
RNF13 Dileucine Motif Variants L311S and L312P Interfere with Endosomal Localization and AP-3 Complex Association



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies
2.2. Molecular Biology
2.3. Cell Culture and Transfection
2.4. Immunostaining
2.5. Uptake and Trafficking of EGF and Transferrin
2.6. Fluorescence Microscopy Image Acquisition
2.7. Image Analysis
2.8. Statistical Analysis
2.9. TOPP2 E. coli Transformation and Culture
2.10. Purification of Recombinant C-Terminal RNF13 from TOPP2
2.11. GST Pulldown and Immunoprecipitation Assays
2.12. SDS–PAGE and Western Blot
2.13. Predicted Structure
2.14. AP3D1 Knockdown
3. Results
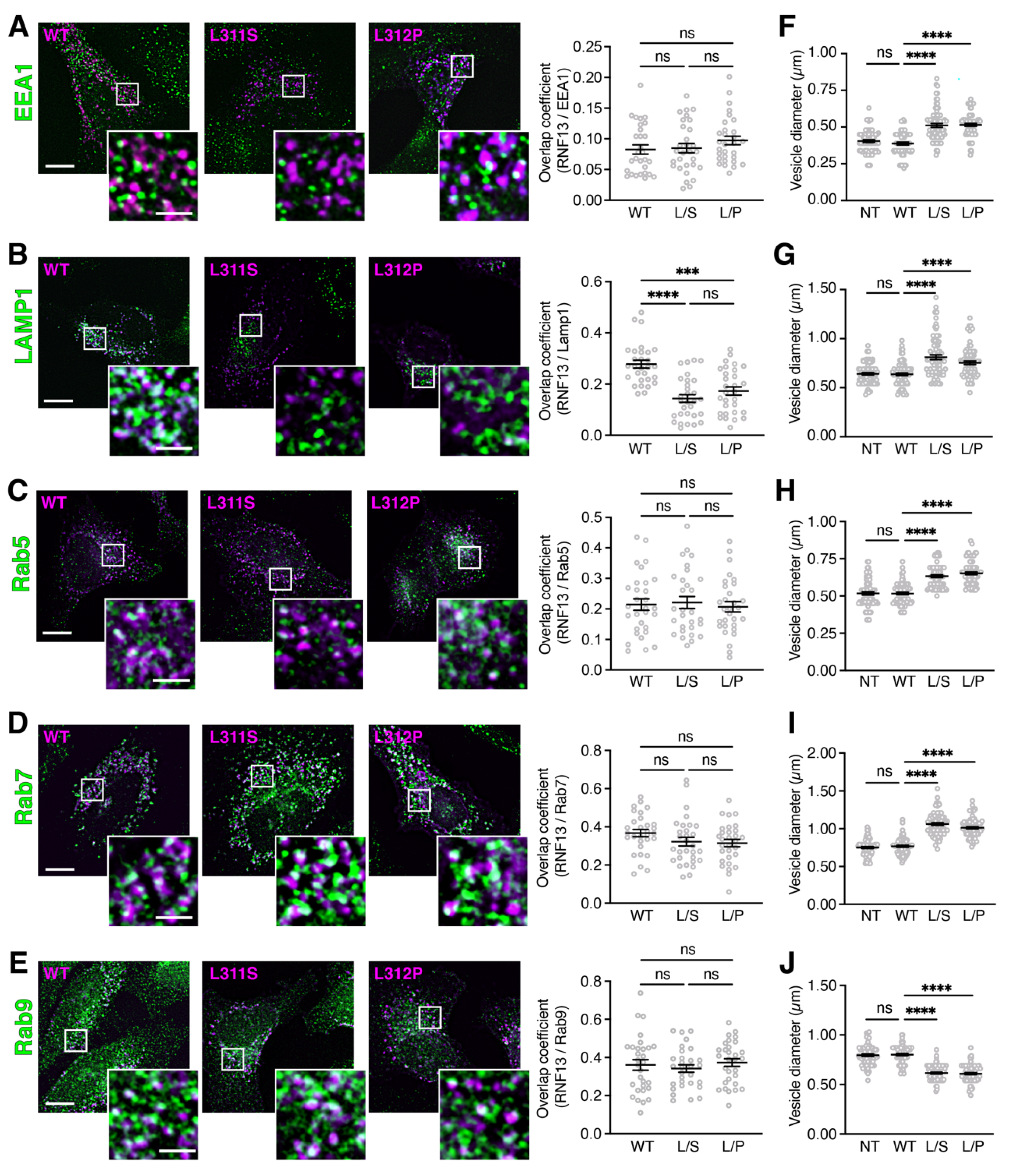
3.1. RNF13 Variants L311S and L312P Show Reduced Presence in Lysosomes and Alter Endosomal Vesicle Size
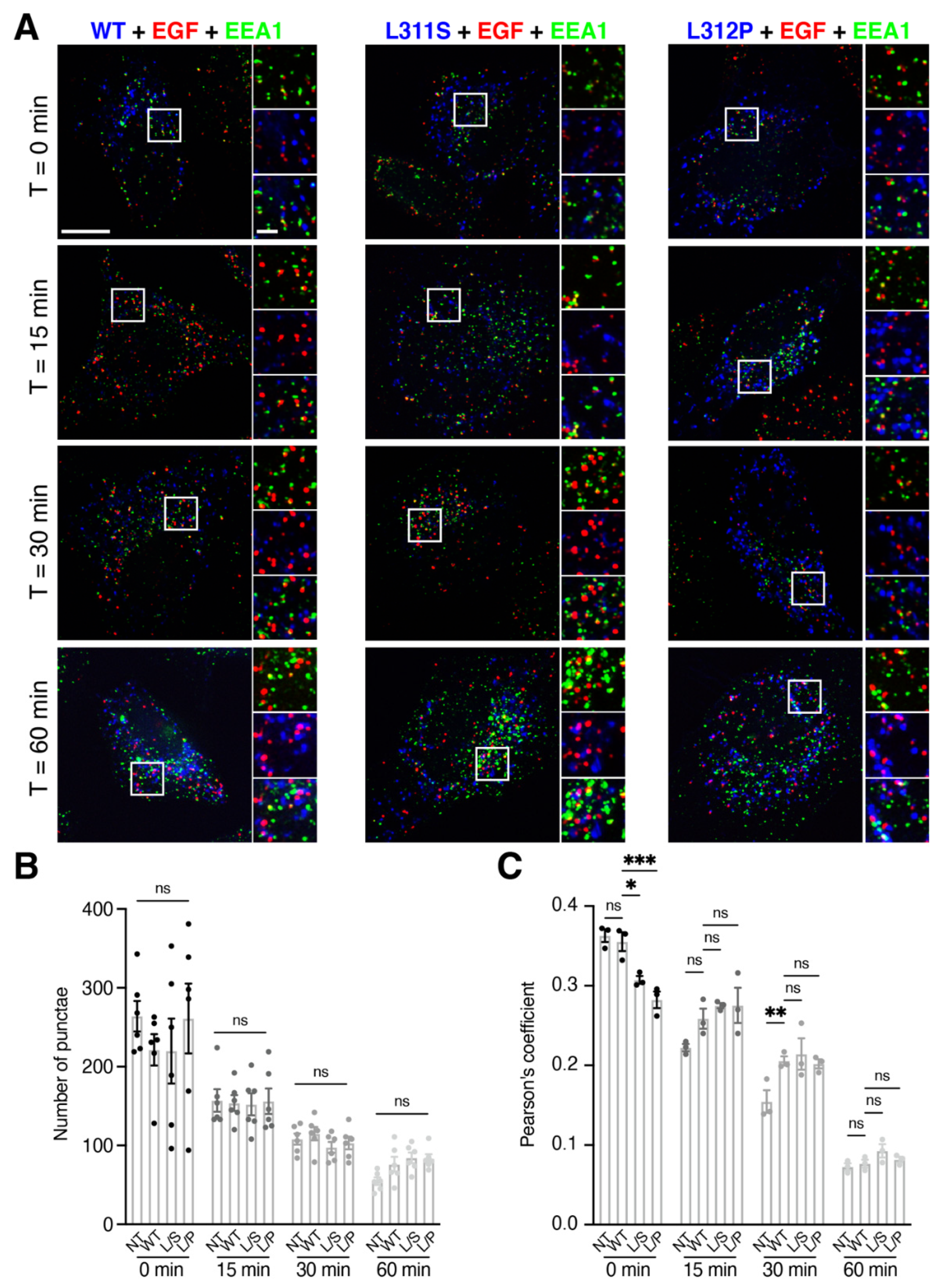
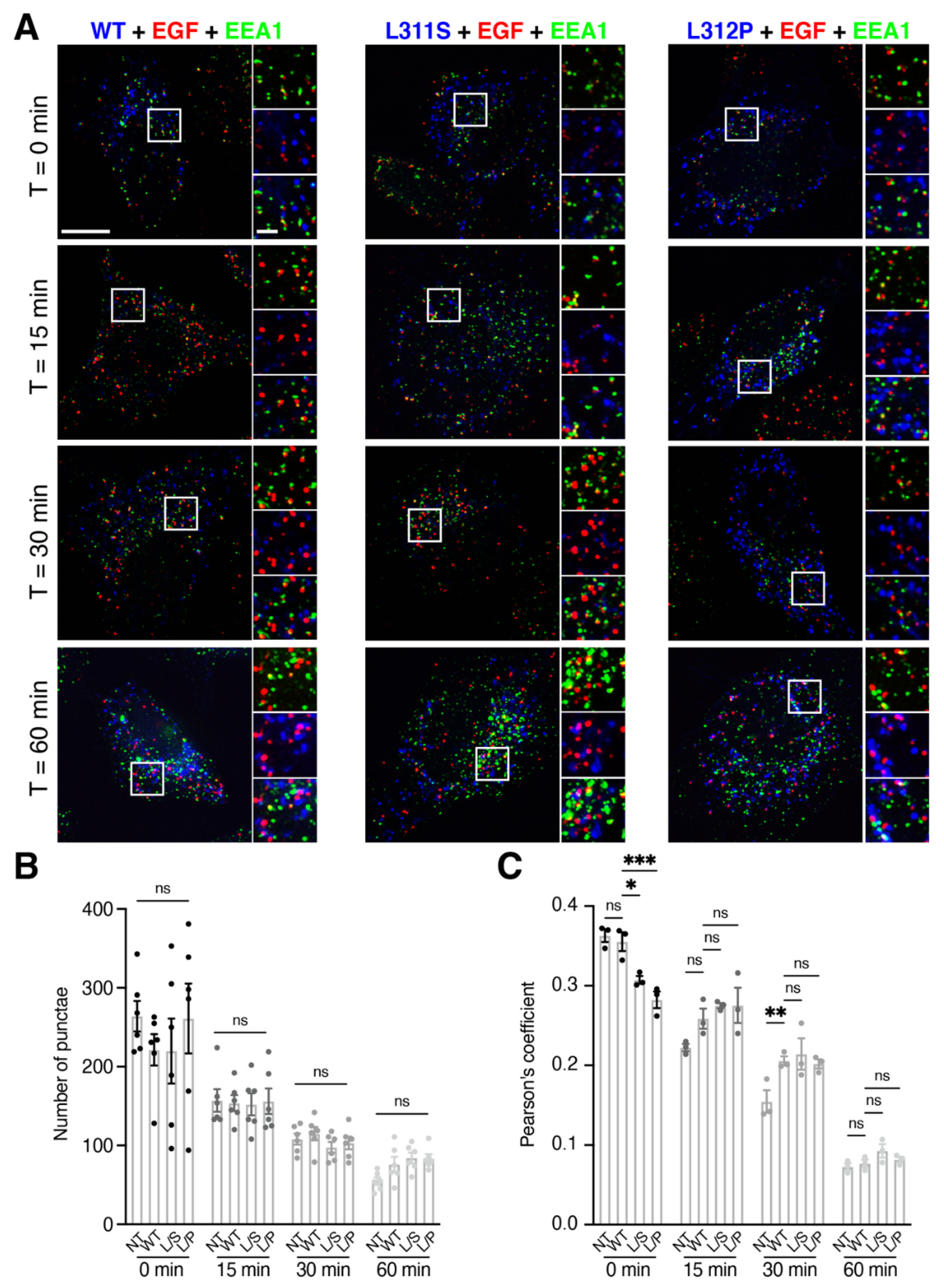
3.2. RNF13 Variants L311S and L312P alter EGF Sorting at Early Endosomes
3.3. RNF13 Variants L311S and L312P Do Not Alter Transferrin Trafficking
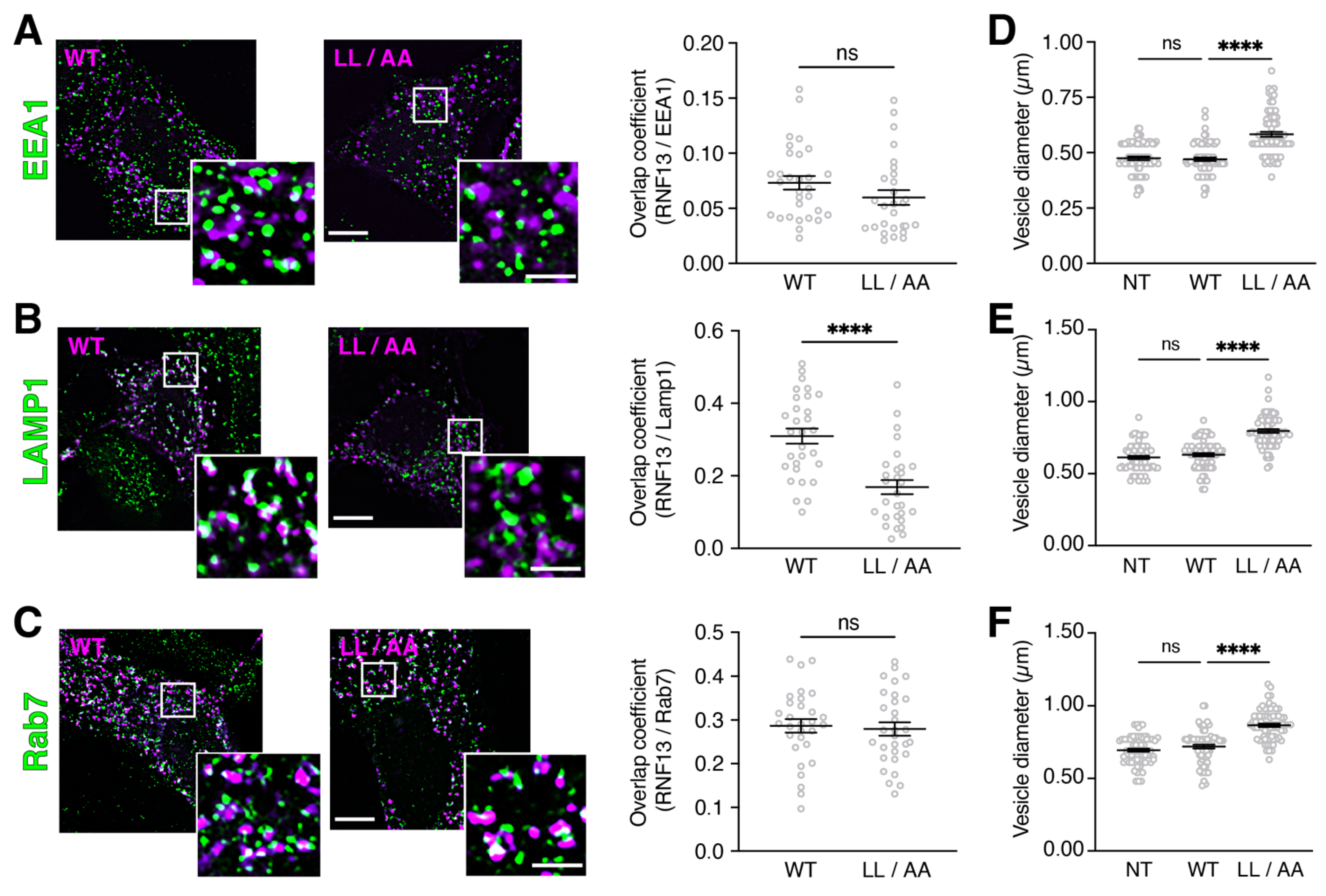
3.4. RNF13 Variant L311L312/AA Shows Reduced Presence in Lysosomes and Increases Endosomal Vesicle Size
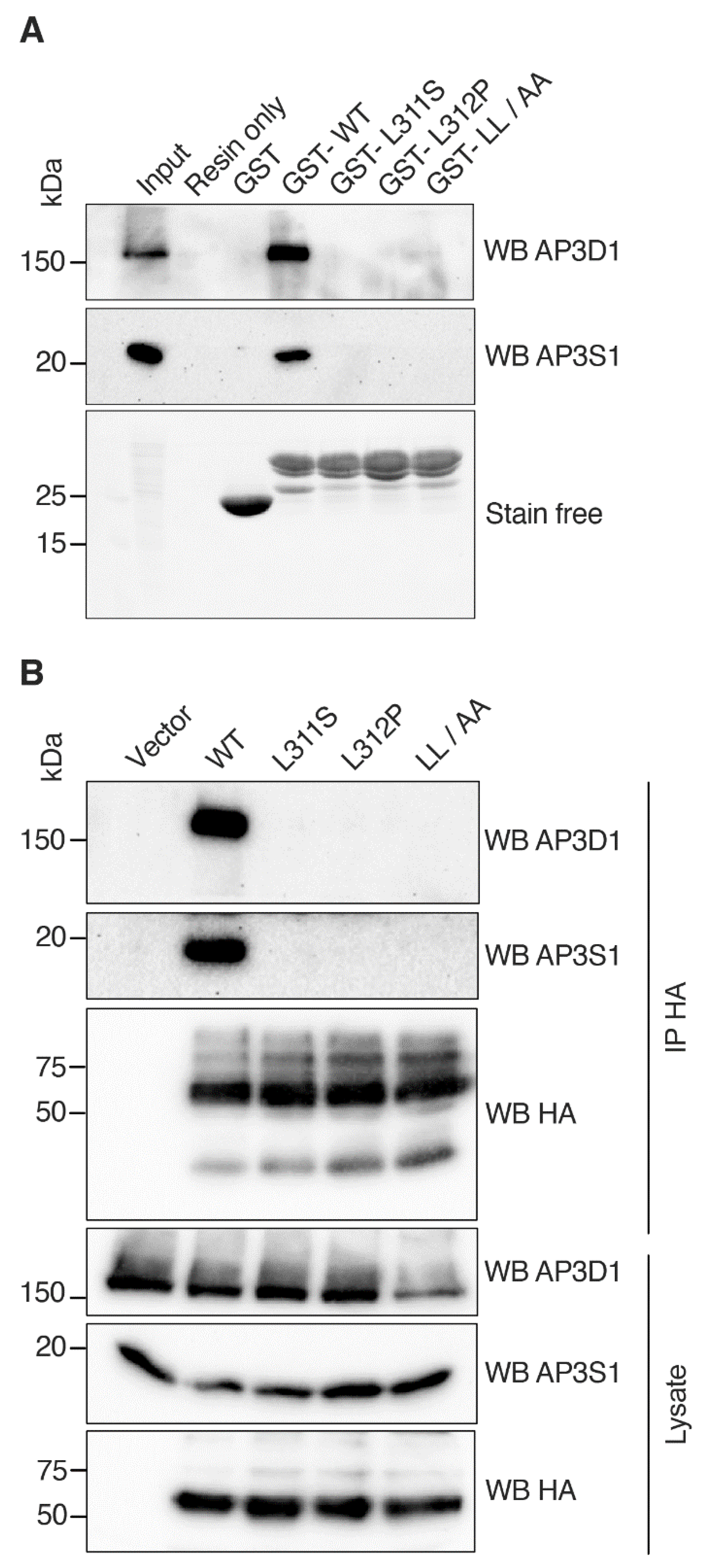
3.5. RNF13 Variants L311S and L312P Do Not Interact with AP-3 Complex
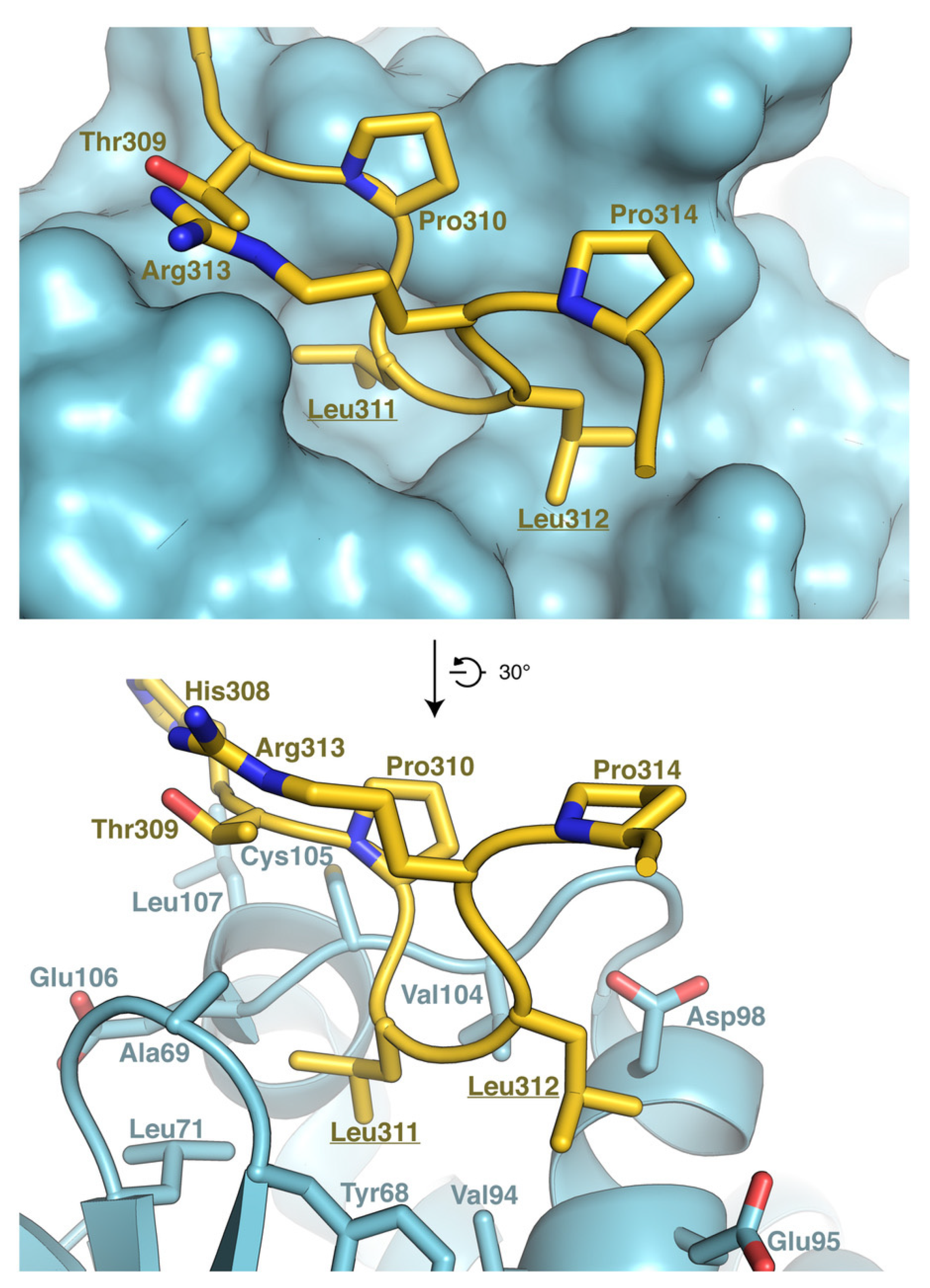
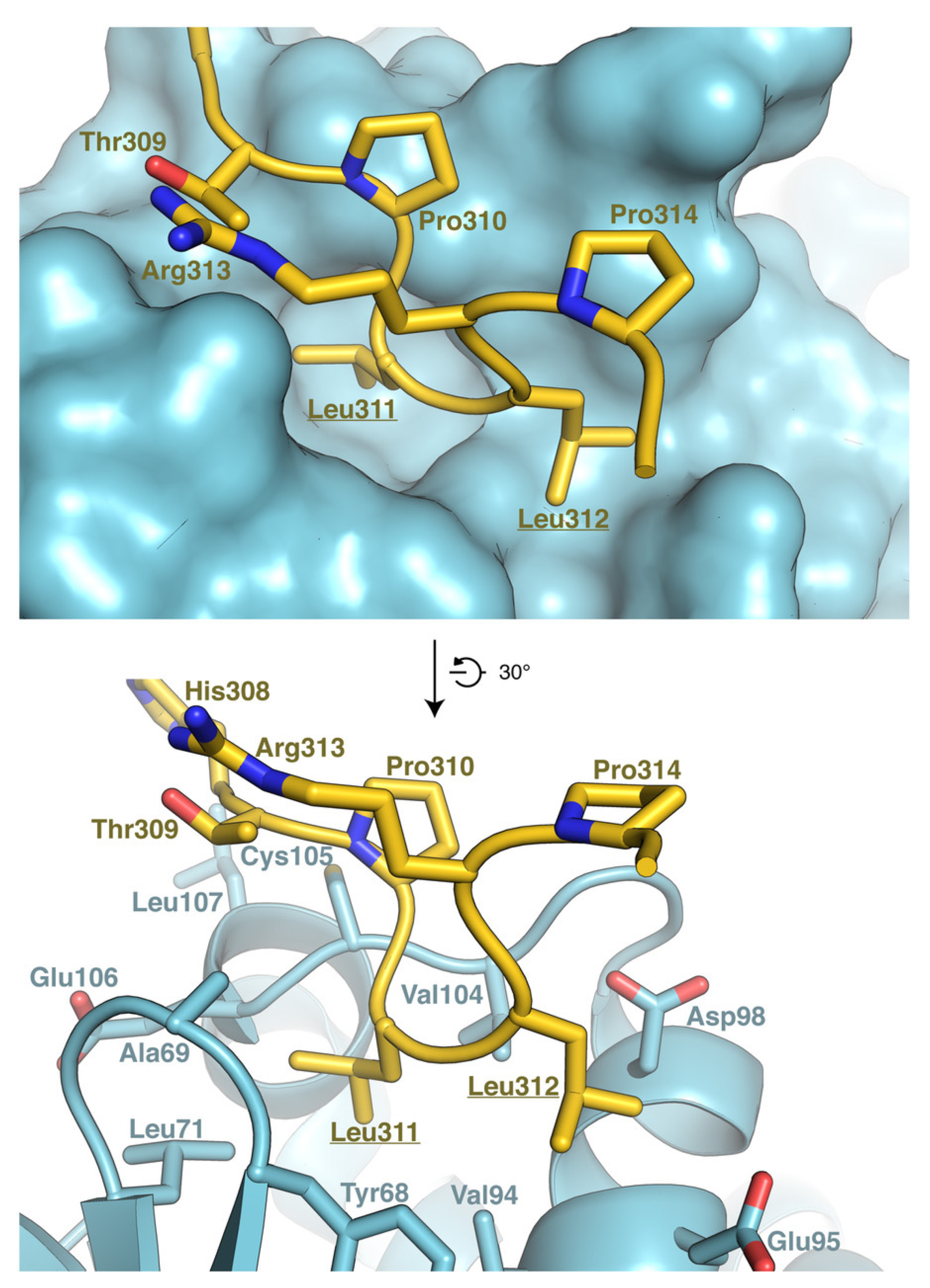
3.6. Model of RNF13 and AP3S1 Complex Association
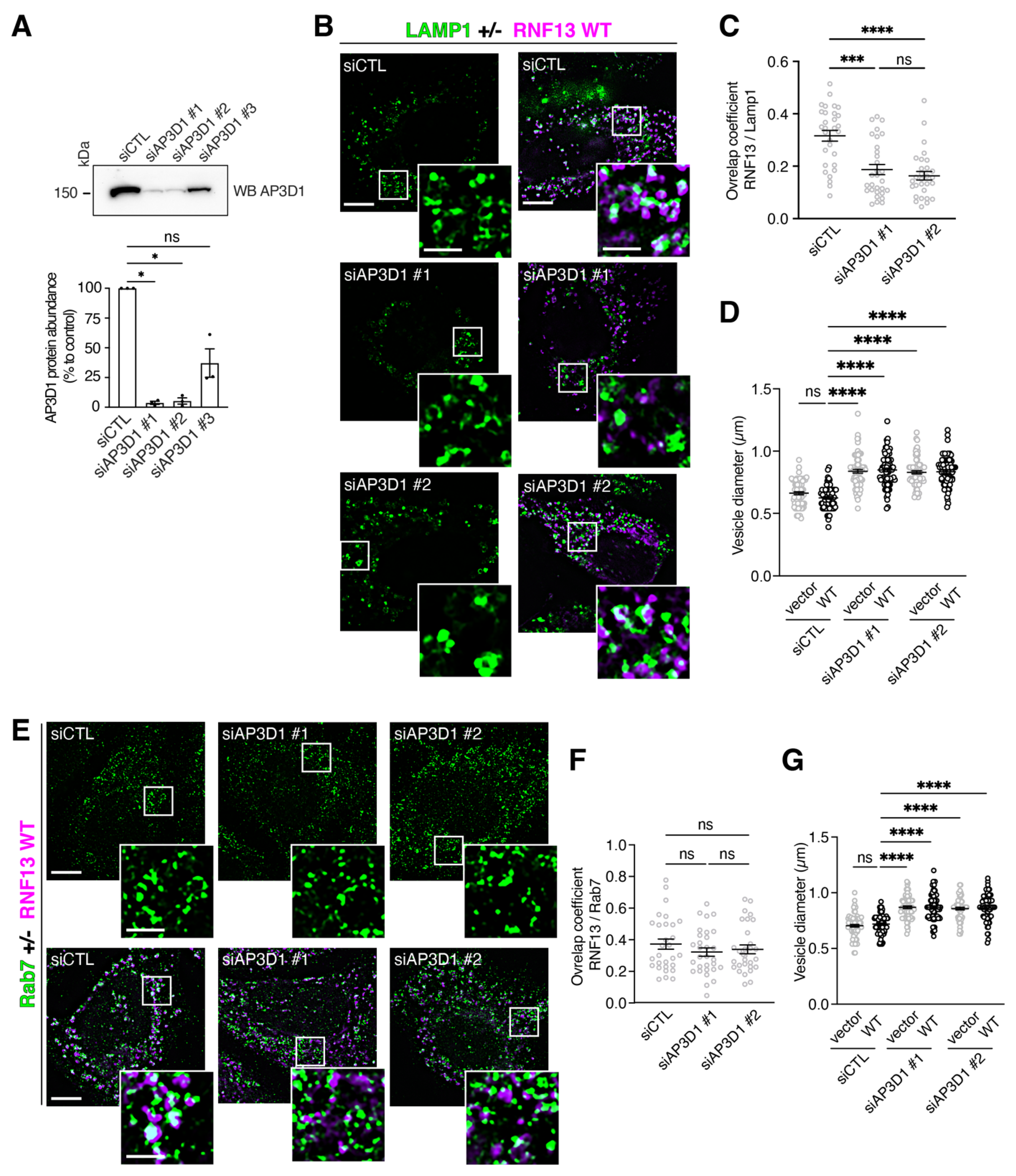
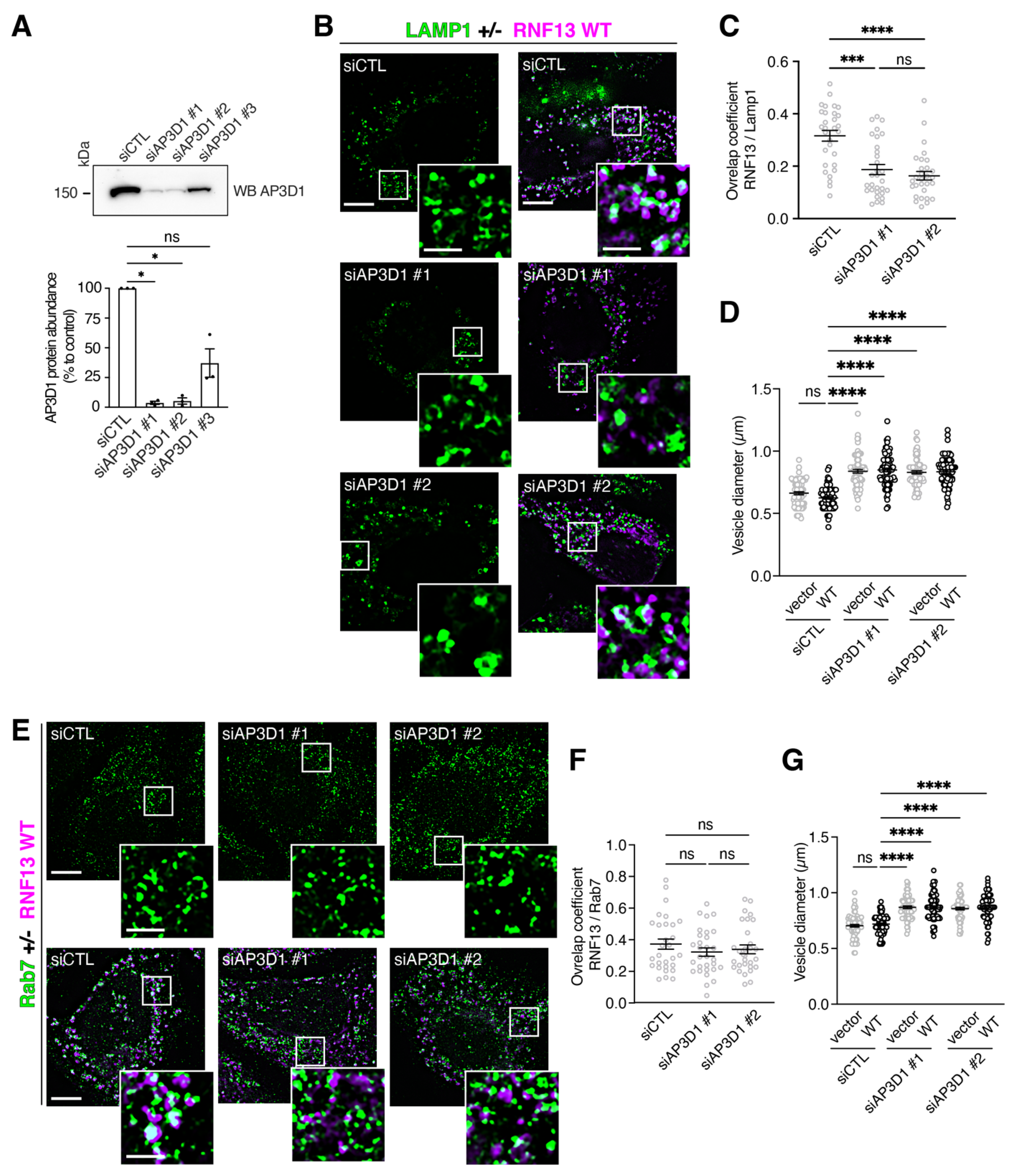
3.7. Knockdown of AP-3 Gene Expressions Reduces RNF13 Localization to Lysosomes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Komander, D.; Rape, M. The ubiquitin code. Annu. Rev. Biochem. 2012, 81, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Hicke, L.; Dunn, R. Regulation of membrane protein transport by ubiquitin and ubiquitin-binding proteins. Annu. Rev. Cell Dev. Biol. 2003, 19, 141–172. [Google Scholar] [CrossRef]
- Ciechanover, A.; Iwai, K. The ubiquitin system: From basic mechanisms to the patient bed. IUBMB Life 2004, 56, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Komander, D. The emerging complexity of protein ubiquitination. Biochem. Soc. Trans. 2009, 37, 937–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhopadhyay, D.; Riezman, H. Proteasome-independent functions of ubiquitin in endocytosis and signaling. Science 2007, 315, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Culley, M.K.; Zhao, Y.; Zhao, J. The role of ubiquitination and deubiquitination in the regulation of cell junctions. Protein Cell 2018, 9, 754–769. [Google Scholar] [CrossRef]
