
Lamin A/C-Dependent Translocation of Megakaryoblastic Leukemia-1 and β-Catenin in Cyclic Strain-Induced Osteogenesis


 and
and
Abstract
:1. Introduction
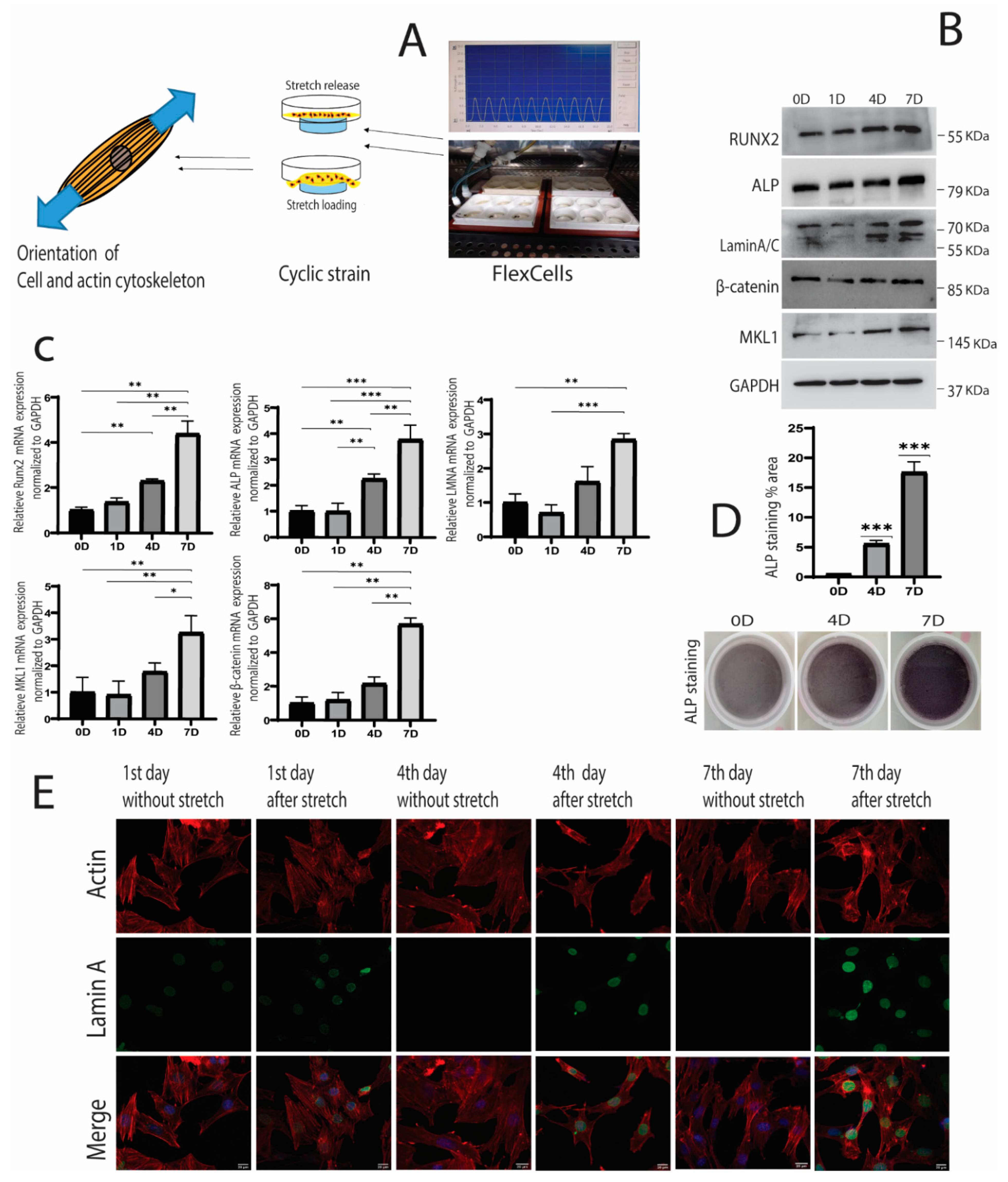
2. Results
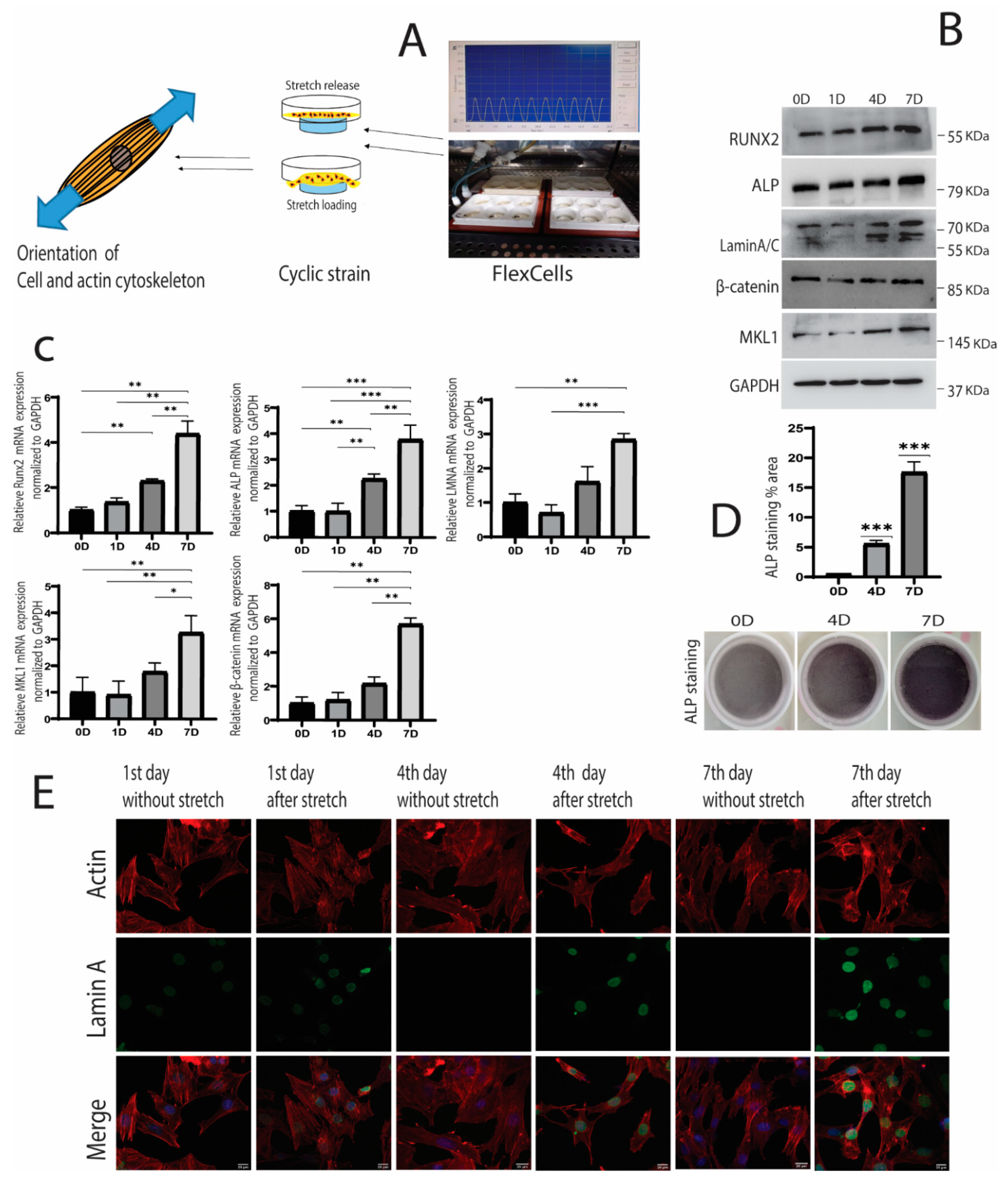
2.1. Cyclic Strain and Osteogenic Differentiation
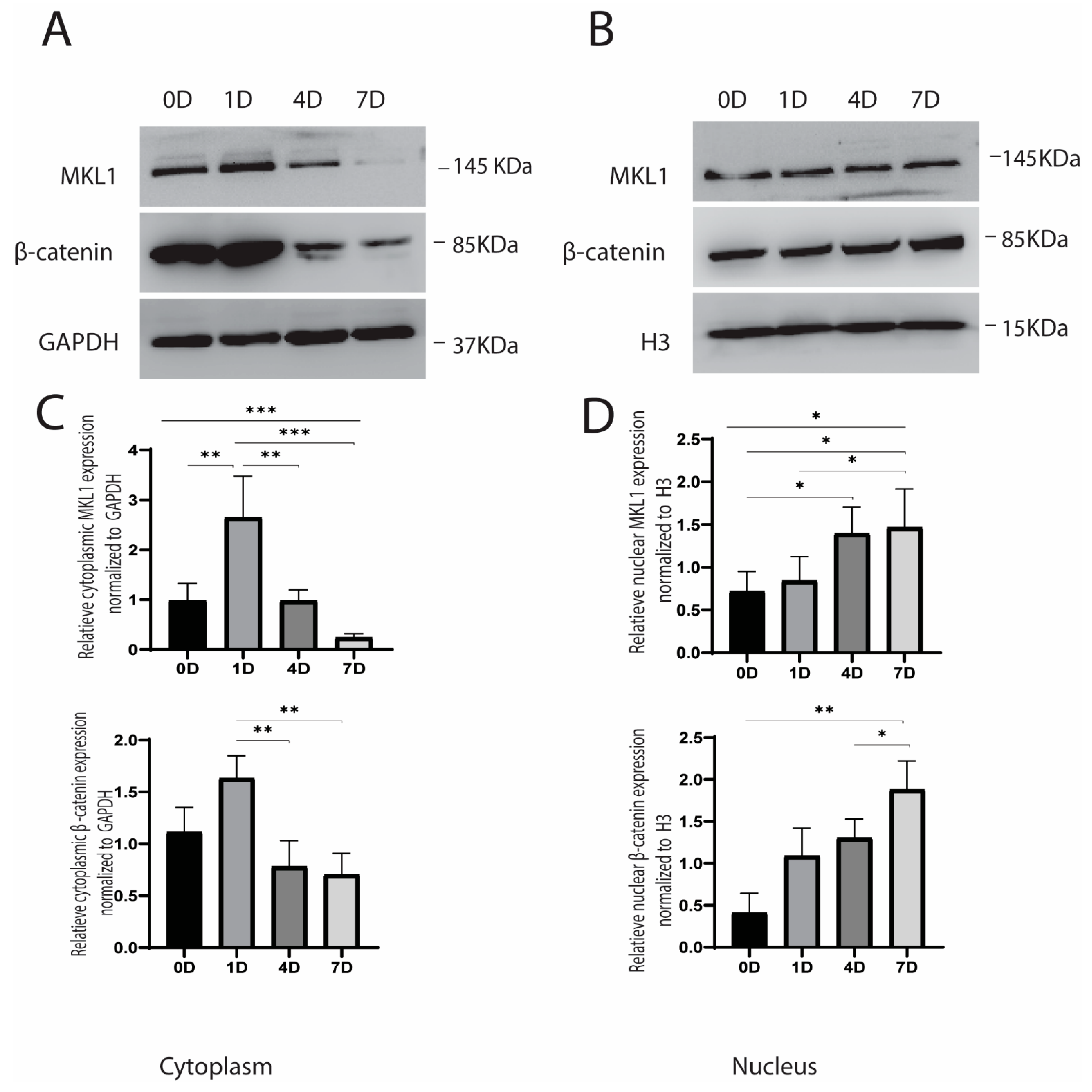
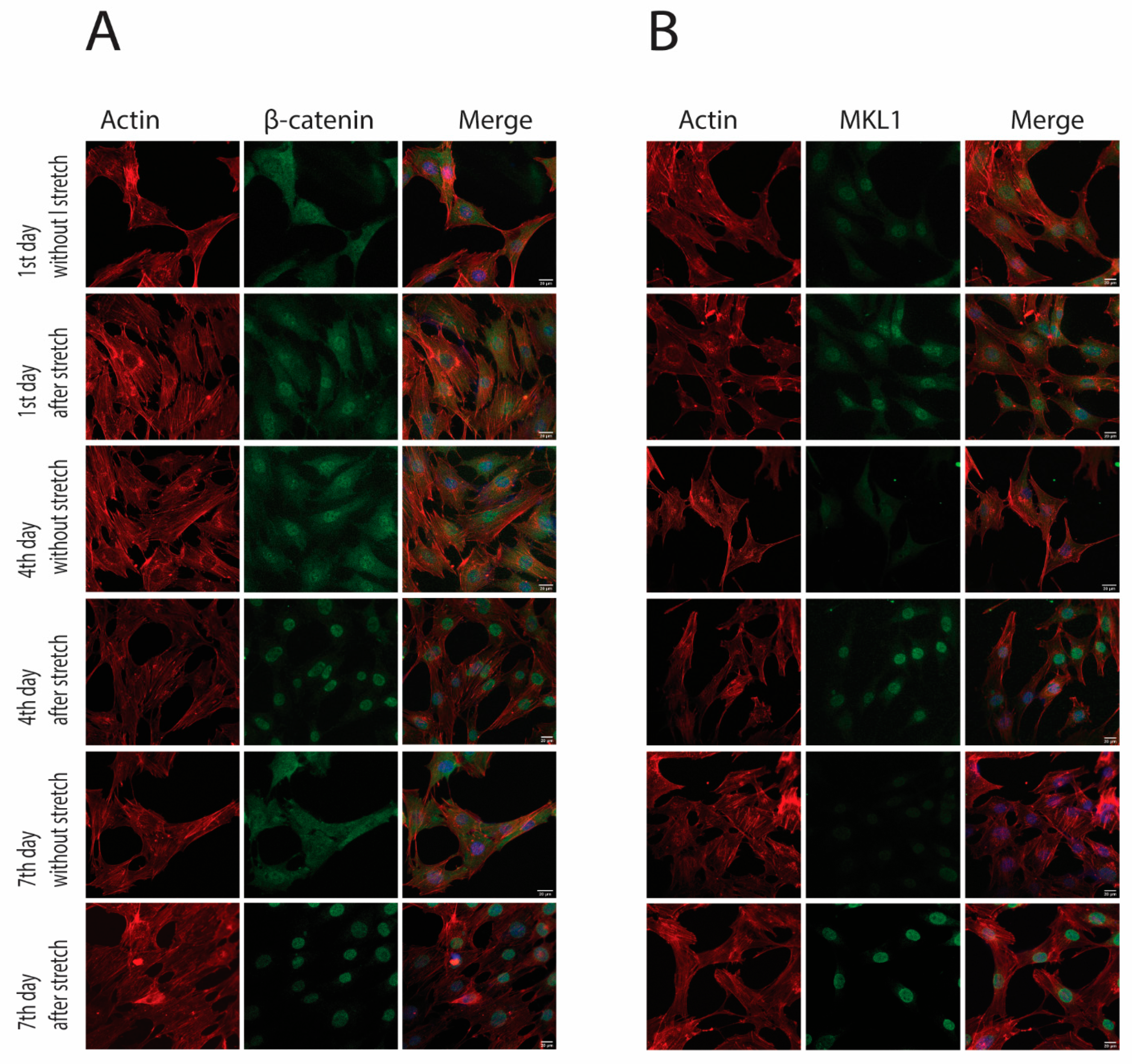
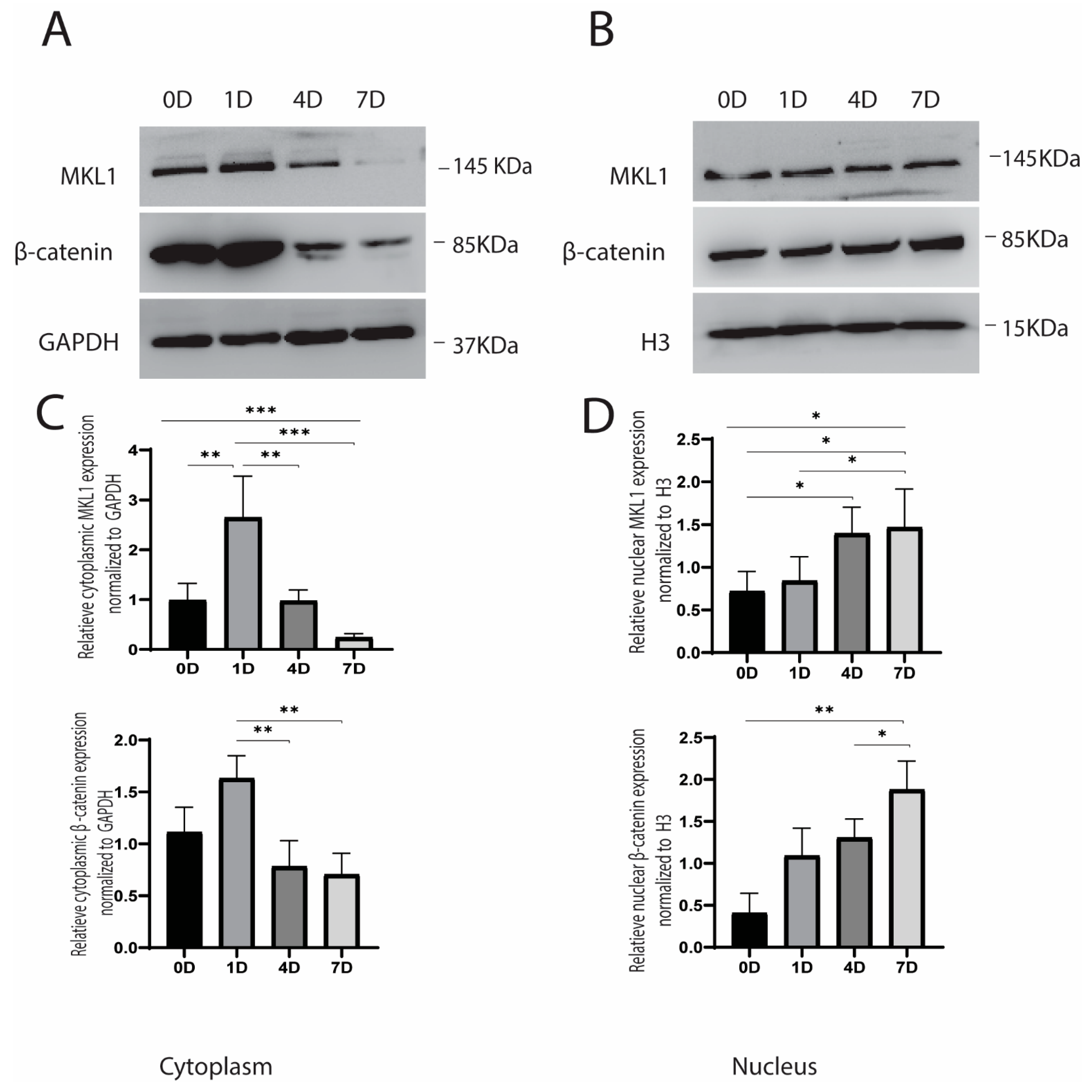
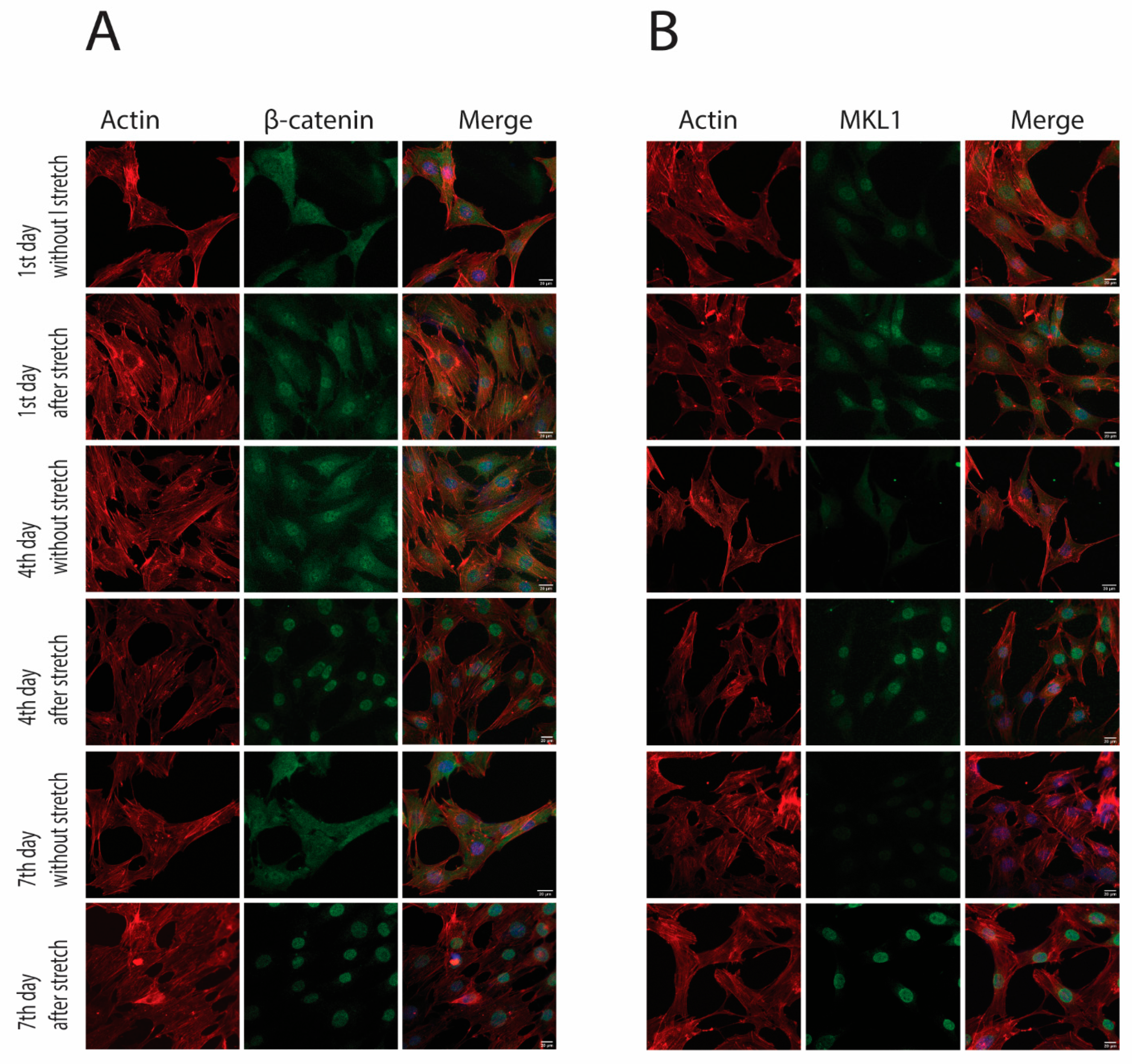
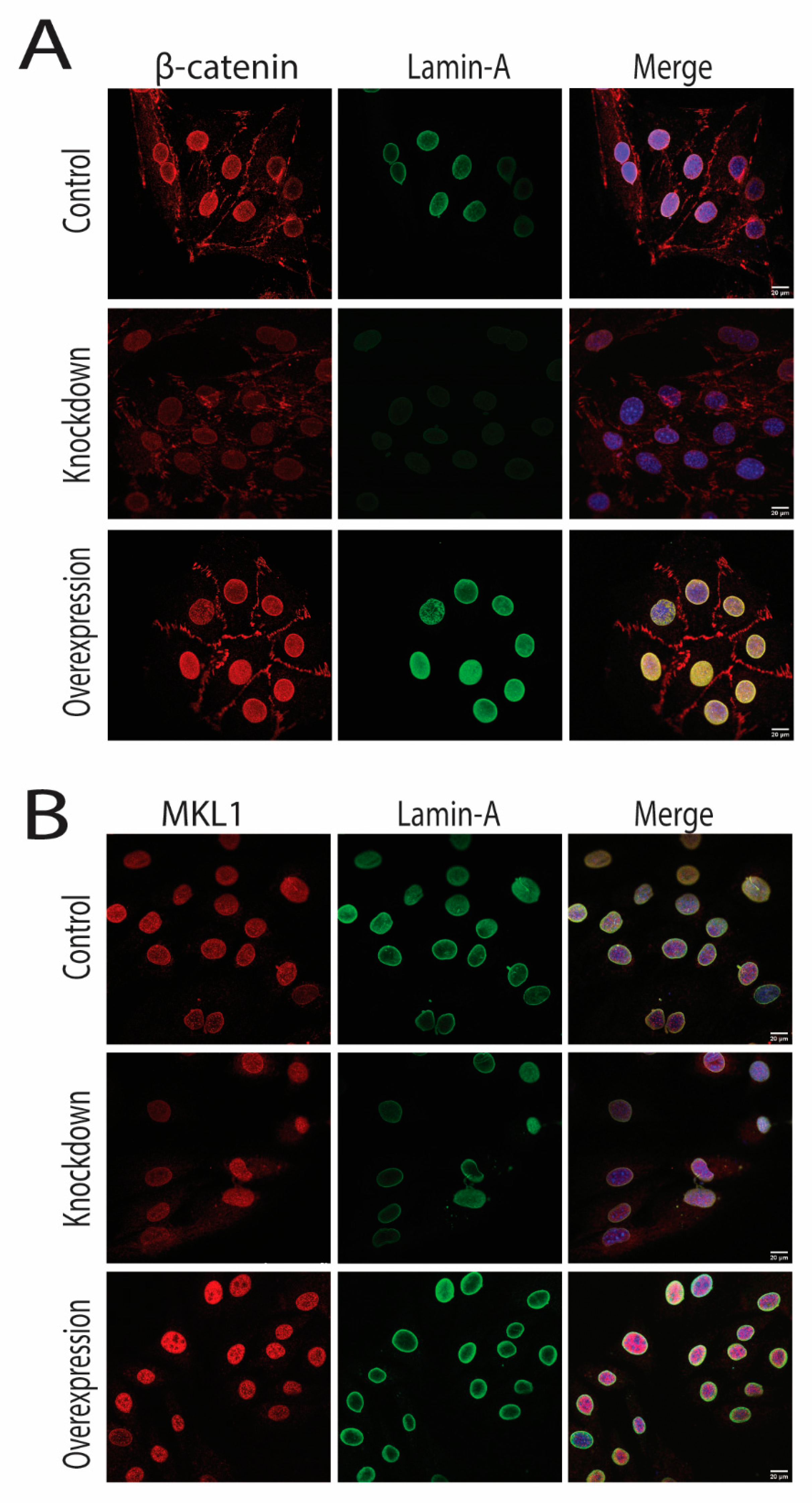
2.2. Cyclic Strain and Translocation of Signaling Molecules
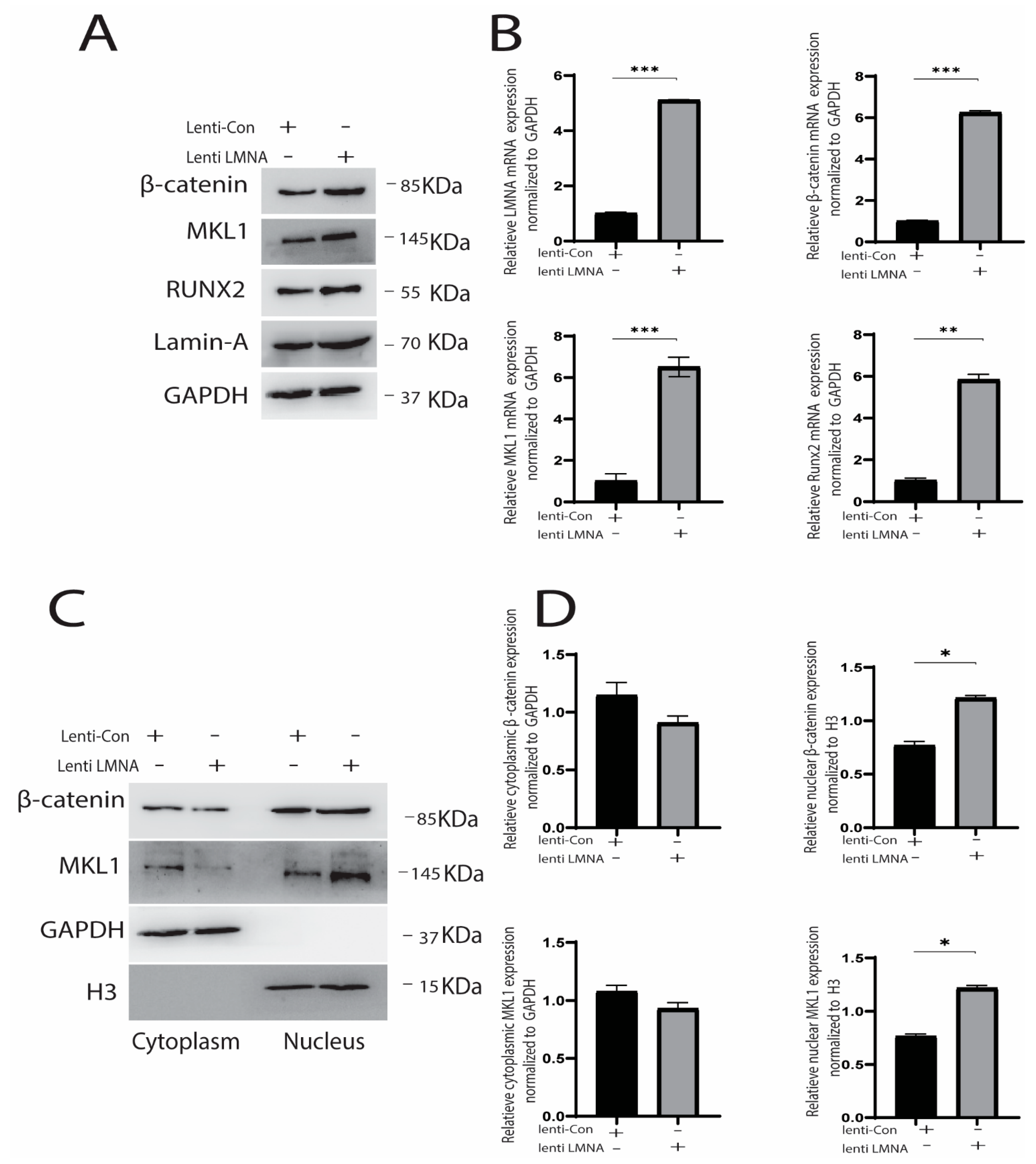
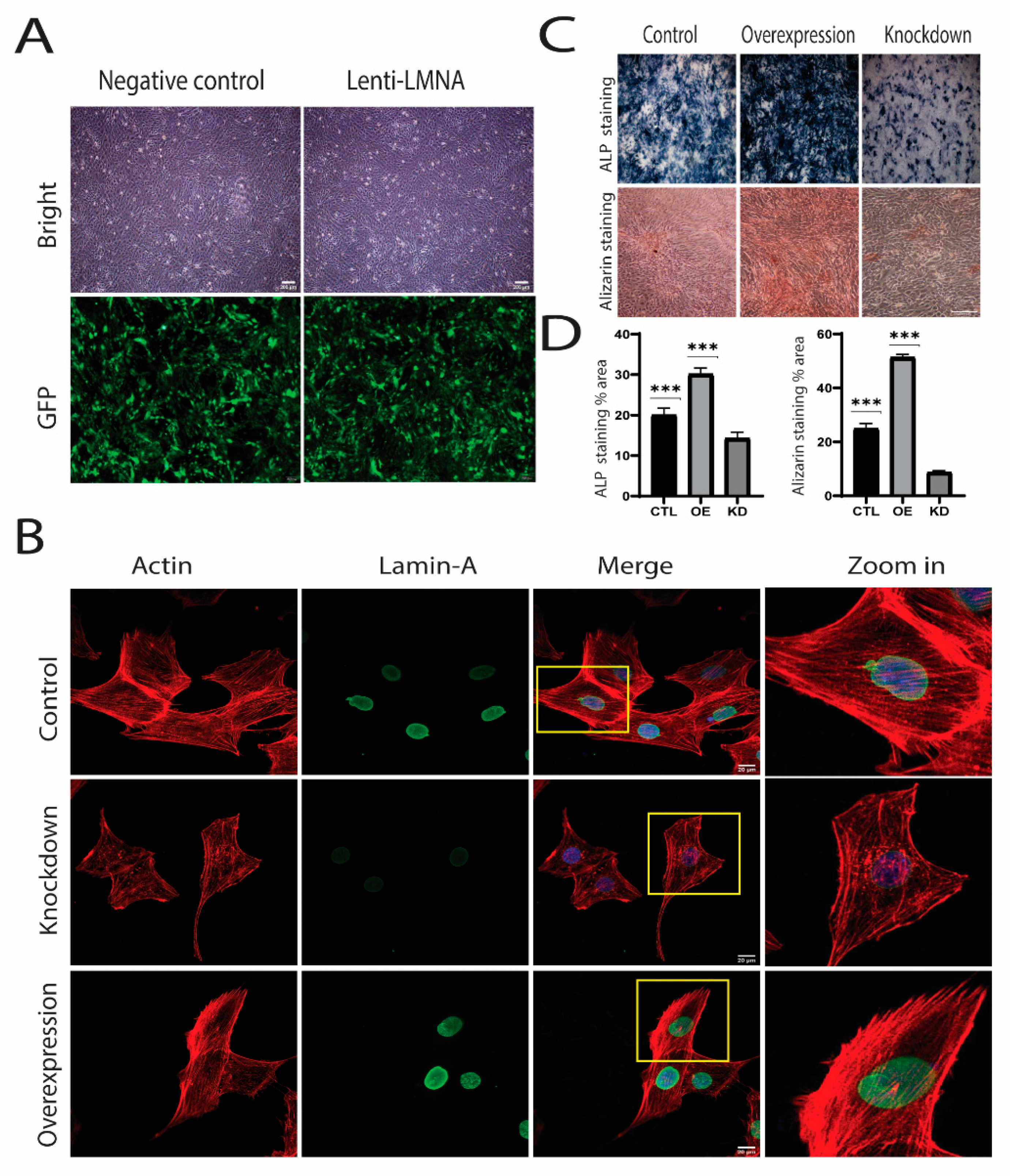
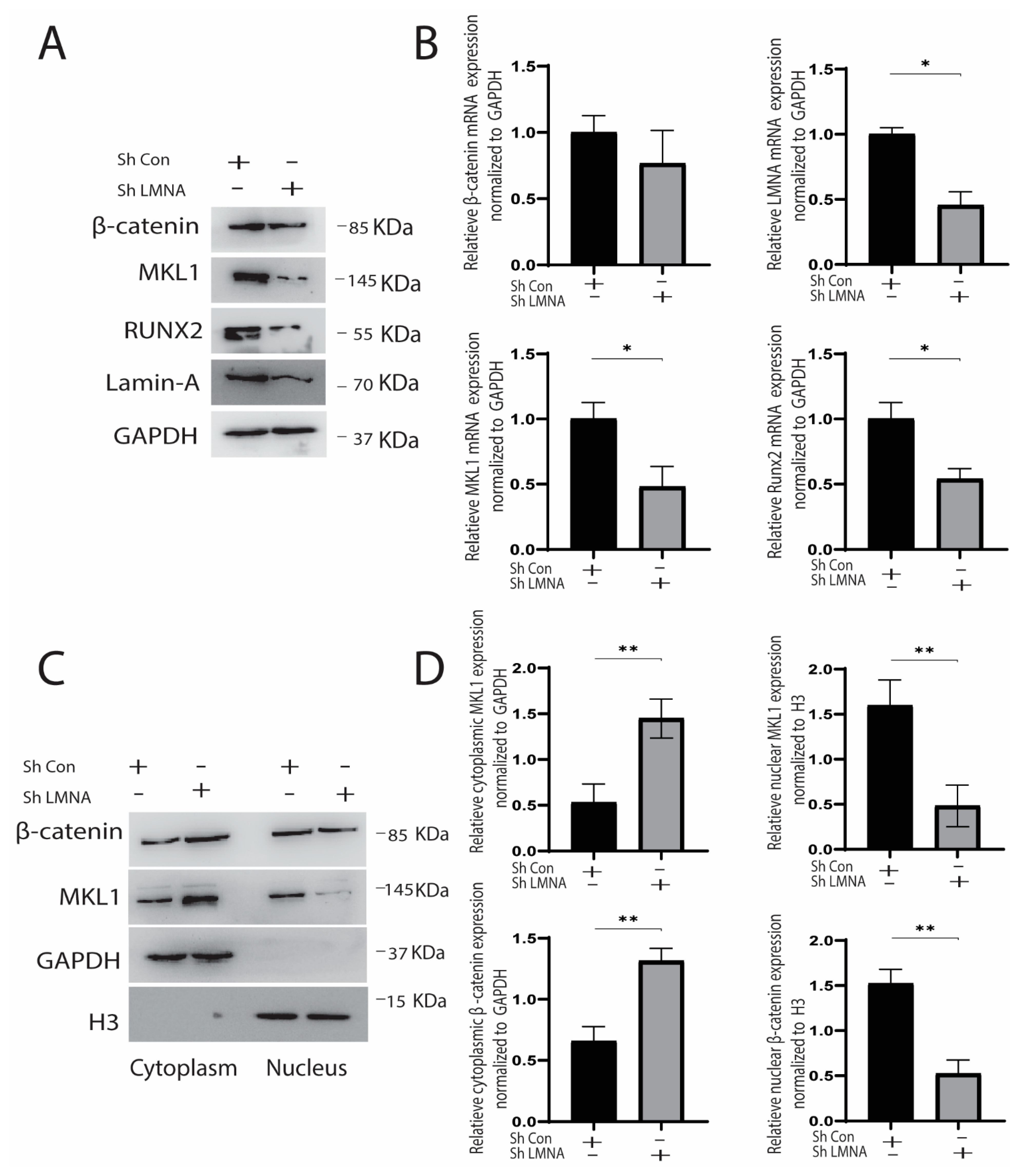
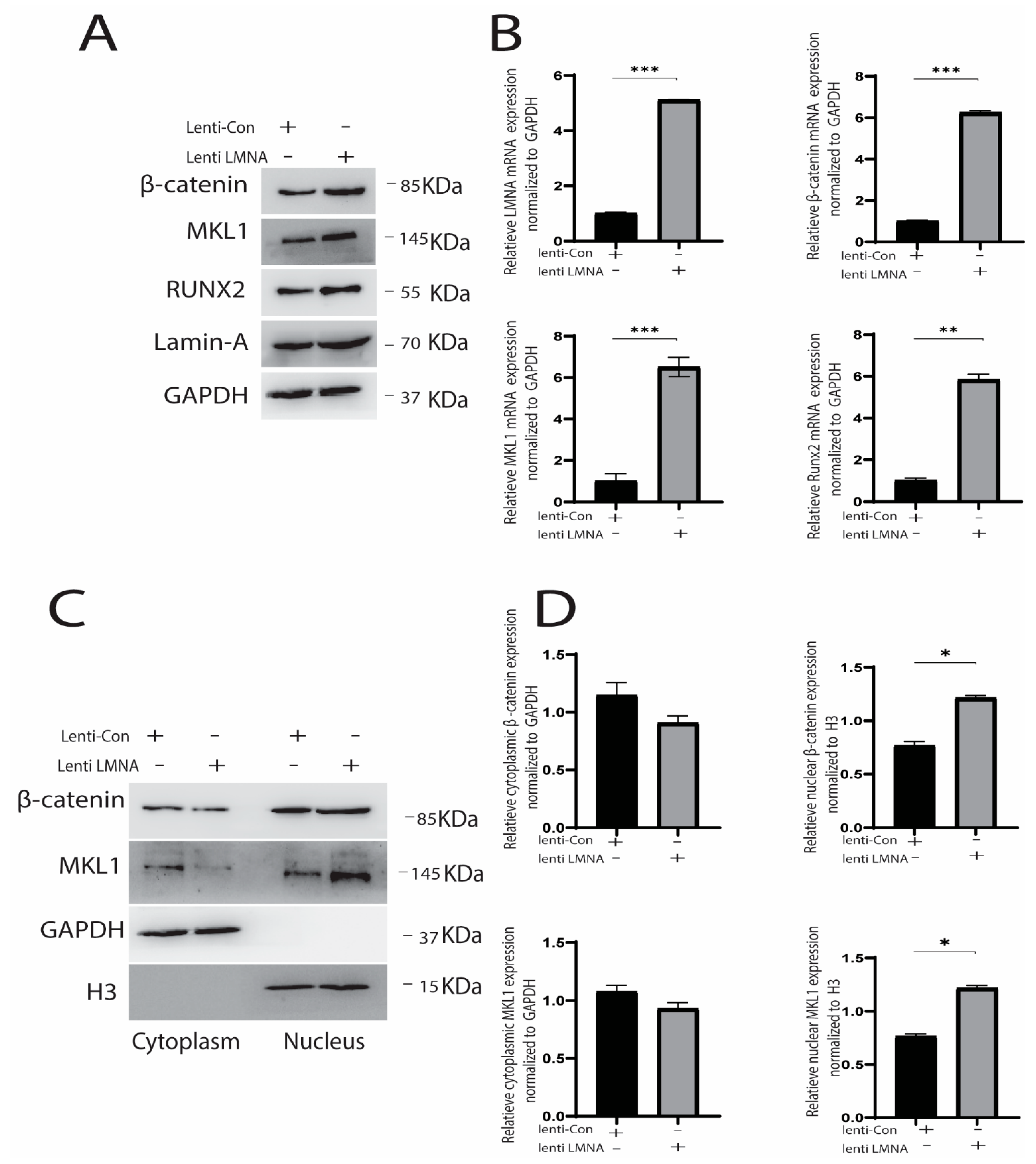
2.3. Knockdown and Overexpression of LMNA
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cyclic Strain Loading
4.3. ALP Staining
4.4. Alizarin Staining
4.5. Immunofluorescence
4.6. Cytoplasmic and Nuclear Protein Extraction
4.7. Western Blotting
4.8. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
4.9. Lentivirus Transduction for Overexpression
4.10. Knockdown LMNA
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heessen, S.; Fornerod, M. The inner nuclear envelope as a transcription factor resting place. EMBO Rep. 2007, 8, 914–919. [Google Scholar] [CrossRef] [Green Version]
- Andres, V.; Gonzalez, J.M. Role of A-type lamins in signaling, transcription, and chromatin organization. J. Cell Biol. 2009, 187, 945–957. [Google Scholar] [CrossRef] [Green Version]
- Worman, H.J. Nuclear lamins and laminopathies. J. Pathol. 2012, 226, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Paro, F.M.; Steiner, A.A.; De Paula, P.M.; Branco, L.G. Central heme oxygenase–carbon monoxide pathway in the control of breathing under normoxia and hypoxia. Respir. Physiol. Neurobiol. 2002, 130, 151–160. [Google Scholar] [CrossRef]
- Scaffidi, P.; Misteli, T. Reversal of the cellular phenotype in the premature aging disease Hutchinson-Gilford progeria syndrome. Nat. Med. 2005, 11, 440–445. [Google Scholar] [CrossRef]
- Hennekam, R.C. Hutchinson–Gilford progeria syndrome: Review of the phenotype. Am. J. Med. Genet. Part A 2006, 140, 2603–2624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novelli, G.; Muchir, A.; Sangiuolo, F.; Helbling-Leclerc, A.; D’Apice, M.R.; Massart, C.; Capon, F.; Sbraccia, P.; Federici, M.; Lauro, R.; et al. Mandibuloacral dysplasia is caused by a mutation in LMNA-encoding lamin A/C. Am. J. Hum. Genet. 2002, 71, 426–431. [Google Scholar] [CrossRef] [Green Version]
