
Association of FOXO3 Blood DNA Methylation with Cancer Risk, Cancer Survival, and Mortality

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Samples
2.2. Methylation Case-Control Studies
2.3. Methylation Longitudinal Study
2.4. Quality Control of DNA Methylation Data
2.5. Genetic Data
2.6. Vital Status
2.7. Statistical Analysis
2.7.1. Effect of FOXO3 SNPs on Methylation
2.7.2. Cancer Risk
2.7.3. Cancer Survival
2.7.4. Overall and Cause-Specific Mortality
3. Results
3.1. Associations with Cancer Risk
3.2. Associations with Cancer Survival
3.3. Associations with Overall Mortality
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kenyon, C.; Chang, J.; Gensch, E.; Rudner, A.; Tabtiang, R. A C. elegans mutant that lives twice as long as wild type. Nature 1993, 366, 461–464. [Google Scholar] [CrossRef]
- Murakami, S.; Johnson, T.E. A genetic pathway conferring life extension and resistance to UV stress in Caenorhabditis Elegans. Genetics 1996, 143, 1207–1218. [Google Scholar] [CrossRef]
- Lin, K.; Dorman, J.B.; Rodan, A.; Kenyon, C. Daf-16: An HNF-3/fork head family member that can function to double the life-span of Caenorhabditis elegans. Science 1997, 278, 1319–1322. [Google Scholar] [CrossRef] [Green Version]
- Ogg, S.; Paradis, S.; Gottlieb, S.; Patterson, G.I.; Lee, L.; Tissenbaum, H.A.; Ruvkun, G. The fork head transcription factor DAF-16 transduces insulin-like metabolic and longevity signals in C. elegans. Nature 1997, 389, 994–999. [Google Scholar] [CrossRef]
- Willcox, B.J.; Donlon, T.A.; He, Q.; Chen, R.; Grove, J.S.; Yano, K.; Masaki, K.H.; Willcox, D.C.; Rodriguez, B.; Curb, J.D. FOXO3A genotype is strongly associated with human longevity. Proc. Nat. Acad. Sci. USA 2008, 105, 13987–13992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anselmi, C.V.; Malovini, A.; Roncarati, R.; Novelli, V.; Villa, F.; Condorelli, G.; Bellazzi, R.; Puca, A.A. Association of the FOXO3A locus with extreme longevity in a southern Italian centenarian study. Rejuvenat. Res. 2009, 12, 95–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flachsbart, F.; Caliebe, A.; Kleindorp, R.; Blanché, H.; von Eller-Eberstein, H.; Nikolaus, S.; Schreiber, S.; Nebel, A. Association of FOXO3A variation with human longevity confirmed in German centenarians. Proc. Nat. Acad. Sci. USA 2009, 106, 2700–2705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wang, W.J.; Cao, H.; Lu, J.; Wu, C.; Hu, F.Y.; Guo, J.; Zhao, L.; Yang, F.; Zhang, Y.X.; et al. Genetic association of FOXO1A and FOXO3A with longevity trait in Han Chinese populations. Hum. Mol. Genet. 