- Haglund, K.; Dikic, I. The role of ubiquitylation in receptor endocytosis and endosomal sorting. J. Cell Sci. 2012, 125, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Jana, N.R. Protein homeostasis and aging: Role of ubiquitin protein ligases. Neurochem. Int. 2012, 60, 443–447. [Google Scholar] [CrossRef]
- Basel-Vanagaite, L.; Dallapiccola, B.; Ramirez-Solis, R.; Segref, A.; Thiele, H.; Edwards, A.; Arends, M.J.; Miró, X.; White, J.K.; Désir, J.; et al. Deficiency for the ubiquitin ligase UBE3B in a blepharophimosis-ptosis-intellectual-disability syndrome. Am. J. Hum. Genet. 2012, 91, 998–1010. [Google Scholar] [CrossRef] [Green Version]
- Gu, B.; Carstens, K.E.; Judson, M.C.; Dalton, K.A.; Rougié, M.; Clark, E.P.; Dudek, S.M.; Philpot, B.D. Ube3a reinstatement mitigates epileptogenesis in Angelman syndrome model mice. J. Clin. Investig. 2019, 129, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Hong, L.; Huang, H.C.; Jiang, Z.F. Relationship between amyloid-beta and the ubiquitin-proteasome system in Alzheimer’s disease. Neurol. Res. 2014, 36, 276–282. [Google Scholar] [CrossRef]
- Cao, J.; Zhong, M.B.; Toro, C.A.; Zhang, L.; Cai, D. Endo-lysosomal pathway and ubiquitin-proteasome system dysfunction in Alzheimer’s disease pathogenesis. Neurosci. Lett. 2019, 703, 68–78. [Google Scholar] [CrossRef]
- Kumar, V.; Singh, D.; Singh, B.K.; Singh, S.; Mittra, N.; Jha, R.R.; Patel, D.K.; Singh, C. Alpha-synuclein aggregation, Ubiquitin proteasome system impairment, and L-Dopa response in zinc-induced Parkinsonism: Resemblance to sporadic Parkinson’s disease. Mol. Cell Biochem. 2018, 444, 149–160. [Google Scholar] [CrossRef]
- Olanow, C.W.; McNaught, K.S. Ubiquitin-proteasome system and Parkinson’s disease. Mov. Disord. 2006, 21, 1806–1823. [Google Scholar] [CrossRef]
- Paul, S. Dysfunction of the ubiquitin-proteasome system in multiple disease conditions: Therapeutic approaches. Bioessays 2008, 30, 1172–1184. [Google Scholar] [CrossRef]
- Scheffner, M.; Huibregtse, J.M.; Vierstra, R.D.; Howley, P.M. The HPV-16 E6 and E6-AP complex functions as a ubiquitin-protein ligase in the ubiquitination of p53. Cell 1993, 75, 495–505. [Google Scholar] [CrossRef]