- Hishiya, A.K. Watanabe, Progeroid syndrome as a model for impaired bone formation in senile osteoporosis. J. Bone Miner. Metab. 2004, 22, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Carrero, D.; Soria-Valles, C.; Lopez-Otin, C. Hallmarks of progeroid syndromes: Lessons from mice and reprogrammed cells. Dis. Model. Mech. 2016, 9, 719–735. [Google Scholar] [CrossRef] [Green Version]
- Ho, C.Y.; Jaalouk, D.E.; Vartiainen, M.K.; Lammerding, J. Lamin A/C and emerin regulate MKL1–SRF activity by modulating actin dynamics. Nature 2013, 497, 507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcorta-Sevillano, N.; Macías, I.; Rodríguez, C.I.; Infante, A. Crucial role of Lamin A/C in the migration and differentiation of MSCs in bone. Cells 2020, 9, 1330. [Google Scholar] [CrossRef]
- Malashicheva, A.; Perepelina, K. Diversity of Nuclear Lamin A/C Action as a Key to Tissue-Specific Regulation of Cellular Identity in Health and Disease. Front. Cell Dev. Biol. 2021, 9, 2834. [Google Scholar] [CrossRef]
- Akter, R.; Rivas, D.; Geneau, G.; Drissi, H.; Duque, G. Effect of lamin A/C knockdown on osteoblast differentiation and function. J. Bone Miner. Res. 2009, 24, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Rauner, M.; Sipos, W.; Goettsch, C.; Wutzl, A.; Foisner, R.; Pietschmann, P.; Hofbauer, L.C. Inhibition of lamin A/C attenuates osteoblast differentiation and enhances RANKL-dependent osteoclastogenesis. J. Bone Miner. Res. 2009, 24, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Scaffidi, P.; Misteli, T. Lamin A-dependent misregulation of adult stem cells associated with accelerated ageing. Nat. Cell Biol. 2008, 10, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Swift, J.; Ivanovska, I.L.; Buxboim, A.; Harada, T.; Dingal, P.D.P.; Pinter, J.; Pajerowski, J.D.; Spinler, K.R.; Shin, J.-W.; Tewari, M. Nuclear lamin-A scales with tissue stiffness and enhances matrix-directed differentiation. Science 2013, 341, 6149. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Wan, J.; Wang, H. Mechanical strain triggers differentiation of dental mesenchymal stem cells by activating osteogenesis-specific biomarkers expression. Am. J. Transl. Res. 2019, 11, 233. [Google Scholar]
