | p = 0.65 |
| CEL (µg/mL) | 17.73 (±10.66) | 19.11 (±7.94) | p = 0.35 |
| pentosidine (ng/mL) | 4.54 (±1.84) | 6.71 (±3.360) | p = 0.06 |
| sRAGE (pg/mL) | 36.36 (±15.71) | 47.18 (±19.41) | p < 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowak, A.; Przywara-Chowaniec, B.; Damasiewicz-Bodzek, A.; Blachut, D.; Nowalany-Kozielska, E.; Tyrpień-Golder, K. Advanced Glycation End-Products (AGEs) and Their Soluble Receptor (sRAGE) in Women Suffering from Systemic Lupus Erythematosus (SLE). Cells 2021, 10, 3523. https://doi.org/10.3390/cells10123523
Nowak A, Przywara-Chowaniec B, Damasiewicz-Bodzek A, Blachut D, Nowalany-Kozielska E, Tyrpień-Golder K. Advanced Glycation End-Products (AGEs) and Their Soluble Receptor (sRAGE) in Women Suffering from Systemic Lupus Erythematosus (SLE). Cells. 2021; 10(12):3523. https://doi.org/10.3390/cells10123523
Chicago/Turabian StyleNowak, Agnieszka, Brygida Przywara-Chowaniec, Aleksandra Damasiewicz-Bodzek, Dominika Blachut, Ewa Nowalany-Kozielska, and Krystyna Tyrpień-Golder. 2021. "Advanced Glycation End-Products (AGEs) and Their Soluble Receptor (sRAGE) in Women Suffering from Systemic Lupus Erythematosus (SLE)" Cells 10, no. 12: 3523. https://doi.org/10.3390/cells10123523
APA StyleNowak, A., Przywara-Chowaniec, B., Damasiewicz-Bodzek, A., Blachut, D., Nowalany-Kozielska, E., & Tyrpień-Golder, K. (2021). Advanced Glycation End-Products (AGEs) and Their Soluble Receptor (sRAGE) in Women Suffering from Systemic Lupus Erythematosus (SLE). Cells, 10(12), 3523. https://doi.org/10.3390/cells10123523