- Hashizume, R.; Fukuda, M.; Maeda, I.; Nishikawa, H.; Oyake, D.; Yabuki, Y.; Ogata, H.; Ohta, T. The RING heterodimer BRCA1-BARD1 is a ubiquitin ligase inactivated by a breast cancer-derived mutation. J. Biol. Chem. 2001, 276, 14537–14540. [Google Scholar] [CrossRef] [Green Version]
- Ardley, H.C.; Robinson, P.A. E3 ubiquitin ligases. Essays Biochem. 2005, 41, 15–30. [Google Scholar] [CrossRef]
- Deshaies, R.J.; Joazeiro, C.A. RING domain E3 ubiquitin ligases. Annu. Rev. Biochem. 2009, 78, 399–434. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Bengtson, M.H.; Ulbrich, A.; Matsuda, A.; Reddy, V.A.; Orth, A.; Chanda, S.K.; Batalov, S.; Joazeiro, C.A.P. Genome-wide and functional annotation of human E3 ubiquitin ligases identifies MULAN, a mitochondrial E3 that regulates the organelle’s dynamics and signaling. PLoS ONE 2008, 3, e1487. [Google Scholar] [CrossRef]
- Nakamura, N. The Role of the Transmembrane RING Finger Proteins in Cellular and Organelle Function. Membranes 2011, 1, 354–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neutzner, A.; Neutzner, M.; Benischke, A.S.; Ryu, S.W.; Frank, S.; Youle, R.J.; Karbowski, M. A systematic search for endoplasmic reticulum (ER) membrane-associated RING finger proteins identifies Nixin/ZNRF4 as a regulator of calnexin stability and ER homeostasis. J. Biol. Chem. 2011, 286, 8633–8643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dijk, J.R.; Yamazaki, Y.; Palmer, R.H. Tumour-associated mutations of PA-TM-RING ubiquitin ligases RNF167/RNF13 identify the PA domain as a determinant for endosomal localization. Biochem. J. 2014, 459, 27–36. [Google Scholar] [CrossRef]
- Deshar, R.; Moon, S.; Yoo, W.; Cho, E.B.; Yoon, S.K.; Yoon, J.B. RNF167 targets Arl8B for degradation to regulate lysosome positioning and endocytic trafficking. FEBS J. 2016, 283, 4583–4599. [Google Scholar] [CrossRef] [PubMed]
- Lussier, M.P.; Herring, B.E.; Nasu-Nishimura, Y.; Neutzner, A.; Karbowski, M.; Youle, R.J.; Nicoll, R.A.; Roche, K.W. Ubiquitin ligase RNF167 regulates AMPA receptor-mediated synaptic transmission. Proc. Natl. Acad. Sci. USA 2012, 109, 19426–19431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghilarducci, K.; Cabana, V.C.; Desroches, C.; Chabi, K.; Bourgault, S.; Cappadocia, L.; Lussier, M.P. Functional interaction of ubiquitin ligase RNF167 with UBE2D1 and UBE2N promotes ubiquitination of AMPA receptor. FEBS J. 2021. [Google Scholar] [CrossRef]