- Li, R.; Liang, L.; Dou, Y.; Huang, Z.; Mo, H.; Wang, Y.; Yu, B. Mechanical strain regulates osteogenic and adipogenic differentiation of bone marrow mesenchymal stem cells. BioMed Res. Int. 2015, 2015, 873251. [Google Scholar] [CrossRef] [PubMed]
- Prè, D.; Ceccarelli, G.; Visai, L.; Benedetti, L.; Imbriani, M.; De Angelis, M.C.; Magenes, G. High-frequency vibration treatment of human bone marrow stromal cells increases differentiation toward bone tissue. Bone Marrow Res. 2013, 2013, 803450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pre, D.; Ceccarelli, G.; Gastaldi, G.; Asti, A.; Saino, E.; Visai, L.; Benazzo, F.; De Angelis, M.C.; Magenes, G. The differentiation of human adipose-derived stem cells (hASCs) into osteoblasts is promoted by low amplitude, high frequency vibration treatment. Bone 2011, 49, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Simmons, C.A.; Matlis, S.; Thornton, A.J.; Chen, S.; Wang, C.-Y.; Mooney, D.J. Cyclic strain enhances matrix mineralization by adult human mesenchymal stem cells via the extracellular signal-regulated kinase (ERK1/2) signaling pathway. J. Biomech. 2003, 36, 1087–1096. [Google Scholar] [CrossRef]
- Luo, Y.; Ge, R.; Wu, H.; Ding, X.; Song, H.; Ji, H.; Li, M.; Ma, Y.; Li, S.; Wang, C. The osteogenic differentiation of human adipose-derived stem cells is regulated through the let-7i-3p/LEF1/β-catenin axis under cyclic strain. Stem Cell Res. Ther. 2019, 10, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Charoenpanich, A.; Wall, M.E.; Tucker, C.J.; Andrews, D.M.; Lalush, D.S.; Loboa, E.G. Microarray analysis of human adipose-derived stem cells in three-dimensional collagen culture: Osteogenesis inhibits bone morphogenic protein and Wnt signaling pathways, and cyclic tensile strain causes upregulation of proinflammatory cytokine regulators and angiogenic factors. Tissue Eng. Part A 2011, 17, 2615–2627. [Google Scholar]
- Charoenpanich, A.; Wall, M.E.; Tucker, C.J.; Andrews, D.M.; Lalush, D.S.; Dirschl, D.R.; Loboa, E.G. Cyclic tensile strain enhances osteogenesis and angiogenesis in mesenchymal stem cells from osteoporotic donors. Tissue Eng Part A 2014, 20, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Qi, M.c.; Zou, S.j.; Han, L.c.; Zhou, H.x.; Hu, J. Expression of bone-related genes in bone marrow MSCs after cyclic mechanical strain: Implications for distraction osteogenesis. Int. J. Oral Sci. 2009, 1, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Sumanasinghe, R.D.; Bernacki, S.H.; Loboa, E.G. Osteogenic differentiation of human mesenchymal stem cells in collagen matrices: Effect of uniaxial cyclic tensile strain on bone morphogenetic protein (BMP-2) mRNA expression. Tissue Eng. 2006, 12, 3459–3465. [Google Scholar] [CrossRef] [PubMed]