2009, 18, 4897–4904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawlikowska, L.; Hu, D.; Huntsman, S.; Sung, A.; Chu, C.; Chen, J.; Joyner, A.H.; Schork, N.J.; Hsueh, W.C.; Reiner, A.P.; et al. Association of common genetic variation in the insulin/IGF1 signaling pathway with human longevity. Aging Cell 2009, 8, 460–472. [Google Scholar] [CrossRef] [Green Version]
- Kenyon, C.J. The genetics of ageing. Nature 2010, 464, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Sanese, P.; Forte, G.; Disciglio, V.; Grossi, V.; Simone, C. FOXO3 on the road to longevity: Lessons from SNPs and chromatin hubs. Comput. Struct. Biotechnol. J. 2019, 17, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Grossi, V.; Forte, G.; Sanese, P.; Peserico, A.; Tezil, T.; Lepore Signorile, M.; Fasano, C.; Lovaglio, R.; Bagnulo, R.; Loconte, D.C.; et al. The longevity SNP rs2802292 uncovered: HSF1 activates stress-dependent expression of FOXO3 through an intronic enhancer. Nucleic Acids Res. 2018, 46, 5587–5600. [Google Scholar] [CrossRef] [Green Version]
- Flachsbart, F.; Dose, J.; Gentschew, L.; Geismann, C.; Caliebe, A.; Knecht, C.; Nygaard, M.; Badarinarayan, N.; ElSharawy, A.; May, S.; et al. Identification and characterization of two functional variants in the human longevity gene FOXO3. Nat. Commun. 2017, 8, 2063. [Google Scholar] [CrossRef] [Green Version]
- Deelen, J.; Evans, D.S.; Arking, D.E.; Tesi, N.; Nygaard, M.; Liu, X.; Wojczynski, M.K.; Biggs, M.L.; van der Spek, A.; Atzmon, G.; et al. A meta-analysis of genome-wide association studies identifies multiple longevity genes. Nat. Commun. 2019, 10, 3669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broer, L.; Buchman, A.S.; Deelen, J.; Evans, D.S.; Faul, J.D.; Lunetta, K.L.; Sebastiani, P.; Smith, J.A.; Smith, A.V.; Tanaka, T.; et al. GWAS of longevity in CHARGE consortium confirms APOE and FOXO3 candidacy. J. Gerontol. Ser. A Biomed. Sci. Med. Sci. 2015, 70, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Vousden, K.H. Serine and one-carbon metabolism in cancer. Nat. Rev. Cancer 2016, 16, 650–662. [Google Scholar] [CrossRef] [PubMed]
- Brennan, K.; Flanagan, J.M. Epigenetic epidemiology for cancer risk: Harnessing germline epigenetic variation. Cancer EpiGenet. 2012, 863, 439–465. [Google Scholar]