- Bocock, J.P.; Carmicle, S.; Sircar, M.; Erickson, A.H. Trafficking and proteolytic processing of RNF13, a model PA-TM-RING family endosomal membrane ubiquitin ligase. FEBS J. 2011, 278, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Bocock, J.P.; Carmicle, S.; Chhotani, S.; Ruffolo, M.R.; Chu, H.; Erickson, A.H. The PA-TM-RING protein RING finger protein 13 is an endosomal integral membrane E3 ubiquitin ligase whose RING finger domain is released to the cytoplasm by proteolysis. FEBS J. 2009, 276, 1860–1877. [Google Scholar] [CrossRef]
- Zhang, Q.; Meng, Y.; Zhang, L.; Chen, J.; Zhu, D. RNF13: A novel RING-type ubiquitin ligase over-expressed in pancreatic cancer. Cell Res. 2009, 19, 348–357. [Google Scholar] [CrossRef]
- Arshad, M.; Ye, Z.; Gu, X.; Wong, C.K.; Liu, Y.; Li, D.; Zhou, L.; Zhang, Y.; Bay, W.P.; Yu, V.C.; et al. RNF13, a RING finger protein, mediates endoplasmic reticulum stress-induced apoptosis through the inositol-requiring enzyme (IRE1α)/c-Jun NH2-terminal kinase pathway. J. Biol. Chem. 2013, 288, 8726–8736. [Google Scholar] [CrossRef] [Green Version]
- Tranque, P.; Crossin, K.L.; Cirelli, C.; Edelman, G.M.; Mauro, V.P. Identification and characterization of a RING zinc finger gene (C-RZF) expressed in chicken embryo cells. Proc. Natl. Acad. Sci. USA 1996, 93, 3105–3109. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Wang, K.; Zhang, Y.; Meng, J.; Yu, F.; Chen, Y.; Zhu, D. The myostatin-induced E3 ubiquitin ligase RNF13 negatively regulates the proliferation of chicken myoblasts. FEBS J. 2010, 277, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Cheng, H.; Chen, J.; Zhu, D. RNF13: An emerging RING finger ubiquitin ligase important in cell proliferation. FEBS J. 2011, 278, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Qin, X.; Zhou, X.; Zhou, J.; Wen, P.; Chen, S.; Wu, M.; Wu, Y.; Zhuang, J. Analysis of genes and underlying mechanisms involved in foam cells formation and atherosclerosis development. PeerJ 2020, 8, e10336. [Google Scholar] [CrossRef] [PubMed]
- Ness, C.; Katta, K.; Garred, Ø.; Kumar, T.; Olstad, O.K.; Petrovski, G.; Moe, M.C.; Noer, A. Integrated differential DNA methylation and gene expression of formalin-fixed paraffin-embedded uveal melanoma specimens identifies genes associated with early metastasis and poor prognosis. Exp. Eye Res. 2020, 203, 108426. [Google Scholar] [CrossRef] [PubMed]