- Lohberger, B.; Kaltenegger, H.; Stuendl, N.; Payer, M.; Rinner, B.; Leithner, A. Effect of cyclic mechanical stimulation on the expression of osteogenesis genes in human intraoral mesenchymal stromal and progenitor cells. BioMed Res. Int. 2014, 2014, 189516. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, X.; Meikle, M.C.; Islam, I.; Cao, T. Short periods of cyclic mechanical strain enhance triple-supplement directed osteogenesis and bone nodule formation by human embryonic stem cells in vitro. Tissue Eng. Part A 2013, 19, 2130–2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Qu, R.; Feng, Y.; Huang, X.; Yang, Y.; Fan, T.; Sun, B.; Khan, A.U.; Wu, S.; Dai, J. Regulation of the integrin αVβ3-actin filaments axis in early osteogenesis of human fibroblasts under cyclic tensile stress. Stem Cell Res. Ther. 2021, 12, 1–12. [Google Scholar] [CrossRef]
- Zeng, Z.; Yin, X.; Zhang, X.; Jing, D.; Feng, X. Cyclic stretch enhances bone morphogenetic protein-2-induced osteoblastic differentiation through the inhibition of Hey1. Int. J. Mol. Med. 2015, 36, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Huang, L.; Hao, B.; Li, H.; Zhu, S.; Wang, Q.; Li, R.; Xu, Y.; Zhang, X. Use of an osteoblast overload damage model to probe the effect of icariin on the proliferation, differentiation and mineralization of MC3T3-E1 cells through the Wnt/β-catenin signalling pathway. Cell. Physiol. Biochem. 2017, 41, 1605–1615. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, J.I.; Plotkin, L.I.; Stewart, S.A.; Weinstein, R.S.; Parfitt, A.M.; Manolagas, S.C.; Bellido, T. Osteocyte apoptosis is induced by weightlessness in mice and precedes osteoclast recruitment and bone loss. J. Bone Miner. Res. 2006, 21, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Nordström, A.; Olsson, T.; Nordström, P. Sustained benefits from previous physical activity on bone mineral density in males. J. Clin. Endocrinol. Metab. 2006, 91, 2600–2604. [Google Scholar] [CrossRef]
- Klein-Nulend, J.; Bacabac, R.G.; Bakker, A.D. Mechanical Loading and How It Affects Bone Cells: The Role of the Osteocyte Cytoskeleton in Maintaining Our Skeleton. Eur. Cells Mater. 2012, 24, 278–291. [Google Scholar] [CrossRef] [PubMed]