- Wong, E.M.; Southey, M.C.; Fox, S.B.; Brown, M.A.; Dowty, J.G.; Jenkins, M.A.; Giles, G.G.; Hopper, J.L.; Dobrovic, A. Constitutional methylation of the BRCA1 promoter is specifically associated with BRCA1 mutation-associated pathology in early-onset breast cancer. Cancer Prev. Res. 2011, 4, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brennan, K.; Garcia-Closas, M.; Orr, N.; Fletcher, O.; Jones, M.; Ashworth, A.; Swerdlow, A.; Thorne, H.; Riboli, E.; Vineis, P.; et al. Intragenic ATM methylation in peripheral blood DNA as a biomarker of breast cancer risk. Cancer Res. 2012, 72, 2304–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, S.L. Foxo in the immune system. Oncogene 2008, 27, 2337–2344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Ao, X.; Ding, W.; Ponnusamy, M.; Wu, W.; Hao, X.; Yu, W.; Wang, Y.; Li, P.; Wang, J. Critical role of FOXO3a in carcinogenesis. Mol. Cancer 2018, 17, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornsveld, M.; Dansen, T.B.; Derksen, P.W.; Burgering, B.M.T. Re-evaluating the role of FOXOs in cancer. Semin. Cancer Biol. 2018, 50, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Bullock, M.D.; Bruce, A.; Sreekumar, R.; Curtis, N.; Cheung, T.; Reading, I.; Primrose, J.N.; Ottensmeier, C.; Packham, G.K.; Thomas, G.; et al. FOXO3 expression during colorectal cancer progression: Biomarker potential reflects a tumour suppressor role. Br. J. Cancer 2013, 109, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.; Ren, L.; Wu, D.; Yang, X.; Zhou, Z.; Nie, Q.; Jiang, G.; Xue, S.; Weng, W.; Qiu, Y.; et al. Overexpression of FoxO3a is associated with glioblastoma progression and predicts poor patient prognosis. Int. J. Cancer 2017, 140, 2792–2804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Z.; Zhou, L.; Zhang, C.; Xu, J. Reduction of circular RNA Foxo3 promotes prostate cancer progression and chemoresistance to docetaxel. Cancer Lett. 2020, 468, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Du, W.W.; Li, X.; Yee, A.J.; Yang, B.B. Foxo3 activity promoted by non-coding effects of circular RNA and Foxo3 pseudogene in the inhibition of tumor growth and angiogenesis. Oncogene 2016, 35, 3919–3931. [Google Scholar] [CrossRef]
- Du, W.W.; Fang, L.; Yang, W.; Wu, N.; Awan, F.M.; Yang, Z.; Yang, B.B. Induction of tumor apoptosis through a circular RNA enhancing Foxo3 activity. Cell Death Differ. 2017, 24, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Li, Y.; Zhao, F.; Zhou, L.; Jia, R. Circular RNA Foxo3: A Promising Cancer-Associated Biomarker. Front. Genet. 2021, 12, 405. [Google Scholar] [CrossRef] [PubMed]