- Bazan-Socha, S.; Buregwa-Czuma, S.; Jakiela, B.; Zareba, L.; Zawlik, I.; Myszka, A.; Soja, J.; Okon, K.; Zarychta, J.; Kozlik, P.; et al. Reticular Basement Membrane Thickness Is Associated with Growth- and Fibrosis-Promoting Airway Transcriptome Profile-Study in Asthma Patients. Int. J. Mol. Sci. 2021, 22, 998. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Honma, K.; Kita-Matsuo, H.; Ochiya, T.; Kato, K. Gene expression profiling of cerebellar development with high-throughput functional analysis. Physiol Genom. 2005, 22, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Li, Y.; Zhang, L.; Yang, N.; Meng, J.; Zuo, P.; Zhang, Y.; Chen, J.; Wang, L.; Gao, X.; et al. E3 ubiquitin ligase RNF13 involves spatial learning and assembly of the SNARE complex. Cell Mol. Life Sci. 2013, 70, 153–165. [Google Scholar] [CrossRef]
- Dingerdissen, H.M.; Torcivia-Rodriguez, J.; Hu, Y.; Chang, T.C.; Mazumder, R.; Kahsay, R. BioMuta and BioXpress: Mutation and expression knowledgebases for cancer biomarker discovery. Nucleic Acids Res. 2018, 46, D1128–D1136. [Google Scholar] [CrossRef] [Green Version]
- Edvardson, S.; Nicolae, C.M.; Noh, G.J.; Burton, J.E.; Punzi, G.; Shaag, A.; Bischetsrieder, J.; De Grassi, A.; Pierri, C.L.; Elpeleg, O.; et al. Heterozygous RNF13 Gain-of-Function Variants Are Associated with Congenital Microcephaly, Epileptic Encephalopathy, Blindness, and Failure to Thrive. Am. J. Hum. Genet. 2019, 104, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Lussier, M.P.; Cayouette, S.; Lepage, P.K.; Bernier, C.L.; Francoeur, N.; St-Hilaire, M.; Pinard, M.; Boulay, G. MxA, a member of the dynamin superfamily, interacts with the ankyrin-like repeat domain of TRPC. J. Biol. Chem. 2005, 280, 19393–19400. [Google Scholar] [CrossRef] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Bocock, J.P.; Carmicle, S.; Madamba, E.; Erickson, A.H. Nuclear targeting of an endosomal E3 ubiquitin ligase. Traffic 2010, 11, 756–766. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Y.; Stenmark, H. Cellular functions of Rab GTPases at a glance. J. Cell Sci. 2015, 128, 3171–3176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Press, B.; Wandinger-Ness, A. Rab 7: An important regulator of late endocytic membrane traffic. J. Cell Biol. 1995, 131, 1435–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombardi, D.; Soldati, T.; Riederer, M.A.; Goda, Y.; Zerial, M.; Pfeffer, S.R. Rab9 functions in transport between late endosomes and the trans Golgi network. EMBO J. 1993, 12, 677–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasa, S.; Modica, G.; Sauvageau, E.; Kaleem, A.; Hermey, G.; Lefrancois, S. CLN3 regulates endosomal function by modulating Rab7A-effector interactions. J. Cell Sci. 2020, 133, 4047. [Google Scholar] [CrossRef]