- Buxboim, A.; Buxboim, A.; Swift, J.; Irianto, J.; Spinler, K.R.; Dingal, P.C.; Athirasala, A.; Kao, Y.R.; Cho, S.; Harada, T.; et al. Matrix elasticity regulates lamin-A,C phosphorylation and turnover with feedback to actomyosin. Curr. Biol. 2014, 24, 1909–1917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dingal, P.D.P.; Discher, D.E. Systems mechanobiology: Tension-inhibited protein turnover is sufficient to physically control gene circuits. Biophys. J. 2014, 107, 2734–2743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Liu, X.; Wang, J.; Chen, X.; Zhang, H.; Kim, S.H.; Cui, J.; Li, R.; Zhang, W.; Kong, Y.; et al. Wnt signaling in bone formation and its therapeutic potential for bone diseases. Ther. Adv. Musculoskelet. Dis. 2013, 5, 13–31. [Google Scholar] [CrossRef] [Green Version]
- Torre, A.G.; Shaw, J.E.; Wood, A.; Gilbert, H.T.; Dobre, O.; Genever, P.; Brennan, K.; Richardson, S.M.; Swift, J. An immortalised mesenchymal stem cell line maintains mechano-responsive behaviour and can be used as a reporter of substrate stiffness. Sci. Rep. 2018, 8, 8981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.U.; Qu, R.; Fan, T.; Ouyang, J.; Dai, J. A glance on the role of actin in osteogenic and adipogenic differentiation of mesenchymal stem cells. Stem Cell Res. Ther. 2020, 11, 283. [Google Scholar] [CrossRef]
- Brock, R. The uptake of arginine-rich cell-penetrating peptides: Putting the puzzle together. Bioconjug. Chem. 2014, 25, 863–868. [Google Scholar] [CrossRef]
- Valenta, T.; Hausmann, G.; Basler, K. The many faces and functions of β-catenin. EMBO J. 2012, 31, 2714–2736. [Google Scholar] [CrossRef] [Green Version]
- Gaur, T.; Lengner, C.J.; Hovhannisyan, H.; Bhat, R.A.; Bodine, P.V.; Komm, B.S.; Javed, A.; Van Wijnen, A.J.; Stein, J.L.; Stein, G.S. Canonical WNT signaling promotes osteogenesis by directly stimulating Runx2 gene expression. J. Biol. Chem. 2005, 280, 33132–33140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakar, K.; Carroll, C.W. Mkl1-dependent gene activation is sufficient to induce actin cap assembly. Small GTPases 2019, 10, 433–440. [Google Scholar] [CrossRef]