- Milne, R.L.; Fletcher, A.S.; MacInnis, R.J.; Hodge, A.M.; Hopkins, A.H.; Bassett, J.K.; Bruinsma, F.J.; Lynch, B.M.; Dugué, P.A.; Jayasekara, H.; et al. Cohort profile: The Melbourne collaborative cohort study (health 2020). Int. J. Epidemiol. 2017, 46, 1757–1757i. [Google Scholar] [CrossRef] [PubMed]
- Dugué, P.A.; Bassett, J.K.; Joo, J.E.; Jung, C.H.; Ming Wong, E.; Moreno-Betancur, M.; Schmidt, D.; Makalic, E.; Li, S.; Severi, G.; et al. DNA methylation-based biological aging and cancer risk and survival: Pooled analysis of seven prospective studies. Int. J. Cancer 2018, 142, 1611–1619. [Google Scholar] [CrossRef] [PubMed]
- Dugué, P.A.; Bassett, J.K.; Wong, E.M.; Joo, J.E.; Li, S.; Yu, C.; Schmidt, D.F.; Makalic, E.; Doo, N.W.; Buchanan, D.D.; et al. Biological aging measures based on blood DNA methylation and risk of cancer: A prospective study. JNCI Cancer Spectr. 2021, 5, pkaa109. [Google Scholar] [CrossRef] [PubMed]
- Dugué, P.A.; Hodge, A.M.; Wong, E.M.; Joo, J.E.; Jung, C.H.; Hopper, J.L.; English, D.R.; Giles, G.G.; Milne, R.L.; Southey, M. Methylation marks of prenatal exposure to maternal smoking and risk of cancer in adulthood. Int. J. Epidemiol. 2021, 50, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Dugué, P.A.; English, D.R.; MacInnis, R.J.; Jung, C.H.; Bassett, J.K.; FitzGerald, L.M.; Wong, E.M.; Joo, J.E.; Hopper, J.L.; Southey, M.C.; et al. Reliability of DNA methylation measures from dried blood spots and mononuclear cells using the HumanMethylation450k BeadArray. Sci. Rep. 2016, 6, 30317. [Google Scholar] [CrossRef] [Green Version]
- Du, P.; Zhang, X.; Huang, C.C.; Jafari, N.; Kibbe, W.A.; Hou, L.; Lin, S.M. Comparison of Beta-value and M-value methods for quantifying methylation levels by microarray analysis. BMC Bioinform. 2010, 11, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amos, C.I.; Dennis, J.; Wang, Z.; Byun, J.; Schumacher, F.R.; Gayther, S.A.; Casey, G.; Hunter, D.J.; Sellers, T.A.; Gruber, S.B.; et al. The OncoArray Consortium: A network for understanding the genetic architecture of common cancers. Cancer Epidemiol. Prev. Biomark. 2017, 26, 126–135. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Forer, L.; Schönherr, S.; Sidore, C.; Locke, A.E.; Kwong, A.; Vrieze, S.I.; Chew, E.Y.; Levy, S.; McGue, M.; et al. Next-generation genotype imputation service and methods. Na. Genet. 