- Ceresa, B.P.; Peterson, J.L. Cell and molecular biology of epidermal growth factor receptor. Int. Rev. Cell Mol. Biol. 2014, 313, 145–178. [Google Scholar] [CrossRef]
- Mayle, K.M.; Le, A.M.; Kamei, D.T. The intracellular trafficking pathway of transferrin. Biochim. Biophys. Acta 2012, 1820, 264–281. [Google Scholar] [CrossRef] [Green Version]
- Bonifacino, J.S.; Traub, L.M. Signals for sorting of transmembrane proteins to endosomes and lysosomes. Annu. Rev. Biochem. 2003, 72, 395–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.Y.; Guo, X. Adaptor protein complexes and intracellular transport. Biosci. Rep. 2014, 34, 69. [Google Scholar] [CrossRef]
- Huttlin, E.L.; Ting, L.; Bruckner, R.J.; Gebreab, F.; Gygi, M.P.; Szpyt, J.; Tam, S.; Zarraga, G.; Colby, G.; Baltier, K.; et al. The BioPlex Network: A Systematic Exploration of the Human Interactome. Cell 2015, 162, 425–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huttlin, E.L.; Bruckner, R.J.; Paulo, J.A.; Cannon, J.R.; Ting, L.; Baltier, K.; Colby, G.; Gebreab, F.; Gygi, M.P.; Parzen, H.; et al. Architecture of the human interactome defines protein communities and disease networks. Nature 2017, 545, 505–509. [Google Scholar] [CrossRef]
- Hung, M.C.; Link, W. Protein localization in disease and therapy. J. Cell Sci. 2011, 124, 3381–3392. [Google Scholar] [CrossRef] [Green Version]
- Laurila, K.; Vihinen, M. Prediction of disease-related mutations affecting protein localization. BMC Genom. 2009, 10, 122. [Google Scholar] [CrossRef] [PubMed]
- Girard, E.; Chmiest, D.; Fournier, N.; Johannes, L.; Paul, J.L.; Vedie, B.; Lamaze, C. Rab7 is functionally required for selective cargo sorting at the early endosome. Traffic 2014, 15, 309–326. [Google Scholar] [CrossRef] [Green Version]
- Poteryaev, D.; Datta, S.; Ackema, K.; Zerial, M.; Spang, A. Identification of the switch in early-to-late endosome transition. Cell 2010, 141, 497–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kälin, S.; Hirschmann, D.T.; Buser, D.P.; Spiess, M. Rabaptin5 is recruited to endosomes by Rab4 and Rabex5 to regulate endosome maturation. J. Cell Sci. 2015, 128, 4126–4137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bucci, C.; Parton, R.G.; Mather, I.H.; Stunnenberg, H.; Simons, K.; Hoflack, B.; Zerial, M. The small GTPase rab5 functions as a regulatory factor in the early endocytic pathway. Cell 1992, 70, 715–728. [Google Scholar] [CrossRef] [Green Version]