- Sankaran, J.; Uzer, G.; van Wijnen, A.J.; Rubin, J. Gene regulation through dynamic actin control of nuclear structure. Exp. Biol. Med. 2019, 244, 1345–1353. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Ren, X.; Oberoi, M.K.; Bedar, M.; Caprini, R.M.; Dewey, M.J.; Kolliopoulos, V.; Yamaguchi, D.T.; Harley, B.A.; Lee, J.C. β-Catenin Limits Osteogenesis on Regenerative Materials in a Stiffness-Dependent Manner. Adv. Healthc. Mater. 2021, 20, 2101467. [Google Scholar] [CrossRef] [PubMed]
- Bermeo, S.; Vidal, C.; Zhou, H.; Duque, G. Lamin A/C Acts as an Essential Factor in Mesenchymal Stem Cell Differentiation Through the Regulation of the Dynamics of the Wnt/β-Catenin Pathway. J. Cell. Biochem. 2015, 116, 2344–2353. [Google Scholar] [CrossRef] [PubMed]
- Mendez, M.G.; Janmey, P.A. Transcription factor regulation by mechanical stress. Int. J. Biochem. Cell Biol. 2012, 44, 728–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boeri, L.; Albani, D.; Raimondi, M.T.; Jacchetti, E. Mechanical regulation of nucleocytoplasmic translocation in mesenchymal stem cells: Characterization and methods for investigation. Biophys. Rev. 2019, 11, 817–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Liu, Z.Z.; Chen, X.; Schulz, V.P.; Kumar, A.; Hartman, A.A.; Weinstein, J.; Johnston, J.F.; Rodriguez, E.C.; Eastman, A.E. MKL1-actin pathway restricts chromatin accessibility and prevents mature pluripotency activation. Nat. Commun. 2019, 10, 1695. [Google Scholar] [CrossRef] [PubMed]
- Donnaloja, F.; Carnevali, F.; Jacchetti, E.; Raimondi, M.T. Lamin A/C Mechanotransduction in laminopathies. Cells 2020, 9, 1306. [Google Scholar] [CrossRef]
- Cho, S.; Vashisth, M.; Abbas, A.; Majkut, S.; Vogel, K.; Xia, Y.; Ivanovska, I.L.; Irianto, J.; Tewari, M.; Zhu, K. Mechanosensing by the lamina protects against nuclear rupture, DNA damage, and cell-cycle arrest. Dev. Cell 2019, 49, 920–935.e5. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.S.; Lee, S.J.; Lee, H.S.; Moon, W.; Cho, D.W. Effects of combined mechanical stimulation on the proliferation and differentiation of pre-osteoblasts. Exp. Mol. Med. 2011, 43, 367–373. [Google Scholar] [CrossRef]
- Kearney, E.; Farrell, E.; Prendergast, P.; Campbell, V. Tensile strain as a regulator of mesenchymal stem cell osteogenesis. Ann. Biomed. Eng. 2010, 38, 1767–1779. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Cai, X.; Wang, J.; Tang, H.; Yuan, Q.; Gong, P.; Lin, Y. Mechanical stretch inhibits adipogenesis and stimulates osteogenesis of adipose stem cells. Cell Prolif. 2012, 45, 158–166. [Google Scholar] [CrossRef]