2016, 48, 1284–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howie, B.N.; Donnelly, P.; Marchini, J. A flexible and accurate genotype imputation method for the next generation of genome-wide association studies. PLoS Genet. 2009, 5, e1000529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dugué, P.A.; Yu, C.; McKay, T.; Wong, E.M.; Joo, J.E.; Tsimiklis, H.; Hammet, F.; Mahmoodi, M.; Theys, D.; Hopper, J.L.; et al. VTRNA2–1: Genetic variation, heritable methylation and disease association. Int. J. Mol. Sci. 2021, 22, 2535. [Google Scholar] [CrossRef]
- Houseman, E.A.; Accomando, W.P.; Koestler, D.C.; Christensen, B.C.; Marsit, C.J.; Nelson, H.H.; Wiencke, J.K.; Kelsey, K.T. DNA methylation arrays as surrogate measures of cell mixture distribution. BMC Bioinform. 2012, 13, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dugué, P.A.; Jung, C.H.; Joo, J.E.; Wang, X.; Wong, E.M.; Makalic, E.; Schmidt, D.F.; Baglietto, L.; Severi, G.; Southey, M.C.; et al. Smoking and blood DNA methylation: An epigenome-wide association study and assessment of reversibility. EpiGenet 2020, 15, 358–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dugué, P.A.; Wilson, R.; Lehne, B.; Jayasekara, H.; Wang, X.; Jung, C.H.; Joo, J.E.; Makalic, E.; Schmidt, D.F.; Baglietto, L.; et al. Alcohol consumption is associated with widespread changes in blood DNA methylation: Analysis of cross-sectional and longitudinal data. Addict. Biol. 2021, 26, e12855. [Google Scholar] [CrossRef]
- Geurts, Y.M.; Dugué, P.A.; Joo, J.E.; Makalic, E.; Jung, C.H.; Guan, W.; Nguyen, S.; Grove, M.L.; Wong, E.M.; Hodge, A.M.; et al. Novel associations between blood DNA methylation and body mass index in middle-aged and older adults. Int. J. Obes. 2018, 42, 887–896. [Google Scholar] [CrossRef] [PubMed]
- Therneau, T.M.; Grambsch, P.M.; Pankratz, V.S. Penalized survival models and frailty. J. Comput. Graph. Stat. 2003, 12, 156–175. [Google Scholar] [CrossRef]
- Thiébaut, A.C.; Bénichou, J. Choice of time-scale in Cox’s model analysis of epidemiologic cohort data: A simulation study. Stat. Med. 2004, 23, 3803–3820. [Google Scholar] [CrossRef] [PubMed]