- Stenmark, H.; Parton, R.G.; Steele-Mortimer, O.; Lütcke, A.; Gruenberg, J.; Zerial, M. Inhibition of rab5 GTPase activity stimulates membrane fusion in endocytosis. EMBO J. 1994, 13, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- Hassink, G.; Slotman, J.; Oorschot, V.; Van Der Reijden, B.A.; Monteferrario, D.; Noordermeer, S.M.; Van Kerkhof, P.; Klumperman, J.; Strous, G.J. Identification of the ubiquitin ligase Triad1 as a regulator of endosomal transport. Biol. Open 2012, 1, 607–614. [Google Scholar] [CrossRef] [Green Version]
- Rosenfeld, J.L.; Moore, R.H.; Zimmer, K.P.; Alpizar-Foster, E.; Dai, W.; Zarka, M.N.; Knoll, B.J. Lysosome proteins are redistributed during expression of a GTP-hydrolysis-defective rab5a. J. Cell Sci. 2001, 114, 4499. [Google Scholar] [CrossRef]
- Kimple, M.E.; Brill, A.L.; Pasker, R.L. Overview of affinity tags for protein purification. Curr. Protoc. Protein Sci. 2013, 73, 9.9.1–9.9.23. [Google Scholar] [CrossRef] [Green Version]
- Ferri, K.F.; Kroemer, G. Organelle-specific initiation of cell death pathways. Nat. Cell Biol. 2001, 3, E255–E263. [Google Scholar] [CrossRef]
- De Araujo, M.E.G.; Liebscher, G.; Hess, M.W.; Huber, L.A. Lysosomal size matters. Traffic 2020, 21, 60–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platt, F.M.; Boland, B.; van der Spoel, A.C. The cell biology of disease: Lysosomal storage disorders: The cellular impact of lysosomal dysfunction. J. Cell Biol. 2012, 199, 723–734. [Google Scholar] [CrossRef] [Green Version]
- Holland, P.; Torgersen, M.L.; Sandvig, K.; Simonsen, A. LYST Affects Lysosome Size and Quantity, but not Trafficking or Degradation Through Autophagy or Endocytosis. Traffic 2014, 15, 1390–1405. [Google Scholar] [CrossRef] [PubMed]
- Vitner, E.B.; Platt, F.M.; Futerman, A.H. Common and uncommon pathogenic cascades in lysosomal storage diseases. J. Biol. Chem. 2010, 285, 20423–20427. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Kim, S.J.; Zhang, Z.; Tsai, P.C.; Wisniewski, K.E.; Mukherjee, A.B. ER and oxidative stresses are common mediators of apoptosis in both neurodegenerative and non-neurodegenerative lysosomal storage disorders and are alleviated by chemical chaperones. Hum. Mol. Genet. 2008, 17, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Sanger, A.; Hirst, J.; Davies, A.K.; Robinson, M.S. Adaptor protein complexes and disease at a glance. J. Cell Sci. 2019, 132, jcs222992. [Google Scholar] [CrossRef]