- Miralles, F.; Posern, G.; Zaromytidou, A.-I.; Treisman, R. Actin dynamics control SRF activity by regulation of its coactivator MAL. Cell 2003, 113, 329–342. [Google Scholar] [CrossRef] [Green Version]
- Dupont, S.; Morsut, L.; Aragona, M.; Enzo, E.; Giulitti, S.; Cordenonsi, M.; Zanconato, F.; Le Digabel, J.; Forcato, M.; Bicciato, S.; et al. Role of YAP/TAZ in mechanotransduction. Nature 2011, 474, 179–183. [Google Scholar] [CrossRef]
- Premaraj, S.; Souza, I.; Premaraj, T. Mechanical loading activates β-catenin signaling in periodontal ligament cells. Angle Orthod. 2011, 81, 592–599. [Google Scholar] [CrossRef]
- Benham-Pyle, B.W.; Sim, J.Y.; Hart, K.C.; Pruitt, B.L.; Nelson, W.J. Increasing β-catenin/Wnt3A activity levels drive mechanical strain-induced cell cycle progression through mitosis. Elife 2016, 5, e19799. [Google Scholar] [CrossRef] [Green Version]
- Norvell, S.; Alvarez, M.; Bidwell, J.; Pavalko, F. Fluid shear stress induces β-catenin signaling in osteoblasts. Calcif. Tissue Int. 2004, 75, 396–404. [Google Scholar] [CrossRef]
- Vartiainen, M.K.; Guettler, S.; Larijani, B.; Treisman, R. Nuclear actin regulates dynamic subcellular localization and activity of the SRF cofactor MAL. Science 2007, 316, 1749–1752. [Google Scholar] [CrossRef] [Green Version]
- Chaudhuri, S.; Smith, P.G. Cyclic strain–induced HSP27 phosphorylation modulates actin filaments in airway smooth muscle cells. Am. J. Respir. Cell Mol. Biol. 2008, 39, 270–278. [Google Scholar] [CrossRef]
- Smith, E.C.; Teixeira, A.M.; Chen, R.C.; Wang, L.; Gao, Y.; Hahn, K.L.; Krause, D.S. Induction of megakaryocyte differentiation drives nuclear accumulation and transcriptional function of MKL1 via actin polymerization and RhoA activation. Blood J. Am. Soc. Hematol. 2013, 121, 1094–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobusue, H.; Onishi, N.; Shimizu, T.; Sugihara, E.; Oki, Y.; Sumikawa, Y.; Chiyoda, T.; Akashi, K.; Saya, H.; Kano, K. Regulation of MKL1 via actin cytoskeleton dynamics drives adipocyte differentiation. Nat. Commun. 2014, 5, 3368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masszi, A.; Speight, P.; Charbonney, E.; Lodyga, M.; Nakano, H.; Szászi, K.; Kapus, A. Fate-determining mechanisms in epithelial–myofibroblast transition: Major inhibitory role for Smad3. J. Cell Biol. 2010, 188, 383–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charbonney, E.; Speight, P.; Masszi, A.; Nakano, H.; Kapus, A. β-Catenin and Smad3 regulate the activity and stability of myocardin-related transcription factor during epithelial–myofibroblast transition. Mol. Biol. Cell 2011, 22, 4472–4485. [Google Scholar] [CrossRef] [PubMed]