- Salas-Pérez, F.; Ramos-Lopez, O.; Mansego, M.L.; Milagro, F.I.; Santos, J.L.; Riezu-Boj, J.I.; Martínez, J.A. DNA methylation in genes of longevity-regulating pathways: Association with obesity and metabolic complications. Aging 2019, 11, 1874. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Wei, Y.; Wang, J.; Chen, S.; Cai, J.; Tang, J.; Xu, X.; Long, B.; Yu, G.; Zhang, Z.; et al. Association between SIRT6 methylation and human longevity in a Chinese population. Public Health Genom. 2020, 23, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Vitiello, M.; Zullo, A.; Servillo, L.; Mancini, F.P.; Borriello, A.; Giovane, A.; Della Ragione, F.; D’Onofrio, N.; Balestrieri, M.L. Multiple pathways of SIRT6 at the crossroads in the control of longevity, cancer, and cardiovascular diseases. Aging Res. Rev. 2017, 35, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Szymczak, S.; Dose, J.; Torres, G.G.; Heinsen, F.A.; Venkatesh, G.; Datlinger, P.; Nygaard, M.; Mengel-From, J.; Flachsbart, F.; Klapper, W.; et al. DNA methylation QTL analysis identifies new regulators of human longevity. Hum. Mol. Genet. 2020, 29, 1154–1167. [Google Scholar] [CrossRef]
- Schächter, F.; Faure-Delanef, L.; Guénot, F.; Rouger, H.; Froguel, P.; Lesueur-Ginot, L.; Cohen, D. Genetic associations with human longevity at the APOE and ACE loci. Nat. Genet. 1994, 6, 29–32. [Google Scholar] [CrossRef]
- Gentilini, D.; Mari, D.; Castaldi, D.; Remondini, D.; Ogliari, G.; Ostan, R.; Bucci, L.; Sirchia, S.M.; Tabano, S.; Cavagnini, F.; et al. Role of epigenetics in human aging and longevity: Genome-wide DNA methylation profile in centenarians and centenarians’ offspring. Age 2013, 35, 1961–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenet, F.; Moh, M.; Funk, P.; Feierstein, E.; Viale, A.J.; Socci, N.D.; Scandura, J.M. DNA methylation of the first exon is tightly linked to transcriptional silencing. PLoS ONE 2011, 6, e14524. [Google Scholar] [CrossRef] [PubMed]



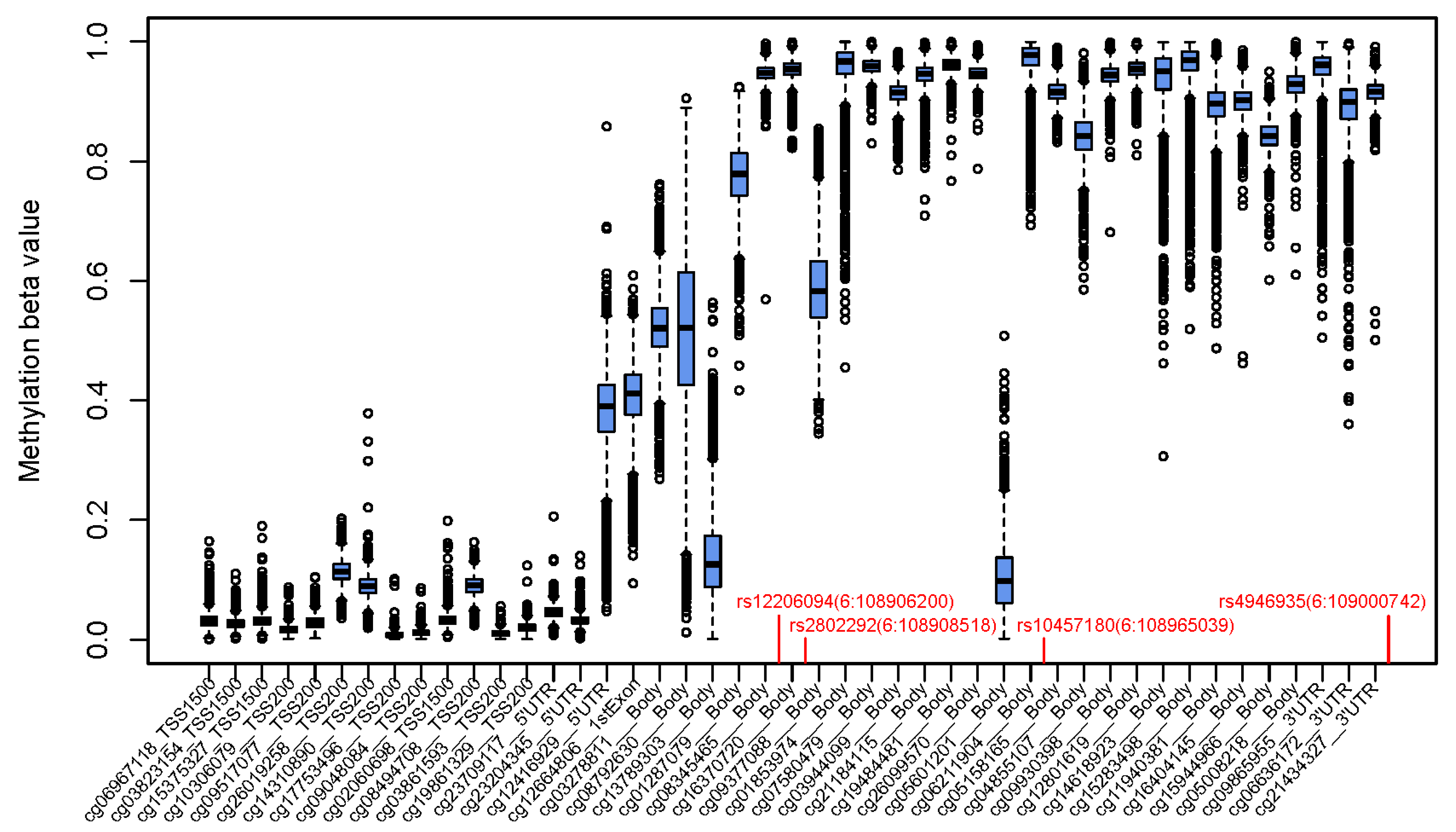{kind=link}
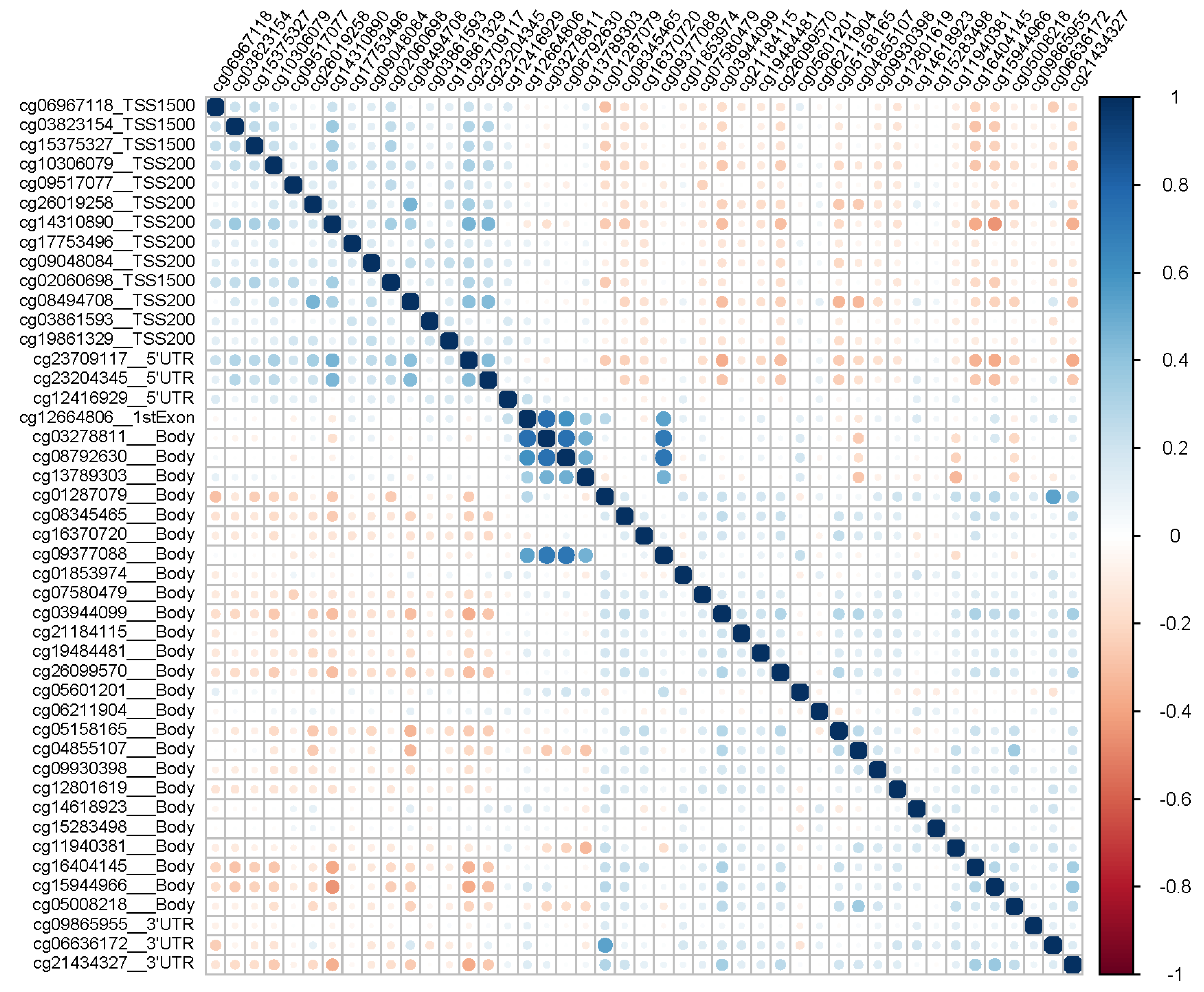{kind=link}
| Participant Characteristic | Matched Case-Control Pairs | Follow-Up Sample (n = 1088) | |
|---|---|---|---|
| Cases of Overall Cancer (n = 3481) | Controls (n = 3481) | ||
| Age at blood draw, median [IQR] | 60.8 [54.1–65.9] | 60.8 [53.9–65.6] | 69.9 [62.8–75.6] |
| Sex: | |||
| Male, n (%) | 2125 (61%) | 2125 (61%) | 740 (68%) |
| Female, n (%) | 1356 (39%) | 1356 (39%) | 348 (32%) |
| Country of birth: | |||
| Australia/New-Zealand, n (%) | 2380 (68%) | 2367 (68%) | 831 (77%) |
| Greece, n (%) | 350 (10%) | 361 (10%) | 44 (4%) |
| Italy, n (%) | 520 (15%) | 520 (15%) | 89 (8%) |
| UK/other, n (%) | 231 (7%) | 233 (7%) | 124 (11%) |
| Blood sample type: | |||
| Dried blood spots, n (%) | 2412 (69%) | 2412 (69%) | |