- Hofmann, M.W.; Höning, S.; Rodionov, D.; Dobberstein, B.; von Figura, K.; Bakke, O. The leucine-based sorting motifs in the cytoplasmic domain of the invariant chain are recognized by the clathrin adaptors AP1 and AP2 and their medium chains. J. Biol. Chem. 1999, 274, 36153–36158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Höning, S.; Sandoval, I.V.; von Figura, K. A di-leucine-based motif in the cytoplasmic tail of LIMP-II and tyrosinase mediates selective binding of AP-3. EMBO J. 1998, 17, 1304–1314. [Google Scholar] [CrossRef]
- Scott, C.; Higgins, M.E.; Davies, J.P.; Ioannou, Y.A. Targeting of NPC1 to Late Endosomes Involves Multiple Signals, Including One Residing within the Putative Sterol-sensing Domain *. J. Biol. Chem. 2004, 279, 48214–48223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, A.C.; Salazar, G.; Styers, M.L.; Newell-Litwa, K.A.; Werner, E.; Maue, R.A.; Corbett, A.H.; Faundez, V. The subcellular localization of the Niemann-Pick Type C proteins depends on the adaptor complex AP-3. J. Cell Sci. 2007, 120, 3640–3652. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabana, V.C.; Bouchard, A.Y.; Sénécal, A.M.; Ghilarducci, K.; Kourrich, S.; Cappadocia, L.; Lussier, M.P. RNF13 Dileucine Motif Variants L311S and L312P Interfere with Endosomal Localization and AP-3 Complex Association. Cells 2021, 10, 3063. https://doi.org/10.3390/cells10113063
Cabana VC, Bouchard AY, Sénécal AM, Ghilarducci K, Kourrich S, Cappadocia L, Lussier MP. RNF13 Dileucine Motif Variants L311S and L312P Interfere with Endosomal Localization and AP-3 Complex Association. Cells. 2021; 10(11):3063. https://doi.org/10.3390/cells10113063
Chicago/Turabian StyleCabana, Valérie C., Antoine Y. Bouchard, Audrey M. Sénécal, Kim Ghilarducci, Saïd Kourrich, Laurent Cappadocia, and Marc P. Lussier. 2021. "RNF13 Dileucine Motif Variants L311S and L312P Interfere with Endosomal Localization and AP-3 Complex Association" Cells 10, no. 11: 3063. https://doi.org/10.3390/cells10113063
APA StyleCabana, V. C., Bouchard, A. Y., Sénécal, A. M., Ghilarducci, K., Kourrich, S., Cappadocia, L., & Lussier, M. P. (2021). RNF13 Dileucine Motif Variants L311S and L312P Interfere with Endosomal Localization and AP-3 Complex Association. Cells, 10(11), 3063. https://doi.org/10.3390/cells10113063