- Corne, T.D.; Sieprath, T.; Vandenbussche, J.; Mohammed, D.; Te Lindert, M.; Gevaert, K.; Gabriele, S.; Wolf, K.; De Vos, W.H. Deregulation of focal adhesion formation and cytoskeletal tension due to loss of A-type lamins. Cell Adhes. Migr. 2017, 11, 447–463. [Google Scholar] [CrossRef] [Green Version]
- Busch, A.; Kiel, T.; Heupel, W.-M.; Wehnert, M.; Hübner, S. Nuclear protein import is reduced in cells expressing nuclear envelopathy-causing lamin A mutants. Exp. Cell Res. 2009, 315, 2373–2385. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5→3) | Reverse Primer (3→5) |
|---|---|---|
| GAPDH | CAATGTGTCCGTCGTGGATCT | GTCCTCAGTGTAGCCCAAGATG |
| LMNA | CCTTCGCACCGCTCTCATCAAC | TCTTCTCCATCCTCGTCGTCATCC |
| RUNX2 | TCCCGTCACCTCCATCCTCTTTC | GAATACGCATCACAACAGCCACAAG |
| ALP | CTTGGTGGTCACAGCAGTTGGTAG | CCAGGCGACAGGTGAAGAAACAG |
| MKL1 | GTGCTGCGTCCTGCTGTCTAAG | GCTCCTCAATCTGCTTGTCCTTCTC |
| β-catenin | GCTGCTGTCCTATTCCGAATGTCTG | GGCACCAATGTCCAGTCCAAGATC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, A.U.; Qu, R.; Yang, Y.; Fan, T.; Peng, Y.; Sun, B.; Qiu, X.; Wu, S.; Wang, Z.; Zhou, Z.; et al. Lamin A/C-Dependent Translocation of Megakaryoblastic Leukemia-1 and β-Catenin in Cyclic Strain-Induced Osteogenesis. Cells 2021, 10, 3518. https://doi.org/10.3390/cells10123518
Khan AU, Qu R, Yang Y, Fan T, Peng Y, Sun B, Qiu X, Wu S, Wang Z, Zhou Z, et al. Lamin A/C-Dependent Translocation of Megakaryoblastic Leukemia-1 and β-Catenin in Cyclic Strain-Induced Osteogenesis. Cells. 2021; 10(12):3518. https://doi.org/10.3390/cells10123518
Chicago/Turabian StyleKhan, Asmat Ullah, Rongmei Qu, Yuchao Yang, Tingyu Fan, Yan Peng, Bing Sun, Xianshuai Qiu, Shutong Wu, Zetong Wang, Zhitao Zhou, and et al. 2021. "Lamin A/C-Dependent Translocation of Megakaryoblastic Leukemia-1 and β-Catenin in Cyclic Strain-Induced Osteogenesis" Cells 10, no. 12: 3518. https://doi.org/10.3390/cells10123518
APA StyleKhan, A. U., Qu, R., Yang, Y., Fan, T., Peng, Y., Sun, B., Qiu, X., Wu, S., Wang, Z., Zhou, Z., Khan, M. A., Dai, J., & Ouyang, J. (2021). Lamin A/C-Dependent Translocation of Megakaryoblastic Leukemia-1 and β-Catenin in Cyclic Strain-Induced Osteogenesis. Cells, 10(12), 3518. https://doi.org/10.3390/cells10123518