| Peripheral blood mononuclear cells, n (%) | 873 (25%) | 873 (25%) | 1088 (100%) |
| Buffy coats, n (%) | 196 (6%) | 196 (6%) | |
| Smoking status: | |||
| Current, n (%) | 490 (14%) | 472 (14%) | 62 (6%) |
| Former, n (%) | 1377 (40%) | 1337 (38%) | 478 (44%) |
| Never, n (%) | 1613 (46%) | 1671 (48%) | 548 (50%) |
| Smoking pack-years, median [IQR] | 1.6 [0–28.5] | 0.75 [0–25.7] | - |
| Body mass index (kg/m2), median [IQR] | 26.8 [24.4–29.6] | 26.7 [24.4–29.3] | 26.8 [24.2–29.4] |
| Alcohol consumption a (g/day), median [IQR] | 4.3 [0–17.7] | 4.3 [0–17.0] | 8.8 [0–23.0] |
| CpG | CHR | MAPINFO | Gene Group | Model 2.1 | Model 2.2 | Cancer Type | ||
|---|---|---|---|---|---|---|---|---|
| HR (95%CI) | p Value | HR (95%CI) | p Value | |||||
| cg12664806 | 6 | 108882981 | 1st Exon | 2.39 (1.60–3.56) | 1.85 × 10−5 | 2.50 (1.67–3.74) | 7.51 × 10−6 | Gastric |
| cg05158165 | 6 | 108965753 | Body | 0.73 (0.64–0.85) | 1.92 × 10−5 | 0.73 (0.63–0.84) | 1.81 × 10−5 | Lung |
| cg03823154 | 6 | 108879922 | TSS1500 | 1.33 (1.16–1.52) | 5.92 × 10−5 | 1.32 (1.14–1.51) | 1.23 × 10−4 | |
| cg16404145 | 6 | 108984834 | Body | 0.76 (0.67–0.87) | 6.08 × 10−5 | 0.77 (0.68–0.88) | 1.12 × 10−4 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, C.; Hodge, A.M.; Wong, E.M.; Joo, J.E.; Makalic, E.; Schmidt, D.; Buchanan, D.D.; Hopper, J.L.; Giles, G.G.; Southey, M.C.; et al. Association of FOXO3 Blood DNA Methylation with Cancer Risk, Cancer Survival, and Mortality. Cells 2021, 10, 3384. https://doi.org/10.3390/cells10123384
Yu C, Hodge AM, Wong EM, Joo JE, Makalic E, Schmidt D, Buchanan DD, Hopper JL, Giles GG, Southey MC, et al. Association of FOXO3 Blood DNA Methylation with Cancer Risk, Cancer Survival, and Mortality. Cells. 2021; 10(12):3384. https://doi.org/10.3390/cells10123384
Chicago/Turabian StyleYu, Chenglong, Allison M. Hodge, Ee Ming Wong, Jihoon Eric Joo, Enes Makalic, Daniel Schmidt, Daniel D. Buchanan, John L. Hopper, Graham G. Giles, Melissa C. Southey, and et al. 2021. "Association of FOXO3 Blood DNA Methylation with Cancer Risk, Cancer Survival, and Mortality" Cells 10, no. 12: 3384. https://doi.org/10.3390/cells10123384
APA StyleYu, C., Hodge, A. M., Wong, E. M., Joo, J. E., Makalic, E., Schmidt, D., Buchanan, D. D., Hopper, J. L., Giles, G. G., Southey, M. C., & Dugué, P.-A. (2021). Association of FOXO3 Blood DNA Methylation with Cancer Risk, Cancer Survival, and Mortality. Cells, 10(12), 3384. https://doi.org/10.3390/cells10123384